Вегетативные органы саговниковых
Листья и крона
Верхушку ствола саговниковых венчает крона перистых листьев, делая этим растения похожими на древовидные папоротники или пальмы. Среди всех голосеменных, живущих в наши дни, только саговниковые сохранили папоротниковидный тип листа. Особенно это относится к стангерии шерстистой (Stangeria eriopus), листья которой настолько напоминают лист папоротника, что сначала она и была описана как один из видов папоротников, получив название ломарии шерстистой (Lomaria eriopus). Еще большее сходство с папоротниками придают этим растениям улиткообразно закрученные на верхнюю сторону стержни (рахисы) молодых листьев и их перышек. Правда, такая особенность свойственна далеко не всем саговниковым. У диоона, цератозамии, энцефаляртоса и макрозамии стержни и перышки в листосложении прямые.
У низкорослых саговниковых с коротким надземным стволом или стеблем, погруженным в почву, листьев обычно мало и образуются они последовательно друг за другом. На крупных экземплярах у других родов их насчитывается значительно больше, в исключительных случаях до 100-150 штук. Появляются они в виде воронки или короны по нескольку сразу, как бы внезапно, с различными интервалами во времени. Так, у саговника завитого (Cycas circinalis) и саговника Туара кроны листьев образуются два раза в год, причем у каждого вида в свои сроки, а у некоторых видов энцефаляртоса - только раз в два года или еще реже.
Между кронами зеленых листьев, чередуясь с ними, нередко развиваются толстые, редуцированные чешуевидные листья - катафиллы. Они довольно короткие, длиной не более 5-10 см. Катафиллы с их густым войлочным опушением служат своеобразной защитой нежным растущим частям кроны. Перед тем как тронутся в рост зеленые листья, верхушки чешуевидных листьев спирально изгибаются, напоминая толстые скрученные веревки.
Давно замечено, что продолжительность существования кроны у саговниковых может быть довольно длительной. Так, на отдельных экземплярах саговников завитого и Румфа ниже листьев самой молодой кроны можно увидеть иногда две или три более ранние кроны, а у саговника поникающего - сосчитать на одном растении до пяти последовательных генераций листовых крон (табл. 38). Со стороны это выглядит довольно любопытно. Вершину растения венчает изящная корона - почти вертикально стоящие молодые светло-зеленые листья. Чуть ниже под ними - несколько отклоненные в сторону жесткие темно-зеленые листья кроны предыдущего года, за которой следует ранее образовавшаяся крона из горизонтально распростертых листьев. Еще ниже размещается крона из опущенных вниз листьев с причудливо изогнутыми концами. Наконец, последняя, в свою очередь, может скрывать от глаз уже высохшие висящие вдоль ствола, перевернутые стержни (рахисы) самых старых листьев. Продолжительность жизни каждого листа саговниковых от 3 до 10 лет.
Листья саговниковых значительно варьируют по величине. У таких крошечных растений, как замия карликовая, они имеют часто всего 5-6 см в длину; у видов саговника или цератозамии длина их может достигать 3 м; некоторые виды энцефаляртоса обладают 5-6 - метровыми листьями, напоминающими перья огромной сказочной птицы. Листья у большинства саговниковых однажды перистые, и лишь у бовении и немногих видов саговника они дважды перистые. У некоторых видов саговниковых не всегда легко установить, имеем ли мы дело с простыми или сложными листьями. Например, листочки микроцикаса прикрепляются к стержню всем основанием, но имеют отделительный слой и опадают нередко раньше, чем отмирает весь лист (рис. 164,7). То же наблюдается и у некоторых видов рода саговник, с той лишь разницей, что листочки у них могут иметь ясно выраженные черешочки.
Число сегментов (их называют также перышками) в одном листе у различных представителей этой группы сильно варьирует. Листья некоторых видов рода замия имеют не более 4-5 перышек, а у крупных экземпляров саговника и энцефаляртоса их может быть более 180-200. Форма и величина сегментов разнообразны, сегменты могут быть овальными, яйцевидными, слегка вытянутыми или узколанцетными, цельнокрайними, пильчатыми или зазубренными и т. д. (рис. 164). Цератозамия узколистная (Ceratozamia angustifolia) и саговник завитой - обладатели наиболее длинных перышек, достигающих в длину 40 см при ширине не более 2-2,5 см.
