Растительная клетка (Н. И. Арронет)
Растение, как и всякий живой организм, состоит из клеток, причем каждая клетка порождается тоже клеткой. Клетка - это простейшая и обязательная единица живого, это его элемент, основа строения, развития и всей жизнедеятельности организма.

Таблица 2. Мхи и печеночник: мхи - политрихум
Существуют растения, построенные из одной-единственной клетки. К ним относятся одноклеточные водоросли и одноклеточные грибы. Обычно это микроскопические организмы, но есть и довольно крупные одноклеточные (длина одноклеточной морской водоросли ацетабулярии достигает 7 см). Большинство растений, с которыми мы сталкиваемся в повседневной жизни, - это многоклеточные организмы, построенные из большого числа клеток. Например, в одном листе древесного растения их около 20 000 000. Если дерево имеет 200 000 листьев (а это вполне реальная цифра), то число клеток во всех них составляет 4 000 000 000 000. Дерево в целом содержит еще раз в 15 больше клеток.
Таблица 3. Лишайники, папоротники, хвощи: пармелия (на стволе дерева)
Растения, за исключением некоторых низших, состоят из органов, каждый из которых выполняет свою функцию в организме. Например, у цветковых растений органами являются корень, стебель, лист, цветок. Каждый орган обычно построен из нескольких тканей. Ткань - это собрание клеток, сходных по строению и функциям. Клетки каждой ткани имеют свою специальность. Выполняя работу по своей специальности, они вносят вклад в жизнь целого растения, которая состоит в сочетании и взаимодействии разных видов работы различных клеток, органов, тканей.
Таблица 3. Лишайники, папоротники, хвощи: ксантория (на ветке дерева)
Основными, самыми общими компонентами, из которых построены клетки, являются ядро, цитоплазма с многочисленными органоидами различного строения и функций, оболочка, вакуоль. Оболочка покрывает клетку снаружи, под ней находится цитоплазма, в ней - ядро и одна или несколько вакуолей. Как строение, так и свойства клеток разных тканей в связи с их разной специализацией резко различаются. Перечисленные основные компоненты и органоиды, о которых речь пойдет дальше, развиты в них в различной степени, имеют неодинаковое строение, а иногда тот или иной компонент может вовсе отсутствовать.

Таблица 3. Лишайники, папоротники, хвощи: папоротник
Главнейшими группами тканей, из которых построены вегетативные (непосредственно не связанные с размножением) органы высшего растения, являются следующие: покровные, основные, механические, проводящие, выделительные, меристематические. В каждую группу обычно входит несколько тканей, имеющих сходную специализацию, но построенных каждая по-своему из определенного вида клеток. Ткани в органах не изолированы друг от друга, а составляют системы тканей, в которых элементы отдельных тканей чередуются. Так, древесина - это система из механической и проводящей, а иногда и основной ткани.

Таблица 3. Лишайники, папоротники, хвощи: папоротник; хвощ

Таблица 4. Голосеменные: можжевельник
Возьмем для примера два органа высшего растения - лист и стебель. И мы увидим, насколько разнообразны по строению и работе клетки, образующие их ткани (рис. 22 и 23). Мы увидим также, как приспособлено строение клеток каждой ткани к выполнению их специальных функций.

Таблица 4. Голосеменные: ель

Таблица 4. Голосеменные: лиственница (осенью)
Покровные ткани - эпидермис, пробка, корка - защищают органы растения от неблагоприятных воздействий: от высыхания, перегрева, переохлаждения, лучистой энергии, механических повреждений, излишнего намокания, от проникновения чужеродных организмов, например бактерий. Эпидермис покрывает обе поверхности листа, молодые побеги и лепестки. Как правило, это один слой клеток. Те стенки клеток, которыми они соприкасаются между собой, обычно извилистые, причем выступ одной заполняет вогнутость соседней. Это способствует прочному соединению клеток, которые в результате образуют единый плотный пласт. Внешние, граничащие с воздухом стенки клеток более толстые.
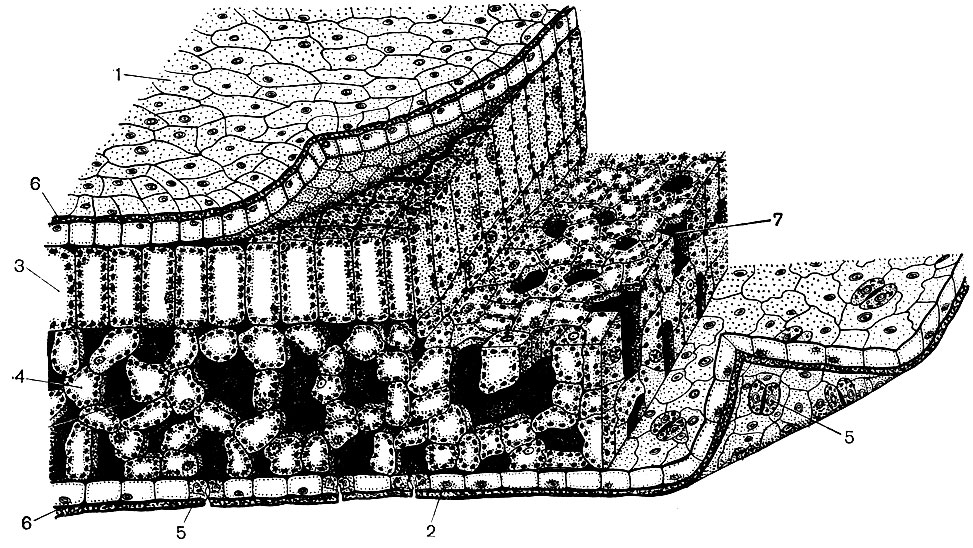
Рис. 22. Клеточное строение части листа двудольного растения. Показаны верхняя и частично нижняя поверхность, продольный и поперечный срезы. Области проводящих путей - жилок - не изображены. В правой части слой верхнего эпидермиса отогнут. 1 - клетки верхнего эпидермиса; 2 - клетки нижнего эпидермиса; 3 - клетки столбчатой паренхимы; 4 - клетки губчатой паренхимы; 5 - замыкающие клетки устьиц, щель между каждой их парой - просвет устьица; 6 - кутикула, покрывающая слой как верхнего, так и нижнего эпидермиса; 7 - межклеточные пространства, т. е. полости между клетками, в данном случае - губчатой паренхимы (они заполнены воздухом, сообщаются между собой, а через просветы устьиц - и с внешней средой; на рисунке они изображены темным). Ядра имеются во всех клетках эпидермиса и листовой паренхимы, но на срезах они видны не везде, так как во многих клетках срез проходит в стороне от ядра, выше или ниже его. В паренхимных и околоустьичных клетках видны хлоропласты. Цитоплазма во всех представленных на рисунке клетках расположена пристеночно (изображена пунктиром)
Эпидермальные клетки выделяют вещества, откладывающиеся на них снаружи в виде пленки (например, кутин, образующий плотную кутикулу, и воск; кутин к тому же пропитывает и сами внешние стенки эпидермальных клеток). Толстая внешняя оболочка и наружная пленка из кутина и воска обеспечивают изоляционные, защитные свойства этой ткани. Сквозь ее слой затруднено проникновение газов, воды, бактерий. Однако воздух обязательно должен входить внутрь зеленых частей растения, а наружу выходят водяные пары и кислород. Обмен газами, а также водяным паром между атмосферой и внутренними тканями молодых органов, покрытых эпидермисом, осуществляется с помощью устьиц.
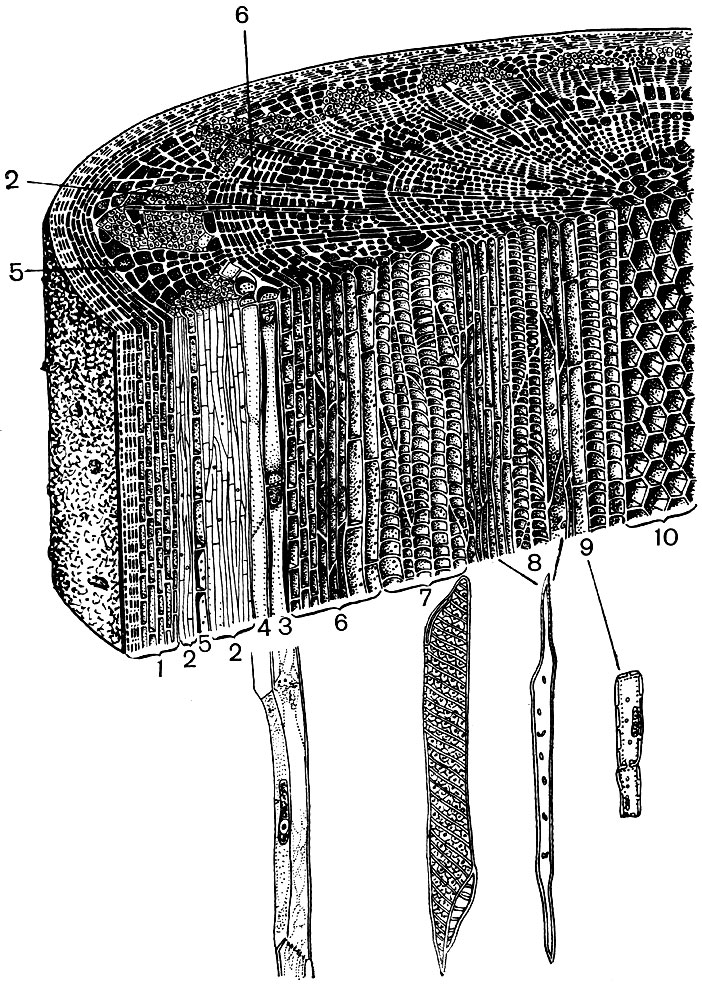
Рис. 23. Клеточное строение однолетнего стебля липы. Продольный и поперечный срезы: 1 - система покровных тканей (снаружи внутрь: один слой эпидермиса, пробка, первичная кора); 2 - 5 - луб: 2 - лубяные волокна, 3 - ситовидные трубки, 4 - клетки-спутники (3 и 4 вынесены на поля, где изображены более крупно), 5 - клетки лубяной паренхимы; 6 - клетки камбия, в крайних слоях растянутые, дифференцирующиеся; 7 - 9 клеточные элементы древесины: 7 - клетки сосудов, 8 - древесные волокна, 9 - клетки древесной паренхимы (7, 8 и 9 показаны также крупно); 10 - клетки сердцевины
Устьица образованы специальными клетками эпидермиса, между двумя из которых (так называемыми замыкающими клетками устьица) имеется отверстие регулируемой величины - устьичная щель. Через нее осуществляется связь между атмосферой и воздухоносными межклеточными пространствами толщи листа или другого органа, покрытого эпидермисом. Замыкающие клетки устьиц под влиянием перемены условий влажности или освещения меняют свою форму, смыкаясь друг с другом или размыкаясь. При этом они открывают или, соответственно, закрывают устьичную щель. На свету, когда растение фотосинтезирует и нуждается в притоке углекислого газа из атмосферы, устьичные щели открыты. Ночью они закрываются; замыкающие клетки закрывают просветы устьиц и в жаркое время дня, что предохраняет растение от большой потери воды, от увядания.
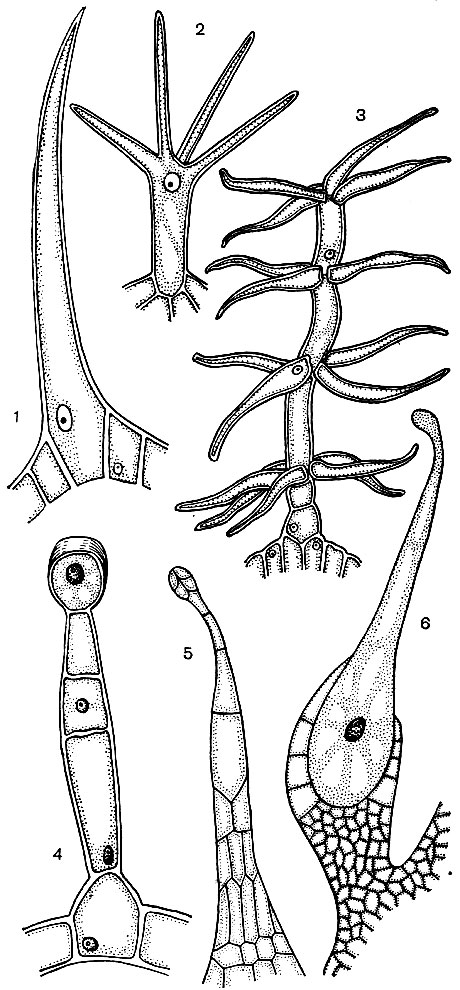
Рис. 24. Различные типы волосков эпидермиса: 1 — 3 — одноклеточные волоски; 4 — 6 — многоклеточные волоски; 1 — волосок подмаренника, 2 — разветвленные волоски обриеции, 3 — клетки-волоски коровяка, 4 — волоски герани, 5 — волоски камнеломки, 6 — жгучий волосок жгучей крапивы
Часто эпидермальные клетки образуют выросты - волоски (рис. 24). Иногда это многоклеточные образования, в других случаях каждый волосок представляет собой отросток одной клетки, лежащей в общем слое эпидермиса. Они играют защитную, опорную (например, у вьющихся или стелющихся растений), выделительную роль. Важны корневые волоски - трубчатые выросты эпидермальных клеток корня. Они увеличивают всасывающую поверхность последнего. Подсчитано, что на 14 млн. боковых корней одного растения ржи развивается до 14 млрд. корневых волосков. Площадь поверхности корней составляет при этом 232 м2, а волосков - 400 м2 . Эпидермис многих семян или плодов образует волоски, способствующие расселению семян, а тем самым растений. Часто волоски использует и человек. Пример этого - хлопок, представляющий собой эпидермальные волоски семян хлопчатника; его волоски имеют толстые целлюлозные оболочки и достигают в длину 70 мм.
В стебле многолетних растений под эпидермисом, на смену ему, развивается более грубая защитная ткань - пробка. Клетки ее отмирают, утрачивают протопласт и состоят только из толстых стенок, которые окружают полость, заполненную воздухом или смолистыми веществами. Стенки пропитаны суберином, делающим их водо- и воздухонепроницаемыми, теплоизолирующими, а также прочными, упругими. Они могут к тому же и одревесневать. Примером этой ткани служит береста. Она довольно тонка, а вот пробка пробкового дуба достигает толщины в несколько десятков сантиметров. Пробка в некоторых местах прерывается чечевичками - образованиями из иной ткани, которая проницаема для воды и воздуха. Через чечевички осуществляется обмен внутренних частей ствола с окружающей средой. Чечевички развиваются на местах устьиц.
В стебле под покровными тканями находятся клетки луба. Луб - это система из элементов нескольких тканей: проводящей, опорной, основной. Важнейший элемент луба - ситовидные трубки. Они построены из удлиненных клеток, вытянутых вдоль стебля, сочлененных друг с другом концами. Это живые, но безъядерные клетки, элементы цитоплазмы в которых расположены пристенно. Оболочки в местах стыка этих клеток имеют многочисленные отверстия, так что перегородки подобны ситу. В результате этого смежные клетки сообщаются между собой и тем самым оказываются объединенными в длинные трубки, тянущиеся сквозь жилки и черешки листьев, по стеблю и корню.
