Репродуктивные структуры саговниковых
Охарактеризованные выше вегетативные органы саговниковых являются органами спорофита, и соответственно на последнем при достижении зрелости формируются спорангии. Саговниковые - разноспоровые растения, т. е. они образуют отдельно мегаспорангии и микроспорангии. Более того, все представители этого семейства являются растениями строго двудомными: на одних особях каждого вида из года в год развиваются только мегаспорангии; на других - только микроспорангии (отдельные сообщения о якобы наблюдавшихся случаях изменения пола в онтогенезе одного растения требуют научного подтверждения).
И мега- и микроспорангии образуются на спорофите не поодиночке, а в большем или меньшем числе на спорофиллах (чешуях), в свою очередь собранных в компактные образования - стробилы.
От всех остальных родов резко отличается род саговник. У видов этого рода спорангии у женских растений развиваются на листовидных мегаспорофиллах прямо на стволе.
Стробилы
Стробилы саговниковых формируются на конце ствола среди листьев кроны. В середине стробилов проходит ось, на которой более или менее плотно расположены чешуевидные спорофиллы, число которых может быть очень велико - до 400 и даже до 600. У большинства саговниковых они расположены спирально и при этом налегают друг на друга, как черепица, но у видов рода замия мегаспорофиллы и микроспорофиллы размещены на оси мутовчато и примыкают друг к другу краями. Сами спорофиллы в большинстве случаев являются более или менее плоскими чешуями, но у видов той же замии они щитовидные, т. е. представляют собой шестиугольную пластинку на ножке, отходящей от оси шишки.
На нижней поверхности микроспорофиллов развиваются эллиптические или почти шаровидные микроспорангии; невооруженному глазу они представляются как мелкие зернышки (табл. 39). Располагаются они обычно не поодиночке, а, как и у папоротников, небольшими группами - сорусами - по 2, 3 спорангия (до 6) в каждом. При этом к каждому сорусу подходит один общий проводящий пучок. Виды саговника имеют наибольшее количество микроспорангиев (до 1000 и больше) на одной чешуе микростробила. Они равномерно покрывают всю нижнюю поверхность микроспорофилла. Самое малое количество микроспорангиев свойственно видам замии. Так, у замии флоридской на отдельных микроспорофиллах (из верхней и нижней частей стробила) наблюдали всего по одному микроспорангию по обе стороны от средней стерильной части чешуи.

Таблица 39. Саговниковые: вверху слева - бовения мелкопильчатая (Bowenia serrulata), справа на снимке молодой лист; вверху справа - лепидозамия Перовского (Lepidozamia peroffskiana) в Ботаническом саду в Сиднее (Австралия); внизу - энцефаляртос Альтенштейна (Encephalartos altensteinii) там же
Мегаспорофиллы в этом отношении значительно однообразнее. Как правило, в их основании развиваются по два, но крупных семязачатка (табл. 40). Впрочем, и здесь имеются исключения, на этот раз в родедиоон. У диоона колючего, например, их иногда может быть 3-4 и даже 5.
Число образуемых одним растением стробилов в известной степени зависит от способа нарастания стебля. При симподиальном нарастании часто образуется всего один стробил, реже 2 или 3. Один огромный мегастробил венчает, например, ствол микроцикаса. При боковом заложении стробилов на моноподиально нарастающем стволе одновременно может сформироваться их большое число, особенно у мужских растений,- до 10 у некоторых видов энцефаляртоса и макрозамии и даже более ста (!) у макрозамии Мора (Macrozamia moorei).
Стробилы саговниковых обычно крупные, а у некоторых видов и огромные. Таковы мегастробилы энцефаляртоса кафрского (табл. 40) и лепидозамии Перовского (длиной до 1 м) микроцикаса (до 95 см) и т. д. Микростробилы обычно мельче. У бовении, например, они в два раза короче, чем мегастробилы. Однако у некоторых саговниковых и микростробилы бывают длинными, например более метра у энцефаляртоса Вуда (Encephalartos woodyi), до 90 см у микроцикаса и т. д. Самые мелкие стробилы (длиной всего 2 см, а массой около 30 г) характерны для женских и мужских растений замии карликовой (рис. 180).
