Ископаемые плауновидные (Н. С. Снигеревская)
Наиболее примитивной и наиболее древней группой плауновидных является порядок астероксиловых (Asteroxylales). До недавнего времени этот порядок относили к риниофитам.
Однако сифоностелическое строение стебля, наличие листьев и в целом более подвинутый тип организации позволяют с полной уверенностью рассматривать его в пределах отдела плауновидных.
Порядок включает одно семейство астероксиловых (Asteroxylaceae) и, по-видимому, два рода - астероксилон (Asteroxylon) и схизоподиум (Schizopodium). Оба рода вымерли около 400 млн. лет назад. Первый известен только в отложениях среднего девона из ряда местонахождений (Шотландия, ФРГ, СССР и КНР), второй - в среднем девоне Австралии и позднем девоне США. Нам известны как отпечатки, так и окаменелости этих растений. Род астероксилон встречается вместе с ринией и хорнеофитом в названных выше кремнистых чертах Шотландии, близ деревни Райни.
Астероксилон был травянистым растением с очень четким расчленением на подземную и надземную части, а также на стебель и листья (филлоиды), что, как мы видели, было не свойственно риниофитам (рис. 56).
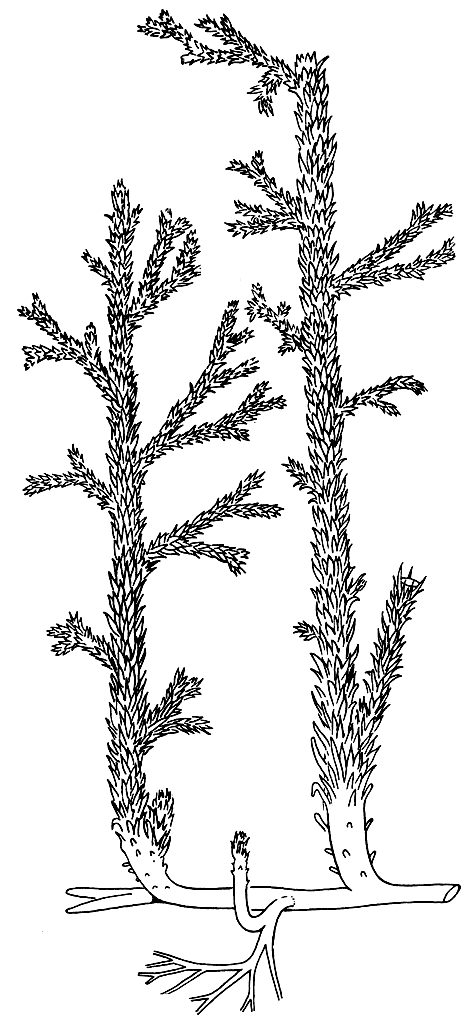
Рис. 56. Астероксилон (Asteroxylon)
Надземная часть астероксилона была представлена прямостоячими, изредка вильчато, или дихоподиально, ветвящимися стеблями, боковые ветви которых многократно дихотомировали. Мелкие шиловидные листья с жилкой, входящей в их основание, но не доходящей до верхушки, густо покрывали стебли, как у современных плаунов. Устьица располагались не только на листьях, но также на стебле.
Стебли отходили от подземной части, представленной горизонтально распростертым дихотомически ветвящимся ризомоидом, лишенным, как у всех плауновидных, корневых волосков.
Стела стебля представлена экзархной или переходной к мезархной актиностелой, которая в основании стебля переходит в сифоностелу со слабо развитой сердцевиной. Это отличает астероксилон от риниофитов и сближает с плауновидными. Ксилема имеет ясно выраженную звездчатую форму, с чем связано научное название рода Asteroxylon (от греч. aster - звезда и xylon - дерево, в данном случае - древесина).
Основную массу стебля (приблизительно 4/5 его диаметра) составляла кора, очень мощная в сравнении с диаметром стелы. Ее средняя часть имела губчатое строение; она состояла из широких воздухоносных каналов (полостей), разделенных перегородками из паренхимных клеток.
Обилие воздухоносных полостей свидетельствует, по мнению ученых, о земноводном, полупогруженном образе жизни астероксилона.
Анатомическое строение ризомоида было очень примитивным и сходным со структурой ризомоида ринии. Интересно, что в коре как стебля, так и ризомоида были обнаружены гифы и споры грибов, с которыми, по-видимому, астероксилон находился в симбиозе.
