Порядок горечавковые (Gentianales)
Семейство логаниевые (Loganiaceae) (Н. Н. Имханицкая)
Это семейство включает немногим более 20 родов и около 500 видов, распространенных главным образом в тропических и субтропических странах, лишь немногие представители заходят в умеренные области. Род гелсемиум (Gelsemium) проникает в Атлантическую Северную Америку и Южный Китай. В Новой Зеландии встречаются виды логании (Logania), гениостомы (Geniostoma) и митрасакмы (Mitrasacme), распространенной на север до Центральной Японии и Кореи. Дефонтэния (Desfontainia), произрастающая в Андах Чили и Перу, достигает мыса Горн. Лишь род стрихнос (Strychnos), самый крупный в семействе (более 200 видов), широко распространен в тропических и субтропических областях земного шара. Большинство же родов ограничено Новым или Старым Светом. Род лабордия (Labordia) эндемичен для Гавайских островов.
Большинство логаниевых - древесные растения: деревья (до 55 м высотой), иногда с досковидными корнями, кустарники, часто лазящие, или лианы, реже полукустарники или травы, однолетние и многолетние. Многие виды стрихноса - лианы, обычно снабженные крючковидно закрученными одревесневающими усиками или шипами (рис. 183); некоторые африканские виды достигают в длину 120 м и в высоту 45 м, взбираясь на верхушки крупных деревьев тропического леса. Многие виды фагрэи (Fagraea, Южная, Восточная и Юго-Восточная Азия, Полинезия, Австралия) - эпифитные или полуэпифитные кустарники или деревца. Поселяясь на стволах высоких деревьев, они плотно обвивают их своими корнями. Если корни достигают почвы, растение становится полуэпифитным. Некоторые виды этого рода - мирмекофильные растения. Они снабжены нектарниками в основании черешков, на листовых пластинках или чашечке. Крупные ушки на черешке листа у некоторых видов, например у фагрэи ушковидной (F. auriculata), дают убежище муравьям.
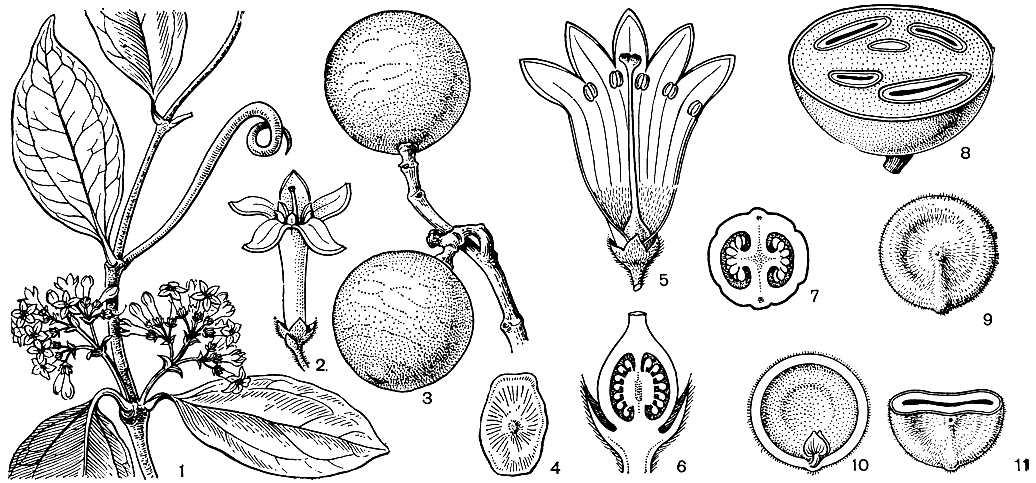
Рис. 183. Логаниевые. Стрихнос Игнатия (Strychnos ignatii): 1 - фрагмент побега с цветками, виден крючковидно закрученный усик; 2 - цветок; 3 - фрагмент побега с плодами; 4 - семя. Чилибуха (S. nuxvomica): 5 - цветок (развернут венчик); 6 - продольный разрез гинецея; 7 - поперечный разрез завязи; 8 - поперечный разрез плода (видны погруженные в мякоть семена); 9 - семя; 10 - продольный разрез семени (виден зародыш); 11 - поперечный разрез семени
Листья у логаниевых обычно кожистые, иногда крупные (более метра в длину у некоторых видов антоклейсты - Anthocleista и фагрэи); они супротивные, реже почти мутовчатые, простые, цельнокрайные, иногда зубчатые, соединенные у основания поперечной стипулярной линией или прилистникоподобным влагалищем, или реснитчатым ободком, иногда с маленькими прилистниками. У американского рода плокосперма (Plocosperma) листья без прилистников. Это растение замечательно своим опушением. Кроме простых одноклеточных волосков, заполненных известью с цистолитами в прилегающих эпидермальных клетках, имеются двуклеточные веретеновидные железистые волоски. У всех представителей семейства, за исключением плокоспермы и дефонтэнии, присутствует интраксилярная флоэма. Многие логаниевые содержат яды - алкалоиды (стрихнин, бруцин, курарины, гелсемин и др.).
Большинство видов семейства - обитатели низинных влажных тропических лесов. Они растут главным образом в подлеске или в нижнем древесном ярусе некоторые лианы достигают верхнего полога леса. Особенно обильны логаниевые по берегам рек, в периодически: затопляемых водой долинах и на болотах. Деревья антоклейсты благородной (Anthocleista, nobilis) из тропической Африки в случаях, когда они растут в заболоченных лесах, образуют ходульные и дыхательные корни. Встречаются логаниевые и в горных лесах, как правило, не выше 2000 м над уровнем моря. Некоторые виды произрастают в областях с сезонным климатом - в галерейных и сухих тропических лесах, зарослях жестколистных кустарников, в саваннах. Невысокие деревца ряда африканских стрихносов (например, стрихноса безвредного - Strychnos innocua и стрихноса колючего - S. spinosa) - обитателей саванн - в сухой сезон сбрасывают листву. Многие виды фагрэи предпочитают открытые местообитания, опушки леса, светлые леса. Логаниевые можно встретить на морских побережьях, в прибрежных лесах, в мангровых болотах. Нередки они и во вторичных лесах, в зарослях аланг-аланга.
Цветки у логаниевых в верхоцветных соцветиях или реже одиночные, иногда крупные, обоеполые, редко однополые, актиноморфные, большей частью 5- или 4-членные. Брактеоли обычно мелкие, иногда чешуевидные, многочисленные (антония - Antonia) или крупные и петалоидные (бониуния - Bonyunia). Чашечка 5-4(2)-лопастная или 5-4(2)-раздельная, лопасти одинаковые или одна из них увеличена и лепестковидная (устерия - Usteria), в бутоне черепитчатые. Венчик сростнолепестный, воронковидный, колокольчатый или колесовидный, с короткой или длинной трубкой, с 5-4, реже 6-5 (плокосперма) или даже 8-16 (антоклейста) лопастями, в бутоне створчатыми, черепитчатыми или скрученными. Тычинки обычно в одинаковом числе с лопастями венчика (редко одиночные - устерия) и чередуются с ними, прикреплены к трубке венчика или к зеву, пыльники 2-гнездные, открываются продольно. Гинецей состоит из 2, реже 3 или 5 (дефонтэния) плодолистиков; короткий столбик с головчатым, 2-лопастным или реже дважды лопастным рыльцем. Завязь верхняя, редко полунижняя, обычно 2-гнездная, редко 1-, 3- или 5-гнездная (дефонтэния); семязачатки в каждом гнезде от многочисленных до одиночных, анатропные или амфитропные. У плокоспермы завязь одногнездная с 4 семязачатками, сидящими попарно.
Крупные одиночные цветки или соцветия из многочисленных мелких невзрачных душистых цветков привлекают насекомых. Цветки фагрэи протандричны. Они раскрываются после захода солнца, испуская сильный аромат. Цветение продолжается два дня. Вначале кремово-белые, на 2-й день цветки приобретают желтую или оранжевую окраску. Они опыляются ночными бабочками или пчелами. Некоторые крупноцветковые виды посещаются только птицами, а возможно, и летучими мышами. Гигантские цветки фагрэи ушковидной с нежным запахом жасмина, белый венчик которых длиной до 30 см и более 30 см в поперечнике, опыляются насекомыми и птицами. У фагрэи душистой (F. fragrans), растения открытых и болотистых низин Малезии, каждый год с исключительной регулярностью наблюдается массовое цветение в мае и начале июня в октябре или ноябре бывает второе, не столь пышное цветение" У родов мостуэя (Mostuea) и гелсемиум известна гетеростилия. Наиболее обычные посетители цветков последнего рода - пчелы и шмели. Спигелия противоглистная (Spigelia anthelmia) - самоопыляемое растение.
Плод у логаниевых - костянка, ягода или коробочка, обычно септицидная, 2-створчатая, редко локулицидная или раскрывающаяся круговой щелью. Семена обычно мелкие, реже крупные, в сухих плодах обычно крылатые, с прямым зародышем в мясистом эндосперме. У плокоспермы линейно-веретеновидная коробочка обычно имеет единственное линейное семя на верхушке с густым хохолком из длинных волосков.
Сочные ягоды видов фагрэи поедают птицы, циветты, летучие мыши, а также, очевидно, плодоядные муравьи. Крупный зрелый плод фагрэи ушковидной (длиной до 15 см), раскалываясь на верхушке на 4 неправильные лопасти, обнажает многочисленные мелкие семена, погруженные в сладкую оранжевую мякоть, привлекая птиц. Мелкими красными или оранжевыми ягодами фагрэи душистой питаются летучие мыши, которые в пору плодоношения этого растения слетаются в огромном количестве.
Ягоды стрихноса диаметром от 1 до 20 см, зеленые, коричневые, белые, черные или часто желтые, оранжевые или красные, покрытые обычно кожистой или твердой, деревянистой кожурой, заключают в нежной мякоти часто крупные (длиной до 2,5 см) плоские семена. У многих стрихносов горькая мякоть плода и особенно семена ядовиты. Однако известны некоторые африканские виды, как стрихнос безвредный, имеющие сладкую съедобную мякоть. Сочной мякотью ягод чилибухи (Strychnos nuxvomica), стрихноса Игнатия (S. ignatii) и некоторых других видов питаются птицы, особенно птицы-носороги, фазан аргус, туканы, а также обезьяны, летучие мыши, циветты, музанги, белки, слоны и другие животные.
Костянки невбургии (Neuburgia), очевидно, переносятся водой. Легкие крылатые семена видов гелсемиума и норисии (Norrisia) приспособлены к распространению ветром.
Логаниевые - в высшей степени гетерогенная группа. Она включает 7 подсемейств, выделяемых главным образом на основании почкосложения лопастей венчика и типа плода. Иногда этим подсемействам придают ранг семейств. Систематическое положение монотипных родов дефонтэния и плокосперма, образующих соответственно подсемейства дефонтэвиевых (Desfontainioideae) и плокоспермовых (Plocospermatoideae), до сих пор остается неясным.
Более половины всех видов включают подсемейства логаниевые (Loganioideae, 5 родов и более 100 видов) и стрихновые (Strychnoideae, 3 рода и более 220 видов), распространенные в тропических и субтропических, реже в умеренных областях земного шара. Виды первого подсемейства - деревья, кустарники, лианы и травы с черепитчатыми или скрученными в бутоне лопастями венчика; плод - коробочка. Виды стрихновых - деревья, кустарники и лианы со створчатыми лопастями венчика. Плод - ягода.
Небольшие подсемейства поталиевые (Potalioideae) и антониевые (Antonioideae), ограниченные тропическими областями, включают древесные растения. У родов поталия (Potalia, Южная Америка), фагрэя (35 видов) и антоклейста (Африка, Мадагаскар, Коморские острова), составляющих первое подсемейство, лопасти венчика скрученные, плод - ягода; у видов норисии (Малезия), устерии (Западная Африка), антонии и бониунии (Южная Америка) - представителей подсемейства антониевых - лопасти створчатые, плод - коробочка.