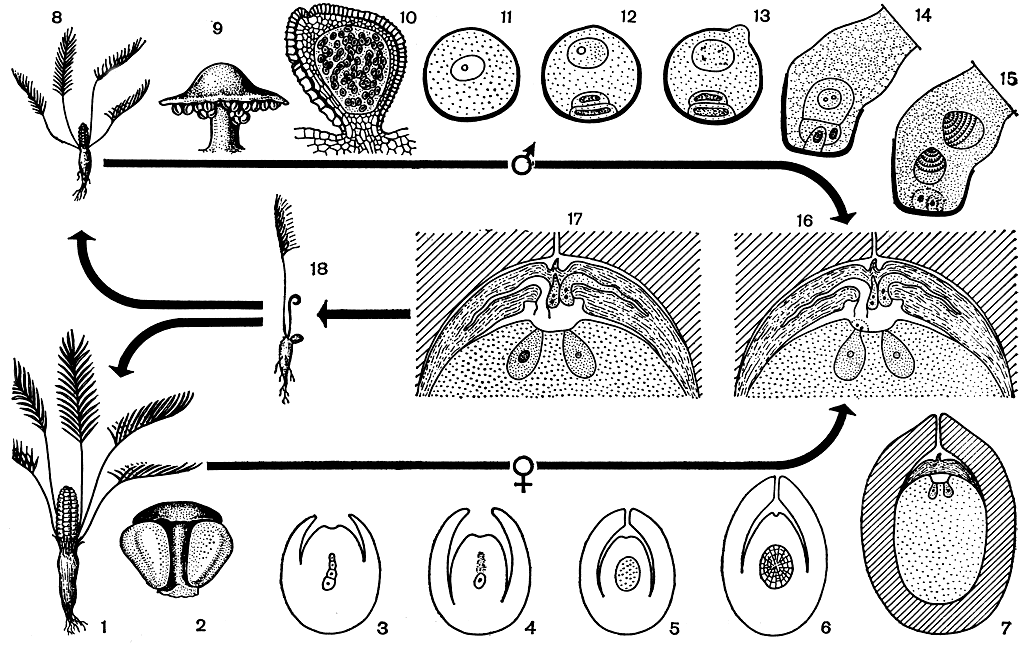
Рис. 168. Жизненный цикл замии (Zamia): 1 - женское растение; 2 - мегаспорофилл с двумя семязачатками; 3 - образование линейной тетрады мегаспор; 4 - семязачаток с единственной сохранившейся мегаспорой; 5 - образование свободноядерного женского гаметофита; 6 - образование клеточного гаметофита; 7 - появление архегониев; 8 - мужское растение; 9 - микроспорофилл; 10 - микроспорангий; 11 - микроспора; 12 - образование трехклеточного мужского гаметофита в микроспоре; 13 - прорастание клетки-гаустории; 14 - образование сперматогенной клетки; 15 - формирование сперматозоидов; 16 - верхняя часть семязачатка перед оплодотворением; 17 - то же после оплодотворения одной (левой) из яйцеклеток; 18 - проросток
Мега- и микростробилы отличаются и по форме. Как правило, микростробилы более узкие, вытянутые в длину. Лишь в редчайших случаях (один из видов энцефаляртоса) стробилы женских и мужских растений почти неотличимы как по размерам, так и по форме. Однако во всех случаях те и другие отличаются по массе. Мегастробилы с плотно упакованными под чешуями семенами намного тяжелее микростробилов с их мелкими микроспорангиями и многочисленными воздушными полостями между чешуями.
Стробилы некоторых саговниковых, достигая массы 45-50 кг (энцефаляртосы поперечно- жильчатый и кафрский), являются самыми крупными и тяжелыми "шишками" во всем растительном мире. Цвет стробилов то невзрачный - зеленый, сизо-зеленый (некоторые виды макрозамии) или коричневатый (виды замии), то яркий - желтый, оранжевый (виды энцефаляртоса, табл. 40). Нарядно выглядят стробилы энцефаляртоса неожиданного (Encephalartos inopinus), чешуи которого густо покрыты мелкими сосочками (папиллами), придающими поверхности этих "шишек" серебристый оттенок.

Таблица 40. Мегастробилы и мегаспорофиллы видов рода энцефаляртос (Encephalartos): 1 - энцефаляртос умбелузийский (Encephalartos umbeluziensis); 2 - энцефаляртос Альтенштейна (Е. altensteinii); 3 - энцефаляртос кафрский (Е. caffra)
Общее происхождение различных групп современных саговниковых и их строгая двудомность позволяет думать, что и далекие предки этих растений были двудомными. Столь давнее разделение полов не могло не отразиться на особенностях строения не только репродуктивных, но и вегетативных структур женских и мужских растений. Отмечено, например, что стволы у мужских особей микроцикаса выше, чем у женских. Предполагают, что это облегчает опыление. Стволы мужских растений цикаса завитого отличаются большей, чем у женских экземпляров, толщиной.
