Класс плауновые, или ликоподиопсиды (Lycopodiopsida) (В. Р. Филин)
Порядок плауновые (Lycopodiales)
К порядку относят многолетние равноспоровые травянистые растения без ризофоров, с цельными листьями без язычков. Побеги не имеют камбия. Спорангии располагаются в пазухе листа или на его внутренней стороне, а спорофиллы образуют на побегах спороносные зоны или собраны в стробилы. Гаметофиты обоеполые, мясистые, подземные и полуподземные, сапрофитные или полусапрофитные, созревающие в течение года или за 5-15 лет.
Семейство плауновые (Lycopodiaceae)
К семейству принадлежат растения, для которых характерно наличие спорангиев, сидящих в пазухе листа или на его внутренней стороне. Споры без воздушных мешков. Из ныне живущих растений к семейству относят всего два рода - плаун (Lycopodium) и филлоглоссум (Phylloglossum).
Некоторые виды вымершего рода ликоподитес (Lycopodites) также принадлежат к этому семейству.
Латинское название рода Lycopodium было введено в науку в XVI в. немецким ботаником Я. Т. Табернемонтаном, который, описывая и изображая плаун, известный сейчас как плаун булавовидный (Lycopodium clavatum), привел в качестве синонима немецких названий этого растения слово lycopodium, которое означает буквально "волчья лапка". Русские названия "плаун", "плавун", "плывун" также были связаны главным образом с плауном булавовидным. Впоследствии название "плаун" перешло на остальные виды этого рода, которые из-за их облика или свойства часто назывались иначе: например, баранец (теперь плаун баранец - L. selago), болотная можжуха (теперь плаун годичный - L. annotinum) и т. д.
Все плауны - вечнозеленые наземные или эпифитные, растущие на стволах и ветвях деревьев многолетние травянистые растения с прямостоячими, полегающими, свисающими, ползучими или лазящими побегами, с придаточными корнями. Высота прямостоячих побегов у наземных растений или свисающих побегов у эпифитов обычно невелика (не более 1-1,5 м), длина ползучих побегов может быть весьма значительной и иногда превышает 10 м.
Среди плаунов есть виды, стебли которых ветвятся равнодихотомически (плаун баранец, табл. 13), неравнодихотомически (плауны годичный и булавовидный, табл. 13) и псевдомоноподиально (плаун поникший - L. cernuum, рис. 55).

Таблица 13. Плауновидные: вверху слева направо - селагинелла селаговидная (Selaginella selaginoides); плаун баранец (Lycopodium selago); плаун годичный (L. annotinum); внизу - плаун сплюснутый (L. complanatum)
У таких видов, как плаун годичный или плаун булавовидный, при неравновильчатом ветвлении возникает главная, или скелетная, ось - ползучий стебель, от которого отходят боковые полегающие побеги обычно с ограниченным ростом.
Стебли плауна радиально- или двусторонне-симметричны, в последнем случае у них можно различить брюшную и спинную стороны. С поверхности стебель покрыт однослойной эпидермой с устьицами, за которой следует широкая зона первичной коры. В коре часто можно выделить несколько зон (рис. 65).
Располагающаяся в центре стебля цилиндрическая протостела занимает сравнительно небольшую часть его объема. Очертания участков, занимаемых трахеидами ксилемы на поперечном срезе стелы, варьируют по форме не только у разных видов, но и в пределах стебля одного растения.
Ксилема плауна первичная, она возникает исключительно из прокамбия. Метаксилема состоит из сетчатых трахеид и трахеид с лестничными или округлыми окаймленными порами, расположенными в 1-4 продольных ряда на стенке (рис. 60).

Рис. 60. Проводящие элементы ксилемы плаунов: вверху - трахеиды с лестничными окаймленными порами в корне плауна сплюснутого (Lycopodium complanatum) (увел, около 2200); в середине - трахеиды с округлыми и овальными окаймленными порами в стебле плауна баранца (L. selago) (увел, около 2100); внизу - трахеиды с овальными окаймленными порами в стебле плауна баранца, видны замыкающие мембраны окаймленных пор (увел, около 6000)
Проводящие элементы флоэмы представляют собой очень длинные клетки с ситовидными участками на продольных стенках.