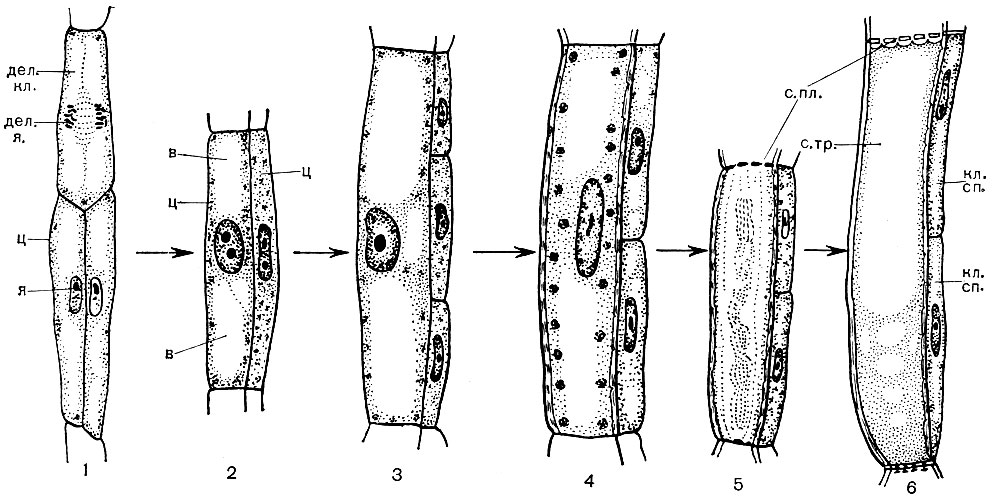
Рис. 25. Процесс дифференциации клетки: образование в стволе из меристематических (камбиальных) клеток специализированных клеток луба - ситовидных трубок и клеток-спутников: 1 - исходные камбиальные клетки: вверху - делящаяся клетка; внизу - две клетки, результат только что закончившегося деления их предшественницы, материнской клетки; 2 - начало дифференциации клеток: из левой клетки начинает образовываться ситовидная трубка, из правой - клетка-спутник. Левая клетка утолщается, в ней возникает и увеличивается вакуоль, цитоплазма занимает лишь пристеночную и околоядерную области. Правая клетка заполнена цитоплазмой с ядром, готовится к поперечному делению; 3 - следующий этап дифференциации: левая клетка выросла в длину и ширину, вакуоли увеличились, правая поделилась; 4 - у левой утолстилась клеточная стенка; 5 - в левой ядро рассасывается, в клеточных стенках, на стыках с соседними клетками (вверху и внизу), образуются поры, т. е. эти стенки превращаются в ситовидные пластинки; 6 - в левой клетке ядро рассосалось, в центре клетки - полость, ситовидные пластинки уже образовались; левая клетка близка к тому, чтобы стать ситовидной трубкой; правые клетки превратились в клетки-спутники. Дел. кл.- делящаяся клетка; дел. л.- делящееся ядро; я - ядро неделящееся; ц - цитоплазма; в - вакуоль; с. пл.- ситовидная пластинка; с. тр.- ситовидная трубка; кл. сп.- клетка-спутник
По системе ситовидных трубок продукты, образующиеся в зеленых частях растений, перемещаются ко всем его другим частям, питают их. Основным из транспортируемых продуктов является сахароза. Ситовидные трубки - это элемент проводящей ткани. Около клеток ситовидных трубок имеются клетки-спутники. Они содержат ядра, и их протопласт имеет непосредственные связи с цитоплазмой безъядерных ситовидных клеток. В лубе встречаются также участки, состоящие из паренхимных клеток (т. е. клеток, длина, ширина и высота которых близки по величине). В них откладываются различные вещества - крахмал, масла, смолы. Это клетки основной ткани.
Кроме того, луб содержит элементы опорной ткани - лубяные волокна. Это очень длинные клетки с утолщенными стенками; длина клетки может превышать ее ширину в тысячи раз. У льна длина тонких вытянутых лубяных волокон достигает 4 см, а у рами - даже 35 см. Обычно это мертвые клетки, без протопласта. Они выполняют механическую функцию, создавая прочность стебля.
Большинство специализированных клеток не способно к размножению. Однако растение растет всю свою жизнь, и в течение всей жизни в нем образуются новые клетки. Они развиваются из клеток образовательных - меристематических - тканей. Размножение делением является специализацией меристематических клеток, их функцией в организме. Происшедшие из них клетки развиваются, превращаясь в те или иные специализированные клетки - клетки ситовидных трубок, клетки пробки, волокна и т. д. Образовательная ткань находится в разных частях растения (например, в точках роста - на верхушках побегов, корней). В толще стебля обычно есть несколько слоев образовательной ткани. Под слоем пробки находятся клетки феллогена, которые, делясь, пополняют число клеток пробки, корки. Внутрь от луба располагается слой клеток камбия. Те из порождаемых ими клеток, которые образуются снаружи от камбия, развиваются в клетки луба (рис. 25). Оказавшиеся внутри, т. е. ближе к оси ствола, дают начало клеткам древесины.
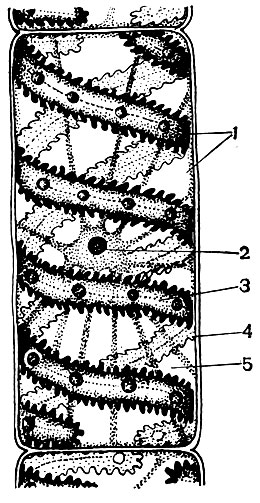
Рис. 26. Клетка многоклеточной нитчатой зеленой водоросли спирогиры сомнительной: 1 - цитоплазма; 2 - ядро с ядрышком; 3 - хроматофор, спиральной лентой вьющийся внутри клетки; 4 - оболочка; 5 - вакуоли
Древесина состоит из элементов проводящей, опорной и основной тканей. К первым относятся волокна древесины - длинные мертвые клетки с одревесневшими стенками; ко вторым - сосуды, представляющие собой результат слияния многих клеток; к третьим - клетки древесной паренхимы. Клетки, дифференцирующиеся из камбиальных элементов в проводящие (сосуды), растут в длину и ширину. Их боковые стенки утолщаются и одревесневают. В стенках, однако, остаются поры, закрытые лишь тонкой оболочкой. Перегородки на стыках смежных клеток исчезают, протопласт отмирает. В результате образуются длинные сосуды, состоящие из одних стенок. Проводящая система тянется сквозь корень и стебель в лист. По таким сосудам осуществляется восходящий ток воды и растворенных в ней солей от корней ко всем органам растения.
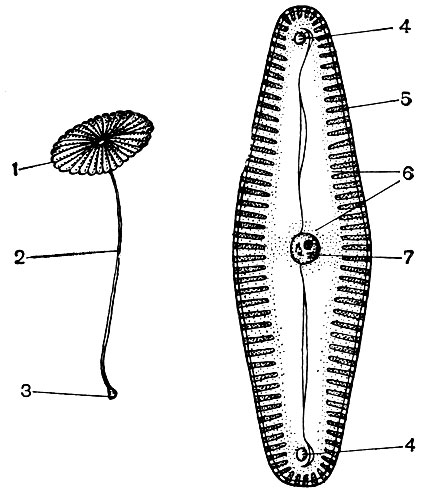
Рис. 27. Одноклеточные растения. Слева - морская зеленая водоросль ацетабулярия средиземноморская, состоящая из гигантской клетки, изображенной в натуральную величину: 1- шляпка; 2 - стебелек; 3 - ризоиды с ядром. Справа - пресноводная диатомовая водоросль пиннулярия зеленая: 4 - конечные узелки шва панциря; 5 - перистая структура панциря; 6 - цитоплазма; 7 - центральный узелок
В центре стебля находятся клетки сердцевины - округлые или многогранные паренхимные клетки. Это элементы основной ткани. Иногда они полые, и в них находится воздух; иногда они заполнены запасными питательными веществами, различными кристаллами, таннинами. Стенки их могут быть одревесневшими. Древесину и луб пронизывают радиальные лучи. Их клетки являются производными камбия и несут запасающую функцию. На уровне луба эти лучи заметно расширяются.
В мякоти листа между верхним и нижним эпидермисом находится основная ткань - клетки с тонкими оболочками и большим количеством зеленых пластид - хлоропластов. В этих клетках происходит фотосинтез. Верхние слои состоят большей частью из продолговатых клеток, плотно прилегающих друг к другу, - эта столбчатая паренхима. Нижние слои рыхлеет между клетками расположены межклетники - пространства, заполненные воздухом; это губчатая паренхима. Паренхима пронизана ветвящимися жилками, которые построены из пучков механической (разнообразные волокна) и проводящей (ситовидные трубки и водоносные сосуды) тканей.
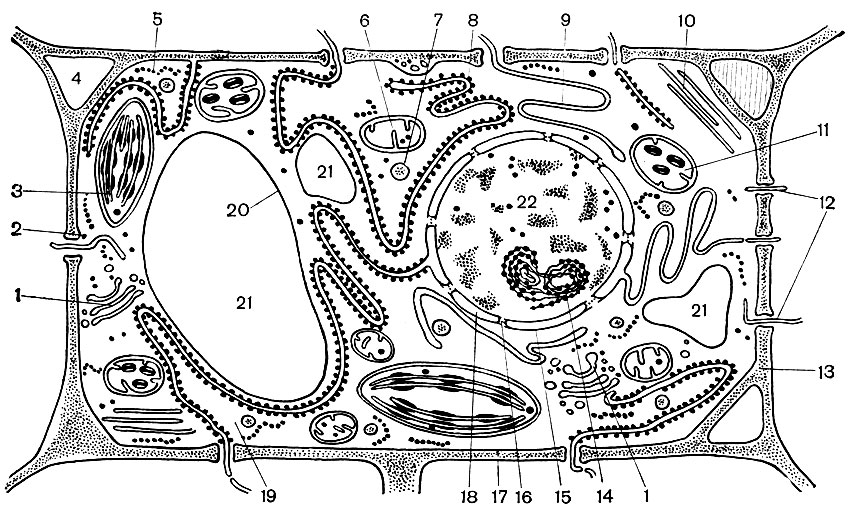
Рис. 28. Современная (обобщенная) схема строения растительной клетки, составленная по данным электронно-микроскопического исследования разных растительных клеток: 1 - аппарат Гольджи; 2 - свободно расположенные рибосомы; 3 - хлоропласты; 4 - межклеточные пространства; 5 - полирибосомы (несколько связанных между собой рибосом); 6 - митохондрии; 7 - лизосомы; 8 - гранулированная эндоплазматическая сеть; 9 - гладкая эндоплазматическая сеть; 10 - микротрубочки; 11 - пластиды; 12 - плазмодесмы, проходящие сквозь оболочку; 13 - клеточная оболочка; 14 - ядрышко; 15, 18 - ядерная оболочка; 16 - поры в ядерной оболочке; 17 - плазмалемма; 19 - гиалоплазма; 20 - тонопласт; 21 - вакуоли; 22 - ядро
Это краткое и схематичное изложение плана строения стебля, листа показывает, насколько разнообразны по величине, форме, строению, функции клетки одного и того же растения. Если взять разные растения, то можно встретиться с еще большим многообразием клеток. Однако, как правило, клетки одноименных тканей даже весьма далеких друг от друга растений сходны, поскольку эти клетки выполняют аналогичные функции. Клетки эпидермиса листа березы и, например, одуванчика более похожи друг на друга, чем эпидермальная клетка одуванчика и его же ситовидная клетка. Отличительные черты клетки связаны в первую очередь с ее специальной функцией. Клетки, специализированные в механической функции, имеют утолщенные и нередко одревесневшие оболочки. Специализация в фотосинтезе ведет к появлению в клетках хлоропластов. Специализация в проводящей функции связана с удлинением клеток, с утратой протопласта, с изменениями в оболочках клеток на стыках, благодаря которым полости смежных клеток сообщаются. Для клеток, специализированных в защитной функции, характерны многообразные изменения внешних стенок, наличие волосков, способность вырабатывать защитные вещества.
Специализация в функции запасания питательных веществ может привести к увеличению размеров клеток, к появлению в них очень крупных вакуолей. В клетках меристематических, усиленно делящихся, особенно развиты те внутриклеточные структуры, которые обеспечивают синтез веществ - составных частей клетки.
В зависимости от характера специализации одни черты строения и работы клетки развиты очень сильно, другие, напротив, мало развиты или вовсе отсутствуют. Как мы видели, некоторые клетки, став специализированными, умирают и именно в мертвом состоянии начинают выполнять свою специальную функцию в многоклеточном организме (клетки пробки, древесинные волокна, сосуды).
Очень разнообразны и своеобразны клетки многоклеточных нитчатых водорослей и одноклеточных растений (рис. 26 и 27). Клетка любого из последних к тому же сильно отличается от клеток многоклеточных растений. Ей одной приходится выполнять несколько функций, которые у многоклеточных растений поделены между клетками разных тканей.
В то же время даже очень отличающиеся между собой клетки обладают глубоким сходством в строении и функциях. Для многоклеточных это связано, во-первых, с тем, что все клетки организма (если исключить вегетативное размножение, при котором от материнского растения сразу отделяется в качестве новой особи или ее зачатка целый многоклеточный комплекс) являются потомками одной и той же клетки-родоначальницы. Поэтому, каким бы образом ни были специализированы клетки, они имеют общий исток, а потому являются родственницами.
Во-вторых, общие черты в строении клеток растений разных видов связаны с тем, что все растения состоят в той или иной степени родства. Все растения развились путем эволюции от одноклеточных общих предков. Общие черты унаследованы клетками современных растений от древних прародительских клеток. С этим связано также наличие общих черт строения и работы растительных и животных клеток.
В-третьих, сходство связано с тем, что все живые клетки, какую бы специальную функцию они ни несли в организме, прежде всего должны обеспечивать собственную жизнь. Клетки поглощают питательные вещества, перерабатывают их, добывая энергию и строя собственное тело, дышат, освобождаются от ненужных веществ, борются с различными повреждениями, реагируют на изменения внешних условий, перестраивая свою жизнедеятельность, растут. Все эти процессы у разных клеток осуществляются сходно и с помощью однотипных структур, общих по плану строения не только для разных растительных клеток, но и для клеток растений и животных. Надо сказать, что выполнение любой специальной функции клетки основывается на ее общих свойствах. Та или иная черта, способность, присущая всем клеткам, у специализированной клетки развивается особенно сильно и обеспечивает выполнение клеткой ее основной, специальной функции. Те общие черты, без которых невозможно выполнение этой специальной функции, в клетке сохраняются, а остальные могут утратиться. Мертвые специализированные клетки - крайний, предельный случай этого. Специальная функция таких клеток связана с их оболочкой; протопласт нужен лишь до тех пор, пока он создает оболочку; после этого он отмирает, и вся клетка состоит только из неживой оболочки, которая и работает на нужды растения.
Рассматривая общие черты строения и жизнедеятельности растительных клеток, удобно говорить о некоей типовой клетке, вобравшей в себя общие черты разных специализированных клеток. Такую клетку, хотя она и не существует в "чистом виде", можно даже изобразить (табл. 7, рис. 28). Из реальных клеток более других похожи на такую "типовую" клетки паренхимы листа (табл. 8, рис. 29).
Перейдем к описанию строения и работы растительной клетки.