Но самые значительные отличия между мужскими и женскими растениями относятся все же к репродуктивным органам, и в этом отношении род саговник занимает совершенно особое положение в семействе, поскольку у его представителей не образуется характерных для всех остальных родов компактных мегастробилов.
Мегаспорофиллы видов саговника
Мегаспорофиллы саговников, особенно у саговников завитого, поникающего и сиамского, похожи на вегетативные листья, но значительно мельче их (длиной не более 40 см). Их нижняя часть представляет как бы черешок, а расширенная верхняя - редуцированную листовую пластинку, у одних видов зубчатую или гребенчатую, у других глубоко-перистораздельную. И "черешок", и "пластинка" мегаспорофиллов лишены хлорофилла и густо покрыты желтоватыми волосками (табл. 38), чем напоминают по виду спорофиллы некоторых папоротников.

Таблица 38. Саговниковые: вверху слева - саговник поникающий (Cycas revoluta) в Ботаническом саду в Сиднее (Австралия); вверху справа - саговник завитой (С. circinalis) в оранжерее Ботанического института АН СССР в Ленинграде, собрание мегаспорофиллов (до их развертывания); внизу - то же растение с развернувшимися мегаспорофиллами
Под "пластинками" мегаспорофиллов, на "черешках", как их боковые выросты, попарно или в очередном порядке развиваются семязачатки. Число их у разных видов саговника различно - от 7 у саговника Туара, 5-6 у саговника поникающего до всего одной пары у ряда австралийских видов этого рода.
Мегаспорофиллы появляются в большом числе, окружая со всех сторон вегетативную точку роста, венчающую ствол женского растения. Первоначально эти спирально расположенные спорофиллы прикрывают верхушку стебля, образуя подобие капустного кочана или рыхлой шишки (табл. 38). Затем они отгибаются, образуется ярко окрашенный "воротничок", заметный издали сквозь воронку из листовых черешков (табл. 38). По мере созревания мегаспорофиллы еще более склоняются вниз и, наконец, повисают вдоль ствола. Выполнив свою функцию, они засыхают и опадают каждый по отдельности.
Таким образом, начиная развертываться в разное время, кроны ассимилирующих листьев и "кроны" мегаспорофиллов чередуются (в своем возникновении) на моноподиально нарастающем стволе видов рода саговник.
Семязачаток
Семязачатки саговниковых отличаются по величине (длиной от 5-6 см у некоторых видов саговника до 5-7 мм у замии карликовой) и по форме. Но при этом они довольно однотипны в основных чертах развития и внутреннего строения. Открыто сидящие на "черешке" листовидного мегаспорофилла у саговников, висячие на нижней стороне щитовидной чешуи у замий, прикрытые более или менее плоской чешуей мегастробила у других родов, семязачатки всегда состоят из нуцеллуса (ядра семязачатка) и одевающего его покрова - интегумента. Толстый покров этот прочно срастается с нуцеллусом, отходя от него лишь на верхушке семязачатка. Здесь, в центре образованного интегументом конусообразного выступа, имеется отверстие - микропиле, а под ним полость - пыльцевая камера.
Главной частью семязачатка является нуцеллус. Это и есть собственно мегаспорангий, который, в отличие от открытых микроспорангиев саговниковых, как и у других голосеменных, заключен в защитный покров.
Образовавшаяся в нуцеллусе в результате мейоза единственная функционирующая мегаспора быстро растет за счет трех остальных, вскоре отмирающих спор тетрады и окружающих клеток нуцеллуса и вырабатывает хорошо выраженную двуслойную оболочку, как это свойственно спорам бессеменных высших растений, переносимым токами воздуха. Но мегаспора саговниковых никогда не покидает мегаспорангия, и названная особенность ее строения сохранилась как пережиток прошлого, унаследована от далеких предков, расселявшихся с помощью спор. У саговников наружная оболочка мегаспоры в дополнение пропитывается кутином, что, очевидно, придает ей еще более архаичный характер.
Итак, спорогенез завершен. К этому времени интегумент разросшегося семязачатка уже дифференцирован на три слоя - мясистые наружный и внутренний и твердый средний, состоящий из омертвевших клеток. Оба мяси-стых слоя пронизаны целой системой проводящих пучков, обеспечивающих разрастающийся семязачаток необходимыми питательными веществами.
Образовавшаяся в нуцеллусе мегаспора тут же и прорастает, образуя женский гаметофит. Процесс этот образно описан еще Ч. Чемберленом. Вслед за первым делением ядра споры происходят многие повторные ядерные деления, при этом не образуется клеточных перегородок (рис. 168,5). Многочисленные свободные ядра (их число, например у диоона, может доходить до тысячи) оказываются в тонком постенном слое цитоплазмы. В дальнейшем начинают обособляться клетки гаметофита, сперва по периферии, затем все ближе и ближе к центру мегаспоры, пока вся она не заполняется многоклеточной тканью (рис. 168, 6). По общему объему и по числу составляющих клеток женский гаметофит саговниковых не уступает даже наиболее крупным свободно живущим гаметофитам (заросткам) папоротников.