Листья зрелого побега у большинства плаунов располагаются поочередно, но у некоторых видов они сидят супротивно, мутовчато (по 4-5 в мутовке) или без особого порядка. Листья прижатые или отстоящие. Они сравнительно небольшие: длиной от 0,7 мм у плауна покрытого (L. vestitum) до 30-35 мм у плауна пильчатого (L. serratum). Форма пластинки у разных видов варьирует и бывает шиповидной, линейной, продолговатой, овальной, вальковатой, треугольной. У одного и того же растения на разных ветвях и даже в пределах одного годичного побега нередко встречаются листья разной величины и формы (рис. 55).
Устьица располагаются беспорядочно только на брюшной (например, у плауна альпийского - L. alpinum), только на спинной (например, у плауна годичного) или на обеих поверхностях листа (например, у плауна булавовидного). Нередко устьица приурочены к той поверхности листа, которая меньше подвергается иссушающему воздействию солнечных лучей и ветра.
Мезофилл листа обычно недифференцированный, состоит из более или менее изодиаметрических клеток. Вдоль листа в его средней части проходит жилка, состоящая из трахеид и паренхимных клеток и отграниченная иногда от мезофилла эндодермой. У плауна заливаемого (L. inundatum) и некоторых других видов на месте элементов протоксилемы возникают слизевые ходы, примыкающие к трахеидам жилки.
У видов с полегающими главными побегами, например у плауна баранца, по мере нарастания побегов в высоту основания их, образовавшиеся несколько лет назад, изгибаются и полегают в такой мере, что высота надземной части зрелого растения остается более или менее постоянной в течение многих десятков лет. Одновременно полегающие участки побегов погружаются в нарастающую моховую дернину или накапливающуюся лесную подстилку, и на них появляются придаточные корни (заложившиеся в выше расположенных участках побега задолго до их полегания). Однако погрузившиеся в почву побеги, конечно, не являются настоящими корневищами. В гораздо большей степени уподобляются корневищам ползучие побеги у плауна булавовидного и плауна годичного, так как придаточные корни появляются на них очень рано, а сами побеги быстро погружаются в моховую дернину или лесную подстилку. Длинные, подземные, залегающие на глубине 5-15 см, не зеленые побеги с расставленными чешуевидными листьями у плауна неясного (L. obscurum) или у плауна сплюснутого (L. complanatum) могут рассматриваться уже как настоящие корневища. И в полегающих и в ползучих побегах, и в корневищах плаунов имеются гифы почвенных грибов, проникающие в ткани растения, по-видимому, через поврежденные участки побега.
Корни закладываются в перицикле или эндодерме стебля. Зачаток корня у эпифитов и у наземных видов с полегающими главными побегами (например, у плауна баранца) после своего появления растет внутри коры по направлению к основанию стебля и лишь спустя некоторое время выходит на поверхность, проникая в почву иногда до глубины 30 см.
Вегетативное размножение у плаунов в большинстве случаев происходит путем отмирания старых участков побегов и корневищ. При этом более молодые участки, снабженные придаточными корнями, продолжают существовать в качестве самостоятельных растений. У некоторых плаунов есть и специализированные органы вегетативного размножения. Так, корни и листья эпифитов легко образуют выводковые клубеньки. У плауна баранца и близких ему видов (плаун светлый - L. lucidum и др.) имеются выводковые луковички или почки, образующиеся на верхушках побегов. По созревании почка опадает или отбрасывается своеобразным катапультирующим механизмом на значительное (до 0,5 м) расстояние, чему способствует также обтекаемая, как у самолета, форма тела почки. На почве при наличии благоприятных условий почка прорастает. Выводковые почки образуются на побеге раз в год, и по числу спороносных зон стебля, разделенных зонами с сохранившимися основаниями выводковых почек, можно легко установить возраст растения и вычислить среднюю величину годичного прироста побега (рис. 61). У плауна заливаемого в конце сезона роста отмирает все растение, за исключением утолщающейся верхушки побега; ее также можно рассматривать как своеобразную почку возобновления.

Рис. 61. Побеги плауна баранца (Lycopodium selago) с выводковыми почками
Бесполое размножение у плаунов осуществляется с помощью спор, которые образуются в спорангиях, расположенных по одному в пазухах спорофиллов или на верхней стороне последних, близ их основания. Реже спорангии образуются прямо на стебле, над пазухой листа (рис. 62).
