Порядок схизейные (Schizaeales)
Семейство схизейные (Schizaeaceae) (Т. Н. Попова)
Схизейные - травянистые, реже лиановидные папоротники, преимущественно тропические и субтропические. Лишь немногие виды встречаются в умеренных областях Северной Америки и Японии, Чили, Новой Зеландии, Тасмании и Южной Африки. Схизейные, несомненно, древняя группа, история ее началась с каменноугольного периода. Благодаря наличию многих примитивных признаков схизейные считаются одним из самых своеобразных семейств папоротников, привлекающим постоянное внимание ботаников-систематиков, палеоботаников и морфологов.
В пределах одного семейства схизейных можно наблюдать разные типы строения проводящей системы корневища - от протостелы до сифоностелы и диктиостелы, различные типы ветвления жилок листьев и переход от дихотомически расчлененных листьев к симподиальному и перистому типу.
Современных родов схизейных пять: лигодиум (Lygodium), насчитывающий примерно 45 видов, схизея (Schizaea)- около 30, актиностахис (Actinostachys) - 13, анемия (Anemia) - 90 и мория (Mohria) - 2 вида. Эти роды столь своеобразны, что рассматриваются (за исключением лишь актиностахиса) некоторыми исследователями и как отдельные семейства.
Для всех родов схизейных характерны одиночные спорангии с поперечным, почти верхушечным кольцом и специфические споры, признаки которых позволяют с большой точностью определить их родовую принадлежность по рассеянным спорам, находимым в ископаемом состоянии. Особенно обильны находки спор и реже отпечатков листьев на севере Евразии, где современные их представители полностью отсутствуют, и в Северной Америке, где до настоящего времени сохранилось лишь несколько видов. Значительно меньше палеоботанических находок в пределах основной части современного ареала семейства.
Внешняя морфология спорофита очень различна у современных схизейных: от огромных лиановидных лигодиумов (длиной до 30 м) до схизей высотой в несколько сантиметров и таких же миниатюрных актиностахисов, у которых как по размерам, так и по продолжительности жизни может даже доминировать гаметофит. Спорофит актиностахиса иногда представляет собой очень короткое подземное корневище с узким неразделенным листом двухсантиметровой длины.
Стебли или корневища схизейных прямостоячие (схизея, актиностахис, мория, большинство видов анемии) или стелющиеся (часть видов анемии и лигодиума), иногда подземные, короткие или удлиненные, дихотомически разветвленные (лигодиум). Корневище может быть радиально-симметричным (у прямостоячих форм) или дорсивентральным, как у лигодиума, листья которого расположены двумя рядами на верхней поверхности корневища, а корни главным образом на нижней.
Молодые листья и основания их черешков покрыты волосками или чешуями (мория). У малезийских видов лигодиума волоски на остановившихся в росте концах стержня листа имеют вздутое основание, сформированное из большого числа клеток (например, лигодиум ушковидный - Lygodium auriculatum). У анемии корневища обычно густо покрыты красными или буроватыми волосками, а листья шелковистоопущенные.
Отличия в облике у различных родов обусловлены прежде всего разнообразием формы и строения листьев. Листья бывают простыми или чаще рассеченными (иногда веерообразно - у схизеи изящной - Schizaea elegans, рис. 103) от дихотомических до перистых. Их жилки обычно свободные, но у некоторых видов лигодиума и анемии могут сливаться у края. Черешок прямостоячий, тонкий, у актиностахиса и схизеи нередко узкокрылатый.

Рис. 103. Схизея изящная (Schizaea elegans), спорангиеносная часть листа
Спороносные листья обычно мало отличаются от вегетативных. Исключение - анемия тысячелистная (A. millifolia), листья которой резко диморфны.
Фотосинтезирующая пластинка листа схизеи и актиностахиса редуцирована до узкога крыла, несущего один (редко 2) правильный ряд устьиц на нижней поверхности. На листьях нередко имеются двуклеточные железистые волоски, конечные клетки которых при старении или высыхании сморщиваются или отваливаются, а базальные сохраняются, образуя маленькие "бородавки".
Спорангиеносные зоны расположены латерально вблизи верхушки листа или его разветвлений (рис. 103, 104). Многочисленные спорангии сближены или покрыты выростом ткани листа (индузиальным флагом) по всей длине каждого спороносного пера. Кроме того, противоположные ряды спорангиев частично защищают друг друга. У актиниостахиса индузиальный флаг развит лучше, чем у схизеи, фертильных перьев три или (из-за последующих глубоких дихотомических делений) гораздо больше. Они почти одинаковой длины, несут значительно больше спорангиев и отходят радиально от места прикрепления, чему и обязано происхождение родового названия Actinostachys (от греч. aktinos - луч и stachys-колос).
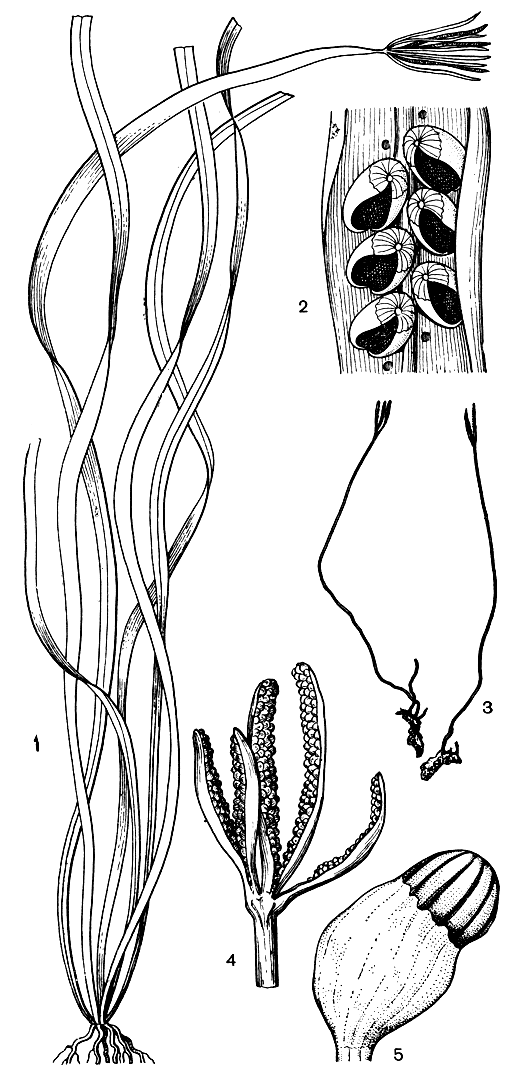
Рис. 104. Актиностахисы. Актиностахис пальчатый (Actinostachys digitata): 1 - общий вид; 2 - фрагмент спорангиеносной части листа. Актиностахис крупнобазальный (A. macrofunda): 3 - спорофиты с прикрепленными гаметофитами; 4 - спорангиеносный аппарат; 5 - спорангий
У первого из ставших известным ботаникам вида схизей - схизеи вильчатой (Schizaea dichotoma) - наблюдается многократное (2-8 раз) дихотомическое расщепление пластинки листа. Отсюда ее родовое название (от греч. schizo - расщеплять), которое иногда переводилось на русский язык как "разрывница".
Листья лигодиума, нередко ошибочно принимаемые за стебли, вследствие их лиановидного облика, представляют собой уникальное образование в растительном мире (рис. 105). Именно листьям обязан лигодиум своим названием (от греч. lygodes - гибкий, извилистый). На первый взгляд они не похожи на узкие жесткие листья схизеи или актиностахиса. Однако молодые листья лигодиума выглядят так же, как листья актиностахиса пальчатого (Actinostachys digitata).