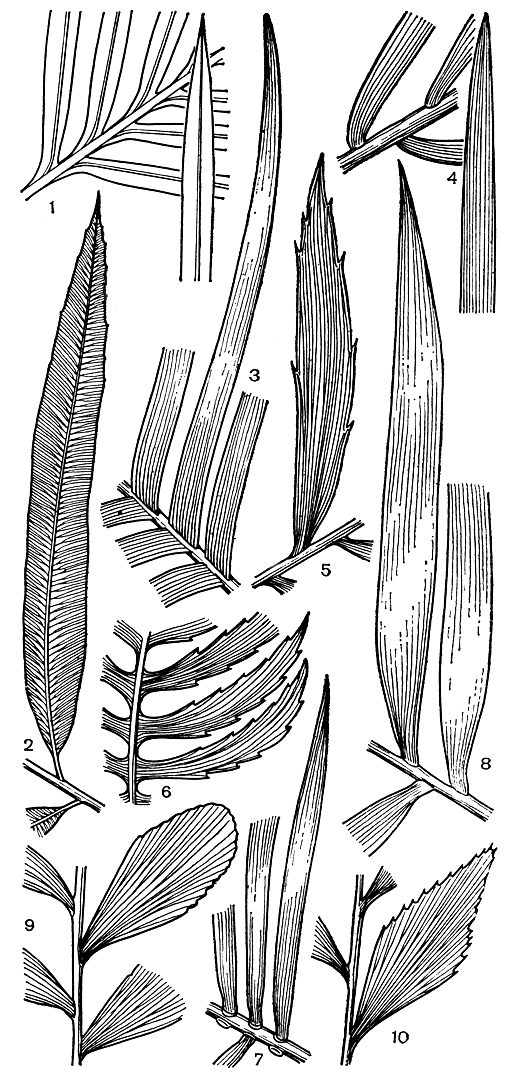
Рис. 164. Форма перышек (листочков, сегментов) у листьев саговниковых: 1 - саговник Румфа (Cycas rumphii); 2 - стангерия шерстистая (Stangeria eriopus); 3 - лепидозамия Перовского (Lepidozamia peroffskiana); 4 - макрозамия Мора (Macrozamia moorei); 5 - энцефаляртос Альтенштейна (Encephalartos altensteinii); 6 - диоон колючий (Dioon spinulosum); 7 - микроцикас красивокронный (Microcycas calocoma); 8 - цератозамия мексиканская (Ceratozamia mexicana); 9 - замия карликовая (Zamia pygmaea); 10 - бовения мелкопильчатая (Bowenia serrulata)
По типу жилкования перышек все саговниковые можно разделить на три группы. К первой относятся виды рода саговник, имеющие единственную неразветвленную среднюю жилку. Перышки стангерии обладают тоже ясно выраженной средней жилкой, от которой, однако, отходят вильчато разветвляющиеся боковые жилки. Для остальных представителей саговниковых характерен третий тип жилкования, при котором главная жилка отсутствует, а имеется большее или меньшее число параллельно идущих жилок, отчетливо заметных на нижней стороне листа.
Стержни, к которым прикрепляются перышки, длинные и довольно толстые. У многих видов они снабжены острыми твердыми шипами, расположенными двумя рядами на верхней стороне или сбоку и переходящими постепенно в перышки (сегменты), с которыми они имеют общее происхождение. Защищены от повреждения животными у многих видов и сами перышки. Их края, участки или лопасти часто заканчиваются острой колючкой. Особой колючестью отличаются виды замии и энцефаляртоса.
Листья саговниковых обладают целым рядом структурных особенностей, позволяющих этим растениям жить в условиях более или менее сухого климата, на открытых солнцу пространствах, иногда почти голых скалах и крутых обрывах. Как правило, листья саговниковых плотные, жесткие, с толстым слоем кутикулы, помогающим не только экономить влагу, но и защитить лист от проникновения в него патогенных грибов и бактерий. Кутикулу можно обнаружить даже внутри листа, например на внутренних оболочках эпидермальных клеток, в местах их контактов с межклетниками. Палисадная ткань в листе развита достаточно хорошо, при этом верхняя часть ее хлорофиллоносных клеток часто бывает сильно лигнифицирована.