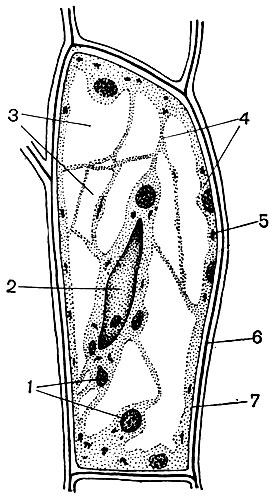
Рис. 29. Клетки из черешка листа сахарной свеклы: 1 - хлоропласты; 2 - ядро с ядрышком; 3 - вакуоли; 4 - цитоплазма; 5 - митохондрии; 6 - клеточная оболочка; 7 - тонопласт
Таблица 5. Покрытосеменные: вверху слева - цветок магнолии; вверху справа - цветки яблони; внизу - лотос.
Таблица 6. Покрытосеменные: вверху (слева направо) - колокольчики, пеларгониум, шафран (крокус); внизу слева - астры; внизу справа - репейник.
Снаружи растительная клетка покрыта оболочкой, неодинаковой по толщине и строению у разных клеток. Образующие ее вещества вырабатываются в цитоплазме и откладываются снаружи от нее, постепенно создавая оболочку. Этими веществами прежде всего являются крупномолекулярные полисахариды - пектин, гемицеллюлоза и в небольших количествах целлюлоза. Они образуют так называемую первичную оболочку. Она довольно эластична, по мере роста клетки растягивается и тоже растет, а потому не препятствует росту клетки. Однако она создает определенную прочность клетки и способна защитить ее от механических повреждений. Есть клетки, которые лишены такой первичной оболочки, - это некоторые клетки, служащие для бесполого и полового размножения (зооспоры и гаметы водорослей и низших грибов, мужские гаметы высших растений). У многих клеток имеется не только первичная, но еще и вторичная оболочка. Она образуется под первичной и построена главным образом из целлюлозы. Целлюлоза - это полисахарид, молекулы которого образуют тончайшие нити - микрофибриллы. В оболочке нити целлюлозы погружены в аморфное вещество, состоящее из пектиновых соединений. У одних клеток эти микрофибриллы целлюлозы расположены поперек длины клетки, кольцами; благодаря этому такие клетки могут растягиваться в длину, но не в ширину (например, клетки сосудов стебля). У других нити лежат продольно; клетки с такой оболочкой эластичны при растягивании поперек, но очень жестки на продольное растяжение. У третьих они расположены наискось, образуя спираль (эпидермальные волоски семян хлопчатника, лубяные волокна). Все это напоминает железобетонные конструкции, причем нити целлюлозы играют здесь роль железных прутьев, а пектиновые вещества - роль цемента. Клетки, имеющие вторичную оболочку, весьма прочны. Они образуют механические, опорные ткани растения. Иногда вторичная оболочка играет и роль склада питательных продуктов: образующие ее вещества могут превращаться в другие, более простые, которые расходуются как питание.
В оболочке имеются неутолщенные места - поры (в первичной оболочке они называются норовыми полями), через которые осуществляется связь между соседними клетками. Сквозь поровые поля и поры проходят тонкие тяжи цитоплазмы. Это плазмодесмы, которые связывают цитоплазму соседних клеток (табл. 9). По ним осуществляется обмен веществами между соседними клетками. Плазмодесмы наряду с элементами проводящей ткани соединяют клетки и ткани организма в единое целое.
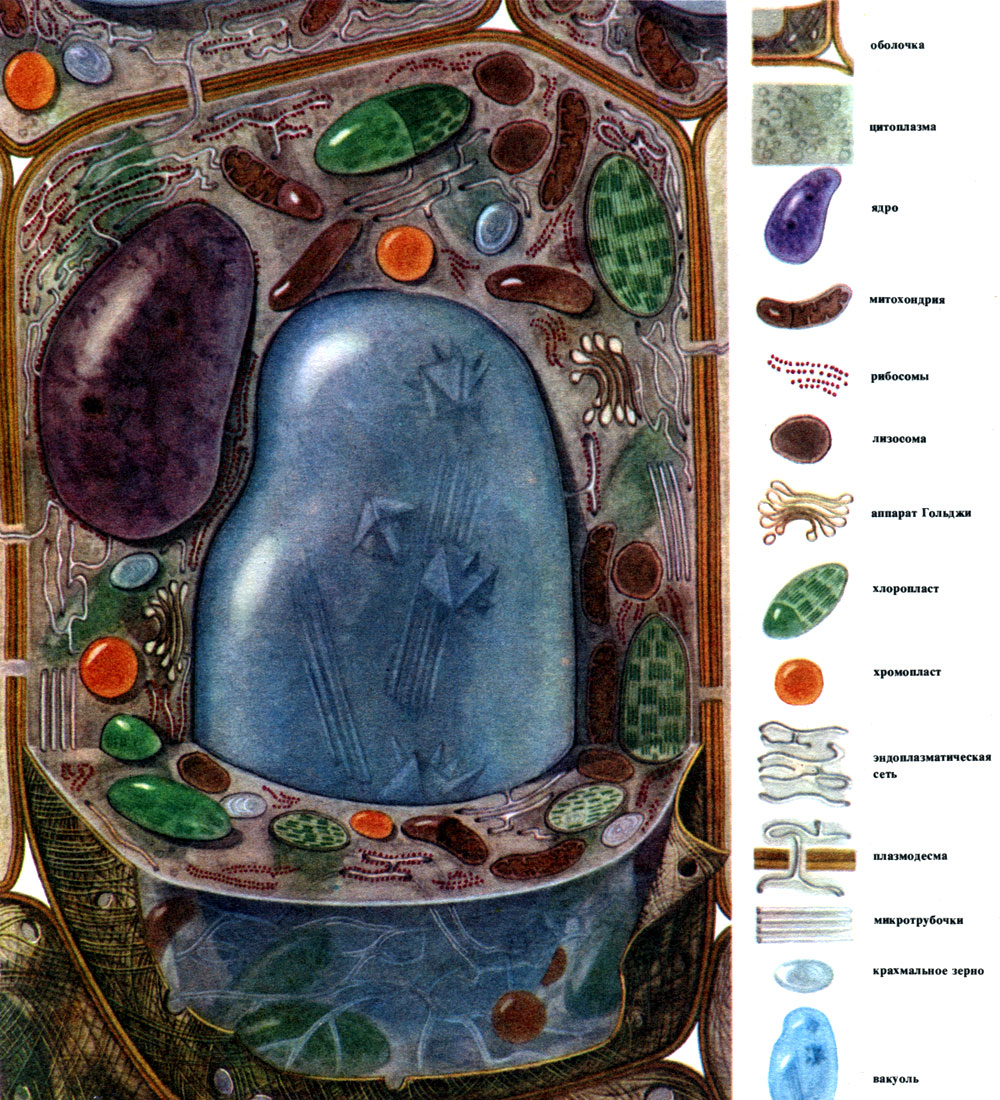
Таблица 7. Схема строения растительной клетки
Обмен веществами и распространение возбуждения позволяют клеткам влиять на развитие и работу друг друга, и каждая ткань влияет на жизнедеятельность всех других тканей. Этим создается координация работы всех частей единого организма-целого растения.
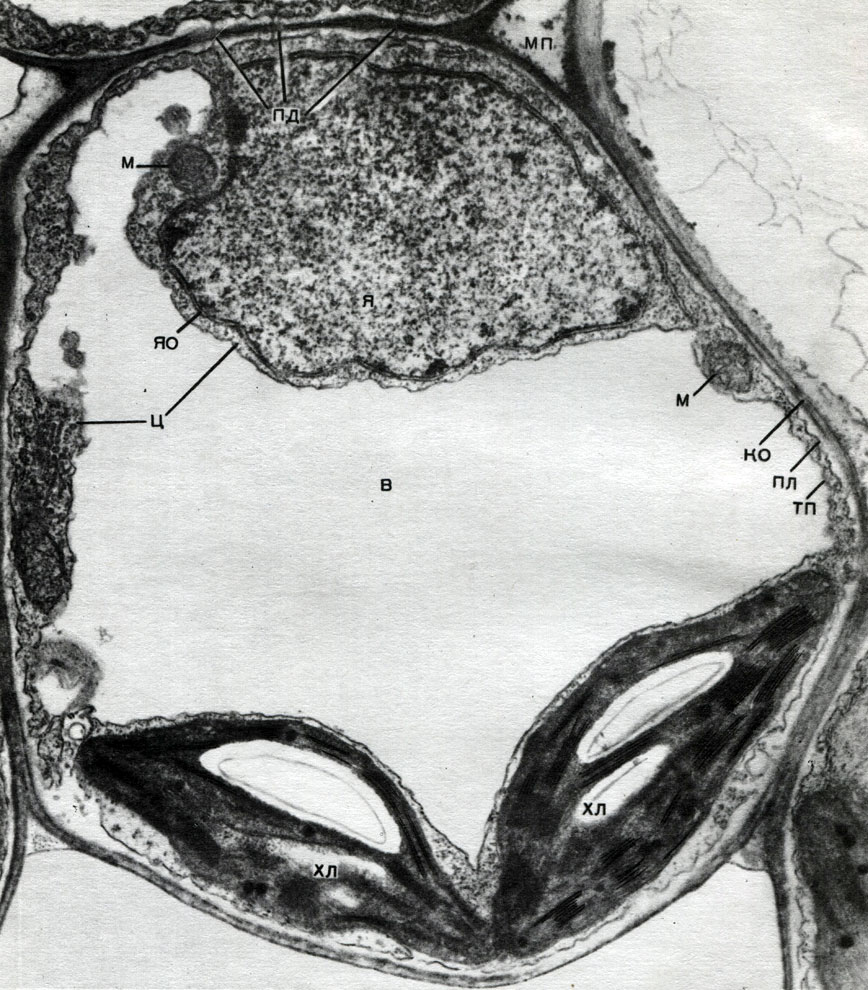
Таблица 8. Клетка из губчатой паренхимы листа коровяка (Verbascum thapsus). Электронная микрофотография (увел, х 25 000) М. Ф. Даниловой: ко - клеточная оболочка; пд - плазмодесмы, соединяющие цитоплазму соседних клеток; мп - межклеточное пространство; ц - цитоплазма; я - ядро; в - вакуоль; яо - ядерная оболочка; м - митохондрии; хл - хлоропласты; пл - плазмалемма; тп - тонопласт
Во многих клетках клеточная оболочка с возрастом пропитывается веществами, еще более укрепляющими ее. Накопление в ней лигнина ведет к одревеснению оболочки. Одревесневают стенки клеток древесины и, часто, лубяных волокон, оболочки клеток кожуры семян, иногда околоплодника (рис. 30) и даже лепестков. Клетки с одревесневшими стенками не только прочнее, но и менее проницаемы для микробов, для воды. Оболочки некоторых других клеток - покровных тканей, а также на поверхностях поранений - опробковевают, пропитываясь суберином. Содержимое клеток после этого отмирает, так как суберин непроницаем для воды и газов, но ткань из таких мертвых клеток защищает внутренние живые части растения от вредных внешних воздействий. Как уже говорилось, наружные оболочки клеток эпидермиса обычно пропитываются или покрываются кутином и воском, и это сохраняет клетки от излишнего испарения воды, от проникновения инфекции. Оболочка может пропитываться и минеральными веществами, преимущественно солями кальция.
Бывают, однако, в жизни клеток и такие моменты, когда их оболочка должна стать менее прочной, чем была до этого. Это происходит, например, при прорастании семян.
Полисахаридная клеточная оболочка - характерная черта строения растительной клетки, отличающая ее от животной клетки.
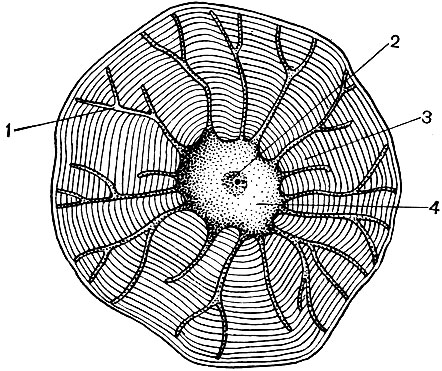
Рис. 30. Живая клетка из скорлупы кокосового ореха с ветвистыми каналами и очень толстой одревесневшей оболочкой: 1 - поровые каналы, заполненные цитоплазмой; 2 - ядро; 3 - слоистая оболочка клетки; 4 - цитоплазма
Оболочка, или клеточная стенка, - это защитное образование. Под оболочкой находится цитоплазма. Самый наружный ее слой, примыкающий к оболочке, - поверхностная клеточная мембрана - плазмалемма. Она представляет собой комбинацию слоев жироподобных и белковых молекул. Такие мембраны называются липопротеиновыми ("липос" - жир, "протеин" - белок). Мембрана подобной конструкции отграничивает цитоплазму от вакуолей, эта мембрана называется тонопластом. Многие органоиды клетки построены из липопротеиновых мембран. Однако в каждом случае мембрана построена из жироподобных веществ (липидов) и белков, присущих именно данной мембране. Качественное разнообразие липидов и особенно белков колоссально, отсюда огромное разнообразие мембран, отличающихся по свойствам, и в пределах одной клетки, и в разных клетках.
Плазмалемма регулирует вход веществ в клетку и выход их из нее, обеспечивает избирательное проникновение веществ в клетку и из клетки. Скорость проникновения сквозь мембрану разных веществ различна. Хорошо проникают через нее вода и газообразные вещества. Легко проникают также жирорастворимые вещества, - вероятно, благодаря тому, что она имеет липидный слой. Предполагается, что липидный слой мембраны пронизан порами. Это позволяет проникать сквозь мембрану веществам, нерастворимым в жирах. Поры несут электрический заряд, поэтому проникновение через них ионов не вполне свободно. При некоторых условиях заряд пор меняется, и этим регулируется проницаемость мембран для ионов. Однако мембрана неодинаково проницаема и для разных ионов с одинаковым зарядом, и для разных незаряженных молекул близких размеров. В этом проявляется важнейшее свойство мембраны - избирательность ее проницаемости: для одних молекул и ионов она проницаема лучше, для других хуже.
Вещество движется в клетку, диффундируя в нее, если его концентрация снаружи больше, чем концентрация внутри. В противном случае оно должно диффундировать из клетки. Но, проникнув в клетку, молекулы данного вещества могут тут же вовлечься в обмен веществ и подвергнуться превращению в другие вещества. Тогда концентрация этого вещества внутри клетки снова падает, и новая порция его молекул диффундирует внутрь клетки. В результате в клетки, активно расходующие то или иное вещество, последнее усиленно поступает из окружающих клеток.
Другой механизм усиленного поглощения вещества клеткой состоит в его связывании белками или другими веществами. Связываясь, вещество уходит из внутриклеточного раствора и не препятствует дальнейшей диффузии этого же вещества. Оно продолжает поступать в клетку и скапливается в ней в связанной форме. Так, в клетках некоторых морских водорослей йод накапливается в концентрациях, в миллион раз превышающих его концентрацию в морской воде.
Во всех этих случаях речь идет о движении веществ путем диффузии. Однако клеточные мембраны располагают и механизмами перемещения веществ через себя от меньшей их концентрации к большей. Среди белков мембраны имеются белковые вещества, работа которых состоит в перемещении различных веществ с одной стороны мембраны на другую. Они называются транспортными ферментами. Вероятно, такой транспортный фермент образует с переносимым веществом промежуточное соединение; последнее проходит сквозь мембрану, расщепляется затем на исходные части, после чего фермент возвращается назад, а перенесенное вещество остается по другую сторону мембраны. Детали этого процесса неизвестны, но сам факт такого активного транспорта веществ несомненен. В отличие от диффузного, пассивного, ферментативный транспорт нуждается в затрате энергии. Вероятно, именно благодаря такому активному транспорту ионов клетки эпидермиса корней способны всасывать из почвы нужные растению неорганические вещества и затем передавать их по растению от клетки к клетке.