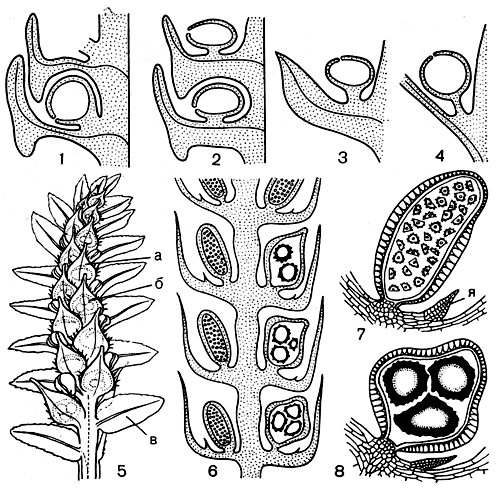
Рис. 62. Схема расположения спорангиев на стебле и щели вскрывания в стенке спорангия у плаунов (1-4), строение стробила и спорангиев у селагинеллы (5-8): 1 - плаун заливаемый (Lycopodium inundatum); 2 - плаун поникший (L. cernuum); 3 - плаун оттопыренный (L. squarrosum); 4 - плаун светловатый (L. lucidulum); 5 - стробил селагинеллы золотистостебельной (Selaginella chrysocaulos), вид с нижней стороны: а - микроспорофилл, б - мегаспорофилл, в - боковой лист; б - схема продольного разреза стробила селагинеллы Крауса (S. kraussiana); 7 - микроспорангий селагинеллы Крауса: я - язычок; 8 - мегаспорангий селагинеллы Крауса
Спорофиллы сходны по цвету, форме и величине с вегетативными листьями или более или менее резко отличаются от них. Сходные с вегетативными листьями спорофиллы обычно образуются на стебле зрелого растения несколько лет подряд, вследствие чего возникает побег со слабо бросающимися в глаза спороносными зонами (табл. 13). Когда спорофиллы отличаются по форме, величине и цвету от вегетативных листьев, они собраны в стробилы - сидячие или на ножках, прямых или поникающих, простых, неветвистых или разветвленных (табл. 13, рис. 55). Длина стробилов варьирует от 0,5 до 40 см, а толщина - от 1 до 6 мм.
Спорангии возникают из группы инициальных поверхностных клеток листа или стебля. От момента заложения спорангия до времени высыпания зрелых спор проходит несколько месяцев или лет. Зрелый спорангий желтоватый, почковидный или почти шаровидный (с наибольшим диаметром 1-2,5 мм). Вскрывается он поперечной щелью (рис. 62). При увеличении влажности воздуха (во время дождя, тумана) створки спорангия закрываются и споры остаются сухими.
Зрелые споры очень мелкие, диаметром около 30 мкм, тетраэдрические с трехлучевым тетрадным рубцом и разнообразной поверхностью (табл. 11).
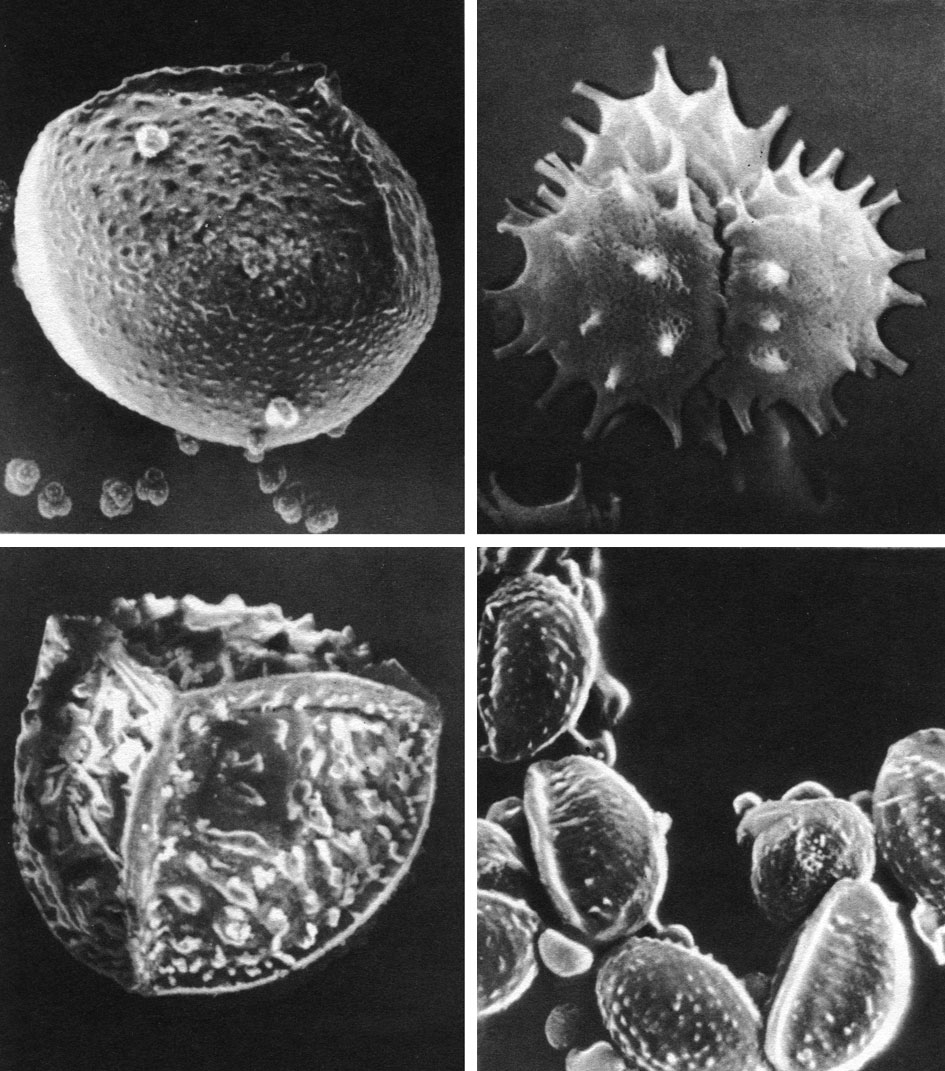
Таблица 11. Споры плауновидных под сканирующим электронным микроскопом: вверху слева - мегаспора и тетрады микроспор селагинеллы селаговидной (Selaginella selaginoides) (увел, около 160); вверху справа - тетрада микроспор селагинеллы селаговидной (увел, около 1200); внизу слева - мегаспора шильника озерного (Isoetes lacustris) (увел, около 120); внизу справ а - микроспоры шильника озерного (увел, около 800)