Глубокое погружение устьиц в ткань листа - еще один способ уменьшить потерю влаги. В углублении над устьицем часто скапливаются пары воды. У бовении, стангерии, многих видов саговника устьица почти непогруженные или слегка погруженные. У других же (лепидозамия, саговник поникающий, многие виды энцефаляртоса и диоона) они, напротив, как бы опущены в мезофилл листа.
В отличие от зрелых молодые листья саговниковых нередко бывают опушены. Предполагают, что такое своеобразное одеяло из волосков, покрывающее нежные ткани растущего листа, защищает их от резких изменений погодных условий.
Хотя листьям саговниковых свойственны многие морфологические и анатомические черты, связанные с засухоустойчивостью, эти растения нельзя считать типичными ксерофитами. Ксероморфную структуру листьев можно объяснить отчасти своеобразием внутреннего устройства всего растения, обладающего слабо развитой проводящей системой, неспособной в должной мере обеспечивать крону водой.
Стебель
Саговниковые - древовидные растения, отличающиеся по форме и высоте стебля, однако никогда не достигающие таких больших размеров, какими славятся многие современные хвойные.
Обычно; когда говорят о саговниковых, представляют себе довольно крупные растения с неветвящимся колонновидным стволом, покрытым толстым шершавым панцирем из остатков черешков листьев.
На самом деле у саговниковых имеются два основных типа ствола: подземный или полуподземный клубневидный, иногда почти сферический, похожий на репу, и надземный колонновидный.
Колонновидный надземный ствол чаще привлекает внимание и лучше изучен. Наиболее высокими из живущих в наши дни саговниковых, имеющих такой тип ствола, считают лепидозамию Хоупа, достигающую высоты 18-20 м. Второе место по высоте принадлежит трем представителям саговниковых - диоону колючему (Dioon spinulosum), саговнику Румфа и одному из видов энцефаляртоса, имеющим ствол высотой до 16 м и диаметром до 1,5 м. Относительно большими размерами отличаются также энцефаляртос поперечно-жильчатый (Encephalartos transvenosus), саговник новокаледонский (Cycas neocaledonica) и микроцикас красивокронный (Microcycas calocoma), достигающие в высоту иногда 10-12 м. Однако все эти цифры исключительны даже для перечисленных видов, имеющих, как правило, высоту не более 6-7 м. Обычно саговниковые, вырастающие до 2,5-3 м, уже считаются достаточно высокими.
Растут саговниковые, за немногими исключениями, очень медленно. Сначала рост в высоту, как обычно, идет у них за счет очень крупной верхушечной точки роста, пожалуй, самой массивной среди всех существующих голосеменных и цветковых растений. Но позже, когда образуются микро- и мегастробилы, характер роста стебля может измениться.
Стебли саговниковых, образующих верхушечный микро- или мегастробил, как бы заканчиваются им. В ось стробила на ранних стадиях его развития входит фактически весь проводящий цилиндр ствола. Верхушечная точка роста с появлением стробила прекращает свое существование. Но как же при этом продолжается рост? Уже в период развития и созревания стробила ниже его на стволе, в пазухе одного из листьев кроны, пробуждается почка. Растущее вверх ответвление отклоняет стробил в боковое положение, а сама ветвь распрямляется и скоро по виду становится как бы прямым продолжением образовавшего ее ствола. Появляется еще один новый стробил, и весь процесс повторяется, а новый участок ствола становится "ветвью ветви". Такой симподиальный способ нарастания характерен для родов микроцикас, цератозамия, диоон, замия (рис. 165).
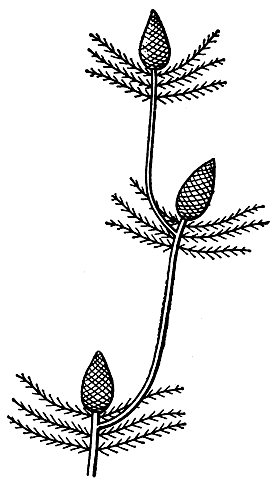
Рис. 165. Схема симподиального нарастания ствола у саговниковых
Австралийские макрозамии и африканские энцефаляртосы характеризуются, напротив, боковым заложением стробилов, которые формируются под точкой роста, в пазухах листьев самой молодой кроны, иногда в виде кольца окружая верхушку ствола. При этом точка роста сохраняет свой вегетативный характер и способность к дальнейшему моноподиальному росту. У женских экземпляров рода саговник пучок мегаспорофиллов на верхушке располагается так рыхло, что почка прорастает его насквозь.