При химических или физических изменениях во внешней и внутренней среде клеточные мембраны изменяют свою проницаемость, а также степень и сам характер ее избирательности. На этом основываются механизмы регуляции движения веществ в клетку и из клетки. Изменение проницаемости мембран для питательных веществ отражается на интенсивности обменных процессов в клетке, на характере протекающих в ней синтезов, на всей ее жизнедеятельности. В изменении проницаемости мембран для ионов натрия и калия состоит механизм распространения клеточного возбуждения - возникновения и перемещения биотока. К поверхности мембраны присоединено большое количество катионов, главным образом ионов калия. Поэтому она несет снаружи положительный заряд. Под влиянием раздражителей ионы калия отщепляются от участка, подвергшегося воздействию, заряд участка падает и он становится электроотрицательным по отношению к соседним участкам мембраны. Эта электроотрицательность является, в свою очередь, раздражителем для соседних участков, тем же путем снимая их заряд, деполяризуя их. Волна электроотрицательности распространяется по мембране - это и есть биоток. Затем ионы калия снова садятся на мембрану, придавая ей исходный заряд, - за волной деполяризации следует волна восстановления. Поверхности плазмодесм, проходящих сквозь клеточные стенки и соединяющих цитоплазму соседних клеток, тоже образованы такими мембранами. Биоток движется и по ним, распространяясь от клетки к клетке. Биотоки растительной клетки пока еще мало изучены. Однако ясно, что они являются способом сигнализации, используемым в растительной клетке для пуска в ход одних химических реакций и торможения других. Дело в том, что все химические процессы, протекающие в клетке и составляющие сущность ее жизнедеятельности, идут с участием биологических катализаторов - ферментов. Каждая реакция возможна лишь тогда, когда фермент, пускающий ее в ход, активен. Большинство ферментов становятся активными под действием тех или иных неорганических катионов: К., Na., Ca.., Mg.., Mn... Биоток, меняя проницаемость мембран для заряженных веществ, создает условия для их проникновения сквозь мембраны и контакта с ферментами. Тем самым биоток включает в действие те или иные ферменты и этим регулирует, направляет обмен веществ в клетке. Регуляция обмена веществ биотоками - это лишь один из многих способов регуляции внутриклеточного метаболизма.
Сказанное о проницаемости поверхностной мембраны клетки - плазмалемме - относится и к другим внутриклеточным мембранам, в том числе к тем, из которых построены многие органоиды клетки.
Цитоплазма, когда-то считавшаяся однородным коллоидным раствором белковых веществ, на самом деле сложно структурирована. По мере развития микроскопической техники исследования выяснялись все более тонкие детали строения цитоплазмы. В цитоплазме были открыты различные органоиды (органеллы) - структуры, каждая из которых выполняет определенные физиологические и биохимические функции. Важнейшими органоидами цитоплазмы являются митохондрии, эндоплазматический ретикулум (эндоплазматическая сеть), аппарат Гольджи, рибосомы, пластиды, лизосомы. У подвижных клеток (зооспоры и гаметы водорослей, сперматозоиды хвощей, папоротников, саговников, некоторые одноклеточные и колониальные водоросли) имеются органоиды движения - жгутики.
Особенно много новых фактов о тонком строении цитоплазмы принесло и приносит использование электронного микроскопа, позволяющего исследовать детали строения самих органоидов. Современные биофизические и биохимические методы позволяют выделять в чистом виде те или иные органоиды цитоплазмы и затем изучать их химический состав и их функции. Вне клетки, в средах сложного состава, многие органоиды способны выполнять ту работу, которую они производят, когда находятся в клетке.
Часть цитоплазмы, в которую погружены органоиды и которая пока что представляется бесструктурной, называется основным веществом цитоплазмы или гиалоплазмой. Гиало-плазма - это отнюдь не пассивный наполнитель, а активно работающая часть цитоплазмы. В ней протекает ряд жизненно необходимых химических процессов, в ее состав входят многие белки-ферменты, при помощи которых эти процессы осуществляются.
Жизнь клетки состоит в непрерывной химической работе, которая в своей совокупности называется обменом веществ. По существу, клетка представляет собой химический завод, вырабатывающий большой ассортимент продукции и самостоятельно добывающий энергию, необходимую для ее производства. Ее продукцией являются вещества, которые необходимы и ей самой для поддержания ее собственной жизни (для построения своего тела при росте и развитии, для замены своих сносившихся частей), и для создания дочерних клеток при размножении, и для нужд других клеток организма .
Все химические реакции, протекающие в клетке, можно разделить на две группы. В результате одних те или иные вещества распадаются на более мелкомолекулярные. В результате других из мелкомолекулярных веществ синтезируются вещества с более крупными молекулами. Молекула любого вещества состоит из атомов, которые удерживаются между собой химическими связями, т. е. тем или иным количеством сконцентрированной потенциальной химической энергии. Когда молекула дробится, связи рвутся и их химическая энергия освобождается. Для синтеза, т. е. для образования более крупной молекулы из мелких, нужно создать новые химические связи, В них необходимо вложить некоторую порцию энергии. Напротив, реакции распада в конечном итоге идут с освобождением энергии, так как при них связи между атомами рвутся. Биологический смысл реакций распада, идущих в клетке, состоит в том, что при них освобождается химическая энергия, используемая затем клеткой для реакций синтеза и для производства иных видов работы (электрической, механической, работы по транспорту веществ). Так как клетка всю свою жизнь синтезирует различные вещества, то ей приходится непрерывно расщеплять другие вещества. Освобождающаяся энергия используется для синтезов не сразу. Сначала она запасается путем образования специальных веществ - аккумуляторов химической энергии - аденозинтрифосфорной кислоты (АТФ) и родственных ей соединений. В нужный момент и в соответствующей точке клетки АТФ расщепляется и отдает энергию для синтеза необходимого клетке вещества.
Все химические реакции, протекающие в клетке, - и синтеза и распада - осуществляются с помощью ферментов. Ферменты - белковые вещества, ускоряющие течение реакций. Ускорение это настолько велико, что без ферментов подобные реакции вообще были бы невозможны в клетке. Известны случаи, когда благодаря ферменту реакция ускоряется в 1011 раз. Это значит, что реакция, заканчивающаяся с участием фермента в течение 0,01 сек, без него протекала бы 31 год. Понятно, что такие реакции без фермента были бы просто нереальными.
Кроме того, благодаря ферментам течение химических реакций в клетке управляемо, регулируемо. Активность ферментов в клетке меняется в соответствии с ее потребностью в определенном веществе или в энергии, т. е. в конечном результате работы этих самых ферментов. Когда появляется потребность в каком-то веществе, включаются в действие или заново синтезируются те ферменты, благодаря которым оно образуется. Образование веществ, освобождение и запасание энергии - это итог последовательных реакций, результат работы целой цепи ферментов. Все структуры живых частей клетки построены именно из ферментов и из веществ, скрепляющих эти ферменты. При этом ферменты, участвующие в смежных, последовательно протекающих реакциях, и расположены рядом. Они передают молекулы превращаемых ими веществ друг другу, как по конвейеру, причем каждый из них совершает над молекулой свою рабочую операцию.
В гиалоплазме содержатся ферменты, расщепляющие молекулы глюкозы на более простые молекулы пировиноградной кислоты. Освобождающаяся при этом энергия запасается путем образования молекул АТФ. Тот же процесс протекает и в клеточном ядре. Однако основная масса энергии добывается в особых органоидах цитоплазмы - митохондриях, так как там происходит более глубокое расщепление веществ.
Митохондрии - мелкие тельца округлой или продолговатой формы, размером 0,5 - 1,5 мк, т. е. величиной с бактерию. Число их в клетке обычно велико, порядка 100 - 3000. Бывают, однако, клетки и с малым количеством митохондрий. Так, в спермин морской водоросли фукуса содержится всего 4 митохондрии, а в одноклеточной водоросли микромонас - одна. Митохондрии видны под световым микроскопом, однако их тонкое строение можно изучать лишь с помощью электронного микроскопа (табл. 10, схема строения - рис. 31). Митохондрии - это образования, построенные из липопротеиновых мембран, погруженных в основное вещество - матрикс. Оболочка митохондрии образована двумя мембранами, между которыми имеется промежуток.
Внутренняя из мембран оболочки дает многочисленные впячивания внутрь, это кристы. Между ними находится матрикс. И внутренняя мембрана оболочки митохондрии, - и образуемые ею кристы построены из упорядочение расположенных ферментов. Благодаря складкам - кристам рабочая поверхность мембран внутри митохондрий очень велика. Ряд ферментов находится в матриксе митохондрии, т. е. между кристами.
Совокупность этих ферментов осуществляет внутриклеточное дыхание и запасание освобождающейся при дыхании энергии в форме АТФ. Работа митохондрий тесно связана с процессами, идущими в гиалоплазме, где протекают первые этапы расщепления глюкозы и других веществ до пировиноградной кислоты. В митохондриях же протекает дальнейшее ее расщепление. Пировиноградная кислота проникает в митохондрии и здесь ступенчато, шаг за шагом, окисляется до углекислого газа и воды, причем одновременно потребляется кислород. Это и есть внутриклеточное дыхание, при котором клетка, расщепляя и окисляя вещества, добывает очень много энергии, которую она потом может использовать для самых разных своих нужд.
Первый этап расщепления молекулы глюкозы, во время которого она дробится пополам и который протекает в гиалоплазме, дает клетке всего лишь две молекулы АТФ.
В результате второго этапа, приводящего к полному "сгоранию" глюкозы, образуется еще 36 молекул АТФ. Поэтому митохондрии по своей функции - это силовые станции клетки, машины для добывания основного количества энергии. Само расщепление продуктов распада глюкозы происходит в матриксе митохондрии, АТФ же образуется благодаря реакциям, разыгрывающимся на внутренних ее мембранах, в состав которых входят дыхательные ферменты и ферменты, обеспечивающие образование АТФ. Количество крист в митохондриях может быть различным. Чем их больше, тем выше биохимическая активность митохондрий.
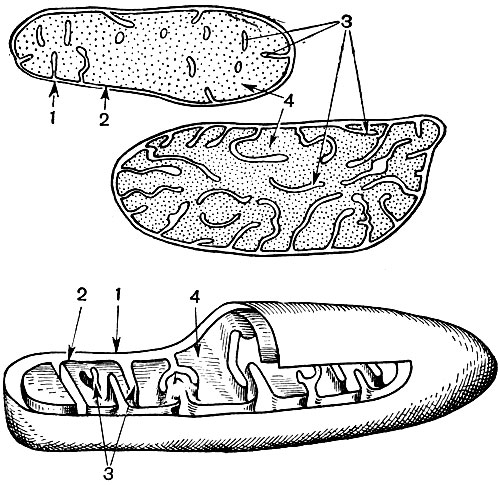
Рис. 31. Строение митохондрии. Вверху и в середине - вид продольного среза через митохондрию (вверху - митохондрия из эмбриональной клетки кончика корня; в середине - из клетки взрослого листа элодеи). Внизу - трехмерная схема, на которой часть митохондрии срезана, что позволяет видеть ее внутреннее строение. 1 - наружная мембрана; 2 - внутренняя мембрана; 3 - кристы; 4 - матрикс
Мы говорим здесь о глюкозе как о веществе, расщепляя которое клетка добывает энергию. Глюкоза является центральным, но не единственным из таких веществ. Молекула ее имеет остов из шести атомов углерода, соединенных между собой. В результате длинной и сложной цепи реакций ее молекула дробится, окисляется и, в конце концов, расщепляется на шесть молекул неорганического вещества - углекислого газа (СO2), каждая молекула которого содержит лишь один атом углерода, причем он предельно окислен. Сложив все последовательные реакции окисления глюкозы и исключив при этом все промежуточные продукты, можно получить суммарную реакцию этого процесса:
Крахмал легко превращается в глюкозу, после чего она подвергается вышеописанному расщеплению. Белки и жиры дают различные органические кислоты, которые превращаются в промежуточные продукты распада глюкозы и далее окисляются таким же образом, как последняя, и с помощью тех же ферментов.
Полное биологическое окисление органического вещества подобно его сгоранию. В обоих случаях результатом являются углекислый газ, вода и выделяющаяся энергия. Однако при горении эта энергия выделяется в виде тепла, причем сразу полностью; при биологическом окислении энергия химических связей освобождается порциями, и основная ее часть связывается, переходя в энергию фосфатной химической связи АТФ. В итоге клетка получает концентрат энергии в такой форме, которая затем в нужный момент и в соответствующей точке может использоваться для создания новых химических связей, для синтеза новых веществ, а также для производства других видов работы - электрической, механической, а также работы по транспорту веществ из среды в клетку, из клетки в среду и от клетки к клетке.
Чем активнее жизнедеятельность клетки, тем больше у нее потребность в энергии и тем больше в ней митохондрий. Они и в пределах одной и той же клетки могут быть распределены неравномерно: их больше в той части клетки, которая в данный момент работает активнее.
Митохондрии способны синтезировать часть тех веществ, из которых состоят они сами. Благодаря этому митохондрии могут размножаться.
Эндоплазматический ретикулум - органоид цитоплазмы, в котором происходит синтез очень многих веществ (табл. 10). Эндоплазматический ретикулум представляет собой систему каналов, которые пронизывают цитоплазму и которые в одних участках сужаются, в других расширяются, образуя то цистерны, то плоские мешки, то ветвящиеся трубки. Стенки всех этих образований построены из мембран, включающих в свой состав ферменты.
Как и в других мембранных образованиях клетки, ферменты в ретикулуме расположены упорядоченно. При этом соседние ферменты осуществляют последовательно протекающие реакции (рабочие операции), а группа их - всю цепь реакций, ведущих к созданию того или иного вещества.
Различают агранулярный (гладкий) и гранулярный эндоплазматический ретикулум. На наружной поверхности каналов гранулярного ретикулума располагаются многочисленные мелкие органоиды - рибосомы, функцией которых является синтез белковых молекул. Агранулярный эндоплазматический ретикулум, который в растительной клетке количественно преобладает над гранулярным, не несет рибосом.
Эндоплазматический ретикулум, кроме того, что он является конвейером для многих видов ферментативного превращения веществ, главным образом для их синтеза, представляет собой и систему магистралей, по которым вещества перемещаются по клетке. Начинается ретикулум от наружной мембраны оболочки ядра и, ветвясь, подходит к различным органоидам цитоплазмы, а также к плазмалемме. Тем самым он связывает между собой все части клетки. Кроме того, каналы эндоплазматического ретикулума проходят через плазмодесмы, соединяя ретикулум соседних клеток.
Далее, мембраны эндоплазматического ретикулума расчленяют цитоплазму на многочисленные отсеки, благодаря чему клетку нельзя представить как однородный массив, в котором перемешаны самые различные вещества. Условия в одном отсеке могут быть совершенно иными, чем в любом другом; процессы, в нем протекающие, могут идти только в нем, тогда как в каждом другом происходят иные процессы.
Наконец, мембраны эндоплазматического ретикулума - это те поверхности, по которым распространяются биотоки, являющиеся сигналами, меняющими избирательную проницаемость мембран и тем самым активность ферментов. Благодаря этому одни химические реакции пускаются в ход, другие тормозятся - обмен веществ подчиняется регуляции и протекает координированно.
