Что такое растение (П. М. Жуковский)
Общая площадь планеты Земля составляет 510 млн. км2. На долю суши приходится 149 млн. км2, Мировой океан занимает 361 млн. км2. И суша и океан заселены растениями и животными. Разнообразие и тех и других очень велико. Ныне установлено около 500 000 видов растений и более 1 000 000 животных.
Среди растений различают разные по окраске и строению водоросли, грибы, слизевики, семенные растения, в том числе голосеменные и цветковые. В подавляющем своем большинстве растения - автотрофные организмы, в основном фототрофные. Но грибы и слизевики гетеротрофны, чужеядны, т е. не сами (как автотрофные растения) строят необходимое для поддержания жизни органическое вещество, а черпают его из окружающей среды. В этом отношении с ними сходны бактерии, тоже относимые обычно к миру растений и в подавляющем большинстве тоже гетеротрофные.
При большом видовом разнообразии трудно найти признаки, общие для всех растений, и притом такие, которые не встречались хотя бы у некоторых животных. Одноклеточные водоросли и некоторые примитивные грибы, сохранившие очень низкий уровень организации, который был свойствен еще, вероятно, древнейшим их предкам, иногда очень сходны с простейшими животными. У ряда растений сходство с животными обнаруживается в строении и поведении специальных клеток размножения и так далее.
Но, отвлекаясь от этих частностей, можно все-таки указать немногие, свойственные подавляющему большинству растительных организмов общие свойства. Среди них прежде всего надо указать специфическое строение клетки - наличие в составе клетки растений твердой, не пропускающей твердых частиц, клеточной оболочки. Эта деталь строения свойственна и клетке гриба, и бактериальной клетке, и клеткам водорослей, и клеткам высших зеленых растений. Клеточная оболочка настолько вошла в программу развития клетки современного высшего растения, что при искусственном удалении целлюлозной оболочки с помощью фермента через несколько часов начинается образование новой клеточной стенки. Подобное явление наблюдается и в тех случаях, когда живое содержимое клетки вследствие нарушения целостности оболочки вытекает наружу - голый протопласт через некоторое время также покрывается оболочкой (водоросль вошерия). Приобретенная еще на очень ранних ступенях исторического развития организмов, оболочка закрепилась, вероятно, как защитное образование, но это сыграло решающую роль в питании клеток. Клетки, покрытые твердой оболочкой, могут воспринимать из окружающей среды необходимое для жизни только в растворенном состоянии.
Приобретение твердой оболочки наложило отпечаток на всю дальнейшую эволюцию растений - питание их стало зависимо от размера поверхности соприкосновения тела растения с окружающей его средой. Вследствие этого строение большинства растений характеризуется более сложным, чем у животных, внешним расчленением. В процессе роста объем тела увеличивается, как известно, пропорционально третьей степени линейного увеличения размера; поверхность - только квадрату этой величины. Поэтому при сохранении геометрического подобия растущего тела величина наружной поверхности, приходящейся на единицу объема, должна уменьшаться, что не может не отразиться на эффективности обмена в сторону ее уменьшения. Это и привело, по всей вероятности, во-первых, к необходимости нарушения в процессе эволюции геометрического подобия за счет расчленения и усложнения наружного контура растения и, во-вторых, к тенденции незавершаемости роста тела растения.
Восприятие питания "всасыванием" было, вероятно, причиной и еще одного свойства растений - их малой подвижности.
Правда, даже некоторые высокоорганизованные растения могут менять свое место в пространстве. Так, корневищные растения, нарастая под землей, со временем развивают надземные побеги и отходящие от них придаточные корни все в новых и новых точках, в то время как первичный побег, возникший некогда из семени, отмирает. Благодаря этому такие растения продолжают свое существование не на том месте, где начиналась их жизнь. Но, во-первых, эти "ростовые" движения несравнимы по скорости и эффекту с теми, которые доступны животным, а во-вторых, они наблюдаются далеко не у всех растений. Есть ограниченные движения и у других растений. Лианы, обвивающие опоры или цепляющиеся за них, иногда перекидывают свои кроны с одного места на другое. Стебель в процессе роста производит так называемые нутационные движения. Листья многих растений из-за неравномерного роста черешков ставят листовые пластинки в наиболее благоприятные условия освещения. Нередки растения, закрывающие цветки на ночь, в то время как у других они закрываются на день. Листочки сложных листьев многих бобовых ежедневно с большой правильностью изменяют свое положение относительно горизонта. Тем не менее не будет ошибкой сказать, что растения в противоположность животным, как правило, "неподвижны". Некоторые долговечные (многие древесные) растения не покидают своего места сотни и даже тысячи лет (секвойи). Это свойство большинства растений - сохранять за собой одно и то же место - относительно общее и, вероятно, тесно связано со способом восприятия питания, а тем самым - с наличием клеточной оболочки.
Дело в том, что, за исключением немногих экологических специализированных групп (галофиты), наземные растения получают минеральное питание из растворов малой концентрации. Извлекаемые из них растениями вещества обычно восполняются в субстрате за счет выветривания и темпами, достаточными, чтобы обеспечить непрерывность питания в данном месте. Кроме того, нарастание и ветвление корней приводят к тому, что их специализированные всасывающие части, оснащенные муфточками корневых волосков, проникают все в новые и новые точки корнедоступного объема субстрата. Быстрая перемена места, сопряженная с тратой энергии, не повысила бы обеспеченности наземного растения ни питательными материалами, ни водой. Что же касается водных растений, то они, окруженные водой, тем более не нуждаются в активном движении - движение и перемешивание воды (ветер, неравномерность нагрева и обусловленная этим конвекция) обеспечивают водные растения всем необходимым. Относительную неподвижность растений, вероятно, можно считать главной причиной особого способа их расселения. Животные обычно расселяются во взрослом, во всяком случае активном, состоянии. В противоположность этому для подавляющей массы растений характерно расселение зачатками.
Зачатки расселения (их называют еще диаспорами) у растений разнообразны. Это могут быть специализированные одиночные клетки, вырабатываемые растением и пассивно или активно отделяющиеся от него, - споры; у других ту же роль выполняют семена - сложно устроенные многоклеточные тела, содержащие в себе зародыш, т. е. маленькое, готовое к росту, но еще не растущее растеньице. У многих растений диаспорами служат специализированные части их вегетативного тела, известные в ряде форм (выводковые почки, метаморфизированные побеги и пр.). В целом разнообразие диаспор очень велико.
Зачатки расселения, в первую очередь споры и семена, во время расселения обычно находятся в состоянии покоя. В это время у них не обнаруживают ни роста, ни других видимых проявлений жизни. Они продолжают оставаться в таком инактивированном (неактивном) состоянии, пока посторонняя растению сила не поставит их в условия, благоприятные для дальнейшего развития. Тогда начинается формирование новой особи растения.
Представляют ли растения филогенетически единую группу - вопрос, по крайней мере, дискуссионный.
В главе "Растения в системе организмов" излагаются аргументы в пользу противоположного воззрения, согласно которому то, что обычно называют растениями, представляет три самостоятельные группы того же ранга, что и группа животных: дробянки (бактерии и сине-зеленые), грибы, растения (фотосинтезирующие).
С этой точки зрения "растения" в обычном понимании надо рассматривать как объединение организмов, обладающих сходными приспособлениями к питанию, т. е. как группу не филогенетическую, а экологическую.