К подсемейству спигелиевых (Spigelioideae, 4 рода и около 100 видов) относятся травянистые растения или реже полукустарники тропических, субтропических и умеренных областей земного шара. Половина видов принадлежит американскому роду спигелия (Spigelia).
Многие виды стрихноса содержат большое количество сильно токсичных алкалоидов. Семена чилибухи, или рвотного ореха (Индия, Цейлон, Индокитай, Малайя), и стрихноса Игнатия (Тонкий, Малезия) - главный источник стрихнина и бруцина. Экстракты из коры нескольких южноамериканских видов, включая стрихнос ядоносный (Strychnos toxifera), используются индейцами как главная составная часть парализующего стрельного яда кураре. Стрельный яд также получают из коры и корней некоторых африканских и азиатских видов. Древесина, кора, корни видов стрихноса, а также других представителей семейства, в том числе сильно ядовитых растений гелсемиума вечнозеленого (Gelsemium sempervirens), спигелии противоглистной, используются в медицине. Семена стрихноса жаждущих (Strychnos potatorum) применяют в Индии для очистки питьевой воды.
Древесина ряда видов фагрэи, твердая и прочная, находит применение при строительстве домов, мостов, каноэ, изготовлении инструментов.
Некоторые логаниевые культивируются как декоративные растения.
Семейство мареновые (Rubiaceae) (Н. Н. Имханицкая)
Мареновые - одно из самых крупных семейств цветковых растений - насчитывает 450-500 родов и 6000-7000 видов, широко распространенных в обоих полушариях, главным образом в тропических странах, хотя немало представителей встречается в умеренных и даже в холодных областях.
Большинство мареновых - древесные растения: деревья высотой до 40-45 м, иногда с досковидными корнями, кустарники, часто вьющиеся или древесные лианы, реже кустарнички, полукустарники или травы, прямостоячие, ползучие или вьющиеся. Высокие деревья митрагины крупноприлистниковой (Mitragyna stipulosa) в болотистых лесах тропической Африки образуют коленчатые дыхательные корни. Среди мареновых немало колючих растений. Лианы, иногда достигающие вершин крупных деревьев тропического леса, обвивают деревья или взбираются, как ункария (Uncaria), с помощью крючкообразно изогнутых, преобразованных в орган лазания цветоносов, которые после сжимания опоры разрастаются и одревесневают. Высоко в кронах деревьев в тропическом лесу Юго-Восточной Азии произрастают виды мирмекодии (Myrmecodia) и гиднофитума (Hydnophytum), курьезные эпифиты. Стебель этих полукустарников образует крупный клубень, часто усаженный шипами, пронизанный многочисленными галереями и ходами, в которых обычно поселяются муравьи (рис. 184). Постоянно населены муравьями также веретеновидно вздутые полые междоузлия стеблей ниже соцветия у видов мир меконауклеи (Myrmeconauclea) и ряда других видов, а также пузыревидные вздутия в основании листьев ремихии пузыреносной (Remijia physophora) и дуройи мешконосной (Duroia saccifera) - растений бассейна Амазонки.
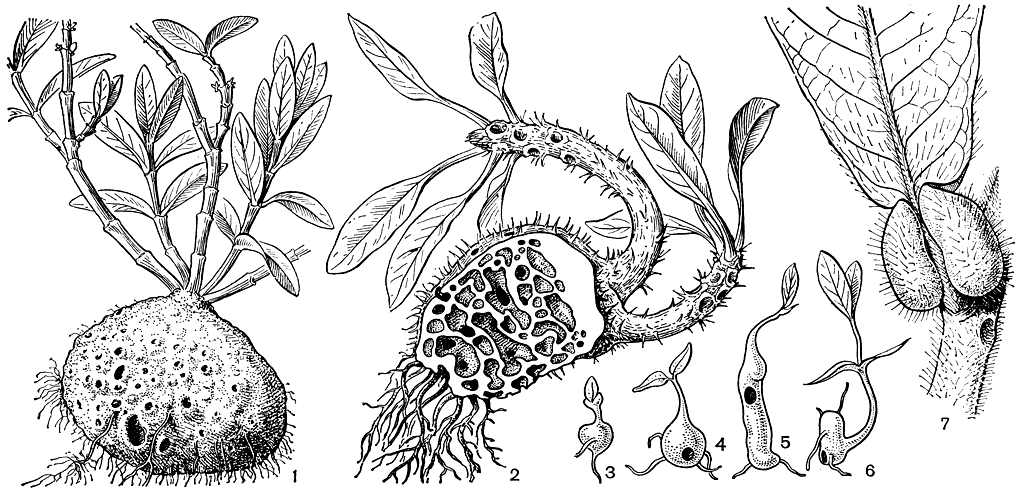
Рис. 184. мареновые. Гиднофитум муравьиный(Hydnophytum formicarum): 1 - общий вид растения с клубнем. Мирмекодия колючая (Myrmecodia echinata): 2 - общий вид растения, на продольном разрезе клубня видны полости, в которых обитают муравьи; 3-6 - проростки на различных стадиях развития. Дуройя мешконосная (Duroia saccifera): 7 - фрагмент основания листовой пластинки с формикариями
Листья у мареновых супротивные или мутовчатые, простые, цельные, редко перистонадрезные, снабженные прилистниками. Они варьируют от крошечных чешуевидных или игловидных до крупных длиной до 140 см. Прилистники расположены или между черешками листьев (интерпетиолярные), или между черешками и стеблем (интрапетиолярные). Часто они, срастаясь друг с другом и черешками, охватывают побег в виде раструба. Иногда прилистники образуют колпачок, защищающий верхушку стебля, позднее сбрасываемый. Прилистники цельные, лопастные, зубчатые или щетинковидные, реже листовидные, неотличимые по форме и величине от листьев, что создает видимость мутовчатого листорасположения (триба мареновых - Rubieae). На листьях видов паветты (Pavetta) и психотрии (Psychotria) имеются "бактериальные узелки". Бактерии обеспечивают растение-хозяина ростовыми веществами, без бактерий оно неспособно к нормальному росту.
Виды мареновых богаты алкалоидами (кофеин, хинин, эметин и др.)- Многие виды способны накапливать алюминий.
Цветки мареновых редко одиночные, как у гардении (Gardenia, табл. 48) и рандии (Randia), обычно в верхоцветных соцветиях, иногда собраны в шаровидные головки, в которых цветки свободные или, как у моринды (Morinda), сросшиеся. У ряда тропических видов известна каулифлория. Цветки обычно обоеполые, иногда однополые или полигамные, актиноморфные или редко слабозигоморфные (двугубые, как у энрикезии - Henriquezia), 5-4-членные. Чашечка развита слабо, (3)4- 5(8)-зубчатая или лопастная, в бутоне открытая, иногда одна или несколько лопастей чашечки листовидно разрастаются и ярко окрашены. Венчик сростнолепестный, 5-4 (редко 8-10)-лопастный, колесовидный, воронковидный, колокольчатый, реже кувшинчатый или трубковидный, в бутоне створчатый, скрученный или черепитчатый. Трубка венчика длинная (до 25 см) и узкая или короткая. Тычинки, как правило, в одинаковом числе с лопастями венчика и чередуются с ними; они прикреплены к трубке венчика или к зеву. Пыльники 2-гнездные, иногда разделены поперечными перегородками, вскрываются продольно, реже верхушечными порами, интрорзные или экстрорзные. В основании столбика имеется нектарный диск.
Гинецей обычно из 2, редко большего числа плодолистиков, с тонким нитевидным столбиком; рыльце 2 -многолопастное с линейными, лопатчатыми или булавовидными лопастями или головчатое. У многих видов известна гете-ростилия. Завязь обычно нижняя, как исключение, почти полностью верхняя или полунижняя, 2-гнездная, редко многогнездная или 1-гнездная с париетальной плацентацией (гардения), с 1 или многими семязачатками в каждом гнезде. Семязачатки висячие или горизонтальные.
Многие виды мареновых - насекомоопыляемые растения. Кроме яркой окраски цветков, обильно выделяющегося нектара, нежного аромата, для привлечения насекомых служат крупные белые прицветники, ярко окрашенные оси соцветия у некоторых мареновых, а также белые, желтые, оранжево-красные или огненно-пурпурные листовидные лопасти чашечки у видов муссенды (Mussaenda), каликофиллума (Calycophyllum), погонопуса (Pogonopus) и других растений.
Приспособления к перекрестному опылению у мареновых разнообразны - от протандрии, обычной в семействе, до диморфной гетеростилии. У некоторых гетеростильных видов наблюдается самонесовместимость. Однополость и двудомность, крайне редкие в семействе, очевидно, возникли как результат крайней гетеростилии. У видов подсемейства иксоровых (Ixoroideae) длиннотрубчатые, опыляющиеся бабочками цветки протандричны. Интрорзные пыльники вскрываются в бутоне, и пыльца отлагается на наружной поверхности рыльцевой головки, которая находится в непосредственном контакте с пыльниками. После распускания цветка насекомые-опылители могут перенести эту пыльцу на цветки, уже достигшие стадии зрелых рылец. Затем происходит развертывание лопастей рыльца, и цветок вступает в женскую стадию, продолжающуюся несколько дней.
У посокерии широколистной (Posoqueria latifolia) из т.ропической Америки крупные белые зигоморфные цветки, приспособленные к опылению ночными бабочками, имеют взрывной механизм выбрасывания пыльцы. Пыльники 5 тычинок, плотно смыкаясь, образуют шар. Раскрываясь в бутоне, они высыпают пыльцу в полость шара. Тычиночные нити находятся в состоянии сильного натяжения, две из них дугообразно изогнуты и особенно раздражимы. При прикосновении хоботка бабочки к их средней части одна пара тычинок отходит направо, другая - налево, а пятая, непарная, изгибаясь вверх, отшвыривает комок пыльцы на хоботок насекомого, закрывая при этом своей нитью вход в трубку венчика. У фуопсиса длинностолбикового (Phuopsis stylosa) выбрасывание пыльцы производится в момент раскрытия венчика благодаря быстрому выпрямлению столбика, несущего на наружной бородавчатой поверхности рыльцевой головки пыльцу из вскрывшихся в бутоне пыльников.
У нертеры (Nertera) обычно самоопыление. У некоторых видов мареновых оно имеет место при отсутствии опылителей. Мелкие клейстогамные цветки мирмекодии клубневой (Myrmecodia tuberosa) завязывают в обилии всхожие семена.
Наиболее обычные опылители цветков мареновых - дневные и ночные бабочки, пчелы и шмели. У многих тропических видов нектар находится на дне длинной трубки венчика и доступен лишь длиннохоботковым насекомым и колибри. Виды мареновых, опыляемые сумеречными или ночными бабочками, имеют белые цветки, хорошо заметные насекомым в темноте. Они раскрываются ночью или в сумерки, распространяя сильный аромат. Мелкие протандричные цветки подмаренника (Galium) с неглубоко скрытым нектаром посещаются чаще всего короткохоботковыми насекомыми, главным образом мухами.
Красные, длиннотрубчатые, лишенные запаха, но с обильным нектаром цветки манеттии вздутой (Manettia inflata) из Южной Америки, бурхелии буйволиной (Burchellia bubalina) из Южной Африки и некоторых других мареновых опыляются колибри. Крупные висячие цветки с сильным ароматом и ночным цветением африканских видов ротмании (Rothmannia) и гардении приспособлены к опылению летучими мышами. Виды копросмы (Goprosma) и антоспермума (Anthospermum), очевидно, ветроопыляемые растения.
Плоды мареновых диаметром от 2 мм до 15-20 см, сочные (табл. 48) или сухие, ягоды или костянки, коробочки (локулицидные или септицидные), сухие, нераскрывающиеся или дробные, иногда образующие соплодия, (1)2 - многосемянные. Семена часто мелкие, обычно с прямым зародышем и с эндоспермом, реже эндосперм почти или полностью отсутствует (подсемейства геттардовых - Guettardoideae и энрикезиевых - Henriquezioideae), они прямые, висячие или горизонтальные, иногда крылатые или с ариллусом (цефалаптус - Cephalanthus), у некоторых мареновых погружены в студенистую плацентарную массу (гардениевые - Gardenieae).