Женский гаметофит развивается как запасающая ткань (первичный эндосперм). Разрастаясь, он вытесняет нуцеллус (рис. 168,7), затем потребляет вещества из внутреннего мясистого слоя интегумента, пока этот слой не превращается в тонкую пленку ковнутри от твердой "скорлупы". Клетки эндосперма постепенно заполняются запасными материалами, среди которых преобладает крахмал (до 65-70% в пересчете на сухую массу у видов макрозамии). Накапливаются в эндосперме также жирные масла (у саговника поникающего их содержание может доходить до 23%), а также белки.
Наконец, в клетках женского гаметофита обнаруживаются лейкопласты. Хотя женский гаметофит саговниковых и утерял в ходе эволюции возможность самостоятельного существования вне семязачатка, он удивительным образом сохранил способность к развитию хлорофилла (позеленению) на свету при извлечении из мегаспорангия. Наблюдали превращение лейкопластов в хлоропласты и на семязачатках, в которых не произошло оплодотворение. В этом случае гаметофит прорастал через микропиле и его выступающий конец зеленел на свету.
В этом отношении интересны опыты по выращиванию эксплантатов (кусочков ткани) от женского гаметофита саговниковых в стерильной культуре. В зависимости от соотношения в питательной среде различных стимуляторов роста растущая клеточная масса гаметофита образует то корни, то стеблевые почки, то, наконец, зародышеподобные образования (эмбриоиды), т. е. проявляет особенности морфогенеза, свойственные спорофиту.
В верхней части нормально сформировавшегося женского гаметофита под микропиле развиваются женские половые органы - архегонии (рис. 168, 7, 16). Материнских клеток архегониев закладывается много, и это еще одна архаическая черта саговниковых. Правда, вполне развитых архегониев у них оказывается обычно не более десяти. Совершенно исключителен в этом отношении микроцикас, у которого образуется несколько десятков архегониев.
Яйцеклетка в архегонии достигает огромных размеров (у микроцикаса она длиной до 6 мм). Необычайно велико и ядро яйцеклетки; будучи иногда диаметром до 500 мкм, оно видимо простым глазом, как точка. Напротив, шейка архегония маленькая и состоит обычно из двух мелких клеток, которые к моменту оплодотворения ослизняются, открывая доступ к яйцеклетке. К этому времени между микропиле и верхней частью гаметофита оказывается довольно большая полость (пыльцевая и архегониальная камеры при прорыве оболочки мегаспоры сливаются), куда и попадают переносимые ветром пыльцевые зерна.
Образование микроспор
Микроспорангий саговниковых по внешнему виду и внутреннему строению значительно меньше отличается от микроспорангиев папоротниковидных, чем это проявляется у мегаспорантиев.
Развитие микроспорангия происходит так же, как у эвспорангиатных папоротников, например у мараттиевых, т. е. он возникает не из одной, а из группы инициальных клеток. Сформировавшийся микроспорангий состоит из "головки" на короткой массивной ножке (рис. 168, 10).
Покрывающий головку эпидермальный слой клеток видоизменен в своеобразную механическую ткань и состоит из омертвевших клеток с утолщенными со всех сторон, кроме верхней (наружной), стенками. Как и у папоротников, с наличием таких клеток связан механизм раскрывания спорангия. Из всех голосеменных саговниковые являются единственной группой с "папоротниковым" типом строения стенок микроспорангия.
Интересна и другая особенность эпидермы микроспорангиев. У многих представителей семейства (за исключением саговника и диоона) в эпидерме оказались устьица, такие же, как у листьев соответствующих видов. Это редкое явление свойственно лишь единичным представителям других групп высших растений - некоторым моховидным, немногим папоротникам.
Полость микроспорангия, выстланная очень мелкими клетками однослойного тапетума, заполнена многочисленными материнскими клетками микроспор, которые в ходе двух делений мейоза превращаются в тетрады микроспор (рис. 168, 11).
Еще находясь в микроспорангии, микроспоры начинают прорастать. При этом, как и в случае с мегаспорой, внешне это прорастание не проявляется; формирующийся мужской гаметофит остается заключенным в оболочку споры. Микроспора, точнее, пыльцевое зерно покидает микроспорангий с трехклеточным гаметофитом (рис. 168, 12), причем вся вегетативная его часть предельно редуцирована и представлена всего одной вегетативной клеткой. Антеридиальная клетка, разделившись, дает начало генеративной клетке и клетке-гаустории.