Спороношения, достоверно принадлежащие астероксилону, до настоящего времени не обнаружены.
Среди древнейших плауновидных, остатки которых обнаружены в отложениях девонской системы, наряду с представителями порядка астероксиловых (Asteroxylales) встречены остатки растений и из другого целиком вымершего в палеозойскую эру порядка протолепидодендровых (Protolepidodendrales). Обладая характерными особенностями отдела, эти растения отличались от остальных плауновидных прежде всего вильчато разветвленными листьями (рис. 57).
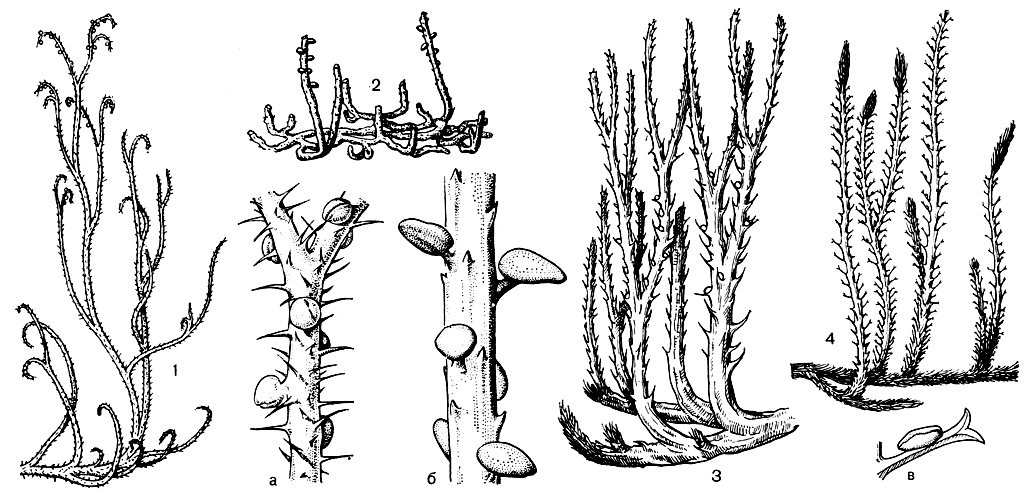
Рис. 57. Ископаемые плауновидные: 1 - содония украшенная (Sawdonia ornata): а - участок побега с шипами и спорангиями; 2 - каулангиофитон иглистый (Kaulangiophyton aicantha): б - участок побега с шипами и спорангиями; 3 - дрепанофикус колючковидный (Drepanophycus spiniformis); 4 - протолепидодендрон Шари (Protolepidodendron scharianum): в - спорофилл
В верхнем палеозое плауновидные достигли своего максимального расцвета: представители многочисленных родов и семейств были широко распространены по земному шару и занимали различные экологические ниши. Но основную роль в сложении растительного покрова многих областей играли, по всем данным, представители класса полушниковых. Среди них особенно выделялись своими размерами лепидодендроны, или чешуедревы (Lepidodendron), из порядка лепидодендровых (Lepidodendrales).
Лепидодендроны обладали колонновидным главным стеблем - стволом - высотой до 30 м и диаметром более 1 м у основания (рис. 58). Первичная и вторичная древесины занимали небольшую часть объема ствола, большая часть которого была сложена паренхимной тканью. Крона дерева формировалась в результате многочисленных последовательных дихотомических ветвлений стебля.