Рис. 1. Участок тропического леса
Растительный покров простирается на огромных площадях континентов земного шара и акватории Мирового океана. Даже в Антарктиде на свободных от льда местах растут (правда, немногие) мхи и лишайники.
Жизнь на Земле в целом обеспечивается воздействием Солнца и атмосферы на зеленый растительный покров суши и океана.
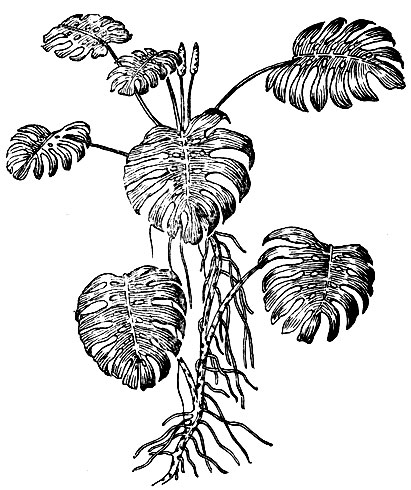
Рис. 2. Монстера
Солнечный свет, распространяясь со скоростью 300 000 км/сек, достигает Земли за 8 мин. Большая часть светового потока, достигшего границы атмосферы, отражается, поглощается и рассеивается ею, и в итоге поверхность суши и морей получает меньше 1/2 (около 48%) энергии, поступающей в атмосферу.
Зеленый экран суши и океанов: лиственные леса тропических и умеренных широт, хвойные леса, луга, степи, полупустыни, пресноводный и морской планктон и бентос Мирового океана, необозримые пространства культурных полевых и садовых насаждений - представляет собой гигантский поглотительный объем для света и реактор фотосинтеза. В дождевых тропических лесах, образованных деревьями разной высоты, кроны их почти полностью смыкаются, а остающиеся небольшие промежутки часто заполнены верхушками богато олиственных лиан; свет почти не проникает под полог леса, и внутри его царят тень и полумрак (рис. 1). Но все же небольшое количество света поступает в виде бликов за счет лучей, многократно отраженных от блестящей поверхности листьев. Да и сомкнутость крон, конечно, не абсолютна - небольшие промежутки между листьями остаются. У некоторых растений дождевого тропического леса листовые пластинки вырезные, например у Monstera deliciosa (неправильно называемой филодендроном); в них образуются как бы окошки, пропускающие солнечный свет (рис. 2).
Высокая степень сомкнутости крон бывает не только в дождевых тропических лесах, но и в горных лесах тропической и субтропической зон, в некоторых вариантах влажного субтропического леса, иногда в таежных лесах. Удивительно не то, что образуются столь сомкнутые насаждения, - в этом проявляется стихия растительной жизни, стремящейся использовать с максимально доступной полнотой весь световой поток, поступающий на Землю. Удивительно, что и в этой глубокой тени все же живут некоторые растения, довольствуясь теми ничтожными порциями света, которые тем или иным путем могут полностью, в виде рассеянного света или бликов пробиваться сквозь зеленую толщу листвы.
Столь полное использование света, как в дождевом тропическом лесу или подобных ему формах лесной растительности, несмотря на присущую растениям способность к неограниченному росту, многократному ветвлению и неограниченному размножению, не всегда возможно. Обычно ограничивающим фактором оказывается недостаточная водообеспеченность, выражающаяся либо в малом количестве осадков, либо в сезонности их поступления. Но и в этих случаях тенденция к использованию светового потока в максимальной степени все же проявляется.
Можно указать в качестве примера хотя бы тропические ксерофильные леса и саванны. В первых обычен густой подлесок при не слишком высокой плотности крон; в саваннах деревья единичны, но зато имеется густой и разнообразный травяной, преимущественно злаковый, покров. Приспособленность растений к наибольшему использованию света можно наблюдать и в растительности умеренных широт. Не случайно, что под пологом древостоя лиственного леса часто наблюдается второй полог из крон кустарников, а под ним травяной покров, в котором иной раз выделяется особый слой совсем приземистых, точно прижатых к почве растений. Свет проходит благодаря такому строению леса как через серию фильтров, поглощающих в совокупности большую часть физиологически активной радиации (рис. 3).
Эту закономерность строения растительного покрова можно наблюдать даже в некоторых чисто травянистых сообществах.

