Оплодотворение (А. Л. Тахтаджян)
Процессу оплодотворения предшествует прорастание пыльцевого зерна на рыльце. Прорастание пыльцы начинается с разбухания пыльцевого зерна и выхода пыльцевой трубки из апертуры - борозды или поры (рис. 28,5). Оболочка пыльцевой трубки состоит преимуществнно из целлюлозы и пектиновых веществ, а также полисахарида каллозы. Рост пыльцевой трубки ограничен ее апикальной зоной, длиной 4 - 7 мкм, состоящей целиком из пектина. Оболочка пыльцевой трубки не сплошная, она пронизана канальцами, через которые проходят цитоплазматические нити - плазмодесмы. Пыльцевая трубка служит ареной самых интенсивных физиологических процессов и очень богата различными ферментами, аминокислотами, гормонами и витаминами, но максимальная напряженность процессов жизнедеятельности сосредоточена в ее кончике. В густой цитоплазме кончика пыльцевой трубки с многочисленными здесь митохондриями и диктиосомами наблюдается наиболее высокая активность ферментов, ростовых веществ, аскорбиновой кислоты, повышенная концентрация белков, аминокислот, жиров, крахмала и т. д., а также интенсивная секреция фосфатаз (ферментов класса гидролаз, катализирующих гидролиз сложных эфиров фосфорной кислоты). Кроме того, выяснилось, что у всех изученных с этой точки зрения цветковых растений кончики пыльцевых трубок имеют утолщенные оболочки, своего рода колпачки, состоящие из тех же веществ, что и оболочка пыльцевых трубок. Предполагают, что в этих колпачках происходит синтез материала оболочки пыльцевой трубки, необходимого для ее роста и, кроме того, они, вероятно, предохраняют кончики пыльцевых трубок от возможных повреждений. Все это характеризует кончик пыльцевой трубки как своего рода гаусторий, активно прокладывающий путь через размягченные им ткани.
В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий (нуцеллус) через микропиле. Это так называемая порогамия (от греч. poros - проход, отверстие и gamos - брак). Реже пыльцевая трубка попадает туда другими, окольными путями (апорогамия). Апорогамия делится на халазогамию и мезогамию. При халазогамии пыльцевая трубка проникает в мегаспорангий через халазу, поднимается вверх и входит в женский гаметофит (зародышевый мешок) поблизости от яйцевого аппарата. Халазогамия (называемая иногда базигамией) наблюдается, например, у березовых, казуарины, ореховых. При мезогамии пыльцевая трубка входит в мегаспорангий сбоку, между халазой и микропйле. Мезогамия наблюдается, например, уцирцеастера (Circaeaster), манжетки, тыквы и других растений. Апорогамия наблюдается исключительно у подвинутых форм со специализированными и обычно упрощенными цветками. Кроме того, семязачатки апорогамных растений не лишены микропиле. Оно всегда имеется, но лишь в виде нефункционирующего, чисто рудиментарного отверстия, которое иногда зарастает. Все это говорит о том, что апорогамия возникла в процессе эволюции из порогамии и является одним из признаков специализации.
По мере роста пыльцевой трубки в нее переходят ядро клетки-трубки и спермин (если генеративная клетка уже разделилась на два спермин). В конце концов они достигают кончика пыльцевой трубки, где, как мы видели, сосредоточено наибольшее количество питательных и физиологически активных веществ. Механизм движения этих клеток не вполне ясен, но, как полагают, они переносятся токами цитоплазмы.
В большинстве случаев ядро клетки-трубки (так называемое "вегетативное" ядро) движется впереди генеративной клетки (или спермиев) и первым доходит до женского гаметофита. Однако во многих случаях ядро клетки-трубки не только не доходит до женского гаметофита, но даже не попадает в пыльцевую трубку и часто дегенерирует еще в пыльцевом зерне. Кроме того, нередко ядро клетки-трубки идет не впереди генеративной клетки или спермиев, а позади них или между ними, иногда сбоку от них.
Если пыльцевое зерно двухклеточное, то генеративная клетка делится только уже внутри пыльцевой трубки по мере ее продвижения к зародышевому мешку. Если пыльца трехклеточная, то в пыльцевую трубку входят уже сформировавшиеся спермин и передвигаются дальше к ее кончику.