Пыльцевые зерна у всех саговниковых мелкие, широкоэллиптические, лодочковидные и имеют на дистальной стороне одну тонкую борозду (табл. 42). У некоторых родов (лепидозамия, энцефаляртос) сохранилась еще чрезвычайно примитивная, состоящая целиком из спорополленина, эктэкзина, свойственная также семенным папоротникам и беннеттитам. В то же время строение спородермы у многих саговниковых весьма специализировано. Для большинства изученных видов характерна удлиненно-ячеистая эктэкзина. В ней имеется, таким образом, много свободных от спорополленина мест. Это уменьшает массу пыльцевого зерна, способствуя переносу пыльцы ветром. Ячеистое строение эктэкзины и утончение экзины на проксимальной стороне облегчает изменение объема пыльцевого зерна, что весьма существенно при образовании многоклеточного мужского гаметофита внутри оболочки микроспоры. Из ныне живущих голосеменных такого типа спородерма характерна только для саговниковых.
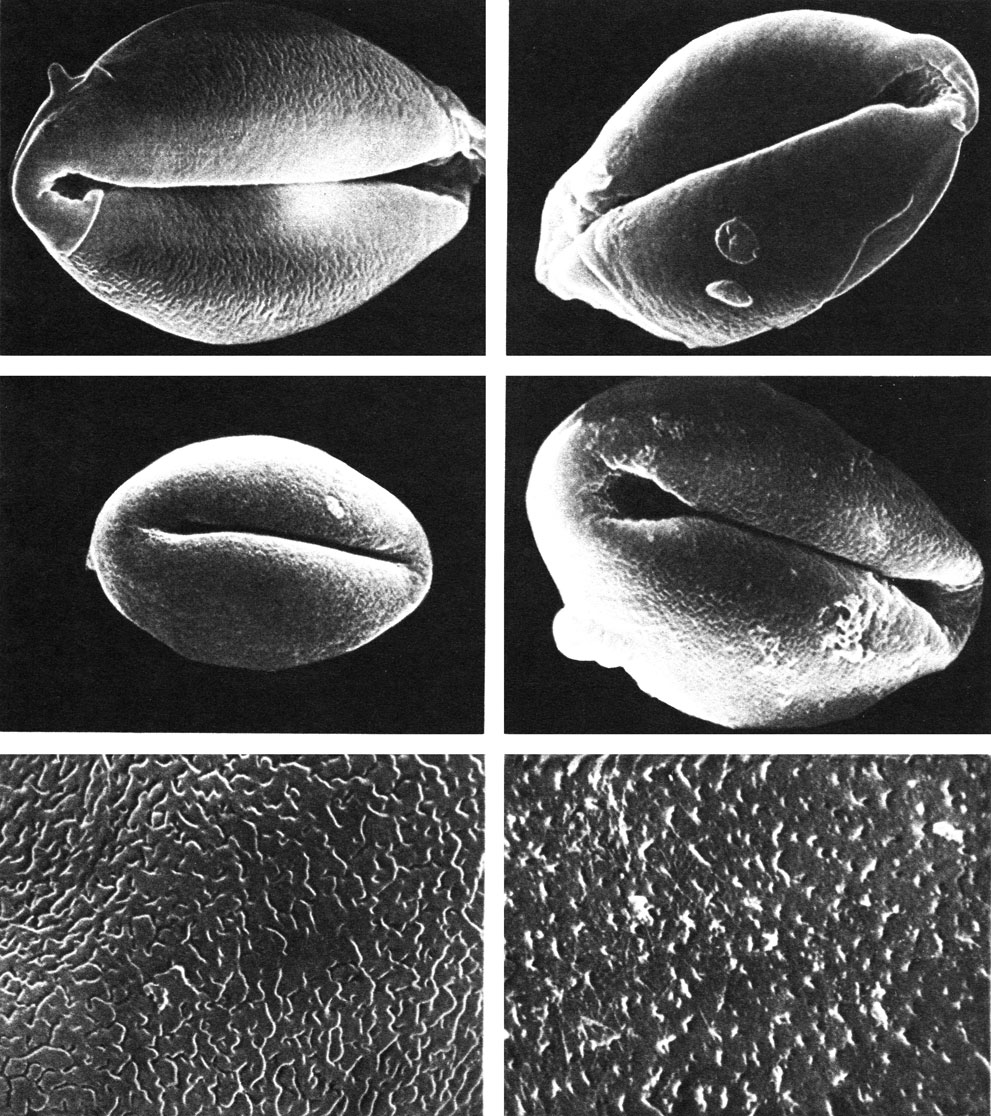
Таблица 42. Пыльцевые зерна саговниковых под сканирующим электронным микроскопом (увел, около 8000): вверху слева - лепидозамия Перовского (Lepidozamia peroffskiana); вверху справа - энцефаляртос Альтенштейна (Encephalartos altensteinii); в середине слева - стангерия шерстистая (Stangeria eriopus); в середине справа - цератозамия мексиканская (Ceratozamia mexicana). Участки поверхности пыльцевых зерен (увел, около 20 ООО): внизу слева - лепидозамия Перовского (Lepidozamia peroffskiana); внизу справа - цератозамия мексиканская (Ceratozamia mexicana)
Саговниковые, как и все другие современные голосеменные, являются растениями ветроопыляемыми. Пыльца из микростробилов мужских особей переносится на мегастробилы женских токами воздуха. К моменту созревания мегастробилов их ось несколько удлиняется и между мегаспорофиллами, до того плотно прижатыми друг к другу, образуются щели, сквозь которые к семязачаткам попадает приносимая ветром пыльца. Одним из приспособлений к ветроопылению является образование очень большого количества пыльцы. Подсчитано, что у одного из видов энцефаляртоса в среднем образуется в каждом микроспорангии 26 тыс. микроспор, на каждом микроспорофилле 618 микроспорангиев, а в микростробиле - 404 микроспорофилла. Таким образом созревший микростробил энцефаляртоса выбрасывает в атмосферу около 7 млрд. (!) очень мелких и легких пыльцевых зерен. Строгая двудомность саговниковых также является одной из черт, свойственных многим ветроопыляемым растениям.