Рис. 3. Участок саванны
Несмотря на обилие солнечного света и большую густоту зеленого покрова, растения используют лишь 1 - 2% падающей на них световой энергии. Но даже при этом в результате фотосинтеза образуются колоссальные количества органической материи.
В процессе фотосинтеза при взаимодействии солнечной энергии, воды и углекислого газа образуется органическое вещество и высвобождается свободный кислород. При этом используется лишь ничтожная часть поступающей в растение воды. Неизмеримо большая доля ее входит в состав цитоплазмы и особенно клеточных вакуолей; вода содержится и в клеточных оболочках. Значительная часть воды заполняет полости сосудов и соответствующих им по функции трахеид, представляя тончайшие водные нити. Они не стабильны - это как бы мельчайшие потоки, постоянно перемещающиеся вверх по растению к листьям. В этом водном потоке перемещаются и растворенные в воде вещества; они, так же как и вода, используются по мере надобности живыми клетками растений.
Вообще содержание воды в растении, хотя и изменяется от вида к виду и в пределах одного вида в зависимости от влажности места обитания, очень велико, составляя, например, в стволах деревьев около половины сырой массы и достигая в листьях некоторых растений (салата, капусты) 90-95%; того же примерно уровня содержание воды в мякоти сочных плодов (томат, арбуз); у водорослей количество воды составляет 98% их сырой массы.
Содержащаяся в растении вода постоянно, хотя бы отчасти, обновляется благодаря, с одной стороны, испарению ее листьями (у растений, погруженных в воду, естественно, испарение отсутствует), с другой - поглощению новых порций из субстрата.
В целом же вода, находящаяся в растении, является той внутренней средой растительного организма, в которой совершаются все химические и физические процессы, в том числе и реакции фотосинтеза. Все эти явления очень сложны, и немалое значение в них принадлежит строению молекул воды и формам связи между ними.
Возвращаясь к фотосинтезу - одному из замечательнейших явлений природы, обратим внимание на то, что необходимый участник этого великого процесса - углекислый газ - содержится в окружающей растение среде в малом количестве. В составе атмосферы на его долю приходится в среднем лишь 0,0321%. Тем удивительнее способность зеленых (в широком смысле) растений (имеются в виду все растения, содержащие хлорофилл, включая и те, зеленая хлорофильная окраска которых маскируется красящими веществами иной цветности - красными, бурыми и др.) создавать огромные количества органических веществ. Достаточно указать, что зеленый покров Земли ежегодно создает сотни миллиардов (4,5 X 1011) тонн органической материи.
Проникая внутрь зеленых растений через устьица на листьях и стеблях, углекислый газ растворяется в воде растительных тканей и при воздействии солнечной энергии и хлорофилла (об этом далее) вступает в сложные реакции. Происходит одно из замечательных явлений в природе - фотосинтез. Фотосинтез - это окислительно-восстановительный процесс. Вода разлагается (окисляется) с выделением кислорода и отдачей водорода на восстановление углекислоты.
В атмосфере есть два источника свободного кислорода: диссоциация молекул водяного пара под воздействием коротких ультрафиолетовых лучей и фотосинтез. Первый процесс был, вероятно, единственным источником кислорода до появления на Земле хлорофиллсодержащих растений; фотосинтез, возникший с появлением зеленых растений, стал основной причиной массового накопления в атмосфере кислорода, содержание которого составляет в настоящее время около 21 % (по объему) и постоянно поддерживается на этом уровне практически. Почти весь кислород современной атмосферы биогенного происхождения.
В нижних частях стратосферы под влиянием коротких ультрафиолетовых лучей (менее 2000А) молекулы кислорода распадаются на атомы, в связи с чем становится возможным образование молекул озона (03). Более длинные ультрафиолетовые лучи, поглощаемые озоном, разрушают молекулы последнего. В зависимости от поглощения энергии кислородом и озоном между ними устанавливается то или иное количественное соотношение. Область максимальной концентрации озона лежит на высотах 25 - 30 км. Здесь находится как бы экран из озона, который поглощает все ультрафиолетовые лучи с длиной волны от 3000А и менее, губительные для жизни. Но этим значение озона не ограничивается, так как он поглощает не только ультрафиолетовые, но и часть инфракрасных лучей, уменьшая этим потерю тепла Землей. Возвращаясь к углекислому газу, напомним, что подавляющее большинство живых организмов, в том числе и растений, выделяет углекислоту в процессе дыхания. Особенно много углекислоты выделяется из почвы благодаря присутствию в ней корневых систем, почвенных водорослей, бактерий и животных. Много углекислого газа выделяется при сжигании топлива, от промышленных процессов, извержений вулканов, лесных пожаров и др.
Растения дышат и днем и ночью, но фотосинтез совершается только в дневные часы. Огромна фотосинтетическая деятельность водорослей, как микроскопических, плавающих взвешенно в толще воды океанов, морей и озер, так и прикрепленных к прибрежным местам крупных водорослей, и даже морских травянистых цветковых растений, прикрепленных ко дну корневищами.
Интенсивность света в воде быстро убывает с глубиной. В озерах на глубине 10 м остается примерно 1%, а в океане на глубине 100 м - около 10% интенсивности света, падающего на поверхность воды. Наиболее продуктивный фотосинтез происходит не в поверхностном фитопланктоне, а на некоторой глубине (около 20 - 25 м). В период полярных ночей фотосинтетическая деятельность растительного планктона почти прекращается, хотя под ледяным панцирем и при очень скудном проникновении света водоросли также жизнедеятельны.
Все организмы, способные к фотосинтезу, содержат хлорофилл (немногие бактерии, способные к фотосинтезу, имеют пигменты, отличающиеся от хлорофилла). Это зеленый пигмент, содержащийся в особых органоидах растительной клетки - хлоропластах. Они взвешены в цитоплазме клетки в виде мельчайших зеленых гранул и обладают способностью движения в зависимости от освещения. В зеленых листьях высших растений хлоропласты представляют собой мелкие белково-липоидные тельца. В клетке их содержится от 20 до 100. Общее количество хлоропластов во взрослом дереве достигает десятков и сотен миллиардов, но у низших одноклеточных водорослей бывает несколько или даже одна зеленая пластида. Величина зеленой пластиды у высших растений обычно 3 - 5 мк.
Зеленые пластиды чутко реагируют на освещение, и хлорофилл образуется в них только на свету. Молекулы хлорофилла поглощают свет. Энергия его преобразуется в процессе синтеза органического вещества в энергию химической связи органических (углеродистых) соединений.
Помимо хлорофилла, пластидам свойственны и другие пигменты, обладающие способностью поглощать лучистую энергию. Вообще все растения, способные к фотосинтезу, содержат, кроме хлорофилла, группу желтых пигментов - каротиноидов (это каротины, ксантофиллы,в том числе фукоксантин бурых и некоторых других водорослей).
Каротиноиды поглощают коротковолновые лучи спектра (синие, фиолетовые и ультрафиолетовые) и передают поглощенную энергию хлорофиллу. У некоторых растений (сине-зеленые и красные водоросли) имеются дополнительные пигменты иной химической природы, чем каротиноиды, но также поглощающие свет и передающие поглощенную энергию хлорофиллу. Это так называемые фикобилины (синий фикоциан и красный фикоэритрин).
Цветковые растения не всегда зеленые, иногда листья у них бывают красные, лиловатые и т. п. Такая окраска вызвана пигментами клеточного сока-антоциана ми, которые маскируют зеленый цвет хлорофилла. Зеленые листья часто располагаются перпендикулярно лучам солнца или под углом к свету. Положение поверхности листьев относительно направленности светового потока в известной степени определяет интенсивность освещения. Иногда у некоторых растений (например, у эвкалиптов) взрослые листья пассивно висят на черешках. Они перпендикулярны лучам только при восходе и закате солнца.
Существенным продуктом фотосинтеза в зеленом листе являются фосфорные эфиры Сахаров (гексозо-фосфаты), которые, освобождая фосфорную кислоту, могут конденсироваться в дисахарид - сахарозу или в полисахарид - крахмал. В некоторых случаях гексозо-фосфаты дефосфорилируются, образуя свободные моносахариды - чаще всего глюкозу и фруктозу. Только первые фазы фотосинтеза световые, остальные не требуют света и являются темновыми реакциями. Первичные продукты фотосинтеза вовлекаются во внутриклеточные химические процессы и служат исходным материалом для построения других, часто очень сложных органических соединений. Результатом фотосинтеза является также образование нуклеиновых кислот, белков и других сложных веществ.
Такова в самом простом изложении схема фотосинтеза. Но наряду с синтезом органических веществ из неорганических элементов в биосфере совершается распад значительной части органических комплексов до исходных неорганических. Этот распад происходит через дыхание, а после гибели зеленых растений и животных - разложение осуществляется благодаря деятельности гетеротрофных растений (бактерий и грибов), населяющих почвы и водные бассейны, обитающих также на трупах животных и т. п.