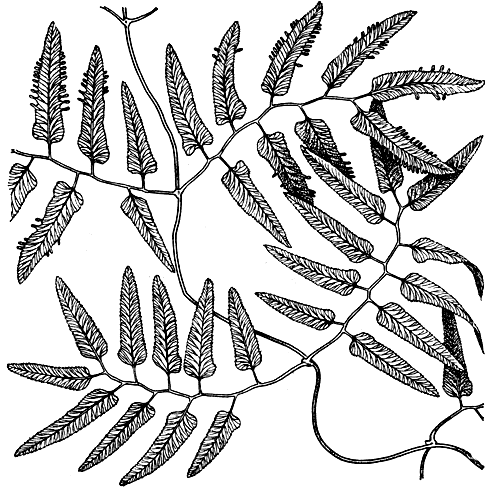
Рис. 105. Лигодиум вьющийся (Lygodium volubile), часть листа
Листья обладают неограниченным ростом в длину и у старых экземпляров имеют тонкий (диаметром 2-5 мм) ветвящийся стержень, образованный серией последовательных, очень неравных дихотомических ветвлений, при которых одна ветвь продолжает рост, а другая, короткая (называемая первичной) оканчивается спящей верхушкой. Они могут начать рост при повреждении дистальной части листа.
Стерильные листочки (или их доли) с цельным или пильчатым краем; только у лигодиума многоколосого (Lygodium polystachyum) они перистораздельные. Фертильные листочки несут узкие спорангиеносные зоны, развивающиеся на концах большинства жилок. Края спорангиеносных зон пильчатые, средняя жилка развивает очередные короткие боковые жилки, каждая из которых заканчивается единственным спорангием (рис. 106). Особенностью лигодиума является специальный кармашкоподобный индузиальный вырост, защищающий спорангий, прикрепленный вдоль жилки и открывающийся спереди. Как и у других родов схизейных, он представляет собой разрастание эпидермальных клеток листа, а спорангии являются краевыми по своему происхождению.

Рис. 106. Лигодиум улиткоообразный (Lygodium circinatum), спорангиеносная часть листа снизу
У анемии и мории листья наиболее соответствуют обычным представлениям о листьях папоротников; они однажды перистораздельные или дважды, трижды перисторассеченные. У анемии листья обычно разделены у основания на 3 пера, из них только два нижние (обычно вертикальные) фертильные, вдоль их конечных жилок в двух рядах располагаются спорангии, иногда столь многочисленные, что полностью закрывают поверхность пластинки фертильной части листа (рис. 107). Спорангии голые или с незначительным защищающим выступом листа, что отражено в родовом названии - анемия - Anemia (от греч. aneimon - голый). Стерильные перья имеют сегменты (перышки), сильно варьирующие по форме и степени рассеченности. У одного из наиболее примитивных видов этого рода (A. colimensis) фертильные перья почти не отличаются от стерильных, как и у видов мории, ближайшего к анемии рода. У мории спорангии распределены по всей длине спороносного листа, похожего по форме на стерильный, по краям каждого его сегмента, вблизи окончания каждой жилки и защищены отогнутым краем листа.

Рис. 107. Анемия листовиковая (Anemia phyllitidis), лист
Спорангии схизейных крупные, обратнояйцевидные или грушевидные, сидячие или на коротких массивных ножках (у лигодиума), асимметрично прикрепленные. Специфическим признаком семейства является наличие почти верхушечного (поперечного) кольца спорангия, раскрывающегося продольно, и состоящего обычно из одного ряда удлиненных клеток с утолщенными стенками. У некоторых видов лигодиума имеется неполный двойной ряд клеток кольца, что считается примитивным признаком.
У актиностахиса крупнобазалъного (Actinostachys macrofunda) кольцо спорангия нередко бывает двойным, с полным или неполным вторым рядом, а изредка развивается третий ряд. Несколько рядов клеток имели кольца спорангиев зенфтенбергии (Senftenbergia), вымершей к началу мезозоя. Поэтому считается, что эволюция схизейных шла в направлении уменьшения числа рядов клеток в кольце и, следовательно, лигодиум и актиностахис являются относительно примитивными родами.
В каждом спорангии содержится довольно большое число спор: 128-256 (редко 64). Споры билатеральные монолетные (схизея, актиностахис) или тетраэдрические трилетные (лигодиум, анемия, мория) с разнообразной скульптурой.
В семействе схизейных можно наблюдать разные типы гаметофитов. У схизеи они нитевидные, у актиностахиса клубневидные, а у остальных родов талломного типа: сердцевидные у лигодиума и более или менее асимметричные у анемии и мории (рис. 108). Считается, что редкие в мире папоротников нитевидные гаметофиты схизеи представляют собой их видоизмененную ювенильную стадию. Интересно, что, например, у лигодиума может продолжительно существовать нитевидная стадия и не образовываться сердцевидная пластинка, характерная для его гаметофита. Это связано, прежде всего, с условиями освещения. В других случаях (например, у актиностахиса) гаметофит не имеет нитевидной стадии: сразу же образуется осевой цилиндрический таллом.
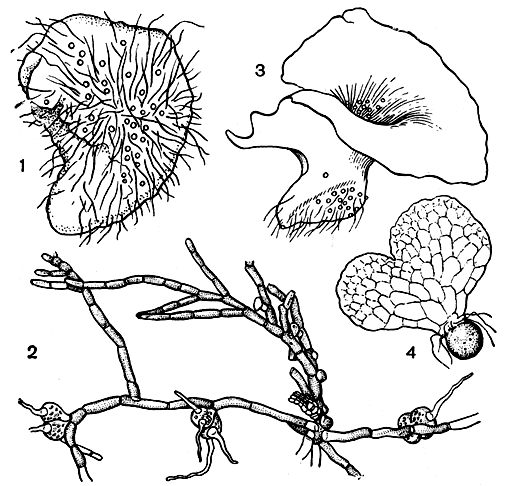
Рис. 108. Гаметофиты схизейных: 1 - актиностахис малоколосый (Actinostachys oligostachys); 2 - схизея маленькая (Schizaea pusilla); 3 - мория кафрская (Mohria caffrorum); 4 - лигодиум японский (Lygodium japonicum)
Развивающиеся на поверхности почвы гаметофиты схизей - зеленые, напоминают сильно ветвящуюся зеленую водоросль. У некоторых же видов они могут быть подземными (и, естественно, бесцветными). Гаметофиты образованы разветвленными однорядными нитями трех типов: стелющимися нитями, кустистыми нитями и ризоидами. Стелющиеся нити относительно длинные (до 5 см), слабо разветвленные, более или менее горизонтальные, промежуточные по толщине между двумя другими типами, несут антеридиофоры и ризоидофоры. Кустистые нити толще, с более короткими клетками, образуют компактные сильно ветвистые пучки, на которых развиваются архегониофоры.
Ризоидофоры - клубневидные клетки, расположенные латерально на стелющихся нитях и отделенные от них короткой клеткой - имеют по 2-5 ризоидов, прочно прикрепляющихся к частицам субстрата. Стенки ризоидов не кутинизированы и легко проницаемы для воды и растворов в отличие от остальных клеток гаметофита. Неразделенный перегородками грибной мицелий имеется в ризоидофорах, ризоидах и иногда в других вегетативных клетках гаметофита.
Гриб состоит в симбиотических отношениях с гаметофитом и способствует абсорбции почвенных растворов.