Наиболее интересные события начинаются с того момента, когда пыльцевая трубка проникает в женский гаметофит, где изливает свое содержимое. При этом судьба двух сестринских спермиев глубоко различна. Как это было впервые установлено в 1898 г. выдающимся русским цитологом и эмбриологом растений С. Г. Навашиным на двух видах растений - рябчике (Fritillaria orientalis) и лилии (Lilium martagon), один из двух спермиев сливается с яйцеклеткой, другой - со вторичным ядром центральной клетки зародышевого мешка или с одним из двух полярных ядер, если последние еще не слились. Поведение второго спермия было большой неожиданностью и главным моментом в замечательном открытии Навашина. В результате оплодотворения яйцеклетки возникает диплоидная зигота, слияние же второго спермия с полярными ядрами (или со вторичным ядром зародышевого мешка), обычно называемое тройным слиянием, приводит к образованию триплоидного первичного ядра специальной питательной ткани - эндосперма (от греч. endon - внутри и sperma - семя), развивающегося из центральной клетки.
Весь этот процесс, как известно, принято называть двойным оплодотворением. Но читатель может спросить: является ли оплодотворение, происходящее в зародышевом мешке, действительно двойным? Ведь согласно общепринятому в литературе определению, оплодотворение (сингамия) - это процесс слияния мужской и женской половых клеток (гамет) с образованием зиготы, из которой в дальнейшем развивается новый организм. Такое определение оплодотворения можно найти в любом учебнике биологии и в любом энциклопедическом словаре (в том числе в советских энциклопедиях, в Большой и Малой). И даже в известном "Словаре русского языка" С. И. Ожегова (1973) мы читаем: "Оплодотворить. 1. Создать зародыш в комчемн слиянием мужской и женской половых клеток. 2. Перен. Послужить источником развития, совершенствования". Слияние одного из спермиев с яйцеклеткой является, несомненно, оплодотворением, но тройное слияние не является, строго говоря, оплодотворением, так как 1) центральная клетка - не гамета и 2) в результате этого слияния не образуется зигота, из которой в дальнейшем развивался бы новый организм. Очевидно, тройное слияние является оплодотворением лишь в указанном Ожеговым втором, переносном смысле. Другими словами, в выражении "двойное оплодотворение" термин "оплодотворение" применяется в двух разных смыслах - прямом и переносном. Тем не менее выражение "двойное оплодотворение" настолько широко вошло в литературу, что было бы нецелесообразно его заменять (а попытки такого рода делались, в том числе известным немецким ботаником Э. Страсбургером). Достаточно, если мы будем помнить, что речь идет здесь о двух разных биологических процессах, условно объединяемых общим названием.
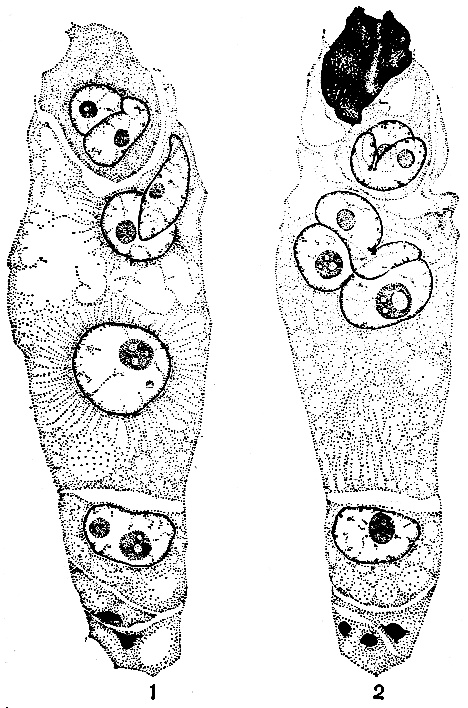
Рис. 45. Двойное оплодотворение в зародышевом мешке гусиного лука Хомутовой (Gagea chomutoviae): 1 - в яйцевом аппарате на срезе зародышевого мешка, изображенного на этом рисунке, оказалась лишь одна яйцеклетка. В ней в тесном контакте с ее ядром находится удлиненный меньший спермий. В центральной клетке более крупный спермий лишь одним своим концом вступил в контакт с верхним полярным ядром; нижнее полярное ядро находится еще в нижней части центральной клетки. Еще ниже видны три антиподы: одна с нормальным ядром, две нижние с дегенерирующими ядрами; 2 - несколько более поздний момент двойного оплодотворения: темная масса в яйцевом аппарате - разрушенная синергида, через которую прошло содержимое пыльцевой трубки; справа от нее - небольшая часть целой синергиды, слева - яйцеклетка. В яйцеклетке в контакте с ее ядром изогнутый меньший спермий. В центральной клетке контакт трех ядер: спермий имеет вид удлиненного ядра, у которого левый конец вздулся вследствие поглощения воды (гидратации), слева верхнее полярное ядро, справа овальное и более крупное нижнее полярное ядро. Внизу антиподы такие же, как на фиг. 1 (увел. 770) (рисунки и текст И. Д. Романова)