Процесс разложения органического вещества сложен и имеет многостепенный характер. Но он всегда имеет, по крайней мере в основном, биологическую природу. Продукты разрушения, возникающие под воздействием одних организмов, "подхватываются" другими, а образуемые ими вещества служат объектом биологической переработки третьих, и так далее. Интересно, что закономерная последовательность участия разных организмов в процессе разрушения органической массы обнаруживается даже в том случае, когда разрушители принадлежат к одной группе организмов: такие явления установлены, например, относительно грибов, разрушающих опад в лесах.
Разложение органического вещества в конечном счете приводит к одному и тому же итогу. Углерод в составе углекислого газа поступает в окружающую среду. При разложении образуется также вода. Таким образом, оба исходных для фотосинтеза вещества вновь возвращаются в окружающую среду. В сложных органических соединениях, создаваемых растениями в процессе их жизнедеятельности, равно как и в веществах животного происхождения, содержатся в том или ином количестве минеральные элементы. Усваиваемые растениями из почвы, эти элементы возвращаются в почву и становятся вновь доступными для использования новыми поколениями растений. Следовательно, почти все вещества, вовлекаемые растениями в жизненные процессы, совершают циклическое движение. Это - биологический круговорот веществ. Очень важно осознать, что запасы минеральных веществ в почве не бесконечны. Поэтому без разложения органического вещества и возвращения в землю элементов минерального питания с течением веков и тысячелетий корнедоступная часть толщи земной коры должна бы стать совершенно бесплодной. Биологический круговорот веществ, обусловленный совокупной деятельностью автотрофных и гетеротрофных организмов, является, как следует из сказанного, необходимым условием постоянства жизни, непрерывности ее существования на нашей планете.
В биологическом круговороте веществ особое место принадлежит азоту. Этот элемент входит в белки, т. е. в материальный субстрат жизни, и поэтому совершенно необходим для всех живых существ. При разложении органических веществ азот уходит в атмосферу либо в виде газообразных соединений, либо в свободном состоянии. Возвращение азота в почву, откуда его черпают растения, осуществляется сложным путем, в результате процесса, в котором участвуют специальные бактерии и некоторые другие организмы, связывающие газообразный азот атмосферы в соединения, доступные для потребления растениями. (Подробнее об этом см. на странице 361 этого тома.)
Надо заметить, что при некоторых условиях процессы разложения органического вещества замедляются, вследствие чего образуются скопления полуразложившихся растительных остатков. Такое происхождение имеют торф, бурый и каменный угли. В этом случае нередко сохранность некогда живших растений бывает достаточной, чтобы составить представление о внешнем облике, а нередко и их внутреннем строении. Залежи торфа и углей представляют исключительный экономический интерес. Эти органогенные породы составляют предмет энергичных поисков, а разведанные их запасы - объекты интенсивной эксплуатации. По мнению ряда ученых, нефть также имеет органогенное происхождение.
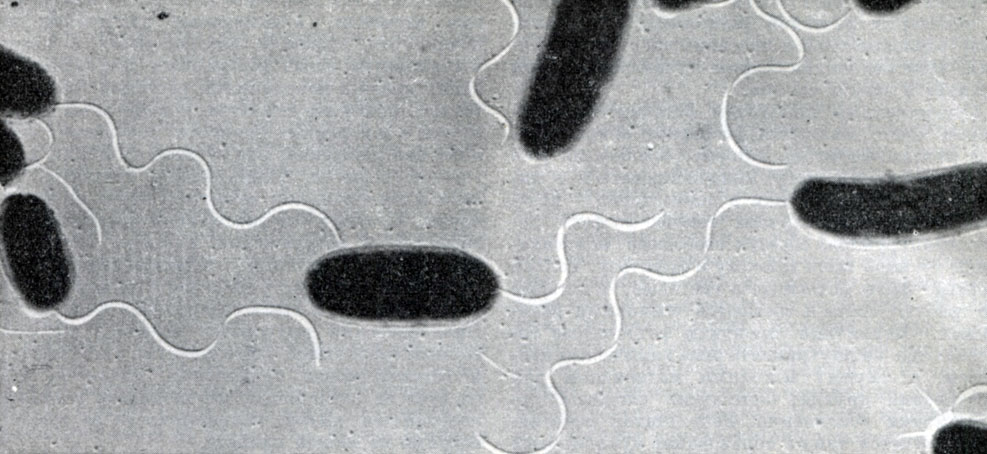
Рис. 4. Представители прокариотов - палочковидные бактерии
Поскольку бесхлорофильные растения способны довести мертвую растительную массу до полного разложения, постольку весь растительный мир в целом, т. е. в совокупности его автотрофных и гетеротрофных компонентов, можно рассматривать как принципиально автономную природную систему, внутренне противоречивую (созидающие органическое вещество и разрушающие его растения), саму себя поддерживающую, т. е. с биогеохимической точки зрения единую.
Развивавшийся в процессе эволюции мир животных, существующий исключительно на гетеротрофной основе, включился в использование как непосредственных продуктов фотосинтеза, так и более сложных органических веществ, создаваемых растениями. Один из результатов этого - удлинение времени существования органических продуктов и увеличение их разнообразия, так как животные не только потребляют, но и преобразуют поглощаемую ими органическую пищу в более сложные вещества.
Важность этого определяется прежде всего тем, что некогда "уловленная" зеленым растением солнечная энергия, заключенная в потенциальной форме в органическом веществе, осуществляет большую работу, выполняемую организмами. Она тем продуктивнее, чем длительнее цикл превращений, испытываемый органическим веществом, созданным зелеными растениями.
Характеристика растений в целом, нарисованная выше, по необходимости схематична. Существующие ныне и ранее жившие на Земле растения очень разнообразны, и более конкретные представления о мире растений можно получить, рассматривая их по группам, объединяющим растения, более сходные между собой, чем с теми, которые относят к другим группам того же ранга. Всякую классификационную группу называют таксоном. Наиболее крупные из них именуются отделами (или типами). Каждый отдел обычно представляет разнообразное по составу объединение растений разного ранга. В него входят классы, подразделяемые далее на порядки, в числе последних различают семейства, которые далее делят на роды. Род включает большее или меньшее количество видов. Вид - основная единица классификации растений, хотя в ряде случаев и она подразделяется на таксоны еще более низкого ранга.
В современной классификации каждый отдел рассматривается как категория, единая в эволюционном отношении, происшедшая от одного общего предка. Все разнообразие видов, родов и других таксонов, относимых в данный отдел, - результат длительной эволюции, осуществлявшейся как процесс приспособления к различным условиям жизни и совместному с другими растениями использованию среды.
В этом издании принято деление растительного мира на отделы, каждый из которых с должной подробностью рассматривается в этом и последующих томах "Жизни растений".
Отделы, в свою очередь, можно подразделить на две категории по организации вегетативного тела растений. У одних из них (у подавляющей части представителей) тело расчленено на лист, стебли и обычно корни. Это так называемые высшие растения. Они почти все автотрофны и связаны в своем существовании с вне-водной (сухопутной) средой. Лишь небольшая часть их обитает в воде.
У низших растений тело не расчленено на листья, корни и стебли. По сложению тела такие растения характеризуются как слоевцовые.
Весь мир живых организмов подразделяется на две большие группы: прокариоты (доядерные) и эукариоты (ядерные).
К прокариотическим организмам относятся (см. стр. 56) бактерии и так называемые сине-зеленые водоросли. Они выделяются отсутствием в их клетках ядер, что, вероятно, свидетельствует о сохранении ими древней организации клетки.
У эукариотических организмов обязательным структурным элементом клетки является ядро. Среди эукариотов различают много отделов (см. стр. 57).
Слоевцовые эукариотические растения бывают и автотрофными, тогда их называют водорослями, и гетеротрофными; объединяющего общепринятого термина для обозначения последних нет. В эту категорию входят грибы и миксомицеты (слизевики). Нередко эту категорию гетеротрофных низших растений понимают в широком смысле, присоединяя к ним бактерии из числа прокариотических организмов. Аналогично к числу водорослей относят прокариотические цианеи, называя их сине-зелеными водорослями.
Рассмотрим кратко некоторые из названных групп.
Бактерии и актиномицеты условно могут быть отнесены к растениям, хотя, может быть, не имеют прямых родственных связей с другими растениями. Огромное большинство бактерий - гетеротрофные организмы. Лишь немногие из них хемотрофны. Они синтезируют органическое вещество за счет химической энергии, освобождающейся при окислении в их теле неорганических соединений. Значительная группа бактерий содержит пигменты (бактериохлорин или бактериопурпурин). Среди бактерий преобладают одноклеточные, но есть и нитчатые многоклеточные. Бактерии способны к очень быстрому размножению делением. Внутри клетки некоторых бактерий, особенно палочковидных, образуется спора, которая высвобождается после разрушения оболочки бактерии и, имея собственную защитную оболочку, сохраняет жизнеспособность даже в крайне неблагоприятных условиях температуры и влажности. Споры лучше переносят очень низкие температуры, нежели высокие. У бактерий существует примитивный тип полового процесса. В их клетках содержится ядерный материал (рис. 4); они способны к конъюгации.
Водоросли (табл. 1, рис. 5) - фототрофные хлорофилл со держащие организмы. Они бывают одноклеточными,колониальными и многоклеточными. У примитивных зеленых водорослей развивается только один хлоропласт, иногда очень крупный. Но такой тип пластиды имеет относительно малую поверхность. У более высокоорганизованных водорослей хлоропласты сильно расчленены или (например, у харовых) хлоропласты мелкие, сферические, линзовидные; в каждой клетке их очень много, вследствие чего совокупность их обладает большой поверхностью световосприятия. Обитают водоросли в океанах, морях, озерах, прудах, а также в почвах, на суше и даже в атмосфере (во взвешенном состоянии).
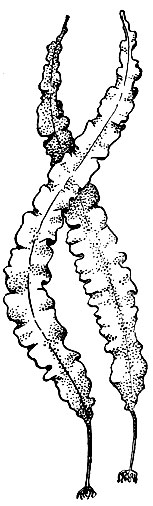
Рис. 5. Водоросль ламинария
Под водорослями, как видно из предыдущего, объединяют несколько отделов слоевцовых автотрофных растений, обычно живущих в воде. Среди водорослей можно различать три объединения отделов, характеризуемых составом пигментов, играющих роль в поглощении световой энергии в связи с фотосинтезом.
В первой группе наряду с хлорофиллом а есть еще хлорофилл b и набор каротиноидов. Последние, однако, как правило, не изменяют своим присутствием чисто-зеленую хлорофильную окраску. Это - зеленые и еще некоторые отделы водорослей, для большинства которых характерно образование в клетках крахмала.