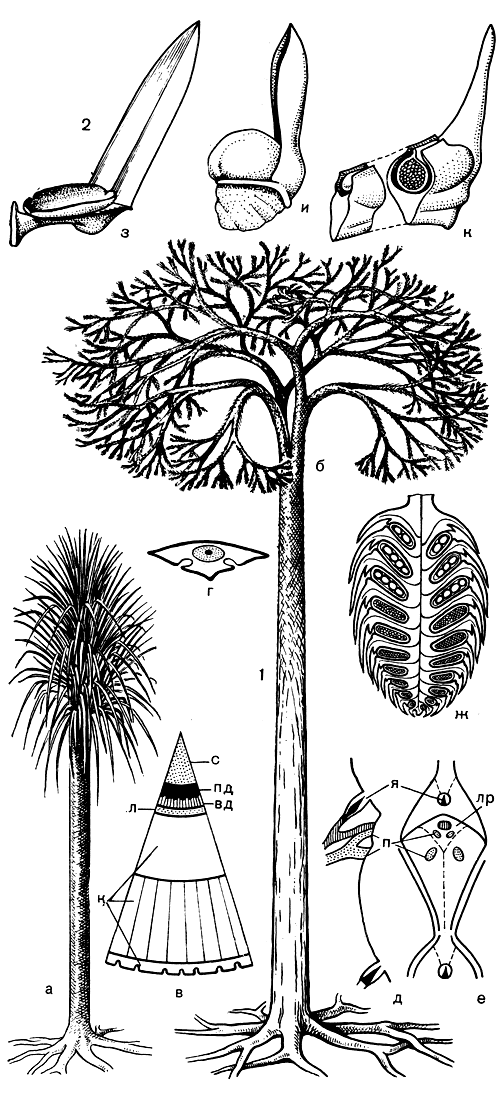
Рис. 58. Лепидодендровые: 1 - лепидодендрон (Lepidodendron): а - молодое растение, б - взрослое растение, в - схема поперечного среза ствола, г - схема поперечного среза листа, д - листовая подушка на продольном разрезе ствола, е - листовая подушка в плане, ж - стробил, вд - вторичная древесина, к - кора, л - луб, лр - листовой рубец, п - парихны, пд - первичная древесина, с - сердцевина, я - язычок; 2 - спорофиллы лепидодендровых: з - спорофилл с тонкостенным вскрывшимся спорангием, внутри которого находилось несколько спор, и - мегаспорофилл с толстостенным спорангием, содержащим мегаспору, к - мегаспорофилл с толстостенным спорангием, прикрытым крыльями пластинки спорофилла, внутри спорангия находится женский гаметофит
Ствол молодого растения был густо покрыт очень длинными шиловидными листьями. У некоторых видов они могли быть длиной до 1 м при ширине более 1 см.
По мере роста растения листья постепенно опадали начиная снизу вверх, и на поверхности ствола оставались лишь листовые подушки, в верхней части которых, в ямке, сидел язычок, а в средней находился листовой рубец с пучком листового следа и выходами воздухоносной ткани - парихнами (от греч. para - около и ichnos - след). Листья были покрыты тонким слоем кутикулы, а под эпидермой располагался мощный слой толстостенных клеток, придававший листьям необходимую жесткость и упругость. Кольцо этой укрепляющей ткани прерывалось только на нижней стороне листа, в области двух устьичных ложбинок, на дне которых располагались многочисленные устьица. При таком погруженном положении устьичного аппарата капельки воды, конденсируемые на поверхности растения из атмосферной влаги, не могли закупорить устьица, и испарение воды (которое и так шло во влажной атмосфере тропических болот с небольшой скоростью) не прерывалось. Это имело большое значение для поддержания восходящего тока воды в гигантском стволе и громадной кроне. Поддержанию такого тока способствовала и деятельность язычков, которые выделяли воду, поставляемую к их основанию мощным тяжем ксилемы.
Газообмен растения с окружающей средой осуществлялся не только с помощью устьиц, но и через парихны. По межклетникам воздухоносной ткани, буквально пронизывающей все органы растения, воздух с кислородом проходил и в находящуюся в анаэробной болотистой почве корневую систему. На подземных осях корневой системы сидели по спирали корни, не имевшие корневых волосков, длиной до 50 см и более. Эти оси, или ризофоры, называют стигмариями (от греч. stigma - клеймо), так как после опадения корней поверхность ризофоров оказывалась покрытой небольшими округлыми рубчиками.
Стробилы лепидодендронов, сидевшие на концах ветвей, могли достигать в длину 50 см и диаметра 5 см. В обоеполых стробилах мегаспорофиллы располагались в нижней, а микроспорофиллы - в верхней их части. В микроспорангиях развивались тысячи мелких микроспор. У более примитивных видов лепидодендронов в мегаспорангиях развивались сотни небольших мегаспор. У более подвинутых их число уменьшилось до 16, даже до 12 и 4, а размер мегаспор увеличился почти до 1 мм. И наконец, в спорангиях наиболее подвинутых видов развивалась всего одна мегаспора, которая тут же прорастала, образуя женский гаметофит (заросток), защищенный от неблагоприятных воздействий внешней среды толстой стенкой спорангия или завернутыми краями спорофилла, создававшими для гаметофита дополнительный покров (рис. 58).