Многие из веществ, синтезированных в клетке, должны быть сконцентрированы и выделены из клетки либо в наружную среду, либо во внутриклеточную вакуоль. Кроме того, клетка концентрирует и вещества, поступающие в нее из других клеток, например если она откладывает их про запас. Эту работу выполняют диктиосомы. Обычно в растительной клетке имеется несколько диктиосом, и вся их совокупность называется аппаратом (или комплексом) Гольдж и данной клетки. Каждая диктиосома представляет собой систему мембран, сложенных стопкой (табл. 10 - срез, рис. 32 - объемная схема). Полости между мембранами, образующими этот органоид, имеют вид то узких щелей, то плоских мешочков - цистерн, то пузырьков. Форма их меняется в ходе работы органоида и, по-видимому, зависит от степени наполнения межмембранных пространств выделяемыми и накапливаемыми веществами. Сформировавшиеся и разросшиеся пузырьки отделяются от органоида. По-видимому, многие клеточные вакуоли, окруженные мембранами - тонопластом, являются продуктом деятельности аппарата Гольджи; это оторвавшиеся от него и затем увеличившиеся пузырьки.
Аппарат Гольджи особенно развит в выделительных (секреторных) клетках, в которых откладываются или из которых выводятся различные вещества. Он синтезирует и выделяет вещества, образующие клеточную оболочку.
Лизосомы - довольно мелкие (около 0,5 мк в диаметре) округлые тельца - еще один органоид цитоплазмы. Они покрыты оболочкой - липопротеиновой мембраной. Содержимое лизосом - ферменты, переваривающие белки, углеводы, нуклеиновые кислоты и липиды. Оболочка лизосомы препятствует выходу ферментов из органоида в гиалоплазму, в противном случае последняя переваривалась бы этими ферментами.
Можно думать, что лизосомы являются продуктами деятельности аппарата Гольджи, оторвавшимися от него пузырьками, в которых этот органоид аккумулировал переваривающие ферменты.
Те части клетки, которые отмирают в процессе ее развития, разрушаются с помощью ферментов лизосом. В умершей клетке лизосомы разрушаются, ферменты оказываются в цитоплазме, и вся клетка, за исключением оболочки, подвергается перевариванию.
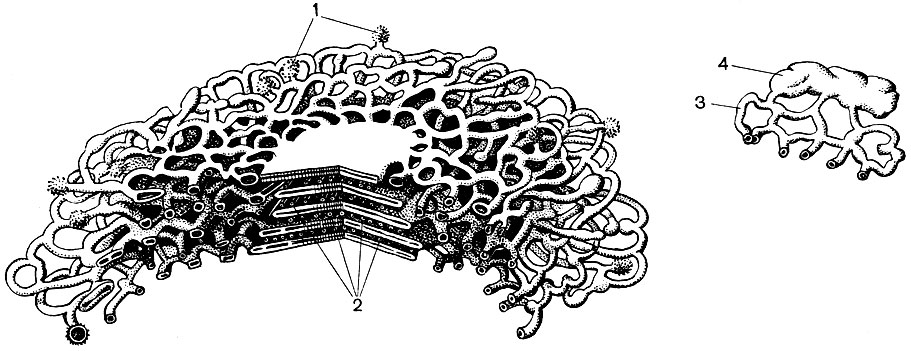
Рис. 32. Трехмерное схематическое изображение строения части диктиосомы из растительной клетки. Слева показана часть пяти смежных цистерн. Справа в более увеличенном виде представлено образование секретируемого аппаратом Гольджи пузырька, еще прикрепленного к каналам - разветвлениям цистерн. 1 - пузырьки; 2 - цистерны; 3 - каналы; 4 - развивающиеся пузырьки
Рибосомы - очень мелкие органоиды, диаметр их около 250А. По форме они почти шаровидны. Часть их прикреплена к наружным (гиалоплазматическим) поверхностям мембран, образующих каналы гранулярного эндоплазматического ретикулума; часть же находится - в свободном состоянии в гиалоплазме. В клетке может содержаться до 5 млн. рибосом. Они представляют собой "аппараты для синтеза белка. Поэтому особенно много их в клетках, активно образующих белок, - в растущих клетках, в клетках, секретирующих белковые вещества. Рибосомы имеются также в митохондриях и хлоропластах, где они синтезируют часть белков, из которых построены эти органоиды.
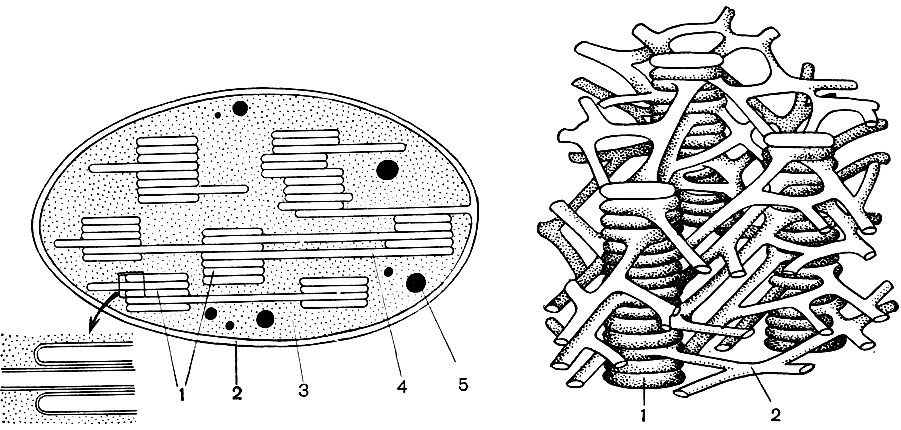
Рис. 33. Строение хлоропласта. Слева - продольный разрез через хлоропласт. Участок внизу показан в увеличенном виде: 1 - граны, образованные ламеллами, сложенными стопками; 2 - оболочка; 3 - строма (матрикс); 4 - ламеллы; 5 - капли жира образованныен в хлоропласте. Справа - трехмерная схема расположения и взаимосвязи ламелл и гран внутри хлоропласта: 1 - граны; 2 - ламеллы.
Во многих клетках обнаружены органоиды, названные микротрубочками. Само их название говорит об их форме - это трубочки с каналом внутри. Внешний их диаметр порядка 250А. Иногда это двойные трубочки - две одиночные, лежащие бок о бок друг с другом и имеющие общую стенку, которая разделяет их полости. Стенки микротрубочек построены из белковых молекул. Считают, что микротрубочки связаны с сократительной (двигательной) активностью цитоплазмы и ее образований. Из них, как из строительных деталей, построены, по-видимому, сократительные структуры жгутика - органоида, при помощи которого перемещаются некоторые одноклеточные и колониальные водоросли, а также клетки, служащие для размножения многих низших растений. Из микротрубочек во время деления клетки образуются нити веретена, о котором речь будет идти дальше. В период деления микротрубочки собираются в группы и образуют эти нити. По окончании деления нити вновь распадаются на отдельные микротрубочки. В клетках или их частях, которые лишены плотной оболочки, микротрубочки, возможно, выполняют опорную функцию, составляя внутренний скелет клетки.
Пластиды - органоиды, присущие только растительным клеткам. Обычно это крупные тельца, хорошо видимые под световым микроскопом.
Различают 3 типа пластид: бесцветные - лейкопласты, зеленые - хлоропласты, окрашенные в другие цвета - хромопласты. Пластиды каждого типа имеют свое строение и несут свои, им присущие функции. Однако возможны переходы пластид из одного типа в другой. Так, позеленение клубней картофеля вызывается перестройкой их лейкопластов в хлоропласты. В корнеплоде моркови лейкопласты переходят в хромопласты. Пластиды всех трех типов образуются из пропластид.
Пропластиды - бесцветные тельца, похожие на митохондрии, но несколько крупнее их. В больших количествах они встречаются в меристематических клетках. Лейкопласты находятся в клетках неокрашенных частей растений (плодов, семян, корней, эпидермиса листьев). Форма их неопределенна. Чаще всего встречаются лейкопласты, в которых откладывается крахмал (он образуется из Сахаров). Есть лейкопласты, запасающие белки. Наименее распространены лейкопласты, заполненные жиром; они образуются при старении хлоропластов. Существенных различий между лейкопластами и пропластидами нет.
Хлоропласты - пластиды высших растений, в которых идет процесс фотосинтеза, т. е. использование энергии световых лучей для образования из неорганических веществ (углекислого газа и воды) органических веществ с одновременным выделением в атмосферу кислорода Хлоропласты имеют форму двояковыпуклой линзы, размер их около 4 - 6 мк. Находятся они в наренхимных клетках листьев и других зеленых частей высших растений. Число их в клетке варьирует в пределах 25 - 30.
О строении хлоропласта дают представление таблицы 11 и 12 (вид на срезе) и схеме на рисунке 33. Снаружи хлоропласт покрыт оболочкой состоящей из двух липопротеиновых мембран. Под ней, в основном веществе (строме), упорядоченно расположены многочисленные образования - ламеллы. Они образуют плоские мешочки, которые лежат друг на друге правильными стоиками. Эти стоики, напоминающие монеты, сложенные столбиком, называются гранами. Сквозь них проходят более длинные ламеллы, так что все граны хлоропласта связаны в единую систему. В состав мембран образующих граны, входит зеленый пигмент - хлорофилл. Именно здесь происходят световые реакции фотосинтеза - поглощение хлорофиллом световых лучей и превращение энергии света в энергию возбужденных электронов. Электроны, возбужденные светом, т. е. обладающие избыточной энергией отдают энергию на разложение воды и синтез АТФ. При разложении воды образуются кислород и водород. Кислород выделяется в атмосферу, а водород связывается белком ферредоксином. Ферредоксин затем вновь окисляется, отдавая этот водород веществу-восстановителю, сокращенно обозначаемому НАДФ. НАДФ переходит в восстановленную форму - НАДФ-Н2. Таким образом, итогом световых реакций фотосинтеза является образование АТФ, НАДФ-Н2 и кислорода, причем потребляются вода и энергия света.
В АТФ аккумулируется много энергии - она затем используется для синтезов, а также для других нужд клетки. НАДФ-Н2 - аккумулятор водорода, причем легко его затем отдающий. Следовательно, НАДФ-Н2 является химическим восстановителем. Большое число биосинтезов связано именно с восстановлением, и в качестве поставщика водорода в этих реакциях выступает НАДФ-Н2.
Далее, с помощью ферментов стромы хлоропластов, т. е. вне гран, протекают темновые реакции: водород и энергия, заключенная в АТФ, используются для восстановления атмосферного углекислого газа (С02) и включения его при этом в состав органических веществ. Первое органическое вещество, образующееся в результате фотосинтеза, подвергается большому числу перестроек и дает начало всему многообразию органических веществ, синтезирующихся в растении и составляющих его тело. Ряд из этих превращений происходит тут же, в строме хлоропласта, где имеются ферменты для образования Сахаров, жиров, а также все необходимое для синтеза белка. Сахара могут затем либо перейти из хлоропласта в другие структуры клетки, а оттуда в другие клетки растения, либо образовать крахмал, зерна которого часто можно видеть в хлоропластах. Жиры тоже откладываются в хлоропластах или в виде капель, или в форме более простых веществ, предшественников жиров, выходят из хлоропласта.
Усложнение веществ сопряжено с созданием новых химических связей и обычно требует затрат энергии. Источник ее - все тот же фотосинтез. Дело в том, что значительная доля веществ, образующихся в результате фотосинтеза, вновь распадается в гиалоплазме и митохондриях (в случае полного сгорания - до веществ, которые служат исходным материалом для фотосинтеза, - С02 и Н20). В результате этого процесса, по своей сути обратного фотосинтезу, энергия, ранее аккумулированная в химических связях разлагаемых веществ, освобождается и - снова через посредство АТФ - тратится на образование новых химических связей синтезируемых молекул. Таким образом, существенная часть продукции фотосинтеза нужна только для того, чтобы связать энергию света и, превратив ее в химическую, использовать для синтеза совсем других веществ.
И лишь часть органического вещества, образующегося при фотосинтезе, используется как строительный материал для этих синтезов.
Продукция фотосинтеза (биомасса) колоссальна. За год на земном шаре она составляет около 1010 т. Органические вещества, создаваемые растениями, - это единственный источник жизни не только растений, но и животных, так как последние перерабатывают уже готовые органические вещества, питаясь либо непосредственно растениями, либо другими животными, которые, в свою очередь, питаются растениями. Таким образом, в основе всей современной жизни на Земле лежит фотосинтез. Все превращения веществ и энергии в растениях и животных представляют собой перестройки, перекомбинации и переносы вещества и энергии первичных продуктов фотосинтеза. Фотосинтез важен для всего живого и тем, что одним из его продуктов является свободный кислород, происходящий из молекулы воды и выделяющийся в атмосферу. Полагают, что весь кислород атмосферы образовался благодаря фотосинтезу. Он необходим для дыхания как растениям, так и животным.
Хлоропласты способны перемещаться по клетке. На слабом свету они располагаются под той стенкой клетки, которая обращена к свету. При этом они обращаются к свету своей большей поверхностью. Если свет слишком интенсивен, они поворачиваются к нему ребром и выстраиваются вдоль отенок, параллельных лучам света. При средних освещенностях хлоропласты занимают положение, среднее между двумя крайними. В любом случае достигается один результат: хлоропласты оказываются в наиболее благоприятных для фотосинтеза условиях освещения. Такие перемещения хлоропластов (фототаксис) - это проявление одного из видов раздражимости у растений.
Хлоропласты обладают известной автономией в системе клетки. В них имеются собственные рибосомы и набор веществ, определяющих синтез ряда собственных белков хлоропласта. Имеются также ферменты, работа которых приводит к образованию липидов, входящих в состав ламелл, и хлорофилла. Как мы видели, хлоропласт располагает и автономной системой добывания энергии. Благодаря всему этому хлоропласты способны самостоятельно строить собственные структуры. Существует даже взгляд, что хлоропласты (как и митохондрии) произошли от каких-то низших организмов, поселившихся в растительной клетке и сперва вступивших с нею в симбиоз, а затем ставших ее составной частью, органоидом.
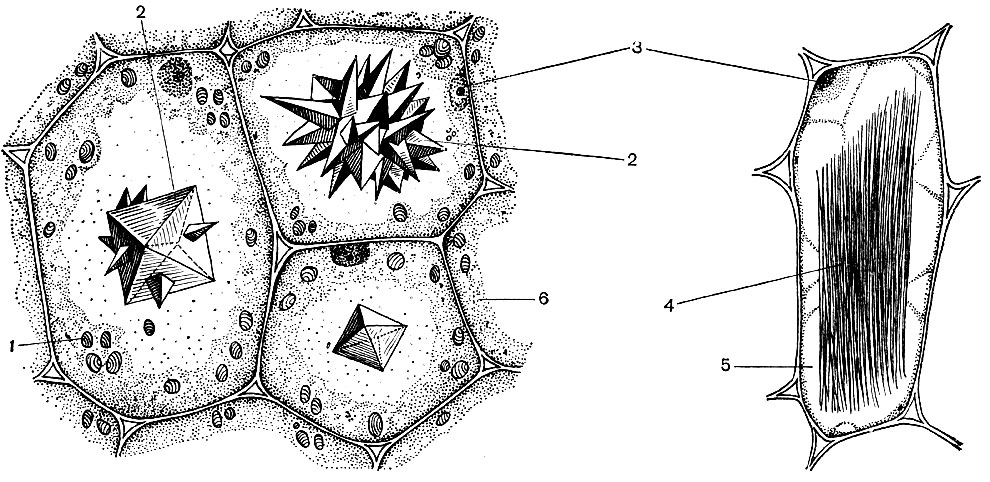
Рис. 34. Кристаллы щавелевокислого кальция в вакуолях клеток. Слева - в клетках из черешка листа бегонии королевской. Справа - в клетке ряски малой. 1 - крахмальные зерна; 2 - друзы; 3 - ядро; 4 - рафиды; 5 - вакуоль; 6 - цитоплазма
У низших растений фотосинтез также осуществляется специализированными, хотя и не столь высокоразвитыми, как в хлоропласте, мембранными структурами. У фотосинтези-рующих бактерий мембраны, содержащие хлорофилл, образуют сеть, которая пронизывает тело бактерии. У сине-зеленых водорослей фотосинтезирующие мембраны слиты в плоские пузырьки. У зеленых и других водорослей система этих мембран отделена от остальной части клетки покрывающей мембраной и образует специальный органоид - хроматофор. Число хроматофоров в клетке невелико, часто клетка содержит всего лишь один хроматофор. Форма их очень различна у водорослей разных видов.