Таблица 48. Мареновые: 2 - марена цветущая (Rubia florida), Западный Копетдаг, урочище Даната

Таблица 48. Мареновые: 3 - адина красноватая (Adina rubella), Сухуми
Белые, желтые, оранжевые, красные, фиолетовые, голубые или черные ягоды, костянки и сочные соплодия многих мареновых охотно поедают различные виды птиц (например, голуби, гуахаро, попугаи, дрозды), летучие мыши и другие животные, распространяя семена. Зеленые или желтоватые плоды гардении трубконосной (Gardenia tubifera, Maлезия), неправильно раскалываясь, обнажают оранжево-алую внутренность перикарпа и яркую сочную душистую массу, поедая которую птицы, обезьяны, белки, музанги проглатывают погруженные в нее семена. Плодами мареновых питаются также жирафы, антилопы, буйволы, лани, дикие козы и овцы, крупный рогатый скот. Семена касасии клузиилистной (Casasia clusiifolia) распространяются игуанами. Сухие плоды подмаренника трехрогого (Galium tricornutum) растаскиваются хомяками. Мелкие сочные плоды мирмекодии и гиднофитума, очевидно, транспортируются муравьями. Мареновые, обитающие на морских побережьях (как геттарда великолепная - Guettarda speciosa, моринда лимонолистная - Morinda citrifolia и др.), берегах илистых лагун в приливно-отливной зоне и мангровых болотах (сцифифора гидрофилаксовая - Scyphiphora hydrophylacea) распространяются морскими течениями. Плавучесть семян этих растений обеспечивается присутствием воздухоносной ткани в семенной кожуре, опробковением плодов или наличием заполненной воздухом полости в косточках. Семена некоторых тропических лесных трав разносятся потоками дождевой воды. Мелкие легкие крылатые семена (хинное дерево - Cinchona, ункария, симира - Simira и др.), а также веретеновидные, на верхушке с хохолком из длинных волосков (у гиллии паразитной - Hillia parasitica) распространяются ветром. У эрнестимейеры (Ernestimeyera) роль крыла выполняют разрастающиеся лопасти-чашечки. Сухие плоды некоторых видов подмаренника, ясменника (Asperula) и видов трибы антоспермовых (Anthospermeae), усаженные крючковатыми щетинками, разносятся млекопитающими. Семена мареновых требуют для прорастания от одной недели до нескольких месяцев. У некоторых видов они прорастают на материнском растении.
Ареал этого обширного семейства охватывает все континенты, за исключением Антарктиды, протягиваясь от арктических стран на севере (подмаренник) до Огненной Земли, Фолклендских островов и острова Тристан-да-Кунья на юге (подмаренник, нертера). В тропиках виды почти космополитного рода подмаренник (400 видов) встречаются высоко в горах, поднимаясь до 4350 м над уровнем моря. Наибольшее разнообразие мареновых наблюдается в тропических областях, где сосредоточено подавляющее большинство родов и видов. Наряду с монотипными или олиготипными родами (а их почти половина в семействе) имеется немало крупных, богатых видами родов, как пантропический род психотрия (700), широко распространенные в тропических странах иксора (Ixora, 400) и рандия (200-300), ограниченные Старым Светом гардения (250) и паветта (400) или приуроченные к Новому Свету ронделетия (Rondeletia, 120) и паликурея (Palicourea, 200).
Многие мареновые произрастают во влажных низинных и горных тропических лесах, встречаясь часто в подлеске или нижнем древесном ярусе, реже достигают верхнего полога леса. Они обычны по берегам рек, на опушках, в заболоченных лесах или на болотах. Некоторые виды растут на морских побережьях, по берегам эстуариев, на мангровых болотах. Многие мареновые приспособились к жизни в областях с резко выраженным сухим сезоном. Они встречаются в африканских саваннах, южноамериканских кампосах, сухих тропических лесах, зарослях жестколистных кустарников и даже в полупустынях и пустынях. Многие виды произрастают в горах, иногда на больших высотах. Мареновые нередки и во вторичных формациях. Некоторые виды широко расселились по земному шару как сорные растения.
Мареновые - вполне естественное семейство. Оно состоит из 5 подсемейств, двум из них - цинхоновым (Cinchonoideae) и мареновым (Rubioideae) - принадлежит большинство видов. Различение подсемейств во многом основано на наличии или отсутствии рафидов (игольчатых кристаллов оксалата кальция), строении волосков, семенной кожуры, морфологии пыльцевых зерен.
Подсемейство цинхоновых включает деревья, кустарники, лианы, редко травы тропических областей обоих полушарий. Важнейшие роды его -хинное дерево, митрагина (Mitragyna), ункария, ронделетия, науклея (Nauclea), муссенда.
У представителей небольшого подсемейства геттардовых (Guettardoideae) семена без эндосперма (или лишь со следами эндосперма). Это деревья или кустарники тропических областей. Важнейший род подсемейства геттарда (Guettarda) включает свыше 80 видов, распространенных главным образом в тропической Америке.
Видам подсемейства иксоровых (Ixoroideae) свойствен особый механизм опыления (см. выше). Это деревья и кустарники тропических стран Старого и Нового Света. Среди важнейших родов подсемейства - кофейное дерево (Coffea, табл. 48), иксора, рандия, паветта.

Таблица 48. Мареновые: 1 - кофейное дерево аравийское (Coffea arabica), оранжереи Ботанического института АН СССР в Ленинграде
Самое крупное подсемейство - мареновые (Rubioideae) отличается от других подсемейств присутствием рафидов и диморфной гетеростилии. Роды психотрия, гамелия (Hamelia), паэдерия (Paederia), моринда, фарамея (Faramea) - деревья, кустарники или лианы тропических областей. Трибы мареновых, антоспермовых и гедиотовых (Hedyotideae) с преобладанием травянистых форм распространены главным образом в умеренной зоне северного и южного полушария.
Подсемейство энрикезиевых (Henriquezioideae) с 2 небольшими бразильскими родами энрикезия и платикарпум (Platycarpum) значительно отличается от остальных мареновых зигоморфным двугубым венчиком, отсутствием эндосперма, а также строением спермодермы и спородермы и иногда рассматривается как отдельное семейство, сближаемое с бигнониевыми (Bignoniaceae).
Семейство мареновых богато полезными растениями, важнейшими из которых являются кофейное дерево и хинное дерево (Cinchona). Род Coffea насчитывает 40 видов в тропических областях Старого Света, главным образом в Африке. Наибольшее экономическое значение имеет кофейное дерево аравийское (C. arabica, рис. 185), широко культивируемое во многих странах земного шара. Родина этого растения - Эфиопия, где и теперь в речных долинах Абиссинского нагорья на высоте 1000-2000 м над уровнем моря можно встретить заросли кофейного дерева. Это вечнозеленый кустарник или деревце высотой не более 8-10 м. Семена содержат алкалоид кофеин, которому кофе обязан своим тонизирующим действием. В небольшом количестве культивируется и некоторые другие африканские виды этого рода.
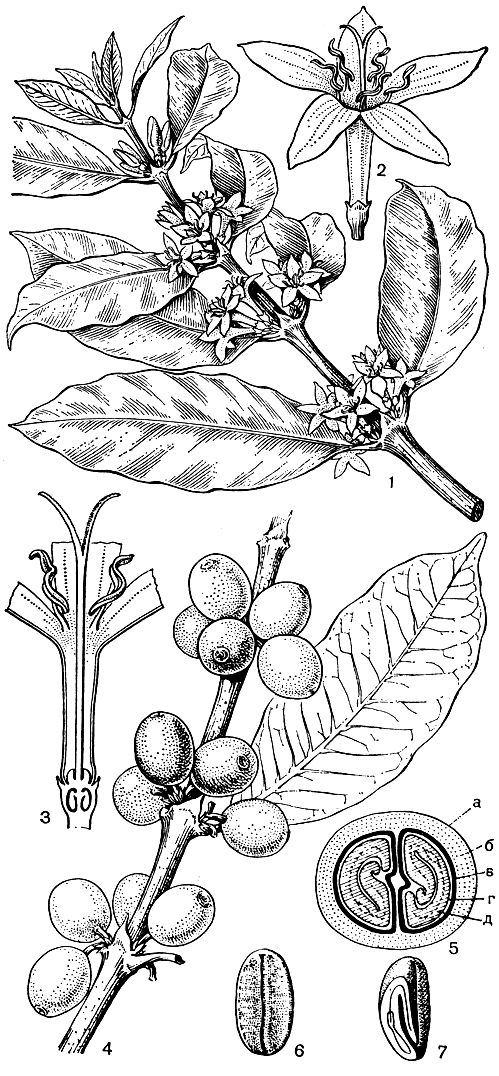
Рис. 185. Кофейное дерево аравийское (Coffea arabica): 1 - цветущий побег; 2 - цветок; 3 - продольный разрез цветка; 4 - фрагмент побега с плодами; 5 - поперечный разрез плода с двумя семенами; а - экзокарпий, б - мезокарпий, в - эндокарпий (пергаментная оболочка), г - семенная кожура (серебристая пленка), д - эндосперм; 6 - семя без семенной кожуры; 7 - продольный разрез семени (виден зародыш)
Хинное дерево включает также около 40 видов. Они произрастают в Андах Южной Америки между 10° с. ш. и 19° ю. ш. на обрывистых, трудно доступных восточных склонах гор на высоте от 1000 до 3300 м над уровнем моря. Это вечнозеленые деревья высотой до 25 м, реже кустарники. Горькая кора содержит до 30 различных алкалоидов, важнейшим из которых является хинин - целебное средство при лечении малярии. Из нескольких ценных культивируемых видов хинное дерево Леджера (Cinchona ledgeriana, рис. 186), родом из Боливии - самый богатый алкалоидами вид. Для лечения малярии применяют также кору некоторых других мареновых.
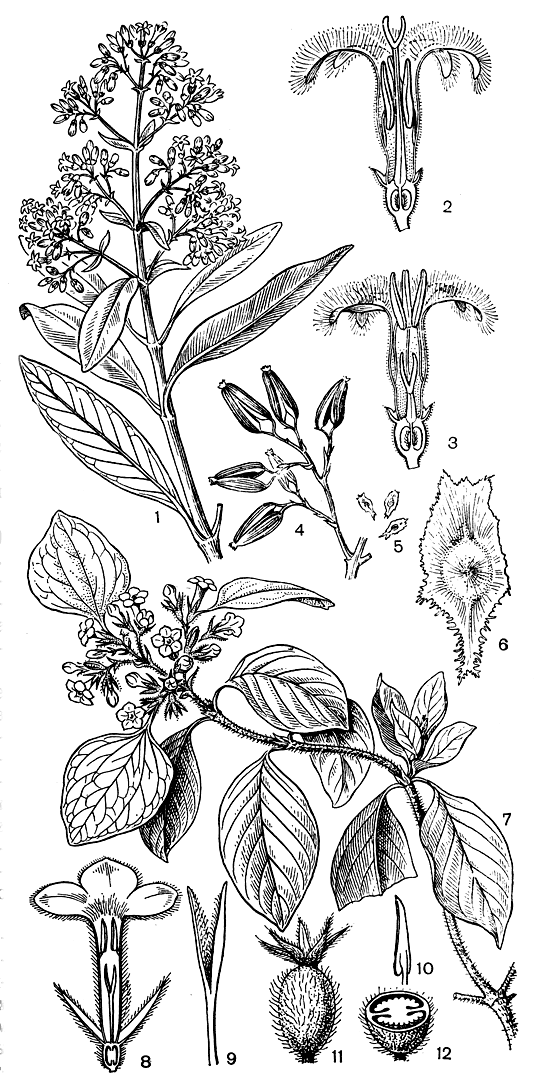
Рис. 186. Мареновые. Хинное дерево Леджера (Cinchona ledgeriana): 1 - побег с цветками; 2 - продольный разрез цветка длинностолбиковой формы; 3 - продольный разрез цветка короткостолбиковой формы; 4 - фрагмент побега с плодами; 5-6 - крылатые семена (5 - натуральная величина; 6 - увелич.). Муссенда краснолистная (Mussaenda erythrophylla); 7 - цветущий побег, видны листовидно разросшиеся лопасти чашечки; 8 - продольный разрез цветка; 9 - столбик с двулопастным рыльцем; 10 - тычинка; 11 - молодой плод; 12 - поперечный разрез завязи
Рвотный корень, или ипекакуана (Cephaelis ipecacuanha, Psychotria ipecacuanha, рис. 187),- важнейшее лекарственное растение. Это невысокий полукустарник девственных тенистых лесов Бразилии, Колумбии и Центральной Америки. Четковидные корни этого растения содержат эметин, цефаэлин и другие алкалоиды. Как заменители ипекакуаны, применяют некоторые другие мареновые, содержащие эметин.