Высокий и толстый ствол некоторых саговниковых содержит много воды и настолько тяжел, что относительно слабая корневая система не всегда в состоянии удержать его в вертикальном положении, особенно в открытых ветрам местах. Постепенно он наклоняется, а иногда и совсем полегает на землю.
Обычно в естественных условиях у саговниковых прямой, неветвящийся стебель, лишь иногда он может слабо вильчато ветвиться. У растений с подземным или полуподземным типом ствола, как, например, у стангерии, такое явление наблюдается исключительно редко. Несколько чаще можно встретить ветвящиеся формы колонновидных саговниковых (рис. 162), особенно выращиваемых в оранжереях и парках.
Ветвление вызывается какими-либо механическими повреждениями стебля, в результате которых трогается в рост почка. Этой особенностью саговниковых искусно пользуются японские садоводы. Нанося специально повреждения растениям саговника поникающего, они заставляют их многократно "вильчато" ветвиться, создавая причудливые карликовые формы, известные во всем мире под названием "ши-ши", "голова льва" или "древовидная пальма". Некоторые такие растеньица достигают возраста нескольких сотен лет и имеют при этом более 20 крон.
Иногда у саговниковых и в природе может проявляться ложнодихотомическое ветвление. На стволе в нижней части старого листового основания начинает развиваться пазушная почка, которая первоначально имеет форму луковички. Такая воздушная луковичка состоит из нескольких чешуевидных листьев, сидящих на крошечном стебле. Это маленькое растеньице по мере роста сминает листовое основание и, развиваясь, дает в конце концов такую же крону, как у взрослой особи. При этом у основания новой ветви нередко появляются придаточные корни, так что ее можно отсадить как самостоятельное растение. Если пазушная почка в момент пробуждения располагалась лишь немного ниже кроны, то образовавшаяся ветвь легко достигает вершины растения и это может выглядеть так, будто имеет место дихотомическое ветвление. Луковички могут появляться в любом месте стебля - от основания до его вершины, и от их положения во многом зависит внешний вид растения.
Развитие пазушных почек на стеблях саговниковых имеет большое значение для сохранения в природе этих удивительных растений. Нередки случаи, когда на старых полегающих стволах на стороне, обращенной к земле, трогаются в рост многочисленные почки, из которых развиваются молодые растеньица и, укореняясь в почве, дают жизнь новому поколению.
Колонновидные стволы саговниковых могут быть гладкими, защищенными лишь перидермой, или чаще покрытыми толстым слоем (в 2-2,5 см) из остатков черешков листьев, имеющих ромбовидную форму. Более крупные из них принадлежат зеленым листьям, а меньшие по размерам - чешуевидным. Расположены эти ромбовидные основания по спирали и обычно так плотно, что поверхность ствола кажется одетой сплошным шершавым панцирем. Длина листовых оснований, составляющих такое защитное образование, с годами уменьшается. От самых старых из них, расположенных в нижней части ствола, начинают отделяться тонкие, как бумага, слои, так что в конце концов диаметр основания стебля может стать заметно тоньше его верхней части.
Поскольку панцирь из остатков листовых оснований может сохраняться в течение всей жизни растения, эта особенность позволяет примерно определить возраст растения. Для этого подсчитывают число полос из крупных и мелких ромбиков, соответствующих числу крон зеленых листьев и катафиллов, и, учитывая периодичность появления крон (два раза в год, каждый год, раз в два года и т. д.), подсчитывают приблизительный возраст растения.