Антеридии возникают на коротких боковых ветвях нитей, архегонии концентрируются вблизи основания главной ветви кустистой нити. Гаметофиты актиностахиса растут в богатых органическими веществами и влажных субстратах на различной глубине. Под пологом леса в перегнившем растительном материале талломы могут находиться почти на поверхности почвы, а в слабо затененных местах - на глубине до 25 см.
На самых ранних этапах развития гаметофит актиностахиса имеет шаровидную форму, затем становится цилиндрическим, а впоследствии - клубневидным. У большинства видов гаметофит тонкий (диаметром 0,3-0,7 мм), ориентирован вертикально в субстрате, несет равномерно распределенные по боковой поверхности ризоиды и антеридии. Архегонии возникают на более старых клубневидных гаметофитах, например у актиностахиса меланезийского (Actinostachys melanesica). Клубневидная форма гаметофита с неправильными гребнями, шишками и углублениями формируется вследствие деятельности периферической меристемы и прекращения апикального роста. Наиболее крупные гаметофиты (актиностахис малоколосый - A. oligostachys), длиной до 15 мм при диаметре 2-5 мм, имеют массивную верхушку, несущую множество антеридиев и архегониев. У актиностахиса крупнобазального необычно большой гаметофит сохраняется в течение всей жизни спорофита, имеющего всего один лист шириной 1 мм и длиной 3-8 см. Спорофит этого вида сильно редуцирован. По степени редукции спорофита актиностахис крупнобазальный завершает ряд, в начале которого располагаются актиностахисы сглаженный и промежуточный. У них спорофит имеет по нескольку относительно больших листьев, а гаметофиты живут не более года, обычно отмирая ко времени образования второго листа молодого спорофита.
Антеридии актиностахиса по форме промежуточны между более крупными и более развитыми антеридиями псилотовых и глейхениевых и более простыми схизейного типа.
У большинства видов анемии (как и у лигодиума) гаметофит имеет удлиненную сердцевидную форму. У некоторых примитивных видов анемии и у рода мория гаметофит асимметричен с ранних стадий развития, так как рост происходит благодаря латеральной меристеме, а не в результате деятельности апикальной клетки. Молодой гаметофит мории сначала стелющийся, но после 7-11 месяцев развития становится прямостоячим. Он состоит из средней колоннообразной части и боковой крыловидной, обе поверхности которых функционально одинаковы. Гаметофиты анемии и лигодиума иногда могут быть однополыми, как в некоторых, более подвинутых семействах папоротников.
Наибольшее число видов лигодиума встречается в Юго-Восточной Азии, Австралии и Океании. С ними связаны виды тропической Африки, острова Мадагаскар и Коморских островов. В частности, лигодиум лазящий (L. scandens) имеет разорванный африкано-южноазиатско-австралийский ареал, что, несомненно, свидетельствует о его древности. Тропики Центральной и Южной Америки населены более примитивными видами, значительно обособленными от азиатских. Северная граница ареала лигодиума в восточном полушарии проходит по Японии, а в западном - по штату Массачусетс (США). На юге лигодиум выходит за пределы тропиков, достигая Новой Зеландии и Южной Африки.
Лиановидные лигодиумы встречаются на опушках девственного тропического леса, в тиковых лесах, в зарослях бамбука, иногда как сорняки на банановых плантациях.
Районы обитания лигодиума относительно влажные и в местах, где бывает продолжительный период засухи, встречается только один вид - лигодиум японский (L. japonicum), но листья в неблагоприятное время отмирают. Другие виды более требовательны к влажности и не выдерживают даже временного иссушения (лигодиум закрученный - L. circinatum).
Большинство видов лигодиума встречается в нижнем поясе гор, не выше 1000 м над у ровнем моря, и только лигодиум японский поднимается до 2550 м.
Лигодиум растет на глинистых, реже известняковых почвах, нередко в болотистых местах, особенно в районах с длительным сухим сезоном, а иногда в солоноватых озерах. Новогвинейский лигодиум Меррила (L. merrillii) - полуэпифит, длиной до 8 м, укореняющийся в моховых подушках на высоких деревьях. Некоторые виды стелющиеся, наземные, например лигодиум красивый (L. venustum) в Коста-Рике.
Известны виды ржавчинных грибов, паразитирующих на спорофите ряда видов лигодиума, при этом вторым хозяином является пихта.
Схизея - широко распространенный род не только в тропиках Старого и Нового Света, но и во внетропических районах южного полушария: в Южной Африке, Чили, Новой Зеландии, Тасмании. В северном полушарии современные виды известны только из Северной Америки.
Аналогичную картину представляет ареал рода актиностахис: большинство видов встречается от Индии и Шри Ланка до Южной части Новой Гвинеи, Новой Каледонии, Австралии и Мадагаскара. Особая группа видов распространена в Вест-Индии, Центральной и Южной Америке. Имеется общий для этих двух территорий вид - актиностахис кисточковидный (A. penicillata).
Необычные по виду папоротники схизея и актиностахис, скорее напоминающие узкими линейными листьями представителей лилейных или ситниковых, а иногда проростки злаков, нередко ускользают от внимания даже опытных ботаников. Например, актиностахис перышковый (A. pennula) не приводился для флоры Французской Гвианы, хотя он обильно растет в островках леса среди саванны вблизи г. Кайенны. Представители этих родов встречаются на бедных кислых, песчаных или торфяных почвах с несомкнутой растительностью, чаще на равнинах и в предгорьях. Есть среди них высокогорные виды, например схизея дудчатая (S. fistulosa), растущая на альпийских болотах и каменистых местах от 2400 до 3750 м над уровнем моря. Схизея малаккская (S. malaccana), также достигающая высот 800-2000 м, встречается на замшелых камнях или как эпифит на деревьях.
Миниатюрная (высотой 3-6 см, изредка до 10 см) северо-американская схизея маленькая (S. pusilla) весьма похожа на злак, и только при микроскопическом исследовании можно установить, что это все-таки папоротник. Из всего семейства она наиболее далеко заходит на север: в США (Нью-Джерси) и Канаду (Ньюфаундленд, Новая Шотландия). Это реликтовое растение, пережившее в Северной Америке периоды оледенения. Действительно, схизея маленькая может выдерживать довольно низкие температуры. Например, гаметофит в культуре оставался жизнеспособным в течение 13 лет, выдерживая зимние периоды в неотапливаемом помещении. В северо-восточных штатах США схизея маленькая растет на пустошах и сфагновых болотах в ассоциации с клюквой и видами росянки. Близкий вид - схизея мощная (S. robusta), отличающаяся большими размерами и формой листьев,- обитатель дождевых лесов и горных болот Гавайи.
В экологическом отношении многие из схизейных могут считаться ксерофильными, но мория и анемия - наиболее устойчивые к засухе папоротники. Мория встречается в горах на высоте от 1300 до 2300 м на лесных опушках у основания крупных камней и скалистых выступов. Ареал мории включает южную и юго-восточную части Африки, Мадагаскар и Маскаренские острова.
Род анемия ограничен преимущественно Неотропическим царством с наибольшим числом видов на сухих плато Южной Бразилии. Только 7 видов (из 90, входящих в род) известны на Антильских островах и в Северной Америке (до Техаса). Восемь видов, изолированно встречающихся в Юго-Восточной Африке, Абиссинии и Индии, по-видимому, реликты. Особая группа видов, центры разнообразия и эндемизма которой в Бразилии и Мексике, наиболее близка к ископаемым представителям этого рода из меловых отложений Патагонии.