Рис. 6. Строение гриба: 1 - внешний вид гриба (шампиньон); 2 - прорастающий 'рожок' спорыньи; 3 - спорынья на ржи
Во второй группе водорослей наряду с хлорофиллом а есть второй хлорофилл, но иной, чем у зеленых, - хлорофилл с. Здесь также имеются каротиноиды, в том числе специфичные, у зеленых не обнаруживаемые. Каротиноиды в пигментном комплексе водорослей этих групп участвуют в фотосинтезе, благодаря им окраска их бывает золотистой, желтой, бурой и буровато-зеленой. Крахмал у этих растений заменен другими углеводами. Сюда относятся отделы: золотистые, диатомовые, бурые водоросли (рис. 5).

Рис. 7. Гриб белый. Плодовое тело
Третью группу составляют водоросли, содержащие, кроме хлорофилла и каротиноидов, особые пигменты - фикобилины: синий фикоциан и красный фикоэритрин. Хлорофилл здесь один (а), но иногда (у некоторых красных водорослей) к нему присоединяется хлорофилл d. Эту группу составляют красные водоросли. По составу пигментов, кроме хлорофилла d, они сходны с прокариотическими цианеями.
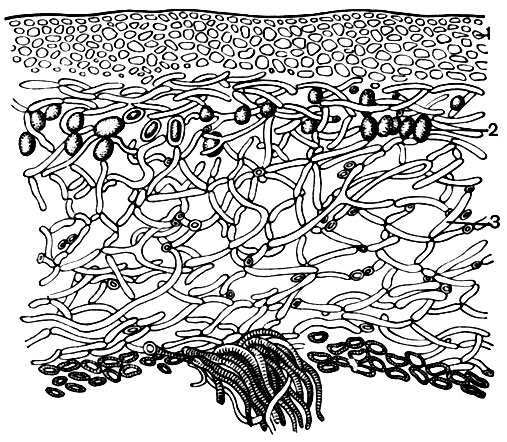
Рис. 8. Поперечный разрез лишайника: 1 - корковый слой; 2 - включения клеток водорослей; 3 - гифы гриба. Внизу видны ризоиды
Многие, особенно одноклеточные и колониальные, водоросли, принадлежащие к разным отделам, представляют важную часть планктона Мирового океана. Они являются первичными продуцентами органического вещества, за счет которого существует весь животный мир океана.
Кроме фитопланктона, в толще морей и океанов есть еще и фитобентос, представленный бурыми, красными, отчасти зелеными водорослями. Бурые образуют у берегов мощные подводные джунгли. Миллионы тонн водорослей после шторма выбрасываются на берег. В Атлантическом океане широко известны огромные скопления дрейфующих саргасс, представляющих сообщество водорослей и различных простейших животных организмов. Многие водоросли являются пищей мелких животных, которых в свою очередь поедают рыбы, поэтому рыбный промысел связан с планктоном и бентосом. В зарослях бентоса укрываются мальки рыб. Водоросли снабжают водные бассейны кислородом. Он, окисляя загрязненные воды, способствует очистке водоемов, например, от гнилостных анаэробных бактерий. Водоросли широко используются человеком в промышленности и сельском хозяйстве, например как удобрение в приморских странах, некоторые используются в пищу.

Рис. 9. Лишайники: вверху - кладония и пельтигера; внизу - цетрария и кладония
Кроме практического значения, водоросли имеют немалый теоретический интерес. В частности, надо отметить, что половой процесс, характерный для высших растений, которые, несомненно, возникли из каких-то водорослей, эволюционно вырабатывался у последних. Сравнивая половые явления у разных водорослей, приходят к выводу, что, вероятно, первичной формой полового процесса была хологами я, состоявшая в слиянии свободноживущих эукариотических клеток, не имевших еще клеточной оболочки. На смену хологамии пришла изогамия; в этом случае организм вырабатывает специальные подвижные за счет жгутиков клетки - гаметы, равной величины и одинаковой формы. Дальнейшая дифференциация гамет по величине привела к анизогамии (или гетерогамии) и, наконец, к оогамии. В последнем случае неподвижная оплодотворяемая клетка - яйцо (яйцеклетка) - оплодотворяется активно подвижной мужской гаметой - сперматозоидом.
У водорослей впервые эволюционно возникло и закрепилось чередование в цикле развития полового и бесполого поколений, т. е. спорофита игаметофита, а соответственно им - диплофазы (двойной набор хромосом) игаплофазы (гаплоидный, т. е. однократный, набор хромосом).
Грибы (табл. 1, рис. 6, 7) - гетеротрофные организмы, лишенные хлорофилла и пластид. Они ведут паразитный или сапрофитный образ жизни. Только немногие из них обитают в водной среде. На суше они достигли огромного разнообразия видов и биотипов, а также широкого географического распространения. Тело большинства грибов состоит из особых нитей, называемых гифами. Всю совокупность гиф гриба называют мицелием. Поперечные перегородки внутри мицелия есть только у высших грибов, однако они не сплошные, а с отверстием в средней части. У многих высших грибов нити мицелия развиваются тесно соединенными пучками, образуя длинные тяжи; они могут пронизывать древесину деревьев, деревянные балки в строениях. Клетки многих грибов многоядерные. Мицелий гриба бывает поверхностным и внутренним, пронизывающим субстрат. Множество грибов живет паразитами зеленых высших растений; в частности, они опаснейшие враги культурных растений, вызывая вспышки болезней - эпифитотии - среди них. В разных странах Земли ежегодно большие площади полевых и древесных насаждений дают низкие урожаи, а нередко и погибают вследствие нападения грибных паразитов (рис. 6).
Грибные паразиты обладают многими приспособлениями для размножения и распространения, для длительного сохранения жизнеспособности. Они не только паразитируют на надземных частях зеленых растений, но и заселяют почвы, поражают корневые системы культурных и диких растений, а споры их в огромном количестве постоянно находятся в биосфере. Заспоренность ее достигает чудовищных размеров. Распространение спор грибов воздушными течениями связано с разнообразными баллистическими механизмами, выбрасывающими споры, которые подхватываются и разносятся токами воздуха. Иногда образуются как бы споровые облака. Особые ловушки на самолетах позволили установить среднюю концентрацию спор многих грибных паразитов на разных высотах. Даже простые ловушки в двух метрах от почвы показали, что в среднем насчитывается 12 500 спор на 1 м3 воздуха. Споры потом оседают на растениях с дождем или приносятся ветром. Споры ржавчины пшениц, разносимые ветром, обнаруживаются даже над океаном, далеко от материков.

Рис. 10. Мох сфагнум

Рис. 11. Мох кукушкин лен
Грибы многих видов, особенно шляпочные, употребляются в пищу человеком и животными (рис. 7).
Положительна роль грибов как продуцентов ценных медицинских антибиотиков. В круговороте веществ (в биосфере) грибы, как и бактерии, играют важную роль разрушителей органического вещества, обеспечивающих возвращение в среду того, что изымают из нее зеленые растения. Принято считать, что грибы не нуждаются в световой энергии. Однако у большинства грибов образование спор и половой процесс происходят под влиянием коротковолновых лучей спектра, которые, вероятно, поглощаются каротиноидами.
Назовем еще одну группу гетеротрофных эукариотических низших растений - миксомицеты, или слизевики. Это совершенно своеобразная группа, отличная от всех других растений тем, что вегетативная их жизнь происходит в состоянии голой многоядерной плазмы, не покрытой оболочкой.