Сведения о предельном возрасте саговниковых крайне противоречивы. Немецкий ботаник Ю. Шустер (1932) указывал в своей монографии, что наиболее рослые растения макрозамий могут достигать двухтысячелетнего возраста. Позднее авторитетный специалист по австралийским саговниковым - Л. Джонсон - высказал убеждение, что самые их крупные экземпляры становятся такими относительно быстро - менее чем за 100 лет. Но это в отношении макрозамий. Представители других родов саговниковых, очевидно, могут быть медленнее растущими и более долговечными, например пятисотлетними. Известны сенсационные сообщения в газетах о саговниках-патриархах, якобы имеющих возраст в 10 или даже 15 тыс. лет, что, несомненно, является огромным преувеличением. Можно думать, что версии такого рода рождаются не только в погоне за сенсацией, но иногда и из стремления защитить эти уникальные растения от истребления. Лет 30 назад близ Брисбена была беспощадно вырублена группа растений лепидозамии Перовского (Lepidozamia peroffskiana). Среди них был особо крупный экземпляр, высотой 7,5 м, любовно названный местными жителями "Прадедушка Питер". Газетное сообщение о варварском уничтожении этого якобы самого древнего растения на Земле, естественно, привлекло внимание к саговниковым в целом как растениям, нуждающимся в очень бережном отношении.
Из всех ныне живущих семенных растений саговниковые являются наиболее типичными представителями маноксильной линии эволюции. На поперечном срезе стебля (рис. 166) бросаются в глаза широкая сердцевина и мощная многослойная кора; древесина представлена узким (0,2-0,5 мм), плохо заметным слоем. Некоторым исключением из этого правила является диоон колючий, толщина слоя древесины которого составляет около 1/3 диаметра ствола. И хотя кольца прироста у него выражены довольно отчетливо, образуются они далеко не каждый год, а иногда и очень редко - один раз в 10-20 и более лет под влиянием различных причин - смены кроны, семеношения и др. У некоторых представителей саговниковых, например у стангерии или замии, слои прироста различить вообще невозможно.
Деятельность камбия у таких родов, как саговник, макрозамия, бовения, а также у некоторых видов энцефаляртоса продолжается очень короткое время, а дальнейшее утолщение стебля вызывается у них новыми камбиальными кольцами, возникающими последовательно в коре и откладывающими дополнительные слои проводящих элементов. Эти добавочные камбиальные кольца (иногда дуги) появляются независимо друг от друга и не связаны онтогенетически с первым, если можно выразиться, "нормальным" камбием. Старые стебли некоторых таких саговниковых могут насчитывать иногда довольно большое число последовательных проводящих цилиндров, как бы продетых один в другой (рис. 166). В этом случае можно говорить о полицикличном типе ствола.

Рис. 166. Поперечный разрез полицикличного стебля энцефаляртоса Альтенштейна (Encephalartos altensteinii)
Камбий других представителей саговниковых (замии, цератозамии, диоона, микроцикаса и стангерии) долгие годы остается активным, создавая единственное проводящее кольцо, поэтому такие саговниковые называют моноцикличными.
Надо сказать, что моноцикличны молодые стебли всех саговниковых. Первичная проводящая система представляет собой слабо развитое кольцо коллатеральных пучков, отделенных друг от друга широкими сердцевинными лучами. От первичных сосудистых пучков под углом в 30° отходят листовые следы которые, пересекая кору, достигают листовых оснований.
У проростков саговниковых протоксилема состоит из трахеид со спиральными утолщениями, но в зрелых стеблях, где рост в длину сильно замедлен, нередко трахеиды в прото- и метаксилеме лестничные. Зрелая древесина большинства саговниковых представлена трахеидами с округлыми окаймленными порами, расположенными на радиальных стенках в 2-4, а иногда 5 рядов. Лишь трахеиды стангерии и замии имеют, подобно папоротникам, лестничную поровость. Саговниковые относятся к тем редким голосеменным, окаймленные поры которых лишены торуса.
Саговниковые отличаются большой длиной волокон, превосходя в этом отношении большинство древесных растений, за исключением араукариевых. Длина их трахеид в молодой древесине достигает 7-7,3 мм, а в зрелой - даже 10 мм. Лишь некоторые ископаемые, относящиеся к кордаитам и беннеттитам, могут сравниться с саговниковыми по длине трахеид.
Центральная часть стебля - сердцевина - занимает примерно 1/3 объема и состоит у всех саговниковых из крупных, тонкостенных, содержащих крахмал паренхимных клеток. В ней имеется самостоятельная система связанных друг с другом коллатеральных проводящих пучков. Каждый из этих пучков связан со слизевыми каналами, в изобилии пронизывающими сердцевину и содержащими водянистую слизь. Иногда их неправильно называют "смоляными ходами", хотя запах слизи совершенно не похож на терпентиновый. Высыхая, слизь становится твердой и прозрачной кристаллической массой.