Анемия в Южной Америке растет в лесах на высоте от 400 до 600 м над уровнем моря, на остепненных участках (в Чили и на севере Аргентины) - до 2700 м, а в Южной Африке - в подлеске сухих лесов от 500 до 1500 м.
В странах Юго-Восточной Азии некоторые схизейные, в особенности лигодиум, находят довольно широкое и разнообразное применение. Жесткие прочные и длинные стержневые части листа лигодиума (с диаметром 1,5-2 мм) используются как материал для плетения. Высушенные на солнце осевые части с удаленными листочками, цельные или расщепленные на полоски, имеют различные цветовые оттенки и пригодны без окраски для изготовления шкатулок, корзинок, сигаретных коробок, браслетов, шляп. Браслетам, изготовленным из листьев лигодиума ушковидного, приписывается магическая сила, предохраняющая от отравления ядами.
Ритуальное значение имеют свежесрезанные растения некоторых видов лигодиума. Они используются в свадебных церемониях, а также празднествах, связанных с различными жизненно важными событиями (постройка дома, посадка или сбор риса, рыбная ловля и т. д.).
Очень молодые листья лигодиумов закрученного и мелколистного съедобны и используются в пищу на острове Ява.
Некоторые виды используются в народной медицине: от лихорадки, кожных болезней, для лечения открытых ран (лигодиум мелколистный). Схизея вильчатая используется при кашле и болезнях горла.
Многие схизейные, особенно лигодиум и анемия, используются в качестве декоративных растений, чаще как комнатных в странах с умеренным климатом, а лигодиумы мелколистный и японский, выращиваемые в садах, даже натурализовались в Северной Америке.
Семейство адиантовые (Adiantaceae) (В. Н. Гладкова)
Адиантовые - обширное семейство, объединяющее морфологически разнообразные типы, главным образом тропических папоротников с ползучими, обычно соленостелическими корневищами, покрытыми чешуями или, реже, волосками. Для всех представителей семейства характерны лишенные настоящего покрывальца сорусы, либо расположенные по всей длине жилок на нижней поверхности листа, либо сосредоточенные на концах жилок ближе к краю и часто объединенные в одну непрерывную линию. В последнем случае они обычно защищены отвернутым краем листа или лежат в особом углублении бороздке (табл. 20). Спорангии их с продольным (вертикальным) кольцом толстостенных клеток содержат обычно тетраэдрические трилетные споры, но в некоторых группах представлены и билатеральные споры. Разнообразию спорофитов этих папоротников соответствует и разнообразие их гаметофитов. В семействе встречаются ремневидные, сердцевидные, почковидные гаметофиты, у многих родов гаметофит асимметричный, с боковым выростом, образующимся благодаря деятельности латеральной меристемы. При таком разнообразии этих папоротников удивительна их константность в отношении основного хромосомного числа: почти все до сих пор исследованные представители семейства являются диплоидами или полиплоидами на основе числа х = 29 или 30.

Таблица 20. Типы спороношепий у папоротников семейств адиантовых и полиподиевых: вверху слева - хемионитис пальчатый (Hemionitis palmata); вверху справа - (Adiantum trapeziforme); внизу - дринария Мейена (Drynaria meyeniana)
Среди птеридологов нет единства в понимании объема семейства и ранга различных групп папоротников, относимых к адиантовым, так как большой полиморфизм этих растений не позволяет разделить их на четкие категории. Особенно это касается групп родов, объединяемых в подсемейство собственно адиантовых.
Подсемейсшво адиантовые (Adiantoideae)
Подсемейство собственно адиантовых охватывает до 47 родов. Наиболее крупным среди них является род адиантум (Adiantum), широко известный в европейских странах также под названием папоротник девичьи волосы или папоротник женские волосы (англ.- maidenhair fern, нем.- Frauenhaarfarn). Название это дано адиантуму за его воздушную изящную листву, неповторимую у других папоротников. Тонкие стержни (рахисы) листьев адиантумов обычно блестящие, темные и упругие, действительно напоминают волосы, а листва в целом с ее изящными вееровидными или иной формы сегментами - женские кудри (рис. 109). Различные легенды связаны с этим растением. Согласно одним на том месте, где упала с утеса прекрасная девушка, возник чистый источник, а волосы девушки превратились в папоротник. Другие легенды связывают происхождение этого папоротника с богиней Венерой. Один из видов его так и называется адиантум венерин волос (Adiantum capillusveneris).

Рис. 109. Адиантум венерин волос (Adiantum capillus-veneris): 1 - часть листа; 2 - сегмент с сорусами; 3 - край сегмента листа
Но не всем видам адиантума свойственна такая воздушная, необычайно красивая листва. Имеются среди них и виды с жесткими грубо рассеченными листьями, а также виды цельнолистные, как, например, растущий на островах Атлантического океана адиантум почковидный (A. reniforme). Всего в роде адиантум насчитывают до 200 видов, растущих в тропических и субтропических странах; особенно много их в американских тропиках.
Адиантумы - наземные растения с ползучими или почти прямостоячими, соленостелическими или диктиостелическими корневищами, обычно покрытыми узкими коричневатыми чешуйками. Листья у большинства видов перистые, от однажды перистых до пятикратно перистых, с дихотомически ветвящимися свободными, реже анастомозирующими жилками. У некоторых видов листья несут на верхушке выводковые почки. Характерной особенностью листьев являются их водоотталкивающие свойства. Капли воды стекают с них, не смачивая поверхность. Этому свойству род и обязан своим названием Adiantum (от греч. а - не и diaino - смачивать).
От других представителей семейства род адиантум отличается своеобразным расположением сорусов. Они лежат здесь обычно вдоль концов жилок, идущих до самого края листового сегмента. Именно эта, покрытая сорусами, часть сегмента отгибается вниз, сегмент в верхней части как бы складывается пополам, и сорусы оказываются внутри особых карманчиков.
Обычное местообитание адиантумов - тенистые влажные леса, где они растут либо на почве, либо на скалистых субстратах. Встречаются адиантумы также по оврагам и речным берегам, возле валунов и термитников. Растут они и на довольно открытых, но влажных местах, а некоторые виды могут выносить и засушливые условия.
Красота и разнообразие этих папоротников издавна привлекали внимание человека, и многие виды адиантума широко вошли в культуру. В России большой интерес к ним возник еще в конце прошлого века. Журнал "Русский вестник садоводства" за 1890 г. рекламировал виды адиантума как одни из лучших растений для разведения в теплицах и комнатах. И до сих пор адиантумы украшают оранжереи, как эффектные единичные экземпляры в корзинках и вазонах, или, умело подобранные по окраске и характеру листвы, они заполняют целые стеллажи в оранжереях, оттеняя красоту друг друга. Адиантумы легко размножаются спорами, часто дают самосев и при достаточной влажности и непрямом освещении хорошо переносят условия культуры. В оранжереях выращивается множество форм американского тропического адиантума Радди (Adiantum raddianum), восточноазиатского и американского адиантума стоповидного (A. pedatum) и многих других.
К числу видов, особенно популярных в культуре, относится адиантум венерин волос, воплощающий в себе все изящество этих папоротников. Листья его очень нежные и тонкие, дважды, трижды перистые с клиновидными сегментами, в свою очередь сидящими на волосовидных черешочках. Темные сорусы просвечивают сквозь тонкую ткань листа, обрамляя его темным пунктиром. Произрастая дико в тропических областях, этот вид достигает и пределов СССР. Он растет в умеренных, субтропических областях, реже в Крыму, на Кавказе и в горах Средней Азии в трещинах скал у просачивающейся воды, у водопадов, по каменистым берегам горных ручьев, на кальцитных породах, на туфах.