Рис. 12. Покров зеленого мха в лесу
К низшим растениям относятся и такие своеобразные организмы, как лишайники (табл. 1, рис. 8 и 9). Слоевище (тело) лишайника составлено двумя организмами - грибом и водорослью, находящимися во взаимно полезном сожительстве - симбиозе. Снаружи лишайник обычно покрыт плотным корковым слоем из тесно сплетенных и видоизмененных гиф гриба. Внутренняя часть состоит из многочисленных гиф, оплетающих отдельные клетки и целые группы водорослей. Грибы, участвующие в симбиозе, относятся к высшим, т. е. к классу сумчатых, реже - базидиальных, а водоросли - из типа зеленых, реже - сине-зеленых водорослей.
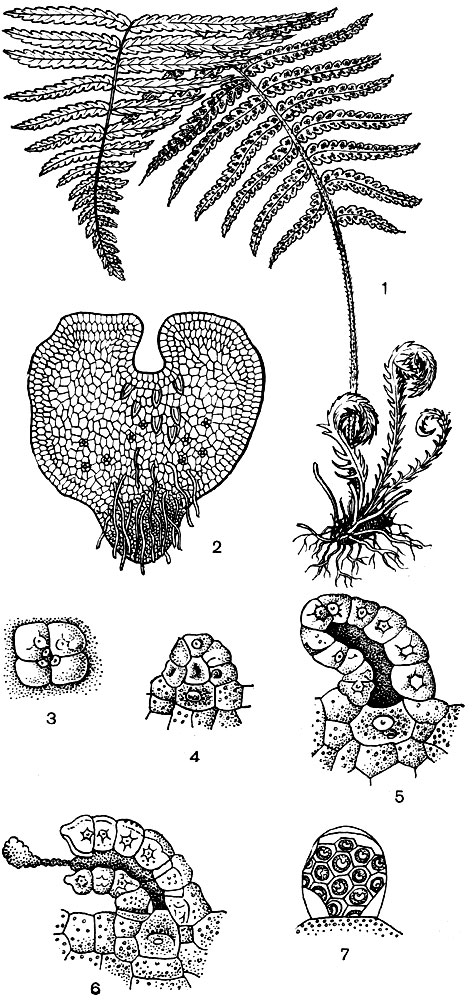
Рис. 13. Развитие папоротника: 1 - взрослое растение (спорофит); 2 - заросток (гаметофит); 3 - 6 - развитие архегония; 7 - зрелый антеридий
Симбиоз возник в природе на такой физиологической основе: гриб, прикрепляющий лишайник к субстрату, обеспечивает водоросль водой и растворенными в ней минеральными веществами, а также системой ферментов; водоросль в процессе фотосинтеза вырабатывает углеводы, которые используются как самой водорослью, так и грибом. В значительной мере водоросль получает воду и пыль, содержащую неорганические вещества, из атмосферы.
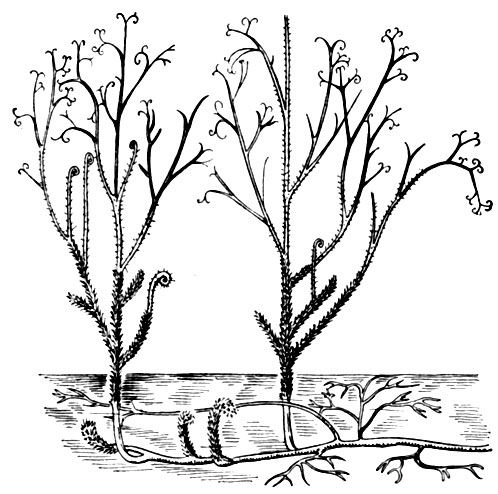
Рис. 14. Псилофит астероксилон (на стеблях видны зеленые выросты - предшественники листьев)
Лишайники - самые неприхотливые растительные организмы. Они поселяются на горных склонах и камнях, на коре деревьев, на поверхности обнаженных необрабатываемых почв и в других местах. Но эти растения крайне чувствительны к загрязненности атмосферы некоторыми газообразными примесями, поэтому их не встретишь в центральной части больших городов.
Рис. 15. Сигиллярия
Фотосинтез лишайников ослаблен, однако они способны к фотосинтезу в зимний период при низких температурах, доходящих до -35°С. В тундрах лишайники занимают огромные площади, и некоторые из них, так называемые ягели, служат основным кормом северных оленей в зимнее время, добываемым оленями из-под снежного покрова (рис. 9).
До сих пор речь шла о низших растениях. Теперь кратко расскажем о высших зеленых растениях. К ним относятся давно вымершие псилофиты (первые поселенцы суши, происшедшие от зеленых или бурых водорослей), затем ныне живущие мхи, папоротники, голосеменные и покрытосеменные (цветковые) растения. Все они, как правило, обитатели суши, но среди них есть и обитатели водоемов.