С наружной стороны проводящего цилиндра располагается мощная кора, сообщающаяся с сердцевиной радиально идущими лучами, которые, переходя из древесины в кору, значительно расширяются. Так же, как сердцевина, кора состоит главным образом из паренхимных клеток, накапливающих крахмал, и содержит большое число слизевых каналов. Луб представлен множеством ситовидных элементов с рассеянными ситовидными полями и косыми конечными стенками. Пробковый камбий, образующийся в коре, наращивает довольно мощную перидерму. Это приводит к постепенному утолщению ствола, благодаря чему отмершие внешние ткани время от времени разрываются и, сшелушиваясь, отпадают.
Стебли саговниковых отличаются мягкой текстурой. Лишь некоторым из них наружную жесткость придают основания листьев, образующие панцирь. Мощная кора со слоями мертвых клеток, пропитанных суберином, служит саговниковым дополнительной механической опорой, надежность которой не может обеспечить слабо развитая древесина.
Саговниковые, обладающие большими массивами паренхимы в надземных органах, по-видимому, не способны длительно переносить низкие температуры, что является одной из причин отсутствия этих растений во флоре умеренных широт.
Корневая система
Корневая система саговниковых еще слабо изучена, и это неудивительно, поскольку речь идет об относительно редких в природе растениях. В сравнении с папоротниками саговниковые обладают более дифференцированными корнями. Именно они являются первой группой высших растений, для которой характерен появившийся в ходе эволюции главный (первичный) корень. Нередко он бывает таким же толстым, как стебель, и имеет веретеновидную форму. У одних саговниковых он короткий; у других достаточно длинный, достигая иногда 10-12 м. Постепенно суживаясь от основания и слабо ветвясь, главный корень углубляется в почву или растет близко к поверхности почти горизонтально.
Однако наблюдаются случаи, когда корневая система такого типа рано отмирает и заменяется придаточными корнями, возникающими из стеблевой ткани. Так, у саговника поникающего еще в стадии проростка, когда растению не более 1-2 лет, могут появиться один-два придаточных корня и добавляться по одному или по нескольку в каждый следующий год. У саговника 7-8-летнего возраста можно насчитать иногда до 17 больших мясистых корней, обладающих способностью сокращаться.
На кончики корней саговниковых, как наконечники, "надеты" массивные корневые чехлики, защищающие нежные меристематические ткани корня от механических повреждений.
Многослойная первичная кора корня состоит из тонкостенных живых паренхимных клеток, заполненных зернами крахмала и нередко содержащих танин. В них в большом количестве встречаются своеобразные полости - вместилища воздуха, недостаток которого постоянно испытывают ткани корня. Самый внутренний слой первичной коры - эндодерма - состоит из одного ряда клеток.
Под эндодермой, в периферической части стелы (центрального цилиндра), у саговниковых образуется многослойный перицикл, включающий до 10 рядов тонкостенных, долго сохраняющих меристематическую активность клеток (перицикл состоит из многих рядов клеток, даже в самых тонких корнях). В перицикле закладываются боковые корни, при участии перицикла осуществляется вторичное утолщение корня.
Большую часть стелы в корне занимает сложный проводящий пучок с двумя-тремя и более радиально вытянутыми ксилемными участками, чередующимися с участками флоэмы. В месте прикрепления главного корня к стеблю обычно имеется до восьми протоксилемных тяжей. Ближе к концу (верхушке) корни становятся диархными, т. е. имеющими два ксилемных участка (рис. 167). Боковые корни всегда диархны.