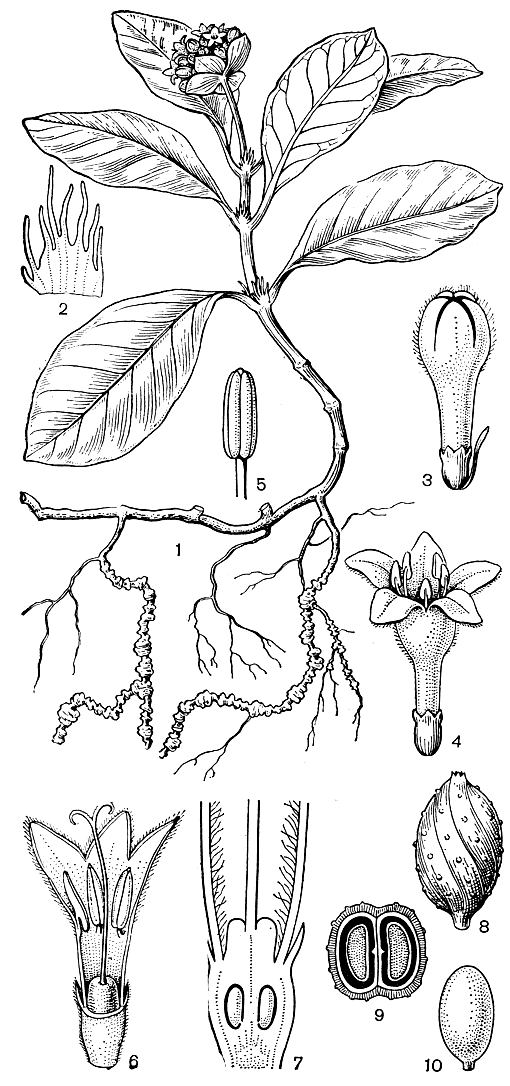
Рис. 187. Ипекакуана (Cephaelis ipecacuanha): 1 - общий вид цветущего растения; 2 - прилистник; 3 - бутон; 4 - короткостолбиковый цветок; 5 - тычинка; 6 - продольный разрез длинностолбикового цветка; 7 - продольный разрез гинецея; 8 - плод; 9 - поперечный разрез плода; 10 - семя
Листья и молодые побеги малайской лианы гамбир (Uncaria gambir) - ценный источник дубильных веществ, используемых при выделке кож. Целый ряд мареновых хорошо известен как красильные растения (например, виды моринды и гардении). С незапамятных времен культивируют марену красильную (Rubia tinctorum) - средиземноморское растение, корневища которого содержат красную краску - крапп. Получаемые тона красного цвета для ковров, тканей и живописи сохраняются в течение столетий.
Некоторые тропические виды культивируют из-за съедобных плодов. Используют древесину ряда мареновых (например, африканской билинги - Nauclea diderrichii).
Многие мареновые известны как красивейшие декоративные растения (виды иксоры, бувардии - Bouvardia, портландии - Portlandia, рандии и др.). К наиболее излюбленным растениям в культуре относится гардения жасминновидная (Gardenia jasminoides) с крупными белыми душистыми, часто махровыми цветками. В Китае и Японии цветки гардении используют для ароматизации чая.
Семейство кутровые (Apocynaceae) (Н. Н. Цвелев)
Жителям внетропических стран семейство кутровые знакомо по двум широко культивируемым декоративным растениям - олеандру (Nerium oleander) и барвинку малому (Vinca minor). И действительно, подавляющее большинство видов этого семейства (их около 2000, принадлежащих к 180-200 родам) сосредоточено в тропических странах обоих полушарий, где они представлены разнообразными жизненными формами. Из последних среди кутровых значительно преобладают деревянистые лианы, обычно вьющиеся, реже лазящие с помощью видоизмененных в усики соцветий (например, у многих видов ландолъфии - Landolphia). Прямостоячих деревьев и кустарников не так уж много, хотя в родах рауволъфия (Rauvolfia), аспидосперма (Aspidosperma) и алъстония (Alstonia) имеются довольно высокие деревья. Из кустарниковых кутровых можно отметить южноавстралийский эрикоидный (верескоподобный) кустарник нотонериум (Notonerium) и колючие растопыренно разветвленные кустарники из рода кариеса (Carissa).
Среди кутровых имеются также крайне своеобразные полудревесные жизненные формы. Таковы виды бразильского кампоса (саванно-подобной растительности) из родов макросифония (Macrosiphonia), родокаликс (Rhodocalyx) и дипладения (Dipladenia) с клубневидно утолщенным основанием ствола, служащим хранилищем воды и крахмала. Не менее замечательны африканские суккулентные виды из родов адениум (Adenium) и пахиподиум (Pachypodium). Так, адениум тучный (A. obesum) из Кении имеет клубнеподобный ствол, от верхней части которого отходят короткие, но довольно толстые ветви, несущие немного мясистые листья и красные цветки в немногоцветковых конечных соцветиях. У пахиподиума Лила (P. lealii) из Анголы толстые, почти конические стволы, как и ветви, густо покрыты колючками, листья ко времени цветения обычно опадают, что делает облик цветущего растения еще более оригинальным.
Многолетние травянистые виды обычного облика имеются лишь в 6 родах кутровых, в том числе встречающихся и в СССР родах барвинок и кендырь (Trachomitum). Растущий на степных склонах юга европейской части СССР барвинок травянистый (Vinca herbacea) имеет ежегодно отмирающие до основания, стелющиеся по земле стебли, в то время как у более известного барвинка малого (V. minor) они в нижней части одревесневают.
Листья кутровых цельные и цельнокрайние, почти всегда без прилистников, расположенные большей частью супротивно, реже мутовчато (например, у раувольфии) или очередно (у адениума и пахиподиума). В пазухах листьев табернемонтапы (Tabernaemontana) имеются особые желёзки, выделения которых как бы лакируют близлежащие части растения. Для кутровых очень характерно наличие в листьях и других органах канальцев, содержащих млечный сок, латекс, нередко с довольно высоким процентом каучука.
Цветки кутровых (рис. 188, 189, табл. 49) всегда обоеполые, обычно актиноморфные (сильно зигоморфные у кондилокарпона - Condilocarpon) и 5-членные (лишь 3 небольших рода имеют 4-членные цветки). Обычно они собраны в метелкообразные, кистевидные или щитковидные соцветия, реже расположены по одному на верхушках ветвей или в пазухах листьев (например, у барвинка). Чашечка обычно почти до основания рассечена и нередко имеет у основания долей с их внутренней стороны железистые чешуйки. Венчик может быть трубчатым, воронковидным, колокольчатым или блюдцевидным и имеет почти всегда скрученные в почкосложении доли. В его обычно волосистой с внутренней стороны трубке часто имеются чешуевидные придатки, иногда лепестковидные и далеко выступающие из зева венчика (например, у олеандра). Тычинки прикреплены к трубке венчика, чередуясь с его долями, и имеют очень короткие нити. Лишь в цветках азиатской лианы бюмонтии (Beaumontia) пыльники на длинных нитях далеко выступают из трубки венчика. У мексиканской лианы тенардии пышноцветущей (Thenardia floribunda) встречается другая особенность в строении тычинок: их нити срастаются в короткую трубку. Пыльники могут быть двух типов, которым придается большое значение при делении кутровых на подсемейства. У более примитивного подсемейства плюмиеровых (Plumieroideae) пыльники вполне свободные или лишь соприкасаются с рыльцевой головкой гинецея; все 4 гнезда у них одинаково развитые и содержат пыльцу. У видов более высоко специализированного подсемейства кутровых (Apocynoideae) тесно сближенные друг с другом пыльники своей внутренней стороной очень тесно примыкают к рыльцевой головке или даже слипаются с ее расширенной частью. Кроме того, их наружные гнезда лишены пыльцы и оттянуты в довольно длинные хвостовидные придатки.

Таблица 49. Кутровые: 1 - плюмерия белая (Plumeria alba), оранжереи Ботанического института АН СССР в Ленинграде; 2 - трахелоспермум жасминовидный (Trachelospermum jasminoides), Сухумский ботанический сад; 3 - строфант жёстковолосистый (Strophanthus hispidus), Сухумский ботанический сад
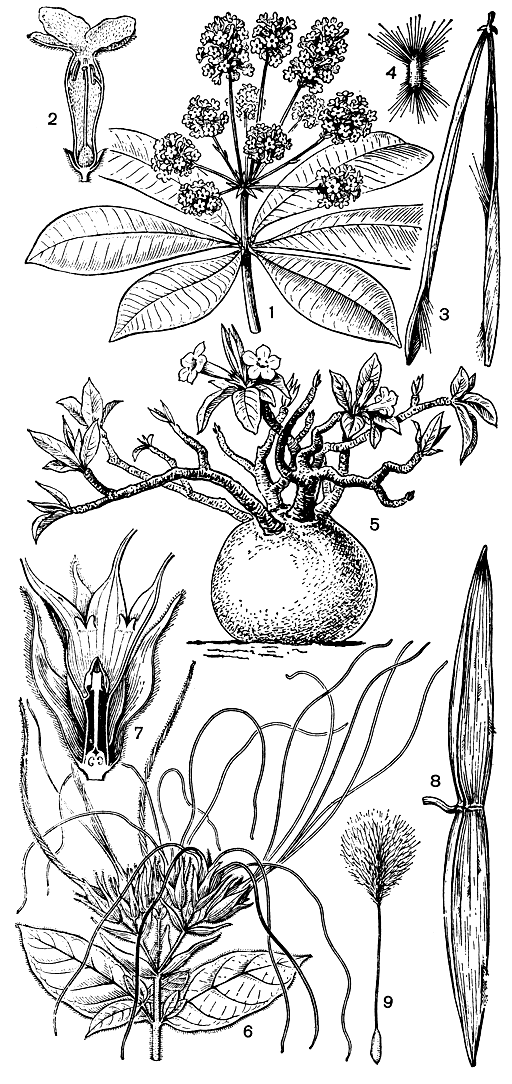
Рис. 189. Кутровые. Альстония малайская (Alstonia scholaris): 1 - верхушка цветущей ветви; 2 - цветок на продольном разрезе; 3 - раскрывшийся плод; 4 - семя. Адениум тучный (Adenium obesum): 5 - общий вид растения. Строфант жестковолосистый (Strophanthus hispidus): 6 - верхушка цветущей ветви; 7 - продольный разрез цветка; 8 - плод; 9 - семя
Апокарпный, реже синкарпный гинецей цветка кутровых образован 2, редко (триба плейокарповых - Pleiocarpeae) 3-5 плодолистиками. Обычно плодолистики по всей или почти по всей длине свободны, но их стилодии всегда срастаются между собой в столбик, заканчивающийся сильно утолщенной рыльцевой головкой - одной из характерных особенностей цветка кутровых. От обычного рыльца она отличается тем, что покрытая клейкими выделениями, воспринимающая пыльцу поверхность головки (т. е. собственно рыльце) находится не в верхней ее части, а по бокам ее или еще ниже, под кольцеобразным расширением рыльцевой головки и обычно против отверстий в конусообразном футляре, образованном над рыльцевой головкой тесно сближенными пыльниками.