У спирогиры хроматофор имеет вид ленты, спирально вьющейся вдоль стенок клетки; у клостридиума - это ребристые цилиндры; у зигнемы - звездчатые тела.
Хромопласты возникают либо из пропластид, либо из хлоропластов, либо из лейкопластов. Их внутренняя мембранная структура гораздо проще, чем у хлоропластов. Гран нет, строма содержит много желтого или оранжевого пигмента. Хромопласты содержатся в клетках лепестков, плодов, корнеплодов.
В типичной растительной клетке имеется крупная вакуоль, наполненная жидким содержимым. Часто вакуоль занимает почти весь объем клетки, так что цитоплазма составляет лишь тонкий слой, прилегающий к клеточной оболочке. У молодых клеток бывает несколько мелких вакуолей, которые по мере развития клетки разрастаются и сливаются в одну. Содержимое вакуоли - клеточный сок - это водный раствор очень многих веществ: Сахаров, аминокислот, других органических кислот, пигментов (красящих веществ), витаминов, дубильных веществ, алкалоидов, гликозидов, неорганических солей (нитратов, фосфатов, хлоридов), иногда - белков.
Все эти вещества - продукты жизнедеятельности клетки. Одни из них хранятся в вакуолярном (клеточном) соке в качестве запасных веществ и со временем вновь поступают в цитоплазму для использования. Другие являются отбросами обмена веществ, выведенными прочь из цитоплазмы. Так, в вакуоль выводится щавелевая кислота; в вакуолярном соке часто откладываются кристаллы щавелевокислого кальция - иногда в форме одиночных кристаллов, в других случаях в виде конгломерата кристаллов этой соли - многогранных (друзы) или игольчатых (рафиды), - изображенных на рисунке 34.
У подавляющего большинства растений (исключение составляют прокариотические организмы) в каждой живой клетке имеется ядро или несколько ядер. Клетка, лишенная ядра, способна жить лишь короткое время. Безъядерные клетки ситовидных трубок - живые клетки. Но живут они недолго. Во всех других случаях безъядерные клетки являются мертвыми.

Рис. 35. Схема строения фрагмента молекулы ДНК. Каждая из двух цепочек образована благодаря тому, что дезоксирибозы соседних нуклеотидов связаны между собой через фосфат. Цепочки соединены друг с другом благодаря водородным (... Н ...) связям между азотистыми основаниями их нуклеотидов (-А ... Н ...Т- или -Г ... Н ... Ц -)
Ядро всегда лежит в цитоплазме. Форма ядра может быть различной - округлой, овальной, сильно вытянутой, неправильно-многолопастной. В некоторых клетках контуры ядра меняются в ходе его функционирования, причем на его поверхности образуются лопасти различной величины. Размеры ядер неодинаковы и в клетках разных растений, и в разных клетках одного и того же растения. Относительно крупные ядра бывают в молодых, меристематических клетках, в которых они могут занимать до 3/4 объема всей клетки. Относительные, а иногда и абсолютные размеры ядер в развитых клетках значительно меньше, чем в молодых.
Снаружи ядро покрыто оболочкой, состоящей из двух мембран, между которыми имеется щель - околоядерное пространство. Оболочка прерывается порами. Внешняя из двух мембран оболочки дает выросты, непосредственно переходящие в стенки эндоплазматической сети цитоплазмы. И поры и прямая связь эндоплазматической сети с околоядерным пространством обеспечивают тесный контакт между ядром и цитоплазмой.
Содержимое ядра - зернистое основное вещество (ядерный сок, или нуклеоплазма), в котором помещаются более плотные структуры - хромосомы и ядрышко. Ядрышко представляет собой аппарат синтеза материала рибосом и место их сборки из этого материала.
Хромосомы построены из большого числа молекул дезоксирибонуклеиновых кислот (ДНК), соединенных с молекулами белков-гистонов. Молекулы ДНК - это длинные сложно упакованные двойные нити. Каждая молекула состоит из двух нитей, заплетенных спирально одна вокруг другой. Нить, в свою очередь, - это цепочка из огромного числа так называемых нуклеотидов. Нуклеотид - соединение азотистого основания, углевода (дезоксирибозы) и фосфорной кислоты.
В состав каждого из нуклеотидов входит одно из четырех следующих азотистых оснований: аденин, гуанин, цитозин или тимин. Соответственно, в ДНК различают 4 разных нуклеотида: адениновый (А), гуаниновый (Г), цитозиновый (Ц) и тиминовый (Т). Нуклеотиды соединены между собой через свои фосфатные группы, благодаря чему образуется длинная цепочка. Две цепочки, закрученные одна вокруг другой и образующие одну молекулу ДНК, скреплены между собой химическим взаимодействием (так называемые водородные связи) азотистых оснований своих нуклеотидов. Основания образуют пары - одно основание из одной цепочки, другое - из второй. Схема на рисунке 35 дает представление о порядке соединения нуклеотидов в цепочку, а двух цепочек - между собой.
Хотя все молекулы дезоксирибонуклеиновых кислот построены по описанному единому плану, конкретный их качественный состав различен; отличаются они и по величине молекул. Молекула ДНК содержит до 50 - 100 тыс, пар нуклеотидов, но число пар их в каждой молекуле свое, отличающееся от числа пар нуклеотидов в любой другой молекуле. Кроме того, нуклеотидов существует 4 разных вида, и в составе каждой молекулы ДНК свой, ей присущий процент нуклеотидов каждого данного вида. Иначе говоря, количественное отношение А : Г : Ц : Т у каждой молекулы ДНК свое. И наконец, в каждой молекуле ДНК порядок чередования нуклеотидов А, Г, Ц, Т характерен только для данной молекулы. Перестановка местами только двух пар из 50 тыс. пар нуклеотидов, пусть даже соседних, резко меняет свойства всей молекулы; то же самое-замена хотя бы одной пары, скажем Из - Г на А - Т или на Г - Из; то же самое - отсутствие одной пары из числа тех же 50 тыс. В действительности молекулы ДНК могут отличаться между собой не в одном звене, не одной-двумя парами нуклеотидов, а в огромном их числе сразу. Количество возможных перестановок пар нуклеотидов в молекуле ДНК бесконечно, и соответственно бесконечно количество разных молекул, у каждой из которых свои свойства. Последовательность нуклеотидов в молекуле ДНК - это зашифрованная запись состава того или иного белка, свойственного данной клетке. На протяжении каждой молекулы ДНК последовательно умещаются записи состава нескольких белков, а во всех молекулах ДНК всех хромосом ядра - записи состава всех белков клетки, которые в ней могут синтезироваться в течение ее жизни. Суть этих кодированных записей состоит в следующем.
Молекулы каждого белка - это цепочки из последовательно соединенных аминокислот. Существует около 20 разных аминокислот, и характер белка определяется тем, из каких именно аминокислот состоит его молекула, каково их общее количество в молекуле и в какой последовательности они соединены друг с другом. Например, участок ДНК, ответственный за состав определенного белка (каждый такой участок называется геном), - это есть запись конкретной последовательности аминокислот, образующих молекулу данного белка, их общего числа в ней. Каждые 3 последовательных нуклеотида цепочки ДНК обозначают (кодируют) одну аминокислоту соответствующей белковой молекулы. Следующие 3 нуклеотида кодируют следующую аминокислоту и т. д. Количества возможных отличающихся сочетаний по 3 нуклеотида из числа четырех разных видов их с избытком хватает для закодирования 20 аминокислот. Интересно, что одни и те же аминокислоты во всем живом мире кодируются одними й теми же сочетаниями нуклеотидов. В итоге в одном гене его нуклеотидным составом зашифрован аминокислотный состав всех белков, способных синтезироваться в данной клетке.
Клеточные белки являются ферментами. Ферменты определяют течение всех реакций, составляющих суть жизнедеятельности клеток. От наличия тех или иных ферментов зависит образование и превращения всех других веществ клетки, будь то жиры, углеводы, алкалоиды, смолы и т. д. Это, в свою очередь, формирует все свойства клетки, отличающие ее от клеток других растений, в клеточных ядрах которых закодирован состав другого набора ферментов. Вот почему сведения о составе всех белков, которые могут образоваться в клетке, - это информация о всех свойствах клетки и организма. Дальше мы увидим, что эта информация - наследственная, т. е. что она в полном объеме передается от клетки к клетке при их размножении и от материнского растения к дочерним.
Хранится эта информация в хромосомах. Однако реализация этой информации - синтез белков - происходит не в них. Гены хромосом выступают только в роли инициаторов этого синтеза. Когда в клетке возникает потребность в образовании того или иного белка, то ген, в котором зашифрован состав этого белка, активируется. Это значит, что на участке нити ДНК, составляющем, данный ген, образуются молекулы так называемой информационной рибонуклеиновой кислоты (и-РНК).
Строение и состав этих молекул, представляющих собой одиночные цепочки из нуклеотидов, отображает нуклеотидное строение того гена, на котором они образовались. Таким образом происходит копирование информации о составе будущего белка.
Образование молекул и-РНК (на каждом гене их образуется много) означает и размножение информации, как бы отпечатывание многих одинаковых матриц, отображающих строение одного и того же гена и тем самым несущих в нуклеотидной последовательности своих молекул информацию об аминокислотном составе заданного белка. Эти отпечатки гена переходят из ядра в цитоплазму. Здесь происходит расшифровка информации, заключенной в матрицах - молекулах и-РНК, реализация информации, перевод ее с языка нуклеотидной последовательности на язык последовательности аминокислот, т. е. синтез белковых молекул заданного состава.
В расшифровке и синтезе, кроме молекулы и-РНК, участвует большое число молекул различных транспортных рибонуклеиновых кислот (т-РНК), рибосомы и ряд ферментов. Аминокислоты связываются с т-РНК - молекула с молекулой. Каждому из 20 видов аминокислот соответствует своя т-РНК. Так, у молекулы т-РНК имеются химические группы, способные узнавать свою аминокислоту, выбирая именно ее из всех наличных аминокислот. Происходит это с помощью специальных ферментов. Узнав свою аминокислоту, т-РНК вступает с ней в соединение. К началу молекулы и-РНК присоединяется рибосома, которая, продвигаясь по и-РНК, соединяет друг с другом в полипептидную цепочку именно те аминокислоты, порядок которых зашифрован нуклеотидной последовательностью данной и-РНК. Это считывание информации, эта ее расшифровка происходит благодаря специальному механизму, имеющемуся в транспортных РНК, молекулы которых выстраивают друг за другом именно те аминокислоты, которые "обозначены", "записаны" нуклеотидной последовательностью молекулы и-РНК. Рибосома - аппарат для химического связывания аминокислот в молекулу белка.
Итак, общая схема такова. На активированном гене - участке одной из цепочек молекулы ДНК - синтезируются нуклеотидные же цепочки, молекулы и-РНК, состав которых точно отображает состав гена, а значит, несет в себе запись состава белка, кодируемого данным геном. В цитоплазме рибосомы на основе информации, перенесенной с гена молекулами и-РНК, с помощью транспортных РНК соединяют разные аминокислоты в заданной последовательности, в результате чего образуется молекула того белка, состав которого закодирован в этом гене.
Образование молекул и-РНК на активированном гене служит и командой и конкретной программой для синтеза строго определенного белка. Одна и та же молекула и-РНК используется как чертеж для создания многих одинаковых молекул белка. Однако она довольно недолговечна, поэтому для длительно продолжающегося синтеза новых молекул того же белка необходимо образование на одном и том же гене одинаковых, но новых экземпляров молекул и-РНК. С переходом гена в неактивное состояние - а это происходит при исчезновении у клетки потребности в данном белке - он блокируется, перестает образовывать и-РНК, и вскоре синтез этого белка прекращается. В ходе жизни клетки у нее возникают потребности в разных белках. Всякий раз активируются определяющие их гены и образуются молекулы и-РНК соответствующего состава.
Каждая рибосома производит за свою жизнь много молекул разных белков. Она может работать на основе любой и-РНК, и результат, характер созданного ею белка зависит только от состава той и-РНК, в контакте с которой рибосома работала на этот раз.
Таким образом, клеточное ядро выполняет следующие взаимосвязанные функции. В нем хранятся сведения о составе всех белков, способных синтезироваться в данной клетке в течение ее жизни. (Исключение представляют собой некоторые белки митохондрий и хлоропластов. Их состав зашифрован в собственных ДНК этих органоидов, где эти ДНК и находятся. Здесь же, на месте, происходят все этапы расшифровки сведений, заключенных в этих ДНК, включая и сам синтез данных белков с помощью собственных рибосом.)
В ядре хранятся сведения о всех свойствах клетки и организма. Ядро организует синтез каждого из этих белков в нужный момент. При делений клетки, сопровождающемся делением ядра, вся эта информация в полном объеме переходит в каждое из вновь образуемых ядер, в каждую новую клетку. Это возможно благодаря тому, что перед делением весь генный материал хромосом самоудваивается, образуются два одинаковых его экземпляра и по одному из них оказывается в каждом из новых ядер. Ядро каждой клетки содержит полный набор генов, свойственных данному организму. Однако в течение жизни разных специализированных клеток работают далеко не все гены. В одних клетках функционирует одна часть генов, в других - другая, в третьих - третья. Именно поэтому клетки разных тканей одного организма отличаются друг от друга. Значительная часть генов так и остается в пассивном, лишь "хранящем информацию" состоянии от рождения до самой смерти клетки, организма. Кроме того, гены в данной клетке работают не одновременно: одни активны в один период жизни клетки, на таком-то этапе ее развития; другие - на другом и т. д.
Почему одни гены так и остаются неактивными, каким образом включаются и выключаются другие - очень сложная и важная проблема, которая сейчас интенсивно исследуется. Большую роль в блокировании и деблокировании генов играют, по-видимому, белки-гистоны. Они входят в состав хромосом, находясь в соединении с ДНК. Возможно, активация, "раскрепощение" гена происходит тогда, когда молекула гистона отсоединяется от соответствующего участка ДНК, тем самым обнажая его цепочки, позволяя им расплестись и начать функционировать химически. Присоединение гистона ведет к блокировке гена. Однако, чем управляется присоединение и отсоединение гистонов, недостаточно ясно.