У плауна отмечены два типа прорастания спор - подземное и наземное. При подземном прорастании споры, обладающие длительным периодом вынужденного покоя, постепенно покрываются слоем падающих сверху минеральных частиц и частиц растительного детрита и как бы погружаются в почву, у эпифитов - внедряются в кору дерева-хозяина. Некоторые споры действительно погружаются в субстрат вместе с токами воды, а, возможно, также и в результате деятельности различных беспозвоночных, например дождевых червей, через пищеварительный тракт которых споры, вероятно, проходят невредимыми. Споры прорастают через 3-8 лет после высыпания из спорангия, находясь уже "под землей", в слое гумуса.
Уже на самых ранних стадиях развития гаметофита в некоторые из его клеток внедряются гифы почвенных грибов. Если этого не происходит, дальнейший рост гаметофита прекращается. При наличии гиф гриба гаметофит, питаясь сапрофитно, медленно растет. У верхушки гаметофита появляется меристема, и его рост приобретает более организованный характер. Спустя 6-15 лет после прорастания спор, находясь уже в почве на глубине 1-8 см, гаметофит созревает, приобретая окончательную форму, характерную, как полагают, для определенных групп видов (рис. 63). После оплодотворения гаметофиты еще долго не отмирают и в течение нескольких лет питают молодые спорофиты.
При наземном прорастании споры плауна, попав на землю, прорастают в течение нескольких дней и дают начало наземному (точнее, полуподземному) гаметофиту. Полагают, что этот гаметофит питается полусапрофитно; в основании погруженной в почву бледно-зеленой части гаметофита содержатся гифы почвенных грибов, а надземная часть гаметофита несет многочисленные зеленые листообразные выросты (рис. 63). Такие полуподземные гаметофиты (диаметром 1-5 мм), как правило, созревают в течение одного сезона и после оплодотворения быстро погибают, но известны случаи их перезимовки в условиях умеренного климата.