Рис. 16. Лист папоротника
Вегетативное тело большинства высших растений расчленяется на корни и побеги, состоящие, в свою очередь, из стеблей и листьев. В осевых органах их спорофита (корнях, стеблях), а иногда и гаметофита (например, зеленые лиственные мхи) имеется стела (внутренняя система проводящих элементов). Типичен для высших растений женский половой орган - архегоний, наблюдаемый у мохообразных, папоротников и большинства голосеменных. В ходе эволюции архегоний упрощался, и уже у некоторых голосеменных он отсутствует. В наиболее молодой по времени появления группе высших растений - у цветковых, или покрытосеменных, от архегония осталась лишь важнейшая его часть - яйцеклетка, развивающаяся в так называемом зародышевом мешке. У мхов нет корней; эти растения прикрепляются к субстрату особыми волосками - ризоидами. Мхи представляют одну из начальных, но слепую (без продолжения) ветвь эволюции, развивающуюся на основе преобладания в цикле гаплоидного гаметофита. Их спорофит не приспособлен или мало приспособлен к фотосинтезу и живет не самостоятельно, а прикрепляется к гаметофиту.
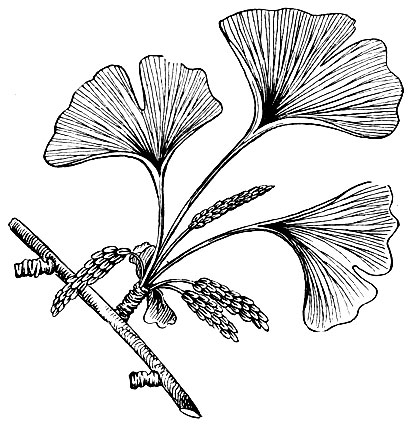
Рис. 17. Гинкго
В классе настоящих мхов (табл. 2, рис. 10, 11, 12) определилось расчленение надземного тела растения на стебель и листья, а одноклеточные ризоиды заменились многоклеточными. Однако сфагновые мхи во взрослом состоянии не имеют ризоидов. Обитая на очень влажных почвах, они накопляют в листьях и в поверхностных слоях стеблей много воды. Эти ткани состоят из лишенных плазменного содержимого клеток, и движение воды по ним совершается по закону капиллярности. Обширные по территории покровы сфагновых мхов обусловливают постепенное заболачивание местности. Из не вполне разлагающихся отмерших частей сфагнов в течение длительных промежутков времени формируются мощные торфяники. Сфагновые мхи распространены в лесной зоне и в тундре северных и умеренных широт СССР до 70° с. ш. и в горах Кавказа. Торф - ценное топливо и удобрение. Сфагновые мхи могут самовозгораться и нередко являются причиной лесных пожаров.
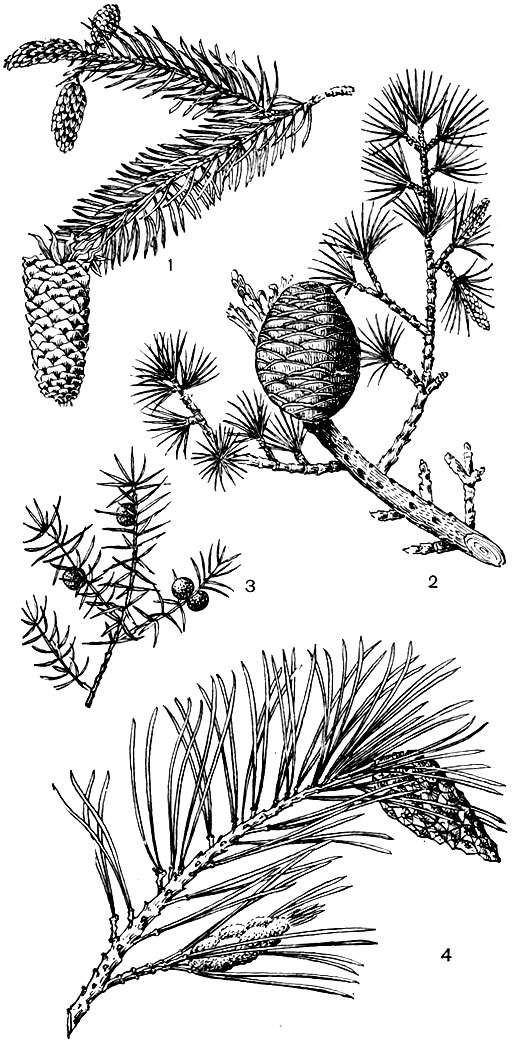
Рис. 18. Хвойные: 1 - ель; 2 - кедр; 3 - можжевельник; 4 - сосна
Зеленые мхи, за немногими исключениями, не имеют на поверхности стеблей тканей, аккумулирующих воду. Зато в стеблях и листьях (жилки) выделяется пучок продольно вытянутых клеток, по-видимому, проводящей функции. Иногда такой пучок дифференцирован на элементы, проводящие органические вещества и проводящие воду. Зеленые мхи обитают преимущественно во влажных местах, но некоторые отличаются замечательной способностью переносить длительное обезвоживание. Зеленые мхи образуют обширные покровы в хвойных лесах, лесотундре, тундре и на некоторых типах болот. Много среди них эпифитов, обитающих на стволах и ветвях деревьев. Папоротники (табл. 2, рис. 13) почти исключительно наземные растения, распространяющиеся спорами, как и мхи; им свойственно чередование поколений. Отличаются от мхов преобладанием в цикле развития спорофита с корнями и олиственными побегами. Стебель спорофита имеет разнообразное и сложное строение. В нем всегда выражена стела, то компактная, то сильно расчлененная на проводящие пучки. Первые представители папоротникообразных появились не позже конца силурийского периода и, вероятно, были сходны с псилофитами, широко распространенными в девонском периоде. Псилофиты имели ось с примитивной стелой (протостелой). На стелющейся части оси были короткие всасывающие волоски (ризомоиды). От стелющейся оси кверху направлялись вертикальные, дихотомически разветвленные оси, которые заканчивались верхушечными спорангиями; таким образом, концы ветвей были спороносными. Такой спорангий со своей ножкой называется теломом. Этот же термин применяется к ветвям последнего порядка, не имеющим спорангиев. Вертикальная ось (прообраз стебля) была зеленой. Наружная ткань представляла кору с фотосинтезирующими клетками, покрытую с поверхности эпидермисом с устьицами и кутикулой. Внутренняя же часть (протостела) состояла из трахеид и окружавших их примитивных ситовидных трубок. У более совершенных псилофитов на стеблях появились небольшие выросты, увеличивающие фотосинтезирующую поверхность. Псилофиты просуществовали на Земле около 50 млн. лет и вымерли. Долго оставались известными только ископаемые спорофиты, а половое их поколение (гаметофит) не был известен. В последнее время появились сообщения о нахождении псилофитов с архегониями, т. е. о гаметофитах этих растений.
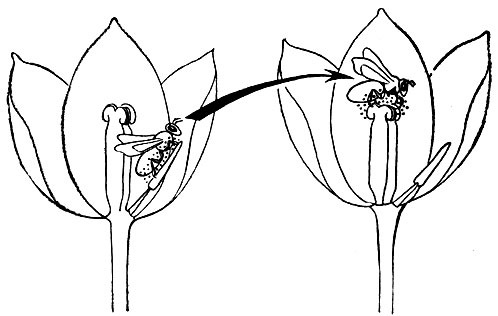
Рис. 19. Перекрестное опыление насекомыми
Псилофиты дали начало развитию разнообразных папоротников, к которым относятся и ныне живущие: плауны (плауны, селагинеллы и др.), хвощи и собственно папоротники. Плауны и хвощи представляют мелколистную линию эволюции, а папоротники - крупнолистную. У всех этих растений резко определились два поколения: одно бесполое (спорофит), другое половое (гаметофит). Спорофит - долговечен, гаметофит - эфемерен. На спорофите образуются спорангии со спорами. Спорангии у папоротников образуются на листьях. Это объясняется тем, что сам лист папоротника по происхождению представляет собой уплощенный, фасциированный стебель - листоветку (рис. 13, 16).
Споры после раскрытия спорангия разносятся ветром и попадают на почву. Из проросшей споры образуется гаметофит, так называемый заросток. У папоротника, например, гаметофит представляет маленькую сердцевидную зеленую фотосинтезирующую пластинку, на нижней стороне ее появляются всасывающие ризоиды, а кроме них, еще половые органы - антеридии и архегонии. Весь заросток, как и оба половых органа, гаплоиден. В антеридиях образуются имеющие жгутики подвижные сперматозоиды, в архегониях - яйцеклетки. Сперматозоиды передвигаются в дождевой воде или росе к архегониям; один из них попадает в яйцеклетку, происходит оплодотворение. Из оплодотворенной диплоидной яйцеклетки возникает диплоидный зародыш, развивающийся далее в крупное, иногда долговечное растение (например, существуют и сейчас древовидные спорофиты папоротников).
Подавляющее большинство древних папоротникообразных растений вымерло в давно минувшие геологические эпохи (рис. 14, 15).
От хвощевидных (членистых) растений в современную эпоху сохранилось всего около двух с половиной десятков видов; несколько больше осталось плаунов, но только травянистых, в то время как в палеозое этот класс, по-видимому, изобиловал деревьями. В лучшем положении оказались папоротники; наиболее молодая ветвь их, развиваясь с позднего карбона, богато представлена и в современной флоре, их известно более 9000 видов.
На территории СССР встречаются папоротники только травянистого типа (рис. 16). Они заселяют овраги, сырые леса, иногда живут в воде. Древовидные папоротники встречаются только в тропических и субтропических странах.
Эволюция папоротникообразных привела к появлению и развитию так называемой разноспоровости. Споры и производящие их спорангии становились разными. Появились микроспорангии, мелкие споры которых прорастают в мужской гаметофит, и мегаспорангии, споры которых прорастают в женский гаметофит. У всех папоротников, плаунов и хвощей нет главного корня, а есть только придаточные корни на корневищах.