Генный материал, как говорилось, находится в хромосомах. Поэтому работа ядра по хранению наследственной информации, по ее удвоению и передаче из клетки в клетку, по организации синтеза различных белков в течение жизни клетки - это прежде всего работа хромосом. Хотя они существуют в течение всей жизни клетки, но в виде четких структур хромосомы различимы в ядре только во время деления клетки. В это время происходит конденсация материала хромосом и хромосомы удается выявить методами микроскопии как индивидуальные, четко очерченные образования.
Хромосомы имеют различную форму. Это либо прямые или изогнутые палочки, либо разнообразные крючки, овальные тельца, шарики (рис. 36). Сильно варьируют они и по размерам. Каждая клетка растений данного вида содержит в своем ядре одинаковый набор (или наборы) хромосом из строго определенного числа разных, но строго определенных хромосом. У всех высших и некоторых низших растений в течение их жизненного цикла чередуются два поколения: с клетками, содержащими в ядрах однократный (гаплоидный, n) набор хромосом, и с клетками, ядра которых имеют двойной (диплоидный, 2 n) набор хромосом. Гаплоидный набор состоит из n разных хромосом, по одной каждого типа. Численная величина строго постоянна для всех гаплоидных клеток всех растений данного вида. Так, для лесной земляники это число составляет 7, для гороха - тоже 7, но для фасоли - 11, для яблони - 17, для земляной груши - 51, для сахарного тростника - 60. Диплоидный набор - это два гаплоидных, сложенных вместе. В нем по две хромосомы каждого типа. Если у гороха в гаплоидном наборе 7 разных хромосом, то в диплоидном 7 разных пар хромосом, причем две хромосомы в пределах каждой пары одинаковы. Легко сосчитать, что для перечисленных растений диплоидный набор (2n) составляет для земляники 14, для гороха - 14, для фасоли - 22, для яблони - 34, для земляной груши - 102, а для сахарного тростника - 120 хромосом.
В жизненном цикле мхов, папоротников чередуются организмы с диплоидными и гаплоидными клетками. Эти организмы у папоротников ведут самостоятельную, изолированную друг от друга жизнь. У мхов диплоидное растение живет на гаплоидном. В диплоидном организме образуются гаплоидные клетки - споры. Каждая 1 из них, прорастая, дает гаплоидный организм, т. е. организм, построенный из гаплоидных клеток. В нем образуются гаплоидные же половые клетки - гаметы.
В момент оплодотворения мужская и женская гаметы сливаются, образуя одну клетку - зиготу, имеющую одно ядро - результат слияния ядер обеих гамет. От каждой из гамет ядро зиготы получает по гаплоидному набору хромосом, и в результате оно имеет двойной, диплоидный набор их. Из зиготы развивается организм, каждая клетка которого имеет диплоидный набор хромосом.

Рис. 36. Диплоидные наборы хромосом в клетках скерды зеленой (Crepis capillaris) - слева и сложноцветного Haplopappus gracilis - справа во время деления клеток. У Haplopappus легко опознать гомологичные хромосомы (хромосомы одной из пар имеют вид дуг; хромосомы другой пары несут на конце по каплевидному образованию, соединенному перемычкой с остальной частью хромосомы, - это так называемый спутник)
В диплоидном наборе две хромосомы каждой пары одинаковы по форме, внутреннему строению, содержат гены, управляющие появлением однородных признаков (рис. 36). Они называются гомологичными хромосомами. Одна из них происходит из гаплоидного набора отцовской гаметы, другая - материнской. Поэтому у раздельнополых организмов одна из них несет гены, определяющие развитие подведомственных ей признаков по отцовскому типу, вторая - по материнскому.
Гомологичные хромосомы другой пары таким же образом определяют развитие другого ряда признаков, третьей пары-третьего ряда и т. д. Хотя отцовский и материнский организмы относятся к одному виду, но наследственные свойства их не тождественны - у них имеются и индивидуальные отличия. Поэтому некоторые гены одной гомологичной хромосомы не тождественны соответствующим генам второй.
Гаплоидный набор, входящий в состав диплоидного и происходящий из отцовской гаметы, несет отцовскую наследственность с ее индивидуальными чертами, а гаплоидный набор из материнской гаметы - материнскую. Сложное взаимодействие однородных, но не всегда тождественных генов двух гаплоидных наборов, в сумме образующих один диплоидный, определяет, какие признаки проявятся у диплоидного потомства, которое, по существу, является гибридом отца и матери.
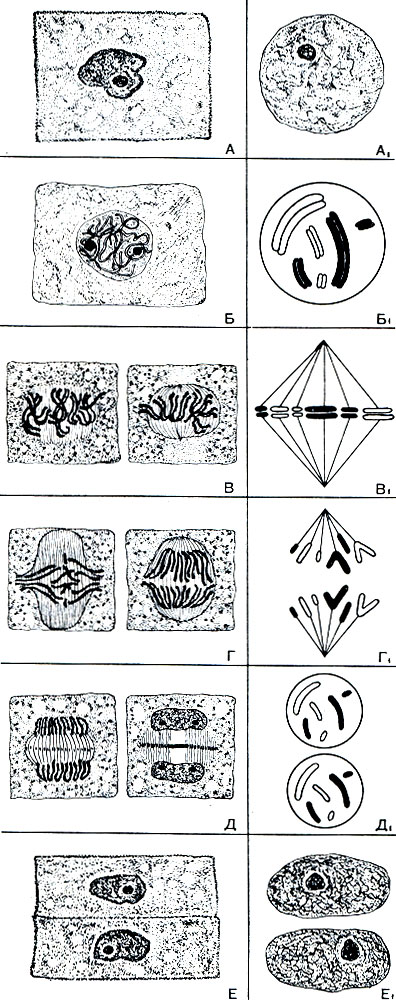
Рис. 37. Митотическое деление клетки кончика корешка лука (слева) и параллельно - схема деления материала ядра при митозе (справ а). На схеме одна из гомологичных хромосом зачернена, другая изображена светлой. А - клетка вне деления, во время интерфазы; хромосомы в это время деконденсированы, деспирализованы, поэтому сами хромосомы не видны как индивидуально обособленные образования; хорошо различимы ядрышки. Б - профаза митоза: клетка готовится к делению; хорошо видны хромосомы, каждая из которых состоит из двух хроматид. В - метафаза митоза: оболочка ядра растворяется; хромосомы располагаются в экваториальной плоскости клетки; появляются нити веретена, прикрепляющиеся к хромосомам. Г - анафаза митоза: хромосомы, расщепившись вдоль, расходятся к противоположным полюсам клетки. Д - тело-фаза: хромосомы деконденсируются, образуются ядра, покрытые каждое своей оболочкой, и появляются ядрышки; исчезают нити веретена, а в середине клетки появляется фраг-мобласт - оболочка, которая, разрастаясь от центра к периферии, перегораживает клетку на две. Е - цитокинез: образование оболочки между двумя дочерними клетками; ядра принимают интерфазный вид - как на А
У голосеменных и покрытосеменных растений чередование поколений происходит в принципе, так же как у мхов и папоротников, но их гаплоидная фаза сильно редуцирована и представлена часто лишь группой клеток. Она живет не самостоятельно, а в теле гаплоидного растения. У покрытосеменных растений женское гаплоидное поколение заключено в зародышевом мешке, находящемся в семяпочке, а мужское- внутри пыльцевого зерна.
Размножаются клетки делением. При этом из одной клетки образуются две дочерние, каждая из которых в свое время тоже может поделиться и т. д. Каждая из дочерних клеток должна нести в своем ядре полный и одинаковый объем наследственного вещества, точно такого же, какой содержится в ядре материнской клетки. Только при этом условии наследственные свойства могут полностью передаваться от клетки к клетке и от растения к растениям-потомкам. Специальный механизм-митотическое деление ядра (митоз)- обеспечивает равное и полное распределение наследственного вещества, вещества хромосом, между дочерними клетками (рис. 37).
Еще до деления клетки каждая молекула ДНК в каждой хромосоме пристраивает около себя свою копию - вторую такую же молекулу. В результате весь наследственный материал клетки удваивается, а каждая хромосома состоит теперь из двух равноценных частей - хроматид. Далее "задача" клетки состоит в том, чтобы, разделив каждую хромосому на хроматиды, строго поровну распределить их между будущими дочерними клетками: в каждую из них нужно направить по одной хроматиде от каждой хромосомы. Это осуществляется следующим образом. Непосредственно перед делением клетки хромосомы сильно уплотняются и сокращаются. Затем они располагаются в одной плоскости - по экватору ядра, причем одна хроматида каждой из них обращена к одному полюсу клетки, другая - к противоположному. Ядерная оболочка исчезает, растворяется и ядрышко. Между полюсами клетки появляются нити, в своей совокупности образующие фигуру веретена. Нити собираются из соединяющихся друг с другом микротрубочек. Веретено состоит из нитей двух родов. Одни- непрерывные, идущие от одного полюса клетки к другому. Другие - тянущие, каждая из которых соединяет полюс с одной из хроматид. В хромосоме имеется участок - кинетохор, к которому и прикрепляются тянущие нити - одна от одного полюса, другая от второго. Далее происходит расхождение хроматид. Хроматиды, составлявшие до этого каждую хромосому, отделяются друг от друга и, подтягиваемые нитями, расходятся к противоположным полюсам клетки. В результате у каждого полюса собирается по одному полному набору хроматид, которые теперь уже являются хромосомами. После этого нити веретена распадаются, вокруг каждого набора хромосом образуется ядерная оболочка, хромосомы сильно разбухают (деспирализуются). В каждом ядре появляется ядрышко. Ядро обретает структуру, свойственную ядрам, неделящихся клеток. В срединной плоскости клетки образуется перегородка, разделяющая клетку на две дочерние.
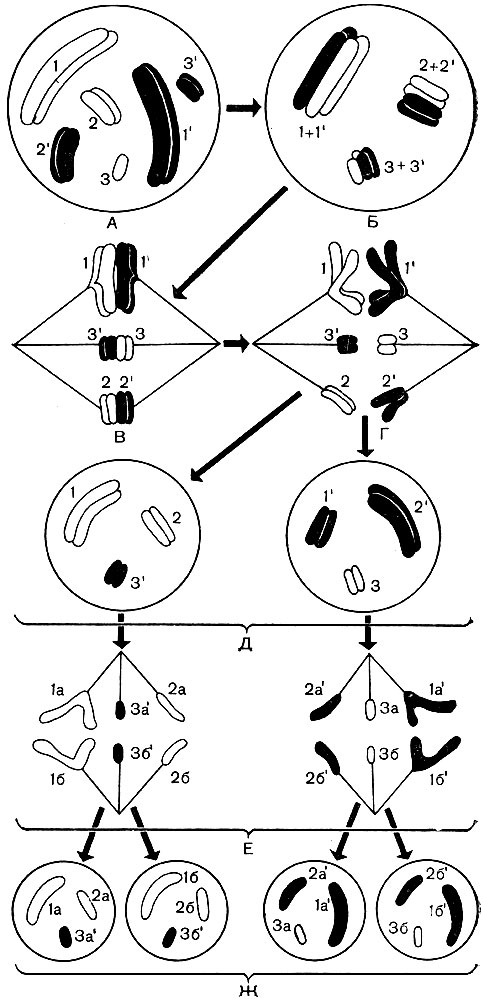
Рис. 38. Схема поведения хромосом 'условной' клетки при мейозе. А - изображено диплоидное ядро клетки, содержащее 3 пары хромосом (1 и 1' - 1-я пара гомологичных хромосом, 2 и 2' - 2-я пара, 3 и 3' - 3-я пара). 2n = 6. 3 хромосомы, происходящие от материнского организма, - светлые (цифры без штриха); парные им гомологичные хромосомы, происходящие от отцовского организма, зачернены (цифры со штрихом). Гомологичные хромосомы обозначены общим номером. Каждая хромосома состоит из двух хроматид. Б - слияние гомологичных хромосом. На этой стадии гомологичные хромосомы обмениваются между собой отдельными участками. В результате происходит некоторое перераспределение материнского и отцовского наследственного материала между хромосомами (кроссинговер). В - образуются нити веретена, прикрепляющиеся к хромосомам, исчезает оболочка ядра. Г - гомологичные хромосомы расходятся к противоположным полюсам клетки, у полюсов оказывается по одной гомологичной хромосоме из каждой пары, общее число хромосом у каждого полюса вдвое меньше, чем в исходном ядре А. Д - образуются два ядра с гаплоидным набором хромосом в каждом; в одно из них попало больше бабушкиных (светлых, материнских по отношению к ядру А) и меньше дедушкиных (зачерненных, отцовских по отношению к ядру А) хромосом, в другое - наоборот; таким образом, новые ядра не вполне тождественны друг другу по составу своего наследственного вещества; их различие обусловлено также и кроссинговером, происходящим с хромосомами на стадии Б. Е - Ж - митотическое деление каждого из гаплоидных ядер Д: продольное расщепление каждой хромосомы, расхождение хромосом к полюсам, образование двух гаплоидных ядер из каждого ядра Д. В итоге появилось 4 клетки с гаплоидными ядрами вместо одной клетки с диплоидным набором хромосом
Органоиды распределяются между дочерними клетками не строго поровну, но затем в клетках синтезируются их составные части, происходит сборка новых экземпляров органоидов каждого вида, и число их в каждой клетке восстанавливается до нормального. Сами клетки растут. В хромосомах происходит удвоение наследственного материала, после чего хромосомы состоят, как перед делением клетки, из двух хроматид. Клетка готова к новому делению.
Рис. 38А. Схема кроссинговера во время мейоза, происходящего на стадии Б рис. 38. 1 - две гомологичные хромосомы, каждая из которых состоит из двух сестринских хроматид; 2 - гомологичные хромосомы сплелись друг с другом; 3 - каждая хромосома расщепляется на составляющие ее хроматиды; хромосомы начинают отталкиваться друг от друга и расходиться, но в некоторых точках {на рисунке - в четырех) они остаются связанными; в центральной петле сдвоены сестринские хроматиды, а в обеих крайних петлях - несестринские; точки, где возможен кроссинговер, показаны стрелками; 4 - хроматиды обменялись участками, и хромосомы, претерпев как бы взаимную гибридизацию, расходятся; 5 - две разошедшиеся, но уже 'гибридные' (в отличие от показанных на 1) хромосомы
При смене диплоидного поколения клеток гаплоидным происходит так называемое редукционное деление ядра - мейоз. Во время мейоза (рис. 38) гомологичные хромосомы каждой пары сближаются, тесно прилегают друг к другу по своей длине, перекручиваются. Между соприкасающимися гомологичными хромосомами происходит обмен отдельными участками. В результате этого часть генов отцовских хромосом переходит в состав материнских хромосом, а соответствующие им гены материнских хромосом занимают освободившиеся места в отцовских хромосомах (явление кроссинговера - рис. 38А). Внешний вид тех и других хромосом в результате этого не меняется, но их качественный состав становится иным. Отцовская и материнская наследственности перераспределяются и смешиваются. Далее ядерная оболочка и ядрышко растворяются, образуется аппарат веретена, такой же, как при митозе. Гомологичные хромосомы разъединяются и с помощью нитей веретена расходятся к полюсам клетки. У одного полюса оказывается один гаплоидный набор хромосом (по одной гомологичной хромосоме из каждой пары), у другого - второй гаплоидный набор.