Рис. 63. Заростки (гаметофиты), гаметангии, зародыши и проростки плаунов: 1 - заросток плауна заливаемого (Lycopodium inundatum); 2 - заросток плауна поникшего (L. cernuum) с двумя зародышами; 3 - заросток и проросток плауна булавовидного (L. clavatum); 4 - заросток и проросток плауна сплюснутого (L. complanatum); 5-7 - заростки и проросток плауна баранца (L. selago); 8 - заросток плауна флегмарии (L. phlegmaria); 9 - антеридий плауна булавовидного; 10 - архегоний плауна булавовидного; 11 - зародыш плауна булавовидного; 12 - проросток плауна булавовидного; 13 - схема строения зародыша плаунов, обладающих подземными заростками; 14 - схема строения зародыша плаунов, обладающих наземными заростками; (а - заросток; б - корень; в - лист; г - ножка; д - подвесок; е - протокорм; ж - проросток; з - протофилл; и - яйцеклетка)
Все описанные до сих пор гаметофиты плаунов являются обоеполыми, несут многочисленные антеридии и архегонии. Зрелые антеридии почти полностью погружены в ткань гаметофита или несколько выдаются над его поверхностью (рис. 63). Сперматозоиды плауна наиболее простые по внешней форме и наиболее мелкие среди описанных до сих пор сперматозоидов высших растений. На переднем конце сперматозоида располагаются 2 (редко 3) бичевидных жгутика. Зрелый архегоний состоит из узкого, погруженного в ткань гаметофита брюшка и выступающей над его поверхностью длинной или короткой шейки (рис. 63).
Антеридии обычно созревают раньше архегониев, что на первых этапах созревания гаметофита снижает вероятность самооплодотворения и увеличивает шансы перекрестного оплодотворения. Случаи нахождения гибридов плауна в природе указывают на то, что яйцеклетка может оплодотвориться сперматозоидом от гаметофита другого вида.
Зигота без периода покоя прорастает и дает начало зародышу. Перегородкой, проходящей перпендикулярно длинной оси архегония, зигота делится пополам. Верхняя, обращенная к шейке архегония клетка (подвесок) больше не делится и лишь ограничивает развитие зародыша нижней половиной брюшка архегония. Из нижней зародышевой клетки после трех последовательных делений возникает восемь клеток, расположенных в два яруса по 4. Дальнейшее развитие зародыша у видов с наземными и подземными гаметофитами протекает по-разному. У видов с подземными гаметофитами 4 клетки верхнего, обращенного к шейке архегония яруса дают начало эллипсоидальной ножке диаметром до 1,5-3 мм. Ножка является гаусторией - она всасывает питательные вещества из гаметофита и передает их собственно зародышу, который развивается из четырех клеток нижнего яруса. Две клетки этого яруса дают начало зародышевому стеблю, две - зародышевому листу.
По мере роста стебля на нем появляются листья, а у основания зародышевого листа образуется придаточный корень (рис. 63). Первый побег проростка иногда несколько лет пробивается к поверхности, и все эти годы проросток или даже несколько проростков получают питательные вещества с помощью ножки из гаметофита, который продолжает увеличиваться в объеме, питаясь сапрофитно. После того как побег окажется на свету, он зеленеет и переходит на самостоятельное питание.
У видов с наземными гаметофитами ножка развита очень слабо, а из четырех клеток нижнего яруса предзародыша образуется клубневидное тело - протокорм, или первичный стебель (рис. 63). От нижней стороны протокорма отходят ризоиды, а на верхней стороне беспорядочно располагаются немногочисленные зеленые первичные листья, или протофиллы. Хотя в клетках тела протокорма могут развиваться хлоропласты, он, как и гаметофит, живет в симбиозе с гифами гриба. На протокорме затем появляется почка, дающая начало облиственному побегу с придаточными корнями. Кончик первичного корешка иногда утолщается и превращается в клубенек типа протокорма.
В пределах рода плаун встречаются виды как со сравнительно небольшим числом хромосом (например, у плауна сплюснутого 2n = 46), так и с очень высокими хромосомными числами (например, у одной из рас плауна поникшего 2n = примерно 340). Между этими крайними величинами можно поместить такие хромосомные наборы, как 2n = 68, 2n = 90, 2n = 156 и 2n = 264. Все известные данные по цитологии говорят о том, что в эволюции рода плаун большую роль сыграли полиплоидия и гибридизация.