На советском Дальнем Востоке (в Приморье и Приамурье) в травостое смешанных и широколиственных лесов обильно представлен другой вид этого рода - изящнейший адиантум стоповидный (Adiantum pedatum). Среди разнообразных представителей травяного яруса он выделяется своими красивыми вееровидными дважды перистыми листовыми пластинками, поднятыми над почвой на 20-40 см на блестящих каштаново-черных черешках.
Виды адиантума в древности (упоминаются уже в сочинениях Плиния) были известны не только как декоративные, но и как лекарственные растения. Сходство их ажурной листвы с кудрями давало повод древним медикам рекомендовать эти растения для лечения волос. В менее отдаленном прошлом индейцы лечили экстрактами из листьев от укусов насекомых, простудные заболевания, втирали их в волосы, употребляли как кровоостанавливающее средство. Черные блестящие черешки некоторых видов адиантума используются индейцами для украшения плетеных изделий.
Птеридологи выделяют в подсемействе адиантовых группы близких родов. Названия групп производят от имени типичного рода.
Гимнограммоидные папоротники являются наиболее примитивной группой подсемейства. К ним относится свыше 20 родов ксерофильных или мезофильных папоротников, очень различных по внешнему облику. Спорангии их нечетко дифференцированы на сорусы и лежат вдоль жилок подобно спорангиям некоторых схизейных.
Род хемионитис (Hemionitis) имеет типичные гимнограммоидные сорусы. Для его листьев характерно сетчатое жилкование, и такую же сеточку образуют простертые вдоль жилок спорангии (табл. 20). В род хемионитис входят 6-8 мезофильных, главным образом тропических, американских видов, только один вид встречается в Азии. Это некрупные наземные растения, с почти прямыми чешуйчатыми корневищами и с простыми листьями. У широко распространенного в Америке хемиопитиса дланевидного (Н. palmata) листья пальчато-лопастные; для азиатского очень своеобразного представителя этого рода - хемиопитиса аронниколистного (Н. arifolia) - характерны стреловидные или сердцевидные листья.
Типичной для хемионитиса чертой является своеобразный способ вегетативного размножения. На боковых жилках ближе к основанию или вдоль края стерильных листьев образуются почки, которые в молодом состоянии закрыты на верхушке несколькими чешуйками. Почки могут долго оставаться покоящимися, но при благоприятных условиях дают начало дочерним растениям.
Некоторые гимнограммоидные папоротники распространены исключительно в Америке. К ним принадлежит джеймсония (Jamesonia) с 20 видами, живущими в горных областях на высоте от 1500 до 5000 м от Южной Мексики до Центральной Боливии и Бразилии. Очень характерны виды джеймсонии для пояса парамо в Андах. Условия произрастания в этих высокогорных областях весьма суровы: низкие температуры (в течение всего года по ночам заморозки), сильная инсоляция, высокая влажность почвы и воздуха, облачность и туманы. Подобно растениям наших северных верховых болот, также существующих в условиях прохладных температур и высокой влажности, джеймсония приобрела черты ксероморфности, в частности редуцированную листовую поверхность. Листья у джеймсонии узкие, линейные, всегда однажды перистые с многочисленными очень мелкими (диаметром до 1 см) сегментами. Сегменты расположены в два ряда и черепитчато налегают друг на друга. В общей массе листья иногда кажутся объемными, а не плоскими. Это впечатление еще более усиливается благодаря их густому опушению. Войлочно опушенные молодые листья плотно прижаты к тонкому ползучему корневищу, также густо покрытому щетинковидными коричневатыми волосками.
Корневища джеймсонии часто глубоко скрыты в дернине, образованной травами или мхом, и, по некоторым наблюдениям, способны выживать после пожаров. Живописную картину представляют колонии джеймсонии. Все листья их, поднимающиеся над землей, имеют слегка свернутые, загнутые вниз верхушки, что у других папоротников наблюдается только на молодых листьях. Дело в том, что апикальная меристема листьев джеймсоний функционирует неограниченно долго, обеспечивая их непрерывный рост. По мере того как сегменты листа у его основания отмирают, на верхушке появляются все новые и новые сегменты, и листья растут в течение всей жизни папоротника.
Сходный с джеймсонией ареал и также с наибольшим разнообразием видов в Андах имеет еще один американский гимнограммоидный папоротник - эриосорус (Eriosorus), прежде известный под названием гимнограмма (Gymnogramme). Он включает около 25 видов. Это в большинстве случаев среднего размера папоротники с дважды, трижды (иногда более) перистыми листьями разнообразной формы. Встречаются среди них и виды с крупными (до 3 м длиной) листьями, цепляющимися или лазающими по опоре.
По расположению сорусов к гимнограммоидным папоротникам принадлежат также два близких рода: анограмма (Anogramma) и питирограмма (Pityrogramma). Виды первого (их всего 5-6) растут в субтропических и умеренно теплых областях, главным образом северного полушария. Маленький изящный папоротничек анограмма тонколистная (Anogramma leptophylla) широко распространен на скалистых затененных местообитаниях. Встречается он в Центральной и Южной Америке, Австралии, Тасмании, Новой Зеландии, в Западной и Южной Европе, в Африке, а в СССР в горах Закавказья и Средней Азии (Копетдаг). В 1910 г. этот папоротник был найден на Южном берегу Крыма на прибрежных скалах горы Кастель, но с тех пор больше никто его там не собирал.
Очень тонкие и нежные, дважды или трижды перистые мелкие листья анограммы тонколистной иногда принимают за молодые спорофиты других папоротников. При внимательном рассмотрении, однако, на их нижней поверхности хорошо видны расположенные вдоль жилок и сливающиеся друг с другом лишенные покрывальца сорусы. Стерильные листья у этих папоротничков меньше фертильных по размеру. Анограмма тонколистная интересна в биологическом отношении. Ее спорофит эфемерный, однолетний, каждый год он полностью (вместе с корневищем) отмирает. Многолетним является ее гаметофит, который выработал замечательные приспособления для перенесения неблагоприятных условий. В частности, он образует особый вырост, дистальная часть которого превращается в запасающий орган - бугорок с запасом питательных веществ, позволяющий гаметофиту переносить период с неблагоприятными условиями.
Виды рода питирограмма распространены главным образом в Америке, из 40 видов только несколько встречается в Африке и, как заносные, в Азии. Они относятся к числу "серебристых" и "золотистых" папоротников, широко распространенных в культуре: их листья снизу покрыты желтоватым или белым восковидным налетом, продуцируемым особыми внутренними головчатыми железками. Мода на культуру питирограмм была столь велика, что это привело к натурализации их во многих влажнотропических странах. Широко натурализовался американский "серебристый" папоротник питирограмма красиво-черная (Pityrogramma calomelanos). Сейчас она встречается повсюду в тропиках, заселяя расчищенные человеком земли, распространившись как сорняк на полях и чайных плантациях.
Питирограмма красиво-черная, как и некоторые другие папоротники этого семейства (Pteris, Acrostichum), была найдена в числе первых растений, поселившихся на острове Кракатау спустя три года после катастрофического извержения 1883 г., уничтожившего всю растительность и покрывшего остров слоем пемзы и пепла толщиной до 30 м.