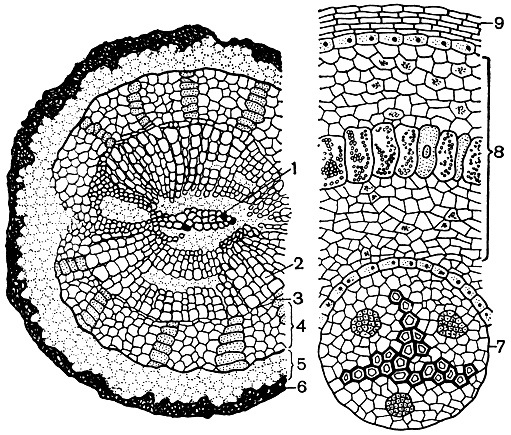
Рис. 167. Строение корней саговника поникающего (Cycas revoluta). Поперечный разрез утолщенного корня: 1 - первичная (диархная) ксилема; 2 - вторичная ксилема; 3 - камбий; 4 - вторичная флоэма; 5 - первичная кора; 6 - перидерма. Поперечный разрез коралловидного корня: 7 - центральный цилиндр с триархным пучком; 8 - первичная кора со слоем клеток, содержащих водоросли; 9 - перидерма
Из перицикла за счет энергичного деления его клеток по всей окружности корня появляется и перидерма. Когда первичная кора корня сшелушивается, наружный слой перидермы (пробка) принимает на себя защитные функции, оберегая корни от повреждений. В таких утолщенных корнях, особенно вблизи их основания, в паренхимных клетках перидермы располагаются слизевые каналы, выстланные изнутри секреторными клетками.
Среди голосеменных только некоторые саговниковые обладают контрактильными, или сокращающимися, корнями. При энергичном росте кончиков корней появляется опасность выталкивания молодых растений из почвы. Но их спасает то, что контрактильные корни периодически укорачиваются (при этом утолщаются) и как бы подтягивают, а иногда и полностью втягивают стебель молодого растеньица в почву, не только прочнее прикрепляя его к субстрату, но и создавая оптимальные условия для его роста и развития придаточных корней. Такая особенность могла развиться у саговниковых благодаря обилию в их корнях паренхимных тканей, клетки которых могут легко менять свою форму и объем.
Примечательной особенностью всех саговниковых являются растущие вверх над землей и дихотомически ветвящиеся коралловидные корни - кораллоиды. Они возникают как разветвления боковых корней эндогенно из многорядного перицикла напротив лучей первичной ксилемы. Благодаря интенсивному дихотомическому ветвлению коротких и тонких боковых корней образуются целые грозди клубеньков, окружающие ствол у его основания и напоминающие внешне кораллы. В кораллоидах, главным образом в протодерме, были обнаружены сине-зеленые водоросли (Nostoc punctiforme, Anabaena cycadae и другие виды), азотфиксирующие бактерии (Bacterium radicicola и Azotobacter sp.) и, наконец, гифы слабопатогенных грибов, образующих фикомицетную эндотрофную микоризу. Какой же из этих микроорганизмов вызывает образование клубеньков? Сначала считали, что это происходит под влиянием бактерий, проникающих в клетки коры корней. В последнее время высказывается предположение, что бактерии, как и сине-зеленые водоросли, являются вторичными поселенцами в сформировавшихся уже клубеньках, а сам клубенек представляет разрастание несущего его корня, вызванное эндофитным грибом, мицелий которого обильно заполняет межклетники в коровой паренхиме этого корня.
Многолетний клубенек может расти неограниченно долго. В нем обнаруживается дифференциация (рис. 167) на центральный цилиндр с диархным радиальным проводящим пучком и кору с хорошо выраженной эндодермой. Клетки коры богаты крахмалом (около 20% по сырой массе). Отдельные сильно разросшиеся клетки внешней части коры и рассеянные в паренхиме слизистые лакуны становятся местом развития сине-зеленых водорослей. В средней и внутренней частях коры, в так называемой "бактериоидной зоне", концентрируются бактерии.
Назначение корневых клубеньков саговниковых состоит прежде всего в усвоении атмосферного азота. Нельзя не связать с этим способность многих саговниковых мириться с крайне бедными азотом субстратами. С другой стороны, по предположению ряда ботаников, кораллоиды по функции аналогичны дыхательным корням некоторых хвойных и цветковых - пневматофорам.
Характер взаимоотношений всех названных микроорганизмов с растением-хозяином остается не вполне выясненным. Но как бы то ни было саговниковые представляют уникальный во всем растительном мире случай сожительства четырех и даже пяти различных организмов: самого растения-хозяина, гриба, двух видов бактерий и сине-зеленой водоросли. Этот симбиоз сложился, по-видимому, уже в глубокой древности.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'