Объем рода плаун (в широком смысле) из-за разногласий в понимании границ видов оценивается по-разному - 200 или даже около 500 видов.
Общепринятой системы рода, находящейся в согласии со всеми известными данными сравнительной морфологии, анатомии, цитологии, хорологии, пока не существует.
Наряду с давней тенденцией к расщеплению рода все еще сильна столь же давняя традиция понимать род плаун широко. При этом по степени родственных отношений виды, входящие в этот род, объединяют в подроды, например в три под рода: уростахис (Urostachys), лепидотис (Lepidotis) и ликоподиум (Lycopodium), каждый из которых, в свою очередь, разбивается на ряд секций.
Большую часть видов рода плаун относят к подроду уростахис. Наземные формы подрода уростахис имеют главные стебли прямостоячие или полегающие, маловетвистые, эпифитные - многократно ветвящиеся стебли, свисающие со ствола растения-хозяина, а частично стелющиеся по нему.
Эпифиты подрода уростахис - растения влажных тропических лесов. Наземные растения этого подрода распространены от тропиков до арктических пустынь и от равнин до снеговой границы в горах (в Андах некоторые виды поднимаются до высоты 4000 и даже 4700 м над уровнем моря). Из упомянутых ранее видов к этому подроду относят плаун баранец, плаун пильчатый, плаун светлый, плаун флегмарию.
Остальные, не относящиеся к подроду уростахис плауны, хотя и составляют в своей совокупности меньшую по числу видов часть рода, более резко отличаются друг от друга, чем виды названного подрода между собой. Их группируют в два или более подродов. Эти виды представляют собой наземные растения с неравновильчато (иногда псевдомоноподиально) ветвящимися ползучими, прямостоячими или лазящими главными стеблями, от которых отходят ограниченные в росте системы боковых побегов. Об облике растений этой группы можно судить по изображенным в книге плаунам булавовидному, годичному, поникшему и сплюснутому. Виды этой группы также широко распространены на земном шаре, встречаясь от тропической зоны (влажные тропические леса) до зоны тундры.
У некоторых плаунов, обладающих ползучими побегами, например у плаунов булавовидного, заливаемого, сплюснутого, растущих на бедных песчаных почвах в редкостойных сосняках, на вересковых пустошах или по берегам озер и окраинам болот в умеренной зоне, отмечено образование "ведьминых кругов", или колец. Эти кольца, появление которых суеверие связывало с местами шабаша нечистой силы, представляют собой группы растений, возникшие при вегетативном размножении одного или нескольких спорофитов, выросших на одном или на нескольких рядом расположенных гаметофитах.
Побеги молодых растений, ветвясь и нарастая в длину, первоначально образуют небольшую куртину. В зависимости от числа проростков, появившихся на одном месте, от скорости роста и степени разветвленности ползучих побегов куртина постепенно приобретает округлое, овальное или вееровидное очертание. Дальнейший центробежный рост главных побегов и ветвей и отмирание побегов в основании приводят к образованию замкнутых или незамкнутых, выпуклых по отношению к месту появления проростков полос - "ведьминых колец". Ширина "ведьминых колец" зависит от скорости роста и продолжительности жизни ползучих побегов.
Встречаясь с различными препятствиями, круги и полосы изменяют свои очертания. Это зависит и от характера почвы, расположенной перед фронтом кольца. У плауна заливаемого, растущего по окраинам болот и на песках по берегам озер, "ведьмины кольца" нередко имеют вид эллипса, длинная ось которого вытянута параллельно краю болота (озера).
Приняв, что увеличение диаметра кольца совершается с более или менее постоянной скоростью, т. е. линейно, определив затем среднегодичный прирост по внешней стороне кольца и измерив диаметр кольца, можно вычислить его приблизительный возраст. Расчеты показывают, что у плауна сплюснутого некоторые кольца, достигающие диаметра 40 м и более, имеют возраст 150-300 лет.
Очень старые клоны (группы растений, возникшие при вегетативном размножении от одного экземпляра) известны и у плаунов с равновильчато ветвящимися побегами из подрода уростахис. У плауна баранца, например, по мере того как верхушка зрелого побега нарастает в длину, основание побега полегает. Отмирание основания побега приводит к тому, что ветви растения обособляются и возникает куртина. Куртина может возникнуть и при росте нескольких рядом расположенных проростков. Диаметр куртины в благоприятных для роста вида условиях (влажные хвойные леса) достигает иногда 2 м, а возраст таких куртин равен нескольким десятилетиям, а иногда и столетиям.