Хейлантоидные папоротники. Особую, со своеобразными признаками группу составляют в подсемействе адиантовых хейлантоидные папоротники, произрастающие главным образом в аридных областях Южной Америки, Евразии, Австралии и особенно Мексики, во флоре которой их насчитывают более 100 видов. Однако, по данным Р. Трайона, они почти не встречаются в местах, где постоянно сухо, они очень характерны для мест с чередованием сухого и влажного сезонов в течение года. Всего в эту группу включают 7-15 родов и до 325 видов.
Хейлантоидные папоротники - ксерофильные растения с прямостоячими или ползучими, у большинства видов соленостелическими корневищами, одетыми непрозрачными чешуями. Листья многих из них кожистые и часто бывают защищены от излишнего испарения густым покровом из чешуй, волосков или восковидным налетом.
До 130 видов входят в род хейлантес, или краекучник (Cheilanthes), характерный для засушливых местообитаний Америки и Африки. Немногие его виды встречаются также в Средиземноморье и горных областях Азии, в Австралии и на островах Тихого океана. Это некрупные растения с короткими корневищами, густо покрытыми узкими, иногда двуцветными (с коричневой полосой по центру) чешуями. Листья их обычно многократно перистые, жесткие, иногда вздутые, края их завернуты на нижнюю сторону и защищают сорусы, расположенные обособленно или непрерывной линией почти по краю сегментов (отсюда название рода). На черешках и на нижней поверхности листьев некоторых видов имеются секреторные волоски, выделяющие кристаллизующееся вещество, покрывающее поверхность золотистым или серебристым налетом. У других видов поверхность листа сплошь покрыта большим количеством чешуй и волосков. Эти и другие приспособления, защищающие папоротник от интенсивного испарения, позволяют краекучнику обитать в скалистых местах (на уступах и в щелях), на открытых солнечных пространствах, по склонам осыпей, по земляным насыпям у дорог. В Австралии виды хейлантеса можно встретить на голой почве полувечнозеленых лесов, где почти ничего не растет, кроме мхов.
Многие виды хейлантеса способны переносить многомесячные засухи. В этот период они становятся совершенно невзрачными и незаметными, но при первом же дожде они впитывают воду в количестве, в несколько раз превышающем их сухую массу, и оживают снова. Такая способность переносить засуху встречается у спорофитов папоротников не столь уж часто, но еще более удивительной является приспособленность гаметофитов хейлантесов. Их гаметофит способен сохранять практически неограниченную жизнеспособность во время засухи и при первом случайном дожде давать жизнь новым спорофитам.
Гаметофиты хейлантеса представляют собой тонкие (толщиной в 1-3 клетки) сердцевидные пластиночки с железистым опушением. Они не имеют специальных приспособлений против потери воды, оболочки их клеток не кутинизированы, и при засухе они теряют практически всю воду. И хотя при длительной засухе большая часть тела гаметофита может оказаться мертвой, где-нибудь по краю или в центре пластиночки сохраняется группа зеленых клеток, которые быстро делятся при увлажнении и образуют выросты, подобные по строению первичному гаметофиту.
После процесса оплодотворения или апогамно из клеток живой ткани развиваются спорофиты. Этот процесс важен не только для выживания растения в условиях засухи, но и для его размножения, так как один гаметофит при увлажнении дает до 10 выростов, каждый из которых становится самостоятельным растением.
Опыты с американским скальным папоротничком хейлантесом изящнейшим (С. gracillima) показали, что гаметофиты его оставались живыми после 9 месяцев высушивания и после помещения их в условия достаточного увлажнения начинали нормально функционировать. В природе были найдены также гаметофиты, не оставляющие сомнения в том, что возраст их около года и, следовательно, они пережили засушливый летний период и последовавшую за ним зиму. Как и в культуре, у этих гаметофитов наблюдались многочисленные пролиферации и вегетативный рост. Хейлантес изящнейший, кроме того, имеет и другое приспособление для перенесения засухи: его спорофит способен задерживать рассеивание спор в сухой период. Споры его созревают в апреле - мае, но в течение всего сухого сезона они сохраняются в спорангиях на листьях, которые свернуты, приобретая почти цилиндрическую форму. Только с приходом дождей, обычно осенью, листья разворачиваются, и тогда происходит рассеивание спор.
В СССР распространены 5 видов хейлантеса. В Сибири и на Дальнем Востоке на скалах и каменистых склонах встречается хейлантес серебристый (С. argentea). В Крыму и на Кавказе растет хейлантес марантовый (С. maranthae), иногда относимый к близкому роду нотолена, или ложнопокровница (Notholaena), отличающемуся от хейлантеса незавернутыми краями листьев (рис. 110). На скалах Аю-Дага в Крыму этот вид иногда покрывает участки до нескольких квадратных метров.
Виды хейлантеса и нотолены относятся к числу наиболее красивых адиантовых, разводимых в оранжерее. Они составляют основную массу "золотистых" и "серебристых" папоротников. Обычно их выращивают в подвешенных корзинках, где они выглядят наиболее эффектно.
Сходные с хейлантесом приспособления к засухе имеет род пеллея (Pellaea), включающий около 80 видов, живущих в аридных и горных областях обоих полушарий. Наибольшее количество видов пеллеи - в Южной Америке и Южной Африке. На юге виды пеллеи достигают Чили и Новой Зеландии, на севере доходят до Канады. В опытах гаметофиты пеллеи оживали и обнаруживали способность к росту и размножению после 18,5 месяца высушивания, а 5% культивируемых гаметофитов сохранили полную или частичную жизнеспособность после пяти лет содержания их в темном, хорошо вентилируемом шкафу. Споры видов пеллеи, взятые с гербарных образцов, собранных от 2 до 5 лет до этого, прорастали в течение 10 дней.
Кожистые темноватые листья пеллеи подчас совершенно не похожи на листья папоротников. Они перистые или многократно перистые, сверху обычно гладкие, снизу с желтоватым или беловатым налетом, сегменты их часто на черешочках, а рахис иногда образует своеобразный зигзаг (рис. 110). И только полоски спорангиев с нижней стороны, прикрытые в молодом состоянии краем листа, выдают их принадлежность к папоротникам. Как и другие хейлантоидные, виды пеллеи растут в сухих местообитаниях. Их корневища, покрытые двуцветными чешуями, глубоко врастают в трещины скал. Иногда пеллеи поселяются на вулканических породах в русле бывших лавовых потоков, в трещинах и ямках, где скапливается немного почвы. Случается, что растут они и на стенах старых домов, и даже на стволах де-ревьев. Скромные и необычные виды пеллеи были очень популярны в английских и американских оранжереях еще в викторианскую эпоху. Многие из них и сейчас входят в состав оранжерейных коллекций, так как они красивы и неприхотливы в культуре. Ценятся некоторые пеллеи и как лекарственные растения: листья пеллеи остроконечной (P. mucronata) мексиканцы заваривают как чай. Напиток этот вкусен и обладает лечебными свойствами.

Рис. 110. Хейлантоидные папоротники. Хейлантес марантовый (Cheilanthes maranthae): 1 - общий вид; 2 - сегмент листа с нижней стороны (часть чешуек удалена, чтобы показать сорусы); 3 - чешуя корневища. Пеллея яйцевидная (Pellaea ovata): 4 - часть листа; 5 - сегмент листа с сорусами; 6 - двуцветная чешуя корневища
Для многих видов хейлантоидных папоротников характерно апомиктическое размножение: спорофиты их часто вырастают из соматической клетки гаметофита, минуя процесс оплодотворения. В спорангиях таких апогамно развившихся спорофитов число спор обычно уменьшено вдвое и равно 32. Апомиктическое размножение, свойственное этим ксерофильным папоротникам, рассматривается некоторыми птеридологами как еще одно приспособление к ксерофитному образу жизни: с переходом к апомиксису этим папоротникам удается стать независимыми от наличия свободной воды, необходимой для осуществления оплодотворения.