Все кутровые - насекомоопыляемые растения. Как уже отмечалось выше, в цветках этого семейства рыльцевая головка обычно прикрыта сверху как бы крышей из пыльников. Насекомые с хоботком, достаточно длинным для того, чтобы достать нектар у основания завязи (обычно перепончатокрылые, мухи или бабочки), просовывают хоботок в одно из 5 отверстий между тычиночными нитями ниже сомкнутых пыльников и прежде всего попадают им на участок рыльцевой головки, покрытый клейкими выделениями. Когда же насекомое вынимает из отверстия обмазанный этими выделениями хоботок, к нему прилипает порция пыльцы из близлежащих пыльцевых гнезд. При посещении насекомым следующего цветка часть этой пыльцы прилипает к воспринимающей пыльцу поверхности его рыльцевой головки. Таким образом осуществляется перекрестное опыление.
Плоды большинства кутровых (особенно из подсемейства кутровых) состоят из 2 вскрывающихся по брюшным швам листовок. Довольно многочисленные семена в них обычно снабжены летучкой из волосков или крыловидной каймой, увеличивающих их парусность, и распространяются с помощью ветра. Однако многие роды подсемейства плюмиеровых имеют мясистые, обычно нераскрывающиеся плоды и распространяются эндозоохорно. Таковы, например, крупные, апельсиноподобные плоды табернемонтаны померанцевой (Tabernaemontana aurantiaca), образующиеся только из одного плодолистика (второй плодолистик у этого вида редуцируется). Экзозоохорно (на шерсти животных) распространяются также невскрывающиеся односеменные части плода кубинской лианы анехитеса липучкового (Anechites lappulacea), покрытые в верхней части мелкими крючковидно изогнутыми шипиками. Не вскрываются также крылаткоподобные половинки плодов камерарии (Cameraria) - прямостоячего кустарника с Антильских островов. У южноамериканских лиан из рода кондилокарпон (Condylocarpon) части плода имеют глубокие перетяжки и распадаются на односемянные членики, снабженные пробковой тканью и распространяющиеся с помощью водных потоков.
Синкарпные плоды, образованные 2 сросшимися по всей длине плодолистиками, имеются лишь у некоторых родов подсемейства плюмиеровых. Они могут быть или невскрывающимися мясистыми, или вскрывающимися коробочко-образными. Мясистые плоды церберы мангас (Cerbera manghas), окутанные сетчато-волокнистыми оболочками, могут долгое время плавать в морской воде, не теряя всхожести. Этим объясняется широкое распространение этого вида по морским побережьям от Мадагаскара до Новой Гвинеи. Из синкарпных плодов-коробочек можно отметить плоды американского рода аллеманда (Allemanda), густо покрытые шипами и напоминающие по облику каштаны.
Многие виды кутровых имеют существенное хозяйственное значение в тропиках. Некоторые из них введены в культуру ради съедобных плодов, отличающихся приятным вкусом. Сюда принадлежат, например, плоды нескольких видов ландольфии, напоминающие по внешнему виду, а часто и по вкусу плоды цитрусовых. В Южной Америке славятся по своим вкусовым качествам плоды видов рода коума (Couma) и ханкорнии прекрасной (Hancornia speciosa). Можно отметить еще культивируемые в Юго-Восточной Азии ради плодов лиану виллугбейю съедобную (Willughbeia edulis) и колючий кустарник кариcсу кавандас (Carissa cavandas), служащий одновременно живой изгородью.
Представители многих родов кутровых прежде использовались для получения каучука. Особенно ценными каучуконосами являются многие виды африканских родов ландольфия и клитандра (Clitandra), ханкорния прекрасная, виды виллугбейи. Очень ценную из-за твердости и красивой белой окраски древесину дает "белое квебрахо" (Aspidosperma quebracho-bianco), кора которого содержит не менее ценные дубильные вещества. Напротив, очень легкую и мягкую древесину дают виды альстонии. Кроме того, кутровые богаты алкалоидами и другими веществами, используемыми для изготовления лекарственных препаратов. Многие виды раувольфии, в особенности индо-малайская рауволъфия змеиная (Rauvolfia serpentina), содержат более 20 различных алкалоидов, которые входят в состав таких лекарств, как резерпин и раунатин. Из семян и корней африканского строфанта приятного (Strophanti us gratus) и некоторых других видов этого рода получают ядовитый гликозид строфантин, входящий в состав лекарств, поддерживающих сердечную деятельность. Туземцы многих стран использовали сильно ядовитые вещества различных представителей кутровых (адениума, строфанта и др.) для ядовитых стрел.
Среди кутровых имеется также немало декоративных растений с красивыми, нередко душистыми цветками, однако значительное большинство их не выходит за пределы тропиков. В Европе, в том числе и на юге европейской части СССР, довольно широко культивируется в качестве газонного и бордюрного растения стелющийся вечнозеленый полукустарничек барвинок малый с довольно крупными голубыми, реже розовыми или белыми цветками. Но особенной известностью среди декоративных кутровых пользуется высокий вечнозеленый кустарник олеандр - одно из старейших культивируемых растений Средиземноморья. В более северных районах он является одним из наиболее распространенных комнатных растений. Содержащийся в листьях олеандра гликозид используют при заболеваниях сердечных мышц. Используемые для изготовления сердечных лекарств гликозиды содержит также культивируемый во многих странах, в том числе и в СССР, американский травянистый вид кутра коноплевая (Apocynum cannabinum), прочные и гибкие стеблевые волокна которой идут на изготовление веревок, рыболовных сетей и грубых тканей. В качестве ценных волокнистых растений используют также виды очень близкого к кутре и нередко объединяемого с этим родом евразиатского рода кендырь.
Семейство ластовневые (Asclepiadaceae) (Н. Н. Цвелев)
Очень близкое к кутровым семейство ластовневых, или ласточниковых, сходно с кутровыми и по своему географическому распространению. Лишь немногие из примерно 250 родов и 2000 видов этого семейства распространены за пределами тропиков, исключая очень богатую ластовневыми Южную Африку. В Арктике и в значительной части северной лесной зоны, а также в Новой Зеландии ластовневые полностью отсутствуют.
Подобно кутровым в семействе ластовневых преобладают вьющиеся и лазящие лианы с более или менее одревесневающими стеблями. Высоких прямостоячих кустарников среди них немного, еще реже встречаются высокие деревья (индийская утлерия иволистная - Utleria salicifolia). Довольно широко представлены жизненные формы, приспособленные к обитанию в условиях засушливого климата. Сюда принадлежат, например, такие безлистные или почти безлистные прутьевидные кустарники с членистыми ветвями, как обвойник хвойникоподобный (Periploca ephedroides) и лептадения пиротехническая (Leptadenia pyrotechnica, рис. 190, 6 и 7). Высушенные стебли последнего вида, широко распространенного в пустынях Африки и Юго-Западной Азии, очень легко воспламеняются и горят ярким пламенем, вполне оправдывая свое видовое название. Не менее оригинальны деканема Боже (Decanema bojerianum) с Мадагаскара - высокий лазящий кустарник с редуцированными до мелких чешуи листьями и виды саркостеммы (Sarcostemma) - безлистные лежачие или вьющиеся кустарники с членистыми стеблями, распространенные в тропиках Старого Света (рис. 190, 2 и 3). В бразильском кампосе (саванноподобной группировке растительности) встречается эрикоидное полудерево хемипогон щетинолистный (Hemipogon setaceus), травянистые ветви которого отходят от толстого одревесневшего стволика и густо покрыты очень узкими, хвоеподобными листьями (рис. 190, 1).
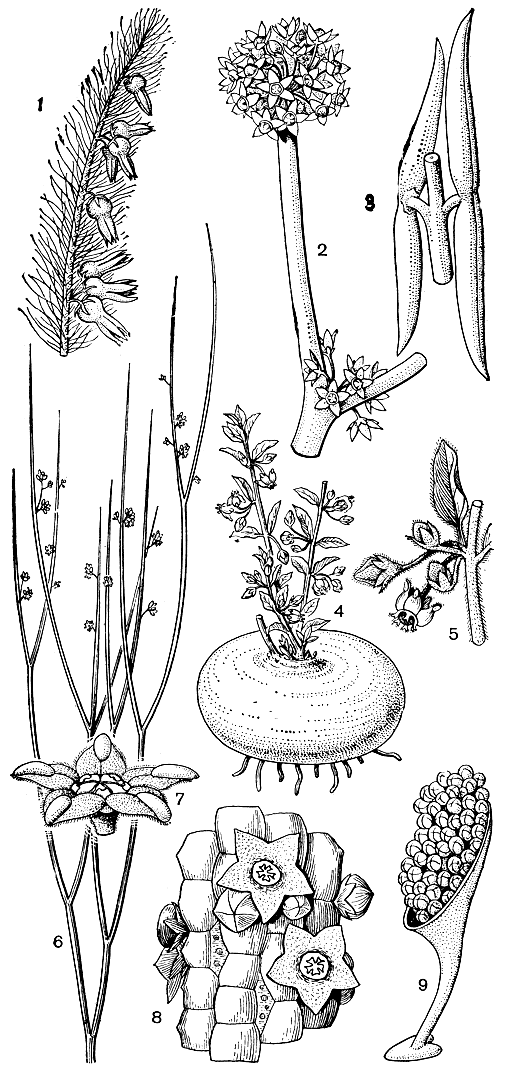
Рис. 190. Ластовневые. Хемипогон щетинолистный (Hemlpogon setaceus): 1 - часть цветущей ветви. Саркостемма кислая (Sarcostemma acidum); 2 - часть цветущей ветви; 3 - плоды. Анизотома Арнотта (Anisotoma arnottii); 4 - общий вид растения; 5 - часть ветви с соцветием. Лептадения пиротехническая (Leptadenia pyrotechnica); 6 - цветущая ветвь; 7 - цветок. Ехиднопсис Вирховa (Echidnopsis virchowii); 8 - часть стебля с цветками. Филлантера двураздельная (Phyllantliera bifida); 9 - транслятор с тетрадами пыльцы
Из довольно многочисленных травянистых многолетников, к которым принадлежит большинство ластовневых флоры СССР, многие африканские и другие виды имеют крупные клубни или репообразно утолщенное основание стебля, например южноафриканская анизотома Арнотта (Anisotoma arnottii, рис. 190, 4 и 5). Очень характерно для ластовневых и широкое распространение в пределах этого семейства суккулентов, как листовых, так и стеблевых, нередко совсем лишенных листьев. К листовым суккулентам могут быть отнесены многие из культивируемых в оранжереях и комнатах видов родов хойя (Hoya) и церопегия (Ceropegia, табл. 50) с относительно небольшими мясистыми листьями. В пределах последнего из этих родов известны и стеблевые суккуленты, однако особенно замечательны в этом отношении африканские роды стапелия (Stapelia, табл. 50), худия (Hoodia), трихокаулон (Trichocaulon), ехиднопсис (Echidnopsis, рис. 190, 8) и другие, имеющие кактусообразную жизненную форму. Листья у этих растений видоизменены в чешуйки или колючки, располагающиеся на более или менее вздутых бугорках, называемых листовыми подушками. Эти бугорки обычно образуют правильные продольные ряды, но у некоторых родов с клубнеобразными стеблями (например, у трихокаулона) эта правильность может нарушаться, бугорки становятся уплощенными в виде щитков, а вся поверхность стеблей выглядит как бы панцирной. В отличие от кактусов одиночные или немногочисленные, иногда довольно крупные (у стапелий диаметром до 30 см) цветки ластовневых - стеблевых суккулентов - расположены не в пазухах видоизмененных листьев, а в бороздах между рядами бугорков.