В крайних для вида условиях существования (тундры, высокогорья) прирост у плауна баранца невелик (2-5 мм в год) и куртины возникают редко, так как побег, достигший 15- 40-летнего возраста, погибает из-за неблагоприятных условий окружающей среды.
Установлено, что некоторые виды плауна содержат сильный парализующий яд, сходный по характеру действия с кураре. Вероятно, поэтому позвоночные травоядные животные не употребляют в пищу плауны. Но беспозвоночные животные иногда поедают плауны и без видимых вредных для себя последствий.
Ядовитые свойства плаунов и их своеобразный способ роста ("ведьмины кольца") издавна привлекали к ним внимание и, может быть, в известной степени обусловили широкое и разнообразное, часто оправданное применение этих растений в народной медицине. Использовали их и в ветеринарии, а также как источник получения зеленой, синей и желтой красок. Народное применение плаунов отразилось, например, в таких названиях плауна булавовидного, как зеленка, зеленица, слабительный плывун.
Плауны входят и в современную фармакопею разных стран. Их используют при лечении никотинизма, алкоголизма, глазных болезней. Следует только помнить, что неосторожное применение этих растений при самолечении может привести даже к смертельному для человека исходу. Богатые жирными маслами споры некоторых широко распространенных в умеренной зоне плаунов раньше использовались в пиротехнике (при изготовлении взрывчатых веществ и для получения световых эффектов), а ныне их продолжают использовать при фасонном литье (при соприкосновении с жидким металлом споры, покрывающие поверхность формы, вспыхивают и образовавшийся газ способствует получению гладкой поверхности детали). Использовались и используются споры в медицине в качестве порошка (ликоподия) для присыпки (как детская присыпка, при пролежнях), а также для обсыпки пилюль. В целях охраны запасов плаунов следует, в частности, рекомендовать при заготовках аккуратно отрезать лишь спороносные "колоски" (стробилы), стараясь не повредить остальные части растения. Следует решительно возражать против использования этих растений для изготовления гирлянд и озеленения помещений.
Род содержит всего один вид - филлоглоссум Драммонда (Phylloglossum drummondii), распространенный на западе и юге Австралии, в Тасмании и Новой Зеландии.
Филлоглоссум - маленький клубневой многолетник, высотой всего в несколько сантиметров (рис. 64). На верхушке клубня в конце осени - начале зимы (мае - июне) развивается пучок немногочисленных листьев длиной до 2 см, которые по форме несколько напоминают узкие языки, что и нашло свое отражение в названии рода (от греч. phyllon - лист и glossa - язык). От верхней части клубня отходят 1-3 корня и ножка, кончик которой в конце зимы - начале весны преобразуется в новый клубень.
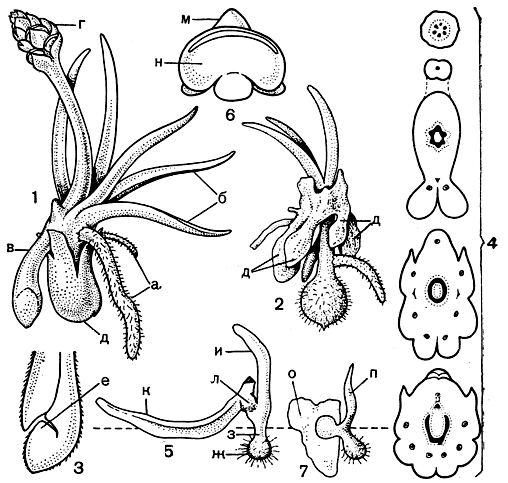
Рис. 64. Филлоглоссум Драммонда (Phylloglossum drummondii): 1 - спороносящее растение: а - корни, б - листья, в - ножка, кончик которой преобразуется в клубень, г - стробил, д - старый, прошлогодний клубень; 2 - молодое растение' видны остатки старых клубней (д), залегавших на разной глубине от поверхности почвы; 3 - схема продольного среза кончика ножки; точка роста (е) молодого клубня, направленная к поверхности почвы, погружена в полость, сообщающуюся с внешней средой с помощью канала; 4 - схемы поперечных срезов растения на разных уровнях, начиная от зоны отхождения листьев и ножки от старого клубня (нижняя схема) и кончая средней частью ножки стробила (верхняя схема) (черным цветом показаны проводящие элементы ксилемы); 5 - вегетативное размножение филлоглоссума: ж - клубень, з - ножка, и - новый лист, к - отломанный и лежащий на поверхности почвы лист, л - протокорм; 6 - спорофилл (м) со вскрывшимся спорангием (к); 7 - заросток (о) и молодой проросток (n)
Вновь образованный клубень располагается обычно на глубине около 1 см от поверхности почвы. Старый клубень и все надземные части растения к весне отмирают, в почве остается лишь молодой клубень.