Криптограммоидные и другие папоротники. По расположению сорусов по краю листа близок к хейлантоидным папоротникам небольшой род криптограмма (Cryptogramma) с 4 видами в умеренных областях северного полушария. Все 4 вида встречаются и в пределах СССР, два из них заходят в Арктику. Это небольшие папоротники, растущие в горах на грубых каменистых россыпях и на скалах. Среди немногочисленных папоротников умеренной зоны эти миниатюрные растения легко узнаются по диморфизму их многократно перистых листьев: спороносные листья у них длиннее и с более узкими, кажущимися сверху линейными сегментами, края которых загнуты вниз. Листья криптограммы либо скучены на коротком корневище, как у криптограммы курчавой (С. crisра), либо расставлены по длинному ползучему корневищу, как у криптограммы Стеллера (С. stelleri).
Чтобы завершить наше далеко не полное описание разнообразия папоротников подсемейства адиантовых, упомянем еще один из них - род актиниоптерис (Actiniopteris), включающий 5 видов, растущих в засушливых местообитаниях в тропической Африке и Азии. Виды актиниоптериса напоминают по внешнему облику миниатюрные пальмы. Пальмовидный облик придают им их скученные жесткие листья, пластинка которых, полукруглая или почковидная в очертании, рассечена на узкие, в свою очередь дихотомически раздельные и расположенные веером сегменты.
Подсемейство виттариевые (Vittarioideae)
Виттариевые - небольшая группа очень специализированных, преимущественно эпифитных папоротников, включающая 7-9 родов, распространенных в тропических и субтропических странах. Все они - некрупные травянистые растения с простыми цельными (очень редко вильчатыми) листьями и с ползучими корневищами, покрытыми решетчатыми чешуями. В чешуях внутренние боковые стенки клеток темные и сильно утолщенные, тогда как остальные стенки тонкие и прозрачные, что придает своеобразный решетковидный облик рисунку на чешуе. Роды виттариевых характеризуются одним основным хромосомным числом 30 и имеют общие черты не только во внешнеморфологическом строении (решетчатые чешуи, расположение сорусов в особых углублениях ткани листа, удлиненный гаметофит), но и в анатомической структуре. Так, все они имеют особые, вытянутые, почти игловидные клетки-идиобласты в эпидерме листьев. (Среди адиантовых такие клетки встречаются также у рода Adiantum.) Характерной особенностью виттариевых является также отсутствие склеренхимных элементов во всех частях растений. Механическую функцию у многих видов выполняет колленхима.
Свое название подсемейство получило от рода виттария (Vittaria). Этот род, самый крупный в подсемействе, включает 80 видов. Все виды виттарии имеют узкие линейные листья, похожие на листья злаков, с очень характерным "виттариоидным" типом жилкования: боковые жилки смыкаются между собой, образуя одну прикраевую жилку и по одному ряду ареол (замкнутых жилками участков поверхности листа) с каждой стороны средней жилки. Вдоль наружной стороны этих ареол в особых углублениях - бороздках - расположены непрерывной линией спорангии. Края бороздок часто бывают приподняты, надежно защищая скрытые в глубине спорангии и выполняя ту же функцию, что покрывальца у других папоротников.
Защитную функцию несут и многочисленные парафизы (свойственные этому подсемейству), особые стерильные образования в сорусе, иногда похожие на спорангии, но большей частью иной формы, у виттарии состоящие из желтоватой или коричневатой разросшейся конечной клетки и простой или ветвистой многоклеточной ножки. Парафизы у некоторых видов виттарии присутствуют в таком количестве, что спорангии среди них теряются.
Листья виттарии иногда расставлены по ползучему корневищу, но обычно они сближены в небольшом числе и у взрослых эпифитных папоротников повисают, придавая оригинальный облик этим растениям, мало похожим на другие виды папоротников. Листья некоторых крупных видов виттарии при ширине 1-5 мм достигают длины 1 м, а у одной из самых маленьких и самых простых по структуре - виттарии наименьшей (V. minima), растущей в тропиках Нового Света, их длина всего 3-6 см.
В тропиках виттария довольно обычна на поросших мхами стволах деревьев по берегам рек, в низинах и на небольших высотах в горах. Виттария мечевидная (V. ensiformis) часто растет на стволах пальм, используя дождевую воду, задерживающуюся в покрове из оснований опавших пальмовых листьев (рис. 111).
Как и другие представители подсемейства, род виттария имеет удлиненные, ремневидные, долго живущие гаметофиты, интересной особенностью которых является их способность размножаться вегетативно, путем образования гемм (выводковых почек). Гаметофит становится как бы независимым и самостоятельным в своем существовании, не переставая в то же время выполнять и свою основную функцию: давать жизнь новым спорофитам. Недавно американские ученые У. Вагнер и А. Дж. Шарп обнаружили в Аппалачах на востоке США гаметофит виттарии, который "забыл" о своем основном назначении: ни в природе, ни в культуре он не образует гаметангиев и затем спорофитов. Его геммы, представляющие собой веретеновидные выросты из 4-10 клеток, разносятся ветром или животными и прорастают в благоприятных местах в новые гаметофиты. Сами гаметофиты не крупные, шириной до 2 мм и длиной до 10 мм, но их скопления на кварцитовых или песчаниковых скалах и на коре деревьев могут покрывать значительные участки, от 10 до 100 см в поперечнике. По своим морфологическим чертам этот гаметофит больше всего напоминает гаметофит виттарии полосчатой (V. lineata), широко распространенной в теплых областях Америки.
Стерильный гаметофит распространен на сотни миль севернее ареала виттарии полосчатой. Таким образом, являясь производным ныне живущего вида или близкородственного ему вымершего вида, этот папоротник демонстрирует явление крайней редукции, когда из биологического цикла исчезло целое поколение - спорофит.
Другой крупный род этого подсемейства - антрофиум (Antrophyum) - распространен в тропиках и субтропиках Старого Света (близкие к нему виды Нового Света выделяются в особый род политениум - Polytaenium). Все 40 видов антрофиума характеризуются особым жилкованием, состоящим из густой сети ареол, при отсутствии (по крайней мере в верхней половине листа) срединной жилки (рис. 111). Сорусы антрофиума либо поверхностные и также образующие сетку, либо погруженные в отдельных, иногда ветвящихся линиях. Корневища у этих папоротников одеты густой массой волосистых ветвящихся корней, активно поглощающих воду, что в сочетании с плотными, иногда мясистыми листьями, способными ее запасать, является адаптацией к перенесению засушливого периода. У некоторых видов листья во время засухи сморщиваются и сворачиваются, но с приходом дождей оживают снова. Растут виды антрофиума не только на стволах деревьев, но и на скалах в тенистых местах. Споры антрофиума, как и других виттариевых, бывают тетраэдрическими и билатеральными. Прорастать они начинают иногда прямо в спорангии, до рассеивания, и, вскрыв спорангий, можно обнаружить там гаметофиты в начальной стадии развития.