Эволюция полушниковых как класса шла далее таким образом, что растения как бы созревали на все более и более ранних стадиях индивидуального развития. На смену сильно разветвленным гигантским лепидодендронам с многочисленными стробилами в пермское время пришли более низкорослые и менее разветвленные сигиллярии, или печатницы (Sigillaria, рис. 59), которых в мезозойское время сменили плевромейи (Pleuromeia, рис. 59) - невысокие (1-2 м), неразветвленные растения с одним верхушечным стробилом. В некоторых районах Земли плевромейи образовывали по берегам морей заросли, возможно напоминающие современные мангры. Одновременно с уменьшением величины и степени разветвленности верхней части оси полушниковых уменьшались размеры и число ветвей ризофора, и в конце концов эти процессы достигли своей крайней степени выражения у современного полушника, который условно можно считать лепидодендроном, созревающим в первый год жизни. Образно говоря, старцы чешуедревы дошли до наших дней в образе преждевременно состарившихся юношей.
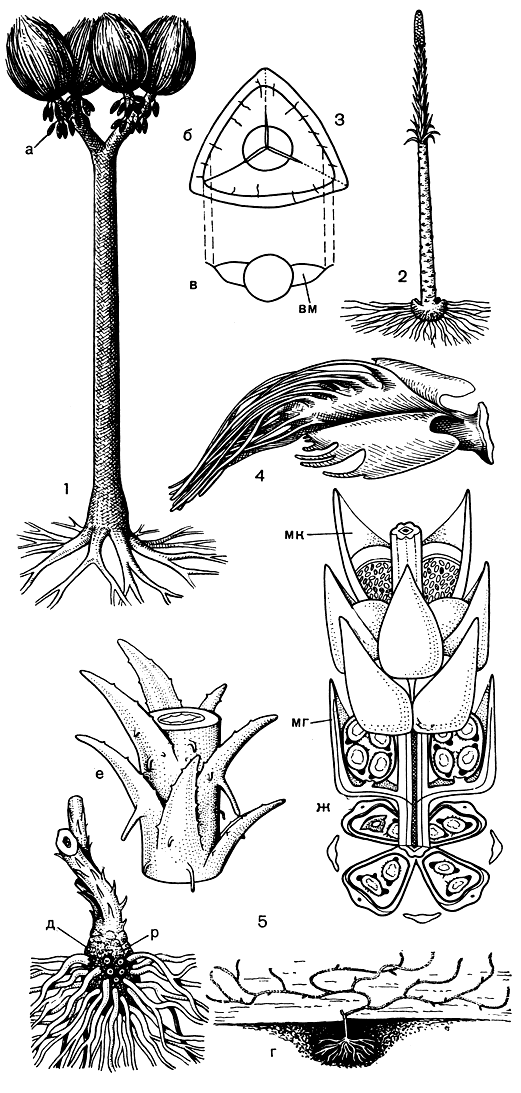
Рис. 59. Ископаемые плауновидные: 1 - сигиллярия (Sigillaria): а - стробил; 2 - плевромейя Штернберга (Pleuromeia sternbergii); з - спора спенсериспоритеса (Spencerisporites): б - вид с полюса, в - продольное сечение споры, виден воздушный мешок (вм); 4 - мегаспорофилл миадесмии кожистой (Miadesmia membranacea); 5 - селагинелла Фрайпонта (Selaginella fraipontii): г - реконструкция растения со стелющимися по поверхности земли побегами, д - основание побега с ризофором (р), е - участок побега с листьями, ж - часть стробила с мегаспорофиллами (мг) и с микроспорофиллами (мк); внутри мегаспорангиев видны тетрады мегаспор, внутри микроспорангиев - многочисленные микроспоры
К отделу плауновидных, возможно, относятся не имевшие еще, как и астероксилон, настоящих листьев с жилками содония и каулангиофитон (рис. 57, 1-2), которые многими палеоботаниками сближаются с зостерофиллофитами. Некоторые каменноугольные плауновые имели споры с воздушными мешками (рис. 59, 3).
В каменноугольных лесах из древовидных представителей полушниковых на почве произрастали не только травянистые представители плауновых, но и травы из порядка селагинелловых (рис. 59, 4-5).
В конце палеозойской эры и в начале мезозойской все крупные высокоспециализированные представители плауновидных вымерли, уступив свое место более конкурентоспособным голосеменным. Но обитавшие под пологом своих древовидных сородичей мелкие травянистые плауновидные смогли не только пережить эру господства голосеменных, но и успешно приспособиться в дальнейшем к жизни в сложно организованных сообществах с преобладанием цветковых растений.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'