Возможно, у некоторых саговниковых имеет место и насекомоопыление, а именно опыление жуками (кантарофилия). Это относится, в частности, к видам энцефаляртоса и саговника. Уже давно отмечено, что стробилы энцефаляртосов Альтенштейна, вильчатого, ощетиненного, кафрского и других в период их "цветения" постоянно посещаются различными насекомыми, особенно жуками из долгоносиков. Этих посетителей может привлекать и сильный и обычно неприятный для обоняния человека запах микростробилов у видов энцефаляртоса, и яркий (желтый, оранжевый) цвет их мегастробилов (табл. 40), и, наконец, образуемая в изобилии пыльца.
Возможно, какую-то связь с кантарофилией имеет и так называемый "тепловой феномен". Значительное повышение температуры внутри созревших микростробилов по сравнению с температурой окружающего воздуха отмечали не только у энцефаляртоса Альтенштейна, но и у представителей рода саговник, макрозамия и цератозамия. Вопрос о кантарофилии у саговниковых остается спорным и требует специальных исследований точными методами.
Там, где совместно произрастают несколько различных видов саговниковых, обычен занос пыльцы с мужского растения одного вида на женский экземпляр другого. Возможна ли при этом гибридизация? Такие факты известны. Сообщают об очевидной гибридной природе некоторых дикорастущих саговниковых, например ряда представителей макрозамии и энцефаляртоса. Проводились также опыты по искусственной межвидовой и даже межродовой гибридизации, в частности, по скрещиванию видов замии и энцефаляртоса, цератозамии и замии. Выращенные из гибридных семян растения проявляли некоторые признаки того и другого родителя. В ботанических садах гибриды образуются и спонтанно. Подчас этим бывает затруднено точное определение выращиваемых саговниковых.
Каким бы путем ни попала пыльца с мужского растения на женское, процесс опыления завершается очень своеобразно. К моменту опыления у созревших семязачатков из микропиле выступает капелька опылительной жидкости, которая улавливает мелкие микроспоры. Позднее эта капелька слизи вместе с захваченными микроспорами всасывается через микропиле в пыльцевую камеру, а ее остающаяся часть, подсыхая, заклеивает пыльцевход.
Оплодотворение
Процессы, связанные с оплодотворением у саговниковых, настолько необычны, что их выявление, особенно открытие у них в конце прошлого века сперматозоидов, произвело среди специалистов настоящую сенсацию. Процессы эти проходят в следующей последовательности.
После опыления семязачатки начинают увеличиваться и скоро достигают размеров семени, хотя оплодотворение в них еще не произошло. Этот период, от опыления до оплодотворения, весьма длителен и занимает обычно полгода (так, у саговника завитого опыление происходит в декабре - январе, а оплодотворение - в мае - июне).