Рис. 20. Однодольные: слева - любка двулистная; справа - ячмень
В своей совокупности на Земле мхи и папоротники создают часть органической материи (как теперь говорят - "биомассы") и уже поэтому заслуживают изучения и внимания.
В верхнедевонском периоде истории Земли возникли и распространились достигшие максимального развития в каменноугольном периоде семенные папоротники. Они известны только по ископаемым остаткам. Эти растения явились как бы новаторами в эволюции наземной флоры. В их внешнем облике много общего с папоротниками, и прежде всего крупные листья с многократно рассеченными листовыми пластинками. Но они существенно отличались от папоротников тем, что на их листьях формировались зачатки семян - семяпочки. По общему плану строения эти органы вполне соответствовали семяпочкам современных голосеменных. Здесь был покров (интегумент) с отверстием наверху (микропиле); в большей своей части он обычно срастался с наружной поверхностью покрываемого им массивного мегаспорангия, оставаясь свободным только наверху. В мегаспорангии, который у голосеменных обычно называют нуцеллусом, развивалась только одна мегаспора, которая здесь же и прорастала. В результате этого внутри мегаспоры образовывался мелкоклеточный заросток. Однажды в нем были найдены окаменевшие архегонии.
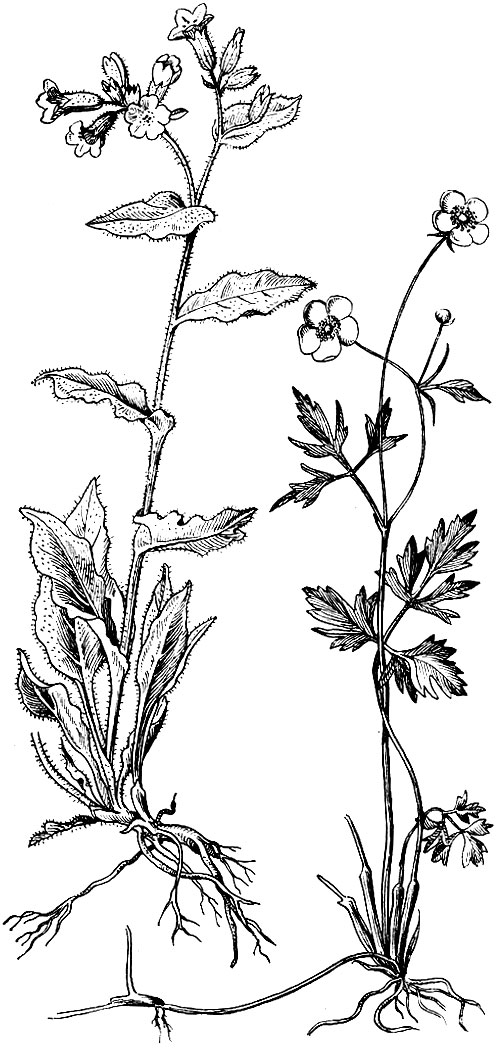
Рис. 21. Двудольные: слева - медуница; справа - лютик
Из сказанного видно, что семенные папоротники заслуженно получили свое наименование. Они могут рассматриваться как родоначальники по отношению к прочим голосеменным. Такое название им дано потому, что их семя-почки располагаются открыто (голо). К голосеменным относятся саговники (сохранилось около 100 видов их до нашего времени), гинкговые (сохранился один вид, главным образом в культуре) (табл. 3, рис. 17), хвойные (табл. 3, рис. 18) - наиболее обширная группа голосеменных современной флоры, оболочкосеменные. Нацело вымерли - беннеттитовые, кордаиты и семенные папоротники. Отметим, что главный корень появился у голосеменных, начиная с саговниковых. Кроме того, саговники и гинкговые - последние в эволюции наземных растений из тех, у которых оплодотворение осуществляется подвижными сперматозоидами, обладающими жгутиками. Отметим также, что семя возникло у растений раньше, чем цветок и плод.

Таблица 1. Водоросли, грибы и лишайники: мухомор
Из некогда господствовавшего отдела голосеменных растений в современной флоре широко развиты хвойные. Из всех голосеменных хвойные наиболее приспособились к современным условиям биосферы. Ныне в этой группе растений насчитывают около 550 видов. Значительно преобладают хвойные в северном полушарии. Сосна, ель, пихта, кедр, лиственница, кипарис, туя, можжевельник и тис широко распространены в северном полушарии, особенно вдоль Тихого океана. Наиболее древний очаг происхождения хвойных - китайский. Наибольшее число видов сосны (около 50) обитает в Мексике. Травянистые растения среди хвойных и голосеменных вообще неизвестны. В южном полушарии из хвойных распространены подокарпус, араукария и некоторые другие.

Таблица 1. Водоросли, грибы и лишайники: опята
Покрытосеменные, или цветковые, растения (табл. 4 - 6) по сравнению с другими высшими растениями в настоящее время значительно преобладают в растительном покрове. Появившись, вероятно, в юрском периоде, около середины мелового периода они стали необыкновенно быстро распространяться, обнаружив исключительную способность к видообразованию. Русский ботаник М. И. Голенкин дал им удачное название "победителей в борьбе за существование". Отличительной особенностью покрытосеменных служит наличие плода, развивающегося, в основном, из завязи цветка. Цветок - это укороченный спороносный побег, в котором женские спороносные листья (мегаспорофиллы) превращены в плодолистики, а мужские (микроспорофиллы) - в тычинки. Цветок служит для образования микроспор - пыльцы - в пыльниках и макроспор в семяпочках. В результате сложного процесса развития в пылинках формируются мужские безжгутиковые гаметы, а в процессе развития макроспоры (зародышевый мешок) - яйцеклетка. После оплодотворения, которому предшествует опыление, образуется зародыш, заключенный в семени. Семена находятся в плодах.
Таблица 1. Водоросли, грибы и лишайники: накипные лишайники (на скалах)
Процессы цветения, оплодотворения, образования плодов и семян - весьма сложные многообразные явления, связанные не только с внутренними генетическими закономерностями, но и с участием в опылении ветра, многих видов насекомых (жуков, пчел, шмелей, ос, мух, муравьев, трипсов и др.), птиц (особенно колибри) и даже некоторых млекопитающих. Широкое распространение перекрестного опыления свидетельствует об эволюционном преимуществе его в сравнении с самоопылением. Перекрестное опыление обеспечивает широкий обмен генетическим материалом (рис. 19).

Таблица 1. Водоросли, грибы и лишайники: нитчатая водоросль (в горячем источнике)
Значительное большинство видов цветковых растений сосредоточено в тропических широтах (около 120 000 видов); в субтропических, умеренных и холодных широтах - не менее 22 000 видов. Тропические виды по преимуществу древесные (даже в тех случаях, когда принадлежат к семействам умеренных широт).
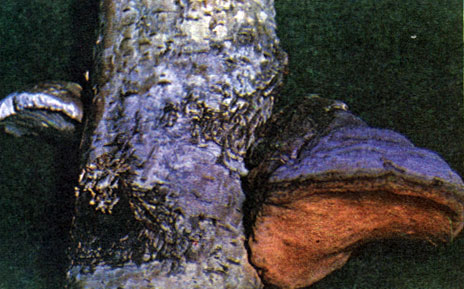
Таблица 1. Водоросли, грибы и лишайники: трутовик
Цветковые растения обычно разделяются на двудольные и однодольные (рис. 20, 21). Первый из этих классов (двудольные) по числу видов в 4 - 5 раз превосходит второй (однодольные). В биохимическом отношении двудольные отличаются многообразием и синтезом более сложных веществ. Но нельзя забывать, что такие однодольные, как пшеница, рис, рожь, кукуруза, сахарный тростник, банан, финиковая пальма, незаменимы в пище человека.

Таблица 1. Водоросли, грибы и лишайники: порфира
Сочетание видов растений на земном шаре сложилось исторически в процессе естественного и искусственного отборов, на основе наследственной изменчивости, гомологических рядов этой изменчивости, на основе мутаций и гибридизации, а также рекомбинаций. В естественной обстановке виды развиваются и успешно размножаются там, где они хорошо приспособлены к среде, в которой обитают. Организация растения, его цикл развития сложились под совокупным воздействием генотипа и среды. На горьком опыте человечество убедилось в огромном влиянии леса на биосферу и ухудшении климата после уничтожения лесов.

Таблица 2. Мхи и печеночник: маршанция
Каждому континенту, его большим физико-географическим районам свойственна своя флора, т. е. совокупность семейств, родов и видов растений. Они сочетаются в так называемые фитоценоз ы, т. е. естественные сообщества видов. Совокупность фитоценозов на определенной большой территории составляет ее растительность, или растительный покров.

Таблица 2. Мхи и печеночник: мхи - плевроциум
Земной шар разделен на ряд флористических царств и областей. Флора их имеет общее историческое происхождение. В состав каждой флоры входят свои эндемические (свойственные только этой флоре) семейства, еще чаще - эндемические роды и значительное число эндемических видов. Конечно, флоры не замкнуты, виды проникают в другие флористические области независимо от человека, но в большей мере благодаря людям (см. стр. 117).
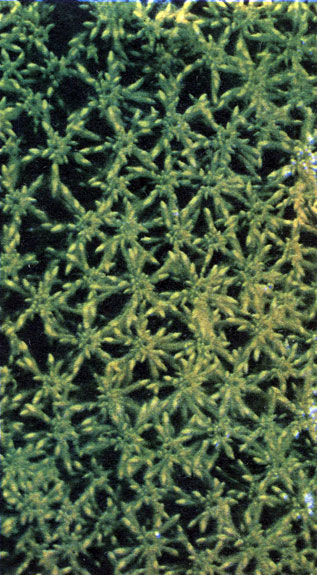
Таблица 2. Мхи и печеночник: мхи - сфагнум
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'