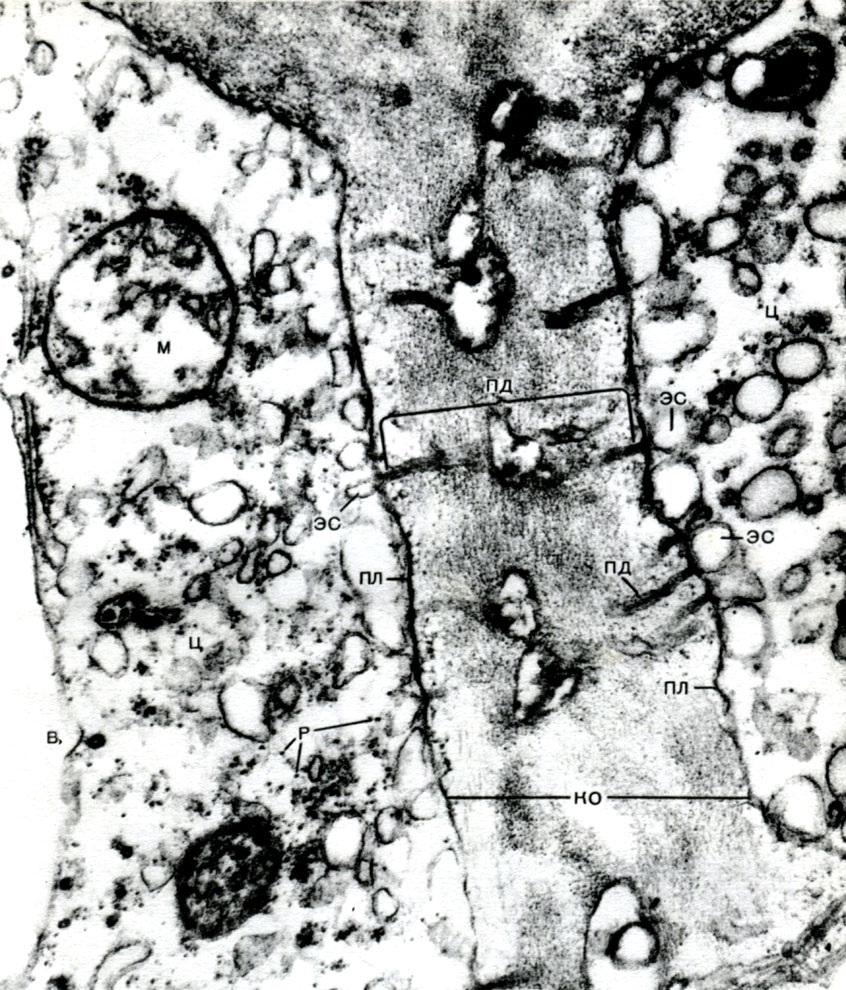
Таблица 9. Участок слившихся оболочек двух смежных клеток и прилегающие к нему участки цитоплазмы этих клеток. Паренхимные клетки коры корня ели (Picea abies). Электронная микрофотография (увел. х 75 000) А. Е. Васильева: ко - клеточные оболочки; пл - плазмалемма; пд - плазмодесмы, некоторые из которых видны не по всей своей длине, так как они тянутся не строго в плоскости среза; ц - цитоплазма; эс - элементы эндоплазматической сети (многие каналы перерезаны поперек); видна связь эндоплазматической сети с плазмодесмами; м - митохондрия; р - рибосомы; в - вакуоль
Вслед за мейозом образовавшиеся гаплоидные ядра делятся путем митоза. При этом каждая хромосома гаплоидного набора расщепляется на две хроматиды, они расходятся и образуются дочерние гаплоидные клетки.
Мейоз отличается от митоза двумя принципиальными моментами. Во-первых, при мейозе происходят слипание каждой пары гомологичных хромосом и обмен между ними частью наследственного материала, чего нет при митозе. Во-вторых, при мейозе к полюсам клетки расходятся гомологичные хромосомы, по одной от каждой пары, а при митозе каждая хромосома расщепляется на хроматиды и к полюсам расходятся хроматиды, по одной от каждой хромосомы. В результате мейоза из диплоидной клетки образуются гаплоидные.
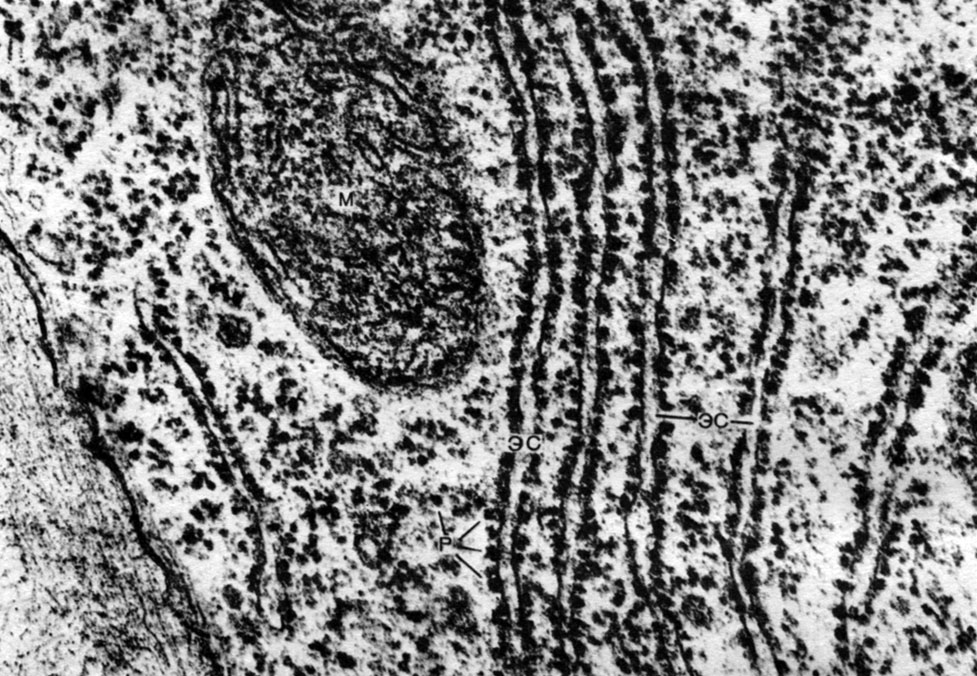
Таблица 10. Структура растительной клетки. Гранулярная эндоплазматическая сеть в цитоплазме развивающегося корневого волоска редиса (Raphanus sativus). Электронная микрофотография (увел. х 103000) М. Ф. Даниловой: эс — каналы эндоплазматической сети; р — рибосомы; м — митохондрия
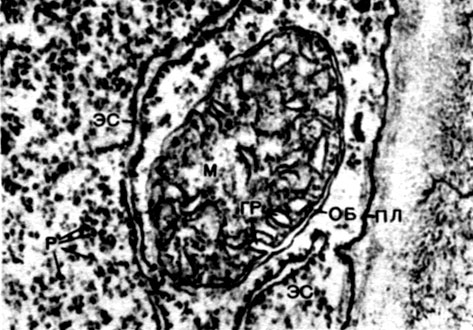
Таблица 10. Структура растительной клетки. Митохондрия в развивающемся корневом волоске редиса (Raphanus sativus). Электронная микрофотография (увел. х 85000) Е. А. Мирославова: м — митохондрия; об — оболочка; гр — гребни; пл — плазмалемма (под ней видна часть оболочки клетки); эс — каналы эндоплазматической сети, на внешних поверхностях которых видны рибосомы; р — свободные рибосомы в цитоплазме
Таблица 10. Структура растительной клетки. Аппарат Гольджи в цитоплазме развивающегося корневого волоска редиса (Raphanus sativus). Электронная микрофотография (увел. х 52000) М. Ф. Даниловой: аг — аппарат Гольджи, видны срезы плоских мешочков и пузырьков
В клетках некоторых тканей при их развитии происходит незавершенный митоз: материал хромосом в ядрах удваивается, хромосомы делятся пополам, но, вместо того чтобы образовать два ядра, остаются в исходном ядре. С этого момента оно заключает в себе не диплоидный, а тетраплоидный (четверной) набор хромосом. Процесс, ведущий к подобному удвоению хромосом внутри одного ядра, называется эндомитозом - внутренним митозом. Если он происходит с одним ядром дважды, то оно становится октоплоидным (восьмикратным) набором и т. д. Клетки, ядра которых несут в себе больше двух наборов хромосом, называются полиплоидными, т. е. многоплоидными. Полиплоидия клеток в ряде случаев повышает их жизнеспособность, поскольку каждый ген дублируется несколькими другими такими же генами. Однокачественные гены действуют в унисон, и повреждение какого-нибудь из них не ведет к выпадению определяемого им признака, так как компенсируется работой остальных однородных генов. Во многих случаях полиплоидные клетки крупнее и богаче содержимым, чем диплоидные. Выведены сорта полиплоидных культурных растений, которые обладают повышенными хозяйственными качествами.
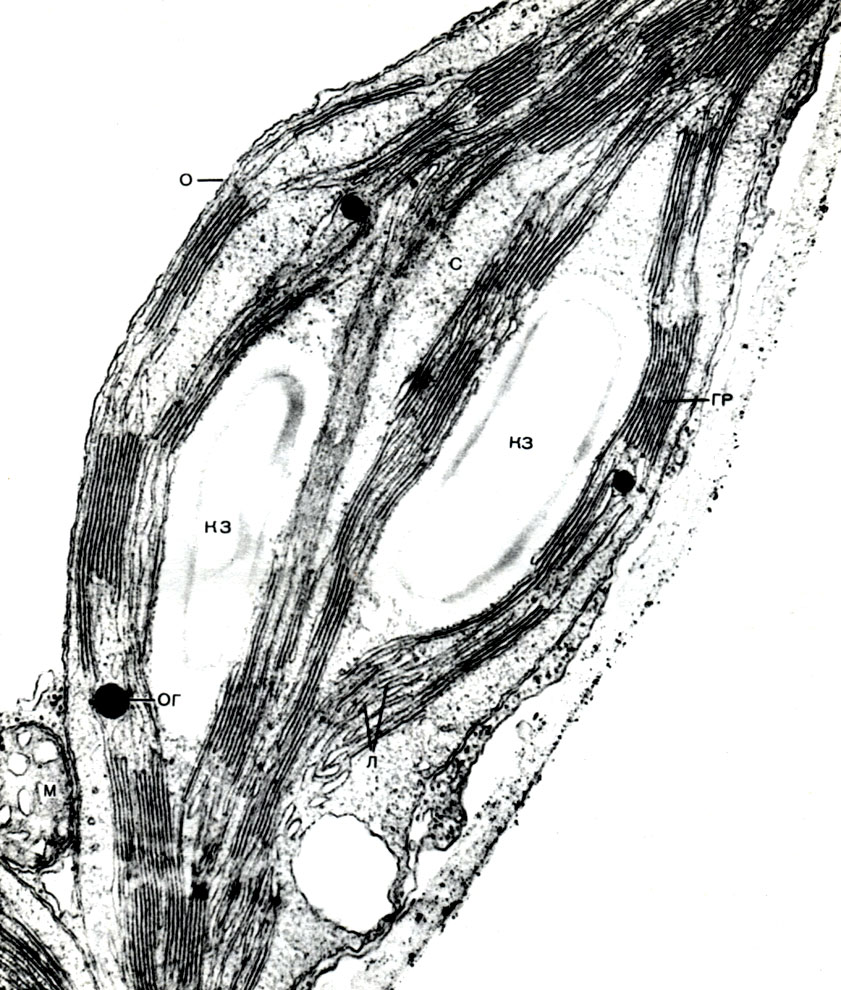
Таблица 11. Хлоропласт из клетки листа огурца (Cucumis sativus). Электронная микрофотография (увел. х 48 000) Е. А. Мирославова: о - оболочка хлоропласта; л - межгранные ламеллы; гр - граны; с - строма; кз - крахмальные зерна; ог - осмиофильные гранулы (капли жироподобных веществ); м - митохондрия
Наука, изучающая клетку, называется цитологией. Клетка представляет собой основу строения всего живого и всех жизненных процессов; поэтому большинство важнейших общебиологических проблем можно решить с привлечением цитологии и методов ее исследования.
С помощью цитологии решаются многие практически важные вопросы: борьба с болезнями растений (грибными, бактериальными), выведение новых сортов культурных растений, преодоление стерильности (бесплодия) гибридных сортов.
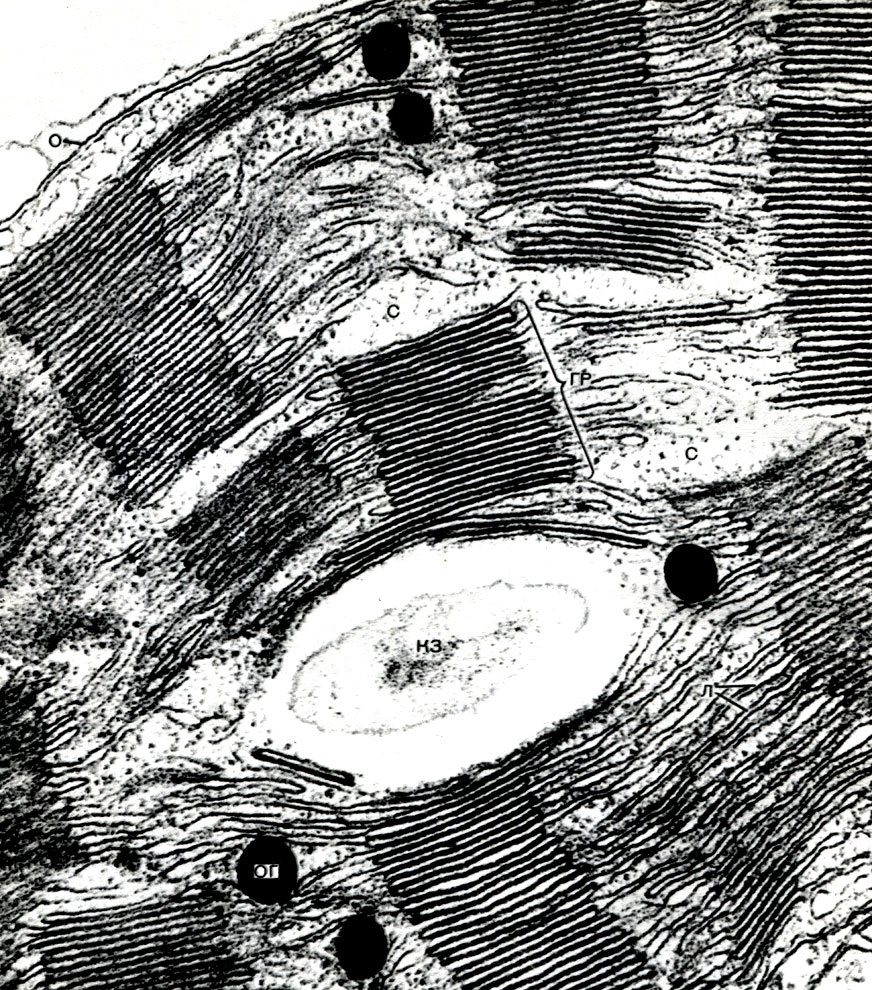
Таблица 12. Фрагмент хлоропласта из паренхимной клетки листа кукурузы (Zea mais). Электронная микрофотография (увел. х 100 000) Е. А. Мирославова: о - оболочка хлоропласта; л - межгранные ламеллы; г - граны; с - строма; кз - крахмальное зерно; ог - осмиофильные гранулы
Эти проблемы современная цитология исследует, применяя разнообразные методы - микроскопические, биохимические, биофизические, генетические - и тесно взаимодействуя с другими смежными биологическими науками.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'