Микроспоры, попавшие с капелькой опылительной жидкости в пыльцевую камеру, прорастают. Экзина при этом лопается, и через разрыв прорастает, растягивая интину, клетка- гаустория (рис. 168, 13). Она внедряется в стенку пыльцевой камеры и высасывает из ткани нуцеллуса питательные вещества (рис. 168, 13). Генеративная клетка в это время делится на две, и одна из образовавшихся клеток - сперматогенная - начинает усиленно расти. В ней и формируются, не сразу, а спустя несколько месяцев, мужские гаметы - сперматозоиды (рис. 168, 15).
В большинстве случаев образуются два сперматозоида. Однако у цератозамий иногда их образуются четыре. Микроцикас и в этом отношении является удивительным исключением: у него в одной проросшей микроспоре образуется не менее 16 (и до 22) сперматозоидов.
У саговниковых сперматозоиды являются самыми крупными мужскими гаметами, известными науке. У замии флоридской их длина достигает 325 мкм, и они видны даже невооруженным глазом.
Сперматозоиды саговниковых имеют кубаревидную форму и опоясаны от середины до переднего конца спиральной лентой блефаропласта - носителя коротких жгутиков, которых у каждого сперматозоида, например замии, примерно 20 тыс.
При электронно-микроскопическом изучении микроструктура этого локомоторного аппарата оказалась очень сложной, обеспечивающей не только поступательное, но и вращательное (эвгленоидное) движение сперматозоида.
К моменту оплодотворения разросшаяся сперматогенная клетка оказывается в непосредственной близости от входа в архегоний. Освободившимся из нее сперматозоидам остается "проплыть" в жидкости, которая излилась вместе с ними из сперматогенной клетки, лишь небольшое расстояние до архегония, в котором происходит слияние содержимого сперматозоида с яйцеклеткой (рис. 168, 17).
Таким образом, у саговниковых в едином процессе соединяются два механизма, один из которых - образование подвижного сперматозоида - свойствен далеким оплодотворявшимся с помощью воды предкам, а второй - формирование пыльцевой трубки (разрастающаяся сперматогенная клетка) - типичен для всех остальных стоящих выше на "эволюционной лестнице" семенных растений.
После оплодотворения зигота - первая клетка нового спорофита - приступает к делению. При этом, как и при прорастании мегаспоры, сначала идет свободное ядерное деление без образования клеточных перегородок. Число ядер в разросшейся зиготе может достигать нескольких сот и даже тысячи, когда, наконец, по прошествии значительного времени складывается многоклеточная структура так называемого предзародыша. В дальнейшем базальные клетки последнего путем многократных делений образуют все удлиняющийся подвесок (суснензор), проталкивающий предзародыш в запасающую ткань эндосперма. Суспензоры у саговниковых достигают иногда длины 5 см (у диоона съедобного даже до 7 см) и, таким образом, являются самыми длинными из всех известных у растений.
К моменту полного созревания семени и его опадения с материнского растения зародыш на конце длинного суспензора остается маленьким и недифференцированным и только на его конце намечаются небольшие выступы - зачатки будущих семядолей.
Семя
Семена саговниковых крупные. Эллиптические, продолговато-яйцевидные или шаровидные по форме, они обычно имеют длину 3-4 см при толщине 2-3 см. Но отдельные виды имеют или более мелкие, или более крупные семена. Так, семена замии карликовой не длиннее сантиметра (их толщина всего 7 мм), а огромные семена некоторых макрозамий и саговников могут достигать в длину 8 см. Мощная семенная
кожура у них двуслойная. Под наружной пленчатой оболочкой семени располагается наружный мясистый слой, или саркотеста, а за ней следует твердый, как косточка, слой из толстостенных клеток - склеротеста (внутренний мясистый слой семязачатка ко времени созревания семени разрушается).
Окраска семян часто более или менее яркая, красная, желтая, оранжевая. Эта особенность многих саговниковых, по-видимому, является результатом их давнего приспособления к распространению животными. Наблюдения показывают, что многие дикие животные охотно используют в пищу сочные семена саговниковых. При этом в большинстве случаев они поедают только мясистую саркотесту и не повреждают склеротесту, не трогая, таким образом, самого семени с его обильным питательным, но в то же время и токсичным содержимым.
Семена диоона съедобного (Dioon edule) поедают медведи и дикие свиньи (пекари), а энцефаляртоса и стангерии - обезьяны. По словам Чемберлена, найти нетронутые зрелые мегастробилы видов энцефаляртоса очень трудно; обезьяны разрывают шишки и выбирают из них семена еще до их созревания. С обезьянами соперничают в этом отношении грызуны и даже слоны. В Австралии в дуплах деревьев находили "запасы" семян макрозамии спиральной. Выше уже отмечалась возможность переноса семян видов саговника с одного острова на другой летучими лисицами.