Таблица 50. Ластовневые: 2 - церопегия стапелиевидная (Ceropegia stapeliiformis), оранжереи Ботанического института АН СССР в Ленинграде

Таблица 50. Ластовневые: 3 - стапелия пёстрая (Stapelia variegata), оранжереи Ботанического института АН СССР в Ленинграде
К наиболее оригинальным в биологическом отношении ластовневым принадлежат также некоторые азиатско-австралийские виды дисхидии, особенно дисхидия Раффлеза (Dischidia rafflesiana, рис. 191). Эта эпифитная, часто растущая высоко на стволах деревьев травянистая лиана имеет листья двух родов: одни - немного мясистые листья обычного облика; другие - видоизмененные в своеобразные мешковидные органы, служащие хранилищами для воды, а иногда также жилищами для муравьев и образованные завернутыми на нижнюю сторону и сросшимися между собой краями листовой пластинки. У обычно обращенного кверху основания такого мешковидного листа имеется широкое, окаймленное валиком отверстие, в которое входят сильно разветвленные воздушные корни, отходящие от стебля близ основания листа и всасывающие попадающую в него во время дождей воду.
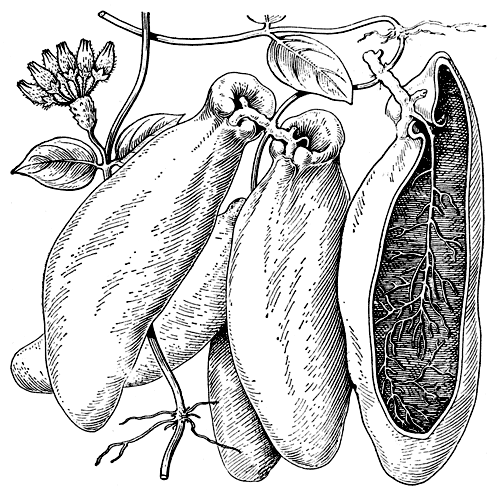
Рис. 191. Мешковидные листья дисхидии Раффлеза (Dischidia rafflesiana): воздушные корни, заходящие в полость листа
Невидоизмененные листья ластовневых довольно однотипны. Они почти всегда супротивные, редко мутовчатые или очередные (у хемипогона щетинолистного), без прилистников или с рудиментами прилистников, с цельными и цельнокрайними, реже близ основания лопастными или неправильно зубчатыми пластинками. Как и для кутровых, для ластовневых очень характерно наличие в листьях и стеблях канальцев, содержащих млечный сок латекс.
Обоеполые и 5-членные, почти всегда актиноморфные цветки ластовневых обычно мелкие и собраны в различного рода соцветия, имеющие вид зонтика или пучкообразной кисти. Как правило, такие соцветия располагаются в пазухе только одного из 2 супротивных листьев, что связано, по-видимому, с широким распространением в пределах семейства жизненной формы - вьющейся лианы. Однако имеются и роды с одиночными или немногими пучковидно расположенными цветками, обычно имеющими более крупные размеры. Чашечка в цветке ластовневых построена очень однообразно, обычно почти до основания рассеченная, и имеет относительно небольшие размеры. Значительно более изменчива форма венчика, который часто бывает колесовидным, но нередко имеет и более длинную трубку, становясь тогда воронковидным или колокольчатым. Его лопасти или доли бывают в почкосложении скрученными или створчатыми. В последнем случае они иногда срастаются своими верхушками, в результате чего образуется крайне своеобразная форма венчика, особенно характерная для многих видов церопегии, венчики которых похожи на ажурные фонарики с 5 боковыми отверстиями в их верхней части (табл. 50).
Очень обычны в семействе ластовневых усложнения в строении цветков, связанные с образованием внутри венчика одной или нескольких коронок из чешуевидных или лепестковидных, нередко мясистых придатков самого венчика в его зеве, сильно укороченных тычиночных нитей или стерильных частей пыльников. Нередко составляющие коронку придатки срастаются между собой, как бы образуя второй, внутренний венчик (например, у южноафриканской диплоциаты реснитчатой - Diplocyatha ciliata). В ряде других случаев эти придатки становятся вместилищами нектара.
Уже формирование коронок в цветках ластовневых связано с их очень высокой специализацией к энтомофилии - опылению с помощью насекомых. Еще в большей степени эта специализация сказывается на других особенностях в строении цветков ластовневых. Это прежде всего образование так называемого гиностегия путем объединения пыльников всех тычинок в кольцо, прирастающее к утолщенной верхушке очень короткого столбика, называемой рыльцевой головкой, или только слипается с ней. Воспринимающая пыльцу поверхность рыльцевой головки, т. е. собственно рыльце, находится не на ее верхушке, как это обычно бывает в других семействах цветковых растений, а на ее нижней стороне в виде 5 ясно отграниченных от остальной поверхности головки участках. Таким образом, в цветке ластовневых рыльце оказывается прикрытым сверху пятигранной конусовидной колонкой (или столбиком) из пыльников.
Очень существенные различия в строении пыльников имеются у 2 подсемейств ластовневых, нередко принимаемых за самостоятельные семейства. У более примитивного подсемейства обвойниковых (Periplocoideae) пыльцевые зерна в 4 гнездах пыльников объединены в довольно многочисленные тетрады - группы из 4 зерен, а у подсемейства ластовневых (Asclepiadoideae) все микроспоры каждого из 2 фертильных гнезд пыльников объединены в мешочковидные комочки, называемые поллиниями. Кроме того, оба подсемейства имеют разное строение свойственных только ластовневым особых переносчиков пыльцы, так называемых трансляторов, расположенных в щелях между пыльниками и образованных из затвердевших выделений специальных желёзок на рыльцевой головке. У обвойниковых трансляторы имеют ложкообразную или воронковидную форму и оканчиваются ножкой с липким диском (рис. 190, 9). У подсемейства ластовневых (рис. 192) они состоят из зажимающего срединного тельца, или "корпускулы", от которой отходит пара ножек, прикрепляющихся к поллиниям правого и левого гнезд 2 соседних пыльников. Детали строения таких трансляторов могут значительно варьировать. Так, у статмостелъмы (Stathmostelma) ножки трансляторов второго типа сильно расширены, вогнуты и значительно крупнее поллиниев. У секамопе (Secamone) все 4 гнезда пыльников развиты и трансляторы несут не 2, а 4 пол линия. В цветках большинства ластовневых нектар доступен короткохоботковым насекомым - обычно перепончатокрылым или мухам. Цветки стапелий и некоторых других суккулентных родов, имеющие запах падали, а нередко и по окраске напоминающие гнилое мясо, опыляются преимущественно крупными мясными мухами. Отмечены даже случаи, когда мухи по ошибке откладывали яички в такие цветки.
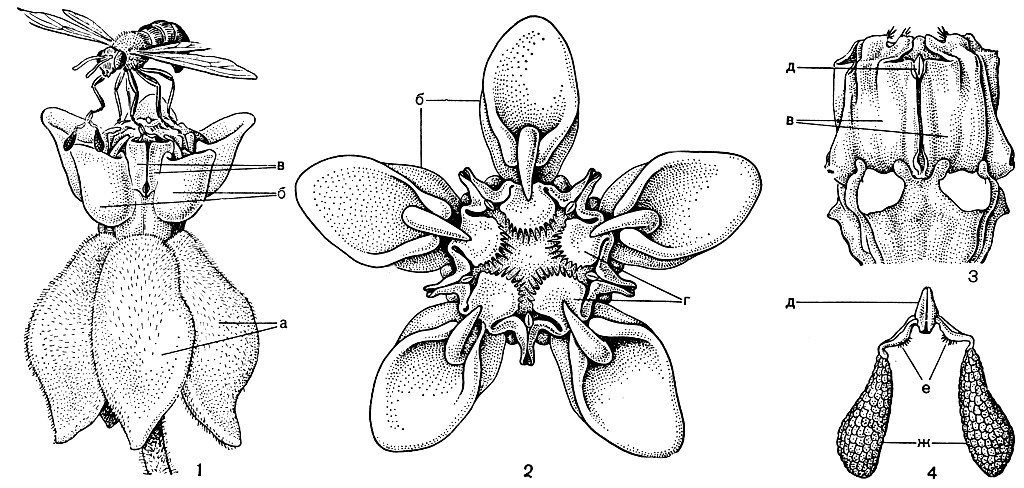
Рис. 192. Ластовень сирийский (Asclepias syriaca): 1 - цветок (вид сбоку); 2 - цветок (вид сверху); 3 - гиностегий с удаленной коронкой сбоку; 4 - транслятор с поллиниями; а - доли венчика; б - листочки коронки; в - пыльники; г - верхушки пыльников; д - корпускула; е - ножки транслятора; ж - поллинии
У обвойниковых (например, у обычного в Средиземноморье обвойника греческого - Periploca graeca) способ опыления напоминает способ опыления цветков у многих орхидей. Насекомое, отыскивая нектар, касается головкой липкого диска транслятора и улетает с цветка, унося на головке приклеившийся к ней транслятор с тетрадами пыльцы в его расширенной части. При посещении другого цветка расширенная часть транслятора оказывается против воспринимающего пыльцу участка рыльцевой головки и снабжает его пыльцой.
Более сложен и совершенно уникален способ "опыления в подсемействе ластовневых (см. рис. 192). Посещая богатый нектаром цветок ластобня сирийского (Asclepias syriaca), насекомое стремится закрепиться на гладкой поверхности венчика и гиностегия. Оно наступает в вырезы между нектароносными листочками коронки, но отсюда его снабженные коготками ноги скользят через щели между пыльниками к скрытым в глубине этих щелей корпускулам и зажимаются ими. Стенки стерильных гнезд пыльников служат при этом как бы направляющими плоскостями. Покидая цветок, насекомое уносит на своих коготках трансляторы с поллиниями. Доступ к воспринимающим пыльцу участкам рыльцевой головки, перед которыми находятся так называемые рыльцевые камеры, также открывается через щели между пыльниками. Посещая другой цветок, насекомое вновь попадает ногами в эти щели и заталкивает в рыльцевые камеры поллинии. При вытаскивании ног из щелей ножки транслятора обрываются и поллинии остаются в рыльцевой камере. При этом к ногам насекомого могут прицепиться новые трансляторы с поллиниями.
Подобным же образом происходит опыление цветков и у большинства других представителей подсемейства ластовневых. Однако у обладающих крайне оригинальным строением венчика церопегий опыление осуществляется несколько иным способом, напоминающим хорошо изученный способ опыления у кирказона (Aristolochia) из семейства кирказоновых. Мелкие насекомые через боковые отверстия в венчике церопегий попадают в нижнюю, расширенную часть его трубки, где находится относительно небольшой гиностегий, но не могут выйти обратно из-за обращенных вниз волосков в суженной части трубки. Только через день после полного распускания цветков волоски отмирают ж насекомые могут выбраться из цветка, унося на себе трансляторы с поллиниями.
Довольно однообразные по строению плоды ластовневых, состоящие из 2 раскрывающихся по брюшному шву листовок, формируются из апокарпного гинецея, 2 плодолистика которого соединены на верхушке рыльцевой головкой. Многочисленные семена с хохолком из более или менее длинных шелковистых волосков распространяются с помощью ветра. У некоторых африканских видов рода дрежея (Dregea) ветром распространяются не только семена, но и рано опадающие листовки, снабженные продольными крыловидными выростами.