Зрелое растение образует короткий, длиной до 7 мм, стробил, сидящий на простой, реже вильчато ветвящейся, обычно безлистной ножке длиной до 3-4 см.
Поверхность надземных частей растения покрыта толстой кутикулой. Устьица располагаются на спорофиллах, ножке и стерильных листьях. Корни и молодой клубень в сезон их образования покрыты волосками и обладают эктотрофной микоризой.
Проводящая система растения в той части клубня, от которой отходят листья и ножка стробила, представлена мезархной сифоностелой, ксилема которой в ножке стробила распадается на отдельные участки.
Обычно растение размножается вегетативно с помощью листочков. Хрупкие листья легко отламываются от растения при механическом воздействии животных, при падении на растение веточек деревьев и кустарников и даже от удара переносимых ветром листьев эвкалипта. Отломанные и попавшие на влажную почву листья образуют у своего основания протокорм, который дает начало углубляющейся в землю ножке. Если весна дождливая, то конец ножки до начала сухого сезона успевает преобразоваться в клубенек, из которого по миновании сухого сезона вырастает молодой филлоглоссум.
Желтые в зрелом состоянии спорофиллы сидят на оси стробила супротивно или в мутовках по три. Спорангии крупные, как и у плаунов, почковидные. Споры, как правило, тетраэдрические, с трехлучевым тетрадным рубцом. Строение стенки спорангия и характер поверхности спор указывают на связь филлоглоссума с плаунами из подрода уростахис.
Бесполое размножение у филлоглоссума, по-видимому, преобладает над вегетативным. Именно с помощью спор филлоглоссум расселяется в новые, подходящие для его жизни места. Гаметофиты филлоглоссума надземные, цилиндрические, длиной 2-6 мм. Нижняя, веретеновидная погруженная в почву часть гаметофита незеленая; верхняя, напоминающая иногда по форме лошадиную голову, зеленая, несет гаметангии того же типа, что и гаметангии плауна. Молодой спорофит в первые 1-3 года образует обычно по одному листу.
Для филлоглоссума характерно наивысшее среди плауновидных число хромосом: 2n = 502 - 510.
Филлоглоссум встречается на территориях, покрытых знаменитым австралийским скрабом - почти непроходимыми зарослями жестколистных вечнозеленых кустарников. Он произрастает также в эвкалиптовых лесах и на участках с господством травянистых однодольных. Вместе с филлоглоссумом часто встречаются луковичные и клубневые многолетники, в том числе и споровые, например плаун змеевидный (Lycopodium serpentinum). Тут же можно встретить и папоротник анограмму тонколистную (Anogramma leptophylla), обладающую гаметофитами с клубнями. Строение филлоглоссума и некоторых сопутствующих ему видов указывает на то, что, как и последние, он относится к группе пирофитов (от греч. pyr - огонь) - растений, приспособленных к жизни в местах с периодически повторяющимися пожарами.
В благоприятных для его жизни местах филлоглоссум очень обилен (до 60 экземпляров на 1 дм2). При отсутствии пожаров травянистый ярус в местах обитания филлоглоссума хорошо развивается, и более высокие растения затеняют филлоглоссум, который в таких случаях находится в угнетенном состоянии, ожидая благодетельного для него пожара. Если травяной покров и ветошь в течение многих лет не выгорают, филлоглоссум исчезает в этом месте. Поэтому успешные меры по предотвращению пожаров, а также превращение значительных участков скраба в пастбища сокращают область обитания этого растения, предки которого отделились от основного ствола плауновых, вероятно, уже в мезозое.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'