Архаичной чертой семян саговниковых является формирование мощной дифференцированной семенной кожуры и обильной запасающей ткани эндосперма, еще до процесса опыления и тем более до оплодотворения. Вот почему здесь нет четкой внешне заметной разницы между семязачатком и семенем. Материнское растение при этом затрачивает питательные материалы на формирование всех семязачатков, хотя лишь часть из них, иногда незначительная, будет опылена и даст в конечном итоге семена.
Другая черта примитивности семян саговниковых заключается в недоразвитии зародыша в морфологически и физиологически зрелом семени. Когда семя отделяется от растения и опадает на землю, зародыш в нем еще слабо дифференцирован. Доразвитие зародыша происходит в отделившемся семени в течение длительного времени за счет запасных веществ эндосперма. При этом сильно удлиняются семядоли, выполняющие функцию всасывающего органа зародыша. Так, у одного из энцефаляртосов семядоли вначале составляют всего около 1/3 длины маленького зародыша, а ко времени прорастания достигают 9/10 его длины; в целом зародыш увеличивается в длину в несколько раз.
Эти замечательные особенности семян саговниковых уже давно привлекали внимание ученых и послужили поводом характеризовать эту группу вместе с гинкго (с таким же типом семени), как "яйцекладущие" растения в противовес "живородящим", каковыми с этой точки зрения может быть названо большинство современных семенных растений с вполне развитым ко времени зрелости семени зародышем.
В готовом к прорастанию семени саговниковых находится дифференцированный и вытянувшийся во всю длину эндосперма зародыш чаще всего с двумя, но у некоторых родов с одной, тремя или даже шестью семядолями.
У большого числа представителей семейства семядоли срастаются в основании в семядольную трубку; у многих (виды саговника, диоона, энцефаляртоса, замии и др.) они имеют неравное развитие: одна семядоля значительно короче другой или (у других видов) совсем не развита, хотя в проводящей системе зародыша обнаруживаются следы ее былого наличия. При проращивании таких односемядольных семян на клиностате, устраняющем действие силы тяжести, развиваются обе семядоли, притом одинаковые.
Период от рассеивания семян до их прорастания в большинстве случаев продолжителен, например, более года у макрозамии Ридла (Масrоzamia riedlei), два года у диоона съедобного и т. д. Но при неизменно благоприятных условиях, в теплицах, срок этот, очевидно, сокращается. Так, сообщают, что семена саговника завитого проращивают за 4 месяца, а замии кремнистой (Zamia silicea) - даже за 2-3 недели. Так или иначе, но настоящего, так называемого органического покоя семена саговниковых, по-видимому, не имеют, и развитие этих растений, по словам Чемберлена, происходит непрерывно от момента оплодотворения до старости и смерти. Наличие индуцированного покоя (покоя, вызванного неблагоприятными условиями) явилось в процессе эволюции как бы предпосылкой, первой ступенью в формировании органического покоя, столь характерного для семян эволюционно более подвинутых семенных растений.
К моменту прорастания семени наряду с формированием крупных семядолей развивается и ось зародыша. На стеблевом полюсе дифференцируется почечка, и на ней появляются выросты - два-три зачатка первых чешуйчатых листьев, в некоторых случаях и зачаток первого перистого листа. На коротком корневом полюсе, как раз под семядолями, рано образуется вздутие; постепенно вокруг слабо развитого зародышевого корня образуется прочное пленчатое влагалище - колеориза, защищающее его нежные ткани. При прорастании семени первой в образовавшийся разрыв склеротесты проходит именно колеориза; растущий корень проходит сквозь нее и углубляется в почву.
Для всех саговниковых характерно подземное прорастание (рис. 168, 18). Сущность его не в том, что прорастающее семя остается глубоко в почве (семя саговниковых, как правило, прорастает у ее поверхности), а в том, что семядоли до конца остаются в семени, обеспечивая поглощение запасных веществ из эндосперма и передачу их развивающемуся проростку. При этом семя может держаться на растении, продолжая его питать в течение года или даже двух лет. После выполнения этой функции опустошенное семя вместе с засыхающими семядолями отделяется от молодого растеньица, ставшего совершенно самостоятельным. Такой подземный, или скрытосемядольный, тип прорастания является очень древним и, можно сказать, исходным у семенных растений.
Очень медленно протекает и дальнейшее развитие проростков. Второй лист в природе обычно появляется лишь спустя год, последующие - по одному за год или даже за несколько лет. По достижении 5-6-летнего, а иногда и большего возраста впервые они появляются в виде "кроны" (у замии свойство поочередного развертывания листьев остается на всю жизнь).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'