Хозяйственное значение представителей ластовневых не так уж велико. Происходящее из Северной Америки многолетнее травянистое растение ластовень сирийский культивируется и дичает во многих внетропических странах, в том числе и в СССР. Волокна его стеблей пригодны для изготовления грубых тканей и веревок, волоски семян могут использоваться как заменители ваты, в семенах содержится более 20% технического масла. Кроме того, это ценный засухоустойчивый медонос. Кору американской кустарниковой лианы кондуранго (Marsdenia condurango), содержащую ценные гликозиды, применяют как лекарственное средство. Молодые побеги, листья и клубни некоторых африканских видов используют в пищу. В странах Средиземноморья, а в СССР в Крыму и на Кавказе в качестве декоративного растения культивируют кустарниковую лиану - обвойник греческий, кора которого содержит гликозиды и алкалоиды. Из многочисленных оранжерейных и комнатных ластовневых следует в особенности отметить "восковый плющ", или хойю мясистую (Hoya carnosa),- вьющийся кустарник с блестящими, немного мясистыми листьями и зонтиками белых или розовых душистых цветков, как бы вылепленных из воска; несколько видов церопегий - лиан с довольно крупными цветками оригинального строения (особенно у церопегий Сандерсона - Ceropegia sandersonii); замечательные африканские стеблевые суккуленты из родов стапелия, трихокаулон, ехиднопсис и других, часто культивируемые в специальных оранжереях вместе с кактусами и другими суккулентными ксерофитами.
Таблица 50. Ластовневые: 1 - ластовень курассавский (Asclepias curassavica), оранжереи Ботанического института АН СССР в Ленинграде
Семейство горечавковые (Gentianaceae) (В. В. Письяукова)
Горечавковые - большое семейство, насчитывающее в настоящее время около 80 родов и более 1000 видов. Распространены они по всей Земле. В умеренных широтах и в горах в семействе господствуют однолетние и многолетние травы, в субтропических и тропических областях представлены и полукустарники, кустарники, лианы, деревца высотой до 5 м и небольшие травянистые сапрофиты. Разнообразие горечавковых и широкий экологический диапазон их видов определяют роль семейства в растительном покрове Земли. Горечавковые можно встретить от тропиков до снегов Арктики; они растут в тундре, в степях, в лесах разных типов и широт, на лугах, болотах, по берегам водоемов, но особенно их много в горах, в альпийских поясах которых они часто господствуют (виды горечавки - Gentiana, табл. 51, сверции - Swertia и др.). Многим горечавковым свойственны разной формы корневища, иногда мясистые, реповидные, всегда несущие запасы питательных веществ. Стебли их часто простые или ложнодихотомически ветвящиеся, что свойственно многим тропическим видам, а также, например, золототысячнику (Gentaurium) и другим в умеренных широтах. Листья простые, цельнокрайные, иногда большие, длиной 10- 20 см, 5-10 см шириной (например, у некоторых сверций), или совсем мелкие, чешуевидные, часто даже неокрашенные, что свойственно сапрофитным формам; они могут быть сидячими или на черешках, часто широко крылатых, всегда с хорошо развитым или сросшимся влагалищем. Расположение листьев обычно супротивное, но у многих видов сверций очередное, реже листорасположение мутовчатое, а у некоторых горечавок листья в прикорневой розетке. Среди тропических форм есть и вечнозеленые, например распространенный в Андах крупноплодник (Macrocarpaea) - деревце высотой 3-5 м, с кожистыми блестящими листьями и др. Характерной анатомической чертой семейства является присутствие в проводящей системе стебля внутренней (интраксилярной) флоэмы.

Рис. 193. Горечавковые. Лагенантус превосходный (Lagenanthus princeps): 1 - цветущий побег. Лизиантус Рассела (Lisianthus russellianus); 2 - цветущий побег; 3 - гинецей. Тахиаденус ладьевидный (Tachiadenus carinatus); 4 - цветуший побег; 5 - цветок в разрезе
Соцветия у горечавковых обычно верхоцветные, но иногда их определяют и как бокоцветные, наконец, полагают, что оба типа здесь сочетаются, образуя тирсоидные, пирамидально-метельчатые соцветия, у которых главная ось - моноподий, а боковые ветвятся симподиально. Цветки обычно обоеполые, большей частью 5-4-членные, редко (у некоторых бразильских представителей) они 6-членные, у крымско-кавказской блэкстонии пронзеннолистной (Blackstonia perfoliata) - 8-членные, наконец, у американского рода сабатия (Sabbatia) - 12-членные. Обычно цветки актиноморфные; лишь у видов палеотропического рода консора (Consora) наблюдается слабая зигоморфия. Чашелистики сросшиеся (иногда лишь у самого основания), венчик сростнолепестный, в почкосложении скрученный. Окраска лепестков поражает разнообразием - белая, желтая, розовая, красная, оранжевая, голубая, синяя, а у мексиканского "цветка смерти" (Flor del Muerte), как там называют лизиантус чернеющий (Lisianthus nigrescens), цветки черные, лишь при некотором освещении они кажутся иногда слабо красноватыми. Часты комбинации цветов окраски венчика. Размеры цветков у горечавковых также разнообразны, чаще длиной от 0,5 до 6-7 см, однако у кустарников из рода симболантус (Symbolanthus), растущих в Андах (Колумбия, Эквадор) в мшистых туманных низкорослых лесах на высоте от 3300 до 4500 м (в так называемых "лесах эльфов"), цветки достигают в длину 12,7 см, а у американского лагенантуса превосходного (Lagenanthus princeps) они длиной до 18 см. Тычинки в числе лепестков и прирастают нитями к трубке венчика, чередуясь с зубцами его отгиба, нити тычинок обычно тонкие, но у видов сверций, например, они более или менее лентовидные и достигают в ширину 2 мм, резко суживаясь лишь под пыльниками; связники часто значительно разрастаются, образуя разной формы выросты над пыльниками (рис. 196). Пыльники прикрепляются к нитям основанием или серединой (качающиеся), стреловидные; тычинки между собой свободные, но у некоторых сапрофитных родов они срастаются пыльниками. Пыльники обычно открываются щелью на своей наружной, т. е. повернутой от центра цветка к трубке венчика, стороне (экстрорзные). Пыльца бывает свободной или в тетрадах, последнее свойственно членам трибы Helieae, состоящей из 60 американских видов. Гинецей состоит из 2 плодолистиков, паракарпный; завязь с коротким или чаще уже довольно длинным столбиком, несущим двулопастное или головчатое рыльце; рыльца комиссуральные, т. е. расположены по спайкам плодолистиков и при открывании плода половинки рыльца отходят с каждым из них. Однако у ломатогониума (Lomatogonium) и у некоторых сверций Гималаев и Тибета (сверция тибетская - Swertia tibetia, сверция Кинга - S. kingii и др.) столбика вообще еще нет, а воспринимающие поверхности просто низбегают по швам завязи (рис. 194). Завязь верхняя, одногнездная, но париетальные плаценты иногда, разрастаясь к ее центру, могут образовать ложную перегородку. Семязачатков много, они обычно анатропные и только у галении (Halenia) ортотропные.
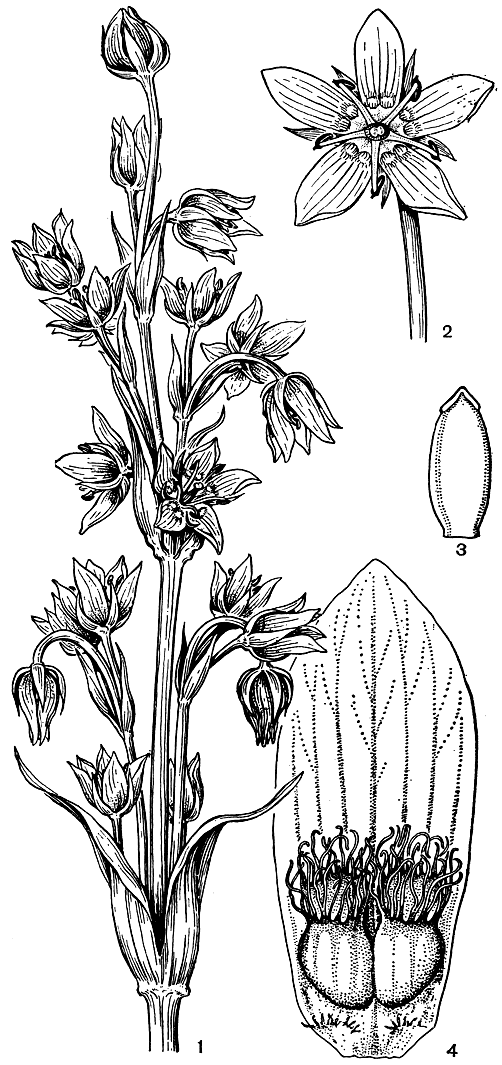
Рис. 194. Сверция Кинга (Swertia kingii): 1 - цветущий побег; 2 - цветок в развернутом виде; 3 - гинецей с низбегающим рыльцем; 4 - лепесток с нектарниками
Цветки у горечавковых с разнообразными по происхождению, расположению и форме нектарниками. У большинства видов нектарники развиваются на цветоложе (торальные) у основания завязи (иногда внизу и на ней самой), образуя разной формы валики или род 4-5-лопастного воротничка. У некоторых видов желёзки развиваются у основания чашелистиков и на их внутренней поверхности (рис. 195). Нектар может выделяться и тычинками. Например, у тропического растения Старого Света себеи (Sebaea) железистая ткань развивается на верхушке связников тычинок, а у африканского лагениаса (Lagenias) эта ткань обособляется на нижних концах пыльников (рис. 196). Интересно, что трубки венчиков таких цветков бывают окрашены светлее; их отгибы (они часто даже просвечивают как "окна") указывают путь к нектару. Это свойственно и альпийским горечавкам, у которых горловина цветка светлая, а лопасти синие, и вход в цветок резко обозначен. Совсем иные нектарники у горечавочки (Gentianella), сверции, офелии (Ophelia), фразеры (Frasera), ломатогониума, галении. Нектарники их эпипетальные, т. е. возникают на лепестках; они располагаются у их основания (иногда выше), обычно парные по обе стороны центральной жилки или одиночные посередине. Это небольшие углубления (ямки), кармашки, огражденные более плотной окраиной и часто прикрытые бахромками или волосками, развивающимися на ней. Одиночные ямки возникают в результате срастания двух, а у галении каждая из них, постепенно разрастаясь, образует на лепестке шпору. Полагают, что эти нектарники возникли на месте пазушных прилистников, приросших к лепесткам. Перекрестное опыление у горечавковых обеспечивается существованием дихогамии, т. е. разновременным созреванием тычинок и рылец. В результате возникают цветки протандрические и протогиничные. В первом случае цветок вначале функционирует как мужской, а затем созревает и его женская часть, во втором - наоборот. Протандрия распространена в семействе вообще очень широко, в то время как протогиния встречается реже, например у горечавки весенней (Gentiana verna), комастомы нежной (Comastoma tenellum) и др. В качестве препятствия самоопылению наблюдается и гетеростилия (разностолбчатость), при этом на растениях одни цветки с очень длинным столбиком и короткими тычинками, другие с их отношениями обратными. Кроме того, в цветках горечавковых тычинки часто перемещаются, т. е. свободные концы их нитей то приближают пыльники к рыльцу, то отклоняются от него к трубке венчика, высыпая пыльцу на ее стенки. Последнее затрудняет самоопыление в том случае, если дихогамия не очень строгая. В связи с приспособлением к перекрестному опылению следует отметить и большую продолжительность жизни цветков у горечавковых; она длится 3-5-7 дней, в течение которых цветки закрываются на ночь (или при плохой погоде) и открываются на восходе солнца. В качестве опылителей цветков горечавковых известны многочисленные представители бабочек, пчел, шмелей, мух, жуков, ос. Для них пищей служит нектар, а теплая и защищающая от ветра трубка венчика - хорошее укрытие в холодные ночи в горах. На альпийском лугу перед восходом солнца температура в трубке цветка, например, у горечавки бесстебельной оказалась на 2,2° выше, чем снаружи. Если открытые цветки (с колесовидным венчиком) посещает большая часть названных насекомых, то опыляют их все же бабочки, пчелы, шмели, обладающие длинными хоботками. Мелкие бабочки посещают эти цветки, забирая пыльцу. Обычными посетителями бокаловидных цветков горечавок являются шмели, и многие из них даже называют шмелиными цветками; осы нектар у горечавок часто воруют, прогрызая цветки. Подчеркнуто энтомофильные цветки у альпийских горечавковых свидетельствуют о том, что даже в очень суровых условиях высокогорий насекомые осуществляют перекрестное опыление.