Впоследствии процесс оплодотворения изучался у многих других растений и в ряде случаев были найдены те или иные отклонения, иногда очень значительные, от того наиболее распространенного нормального типа, который был открыт С. Г. Навашиным у рябчика и лилии. При двойном оплодотворении (рис. 45) события в зародышевом мешке обычно развиваются следующим образом. Пыльцевая трубка проникает в одну из двух синергид и с силой вскрывается. Содержимое пыльцевой трубки обычно изливается в синергиду, которая при этом разрушается (в ней видны остатки ядра синергиды и ядра клетки-трубки); вторая синергида впоследствии также отмирает (рис. 45, 2). При вскрытии пыльцевой трубки происходит освобождение ферментов, которые лизируют цитоплазму спермиев. Поэтому из синергиды спермин выходят в виде голых ядер (Н. С. Беляева, 1976). Далее оба спермия (точнее, их ядра) вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермин разобщаются: один из них проникает в яйцеклетку и вступает в контакт с ее ядром, другой проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и обоими полярными ядрами. Форма и состояние ядер спермиев могут варьировать. У скерды (Crepis) и некоторых других сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлиненные, иногда извитые, но обычно спермин представляют собой округлые ядра, иногда не отличающиеся по структуре от женских ядер. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермин лишь одной из них участвуют в двойном оплодотворении, спермин же остальных дегенерируют. Случаи оплодотворения яйцеклетки двумя спермиями очень редки. Таков ход событий в "нормальном" восьмиклеточном типе женского гаметофита. В уклоняющихся типах многое происходит по-другому.
Слияние одного из спермиев с двумя свободными или слившимися полярными ядрами (число которых может быть и больше, до 14 у акалифы - Acalypha lanceolata из семейства молочайных) центральной клетки женского гаметофита приводит к образованию эндосперма - питательной ткани, служащей для питания развивающегося из оплодотворенной яйцеклетки зародыша (табл. 2). У многих семейств, особенно у семейств относительно более примитивных, эндосперм продолжает существовать также в зрелом семени и часто бывает даже очень хорошо развит и нередко сильно превосходит по размерам зародыш, но у других семейств весь или почти весь эндосперм потребляется в процессе формирования семени. В первом случае эндосперм используется не только зародышем, он потребляется также во время прорастания семени, во втором случае он служит только для образования зародыша. Но в обоих случаях значение эндосперма очень велико. Предполагают, что тройное (а в некоторых случаях даже более чем тройное) слияние служит лишь для увеличения количества ядерного вещества, а это ускоряет деление ядер эндосперма и тем самым стимулирует быстрое развитие этой уникальной питательной ткани. "Оплодотворение" полярных ядер (если это можно назвать оплодотворением), по всей вероятности, возникло в результате крайнего упрощения и миниатюризации женского гаметофита, обычно почти полностью лишенного запаса питательных веществ, и представляет собой своеобразное приспособление для быстрой компенсации недостатка питательных веществ. Следовательно, как уже давно указывалось в литературе, возникновение эндосперма коррелятивно связано с упрощением женского гаметофита и является своего рода компенсаторным приспособлением. Оно позволило достичь максимальной экономии материала на построении женского гаметофита цветковых растений и максимального ускорения его развития.
Однако у подостемовых, рогульниковых и у подавляющего большинства самого большого в растительном мире семейства орхидных эндосперм вовсе не образуется. Это можно объяснить только тем, что вырабатываются новые приспособления, которые исполняют функцию эндосперма более эффективно. У орхидных подвесок зародыша часто развивается в гаусторий, который через микропиле проникает в семяножку и может погрузиться даже в плаценту. Этот гаусторий извлекает из окружающих клеток питательные вещества и доставляет их развивающемуся зародышу и таким образом делает ненужным наличие эндосперма.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'