Рис. 195. Горечавковые. Иксантус клейкий (Ixanthus viscosus): 1 - цветущий побег; 2 - листочки чашечки с нектароносными желёзками на внутренней стороне. Лизиантус мыльнянковидный (Lisianthus saponarioides); 3 - цветущий побег; 4 - листочки чашечки с желёзками. Крупно плодник голый (Macrocarpaea glabra); 5 - листочки чашечки с желёзками
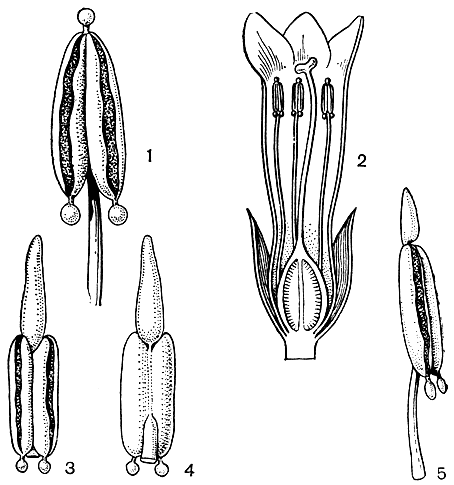
Рис. 196. Горечавковые. Лагениас маленький (Lagenias pusillus): 1 - тычинка с шаровидными нектароносными придатками на верхушке связника и на нижнем конце пыльников; 2 - цветок в разрезе. Бельмонтия первоцветная (Belmontia primuliflora); 3 - пыльник с продолговатым железистым связником на верхушке и округлыми у основания; 4 - вид пыльник' с брюшной стороны. Бельмонтия большая (B. grandis); 5 - тычинка
Тропические представители семейства, обладающие большими цветками, опыляются летучими мышами, в частности, из рода глоссофага (Glossophaga); в Южной Америке они опыляют цветки хелонантуса (Ghelonanthus), прекрасные цветки кустарников из рода симболантус (Symbolanthus), древовидных крупноплодников (Масгосаграеа), лагенантуса превосходного. Эти цветки часто несут на себе следы посещения летучими мышами, которые, устремляясь к нектару, даже разрывают цветки. Есть еще одна интересная особенность в цветке горечавковых, способствующая наилучшему положению на нем посетителя. Тычинки, прирастая нижней частью нитей к трубке венчика, верхней вместе с пыльниками наклоняются к пестику, играющему роль центральной колонны, в результате возникает 4-5 отверстий - трубочек, похожих на "гнезда" барабана револьвера и ведущих на дно цветка. Насекомое, зондируя цветок, принимает позы, удобные для себя, и вращает его. Такие цветки называют вращающимися или "револьверными" цветками. Подобное устройство можно видеть у горной горечавки бесстебельной (Gentiana acaulis), из середины листовой розетки которой поднимается большой (высотой до 6 см) прекрасный синий цветок.
Перекрестное опыление может и не совершиться. Это происходит в отсутствии нужных опылителей, в случае длительной ненастной погоды, туманов, особо суровых в иные годы погодных условий в высокогорьях, когда цветки закрыты. Во всех этих случаях может произойти самоопыление. Оно совершается благодаря способности тычинок и рылец к движениям. К концу цветения нити тычинок поворачиваются и склоняются к столбику, касаясь его рылец пыльниками с остатками пыльцы в них. Это происходит, в частности, у сверций. Иногда активная роль в самоопылении принадлежит рыльцу, лопасти которого перекручиваются и, отклоняясь назад, касаются воспринимающими поверхностями основания столбика, осыпанного пыльцой (горечавки и др.). Наконец, собственная пыльца попадает на рыльце, постепенно проталкиваясь из складок трубки венчика при ежедневном его скручивании и раскручивании, особенно если цветок при этом наклонился и его колеблет ветер. У горечавки береговой обнаружены и клейстогамные цветки.
Плод горечавковых - септицидная коробочка, открывающаяся по швам завязи, но у тропической африканской хиронии (Chironia) и у растущего на Яве триптероспермума (Tripterospermum) плоды ягодообразные. Семена мелкие, с маленьким зародышем и очень обильным эндоспермом; они крылатые, с гладкой кожурой или бескрылые, с сетчатой, морщинистой, бугорчатой поверхностью. Распространяются семена ветром, а бескрылые и водой, при этом их морщинки и бугорки набухают и образуют вокруг семени слизистый футляр, который и защищает его, и облегчает скольжение в потоке.
Географическое распространение горечавковых свидетельствует о большой древности семейства. Самая общая картина их распространения в настоящее время очень выразительна. Наиболее крупные роды, составляющие около 70% видов, такие, как горечавка (в узком смысле, около 250 видов), горечавочка (около 250 видов), галения (около 80 видов), сверция (около 80 видов), золототысячник (50 видов), распространены в пределах всего северного полушария, особенно в горах, в том числе и тропического пояса. Кроме того, золототысячник вместе с первыми тремя родами растет еще и в горах Северной и Южной Америки, Австралии и Новой Зеландии, а виды сверции распространены и в горах Восточной Африки.

Таблица 51. Горечавковые, вахтовые и маслиновые: 2 - горечавка холодная (C.algida), Дальний Восток
Остальные многочисленные роды семейства характеризуются более строгой приуроченностью к определенным областям Земли (горы тропической зоны).
Горечавковые относятся к числу семейств, имеющих большое значение в жизни человека. Это лекарственные растения, которые в народной медицине всех стран, в том числе Китая и Индии, используются уже многие тысячелетия, используются настолько усердно, что горечавка желтая в Европе в диком виде уже почти не существует. Использует их и официальная медицина. Растения эти содержат так называемые горечи. Это глюкозиды (генциопикрин), гликозиды (генцизин, генциамарин и др.), алкалоиды (генцианин), флавоноиды и многие другие вещества. Используются корни, корневища и трава, из экстрактов которых приготовляют настои, порошки и т. д. Горечавковые очень декоративные растения, и наши горечавки, сверции, галении очень украсили бы берега водоемов в парках.

Таблица 51. Горечавковые, вахтовые и маслиновые: 1 - горечавка крестообразная(Centiana cruciata), Южный Крым
Семейство вахтовые (Menyanthaceae) (В. В. Письяукова)
Это небольшое семейство состоит всего из 5 родов (около 40 видов), среди которых только болотноцветник (Nympoides) содержит 25 видов и вилларсия (Villarsia) - 10, а вахта (Menyanthes), фория ( Fauria) и липарофиллум (Liparophyllum) - только по одному виду. Вахтовые относили к горечавковым в качестве подсемейства, и по многим признакам они действительно сходны. Однако обнаружилось большое число серьезных отличий, позволивших выделить вахтовые в качестве самостоятельного семейства. В отличие от горечавковых все вахтовые - многолетние водно-болотные травы с простертыми стеблями, несущими только очередные листья, которые оставляют на них характерные кольчатые рубцы. У болотно-цветника листья и цветоножки развиваются не на главном стебле, а на длинных шнуровидных боковых, которые начинают ветвиться лишь под самой поверхностью воды. Листья почковидные, яйцевидные, продолговатые, цельнокрайные или городчато-зубчатые и тройчатые. В отличие от горечавковых в проводящей системе никогда не бывает внутренней (интраксилярной) флоэмы, а многочисленные сосудистые пучки, идущие к членам цветка, в цветоложе образуют сложную кольцевую систему (в отличие от рассеянного их расположения у горечавковых). Цветки белые, розовые, желтые, всегда 5-членные, в почкосложении створчатые (но никогда не скрученные), краями лепестков внутрь завернутые (рис. 197); нектарники только торальные в виде 5 желёзок, расположенных у основания завязи и чередующихся с тычинками; завязь обычно верхняя, но у вилларсии, например, полунижняя, так как основание гинецея слегка погружено в цветоложе и члены цветка прикрепляются выше его (эпигиния), столбик всегда хорошо развит, с двулопастным рыльцем. Биология опыления у вахтовых и приспособления к нему сходны с этими же процессами у горечавковых, у них также развита гетеростилия, цветки у большинства видов живут долго, открываются и закрываются в зависимости от времени суток или состояния погоды. Если же перекрестного опыления не произошло, происходит автогамия, так же как и у горечавковых, в закрытых цветках (по своему строению хазмогамных, а не специальных клейстогамных). Коробочки при созревании иногда не открываются (у болотноцветника и др.) или открываются короткими зубцами лишь на ее верхушке (у вахты трехлистной). Семена с очень твердой кожурой, на поверхности которой часто возникают крючковатые волоски, шипики и щетинки (у болотноцветника, вилларсии), и в этом случае их могут разносить плавающие вокруг животные. В связи с химическими исследованиями также обнаружены заметные отличия горечавковых и вахтовых. Прежде всего среди глюкозидов, свойственных обоим семействам, у вахтовых никогда не обнаруживается генциопикрин и С-гликофлавон. Исследование всех 5 родов вахтовых и 23 родов горечавковых на содержание L-(+)-борнеситола показало полное их отсутствие у первых, в отличие от вторых. Таким образом, будучи во многом сходны с горечавками и, несомненно, родственны им, вахтовые все же обособлены довольно хорошо.

Таблица 51. Горечавковые, вахтовые и маслиновые: 3 - горечавка семираздельная (C.septemfida), парк Ботанического института АН СССР в Ленинграде
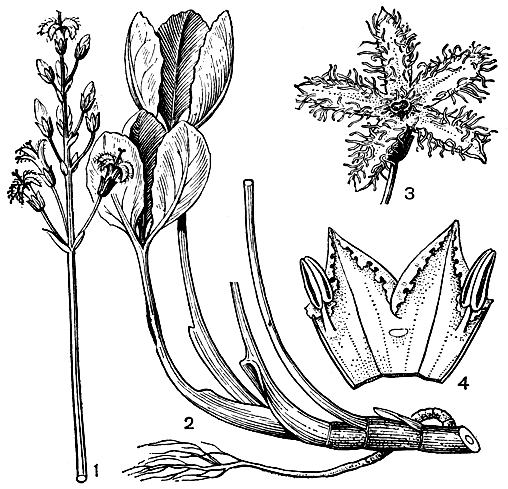
Рис. 197. Вахтовые. Вахта трехлистная (Menyanthes trifoliata): 1 - соцветие; 2 - основание растения; 3 - цветок в развернутом виде, лепестки окаймлены густыми бахромками. Фория гребневая (Fauria cristagalli); 4 - лепестки с надрезанно-зубчатыми и завернутыми краями
Географические позиции вахтовых на Земле представляются в следующем виде: вахта трехлистная (M. trifoliata, табл. 51) занимает все внетропические области северного полушария, т. е. Голарктику; виды болотноцветника от тропической зоны доходят до умеренных широт, и болотноцветник щитовидный широко распространен во всей Евразии; фория растет на острове Итуруп и в Японии и на противоположной стороне Тихого океана в Северной Америке; вилларсия ограничена Австралией, кроме одного вида, распространенного в Южной Африке, а липарофиллум известен только на Новой Зеландии и Тасмании.

Таблица 51. Горечавковые, вахтовые и маслиновые: 4 - вахта трёхлистная (Menyanthes trifoliate), окрестности Киева
Значение вахтовых в жизни людей невелико. Очень широко используется в народной и официальной медицине во всех странах северного полушария вахта трехлистная. Листовые пластинки ее содержат горькие глюкозиды - мениантин, мелиатин, алкалоид генцианин, флавоноиды - рутин, гиперозид и др.; корни также содержат горечь-мелиатин, кроме того, инулин и другие вещества. В траве обнаружено значительное количество йода. Назначение экстрактов из вахты для лечебных целей такое же, как и горечавковых, но особенную ценность имеют общетонизирующие свойства ее горечей.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'