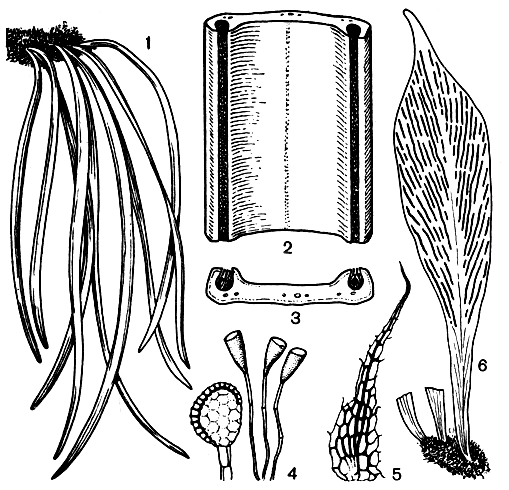
Рис. 111. Виттариевые. Виттария мечевидная (Vittaria ensiformis): 1 - общий вид; 2 - часть листа с полосками сорусов; 3 - разрез листа; 4 - спорангий и парафизы; 5 - чешуя. Антрофиум подорожниковый (Antrophyum plantagineum): 6 - лист с полосками сорусов
Род монограмма (Monogramma) с двумя видами в тропиках Старого Света относится к числу самых маленьких по размеру не только в подсемействе виттариевых, но и среди папоротников вообще. Монограмма дарееплодная (М. dareicarpa), произрастающая в очень влажных лесах и по берегам рек в Юго-Восточной Азии, имеет листья длиной 1-2 см и шириной 1-1,5 мм, с единственной неветвящейся средней жилкой. Ксилема в ней толщиной всего в несколько клеток. Вдоль верхнего конца жилки в бороздке, прикрытой выростом ткани нижней поверхности листа, лежат спорангии. Разрастаясь с помощью своего тонкого ползучего корневища, этот папоротничек, несмотря на миниатюрность, может образовывать плотные скопления на стволах деревьев в их нижней части.
Подсемейство птерисовые (Pteridoideae)
Род птерис (Pteris) и несколько близких к нему небольших родов, составляющих подсемейство птерисовых,- наземные папоротники, распространенные в тропических и субтропических бтранах обоих полушарий, главным образом во влажных областях. К роду птерис принадлежат около 280 видов. Только немногие из них достигают в своем распространении Средиземноморья, Японии и США в северном полушарии, а в южном некоторые доходят до Новой Зеландии, Тасмании и Южной Африки. Но они хорошо знакомы и жителям умеренных стран, так как их часто выращивают в комнатной и оранжерейной культуре.
Жестковатые листья птерисов обычно скучены на коротком корневище, покрытом непрозрачными чешуями и иногда волосками. Любителям папоротников известно, сколь разнообразны могут быть эти листья: от перистых и трехраздельных до многократно перистых, с гладким или покрытым шипиками стержнем, с зелеными, желтыми, коричневатыми черешками, с разнообразной окраски и формы сегментами. У основания листьев сегменты обычно глубоко рассечены, к верхушке они становятся менее рассеченными или цельными.
Общий признак всех птерисов - расположенный по краю листа (маргинальный) ценосорус, прикрытый ложным линейным покрывальцем, образованным из отвернутого края листа. Ценосорус лежит на краевой жилке, соединяющей свободные или с небольшим количеством анастомозов жилки. Составляющие ценосорус спорангии перемешаны с большим количеством парафиз, снабжены ножкой из трех рядов клеток и вертикальным кольцом для вскрывания.
В природе птерисы достигают иногда в высоту 2,5 м и более. Они растут на равнинах и невысоко в горах, в тени лесов или в условиях несильного затенения - на лесных опушках вблизи воды, в заболоченных лесах, часто на скалах у ручьев и по берегам рек, иногда встречаются и в сухих местообитаниях. Колония одного из видов - птериса ленточного (P. vittata), растущего дико в теплых странах Старого Света, была недавно обнаружена в совершенно необычных условиях в Великобритании. Папоротники росли на вершине шлаковой кучи у давно заброшенной шахты, вблизи струек пара, возникающих от испарения воды вследствие внутреннего возгорания шлака. Немногие другие растения смогли соседствовать с этим птерисом в таких условиях, несмотря на наличие тепла и влажности. Предполагают, что эта папоротниковая колония возникла из единственной споры, занесенной ветром от культивируемого растения, и существует там с 1945 г.
Не только птерис ленточный, но и другие культивируемые виды этого рода иногда дичают в теплых странах. Птерис многоразделъный (P. multifida), например, широко распространился на юго-востоке США, процветая там в трещинах старых каменных стен, по берегам
небольших речек, у основания стволов крупных деревьев. Иногда его называют гугенотским папоротником, так как считают, что он был завезен в Америку в XVII в. гугенотами - переселенцами из Франции, где этот папоротник уже тогда культивировали. Родина его - Китай, в некоторых р.айонах которого он является самым обычным растением.
Легкой приспособляемости этих папоротников к необычным условиям существования, быть может, способствует свойственное многим из них апогамное размножение, более надежно обеспечивающее возникновение спорофитов. Среди птерисов имеются виды облигатно апомиктические, например птерис критский (P. cretiса), часто встречающийся в оранжерейной и комнатной культуре. Область его распространения в природе простирается от Средиземноморья до Китая и Японии, растет он также в Африке и на Мадагаскаре. Это единственный вид птериса, который дико произрастает и в пределах СССР, по скалам в горных лесах Закавказья. Он был найден также на стенах старых домов в Крыму.
На окраинах мангровых болот по тропическим побережьям земного шара обитает другой интереснейший представитель подсемейства - акростихум (Acrostichum). Он достигает в высоту 1,5-2 и иногда даже 3-4 м и встречается среди других своеобразных мангровых растений, в зарослях низкорослой пальмы нипа (Nipa) или, наконец, образует чистые заросли на расчищенных открытых местах, столь высокие и плотные, что возобновление других растений в них затруднено. Места произрастания этого папоротника подвергаются регулярному затоплению вследствие морских приливов, и акростихум приспособился к перенесению затопления, засоленности и бедности кислородом грунта. Но он, по-видимому, не является настоящим галофитом, так как может жить также в пресноводных озерцах и болотах, в местах, которые ранее подвергались воздействию приливов, а затем были отрезаны от моря.
Акростихум имеет массивное прямое или восходящее корневище, прочно удерживающее его в субстрате. Корневище покрыто жесткими чешуями и толстыми мясистыми корнями. Фертильные и стерильные листья мало разли-чаются между собой: и те и другие перистые с крупными цельными сегментами (с сетчатым жилкованием), с большим количеством устьиц на их поверхности. У фертильных листьев все сегменты или только верхние из них с нижней стороны густо покрыты перемешанными с парафизами буроватыми спорангиями, и снизу листья кажутся ржавыми. Крупные тетраэдрические споры акростихума попадают иногда и на скалистые морские побережья, и молодые растения вырастают на скалах, но не достигают здесь сколько-нибудь значительных размеров.
Под названием Acrostichum раньше объединяли несколько родов папоротников с "акростихоидным" расположением спорангиев. Сейчас установлено, что такое расположение спорангиев возникало независимо в разных неродственных эволюционных линиях и папоротники с акростихоидными спорангиями, разделенные на несколько родов, включают в разные семейства. В род акростихум теперь включают 3-5 близкородственных, видов, положение которых в семействе адиантовых определяется не только их морфологическим строением (трилетными спорами, типом развития и строения несимметрично-сердцевидного голого гаметофита), но и хромосомным числом, равным 30.
Подсемейство цератоптерисовые (Ceratopteridoideae)
Единственный род этого подсемейства цератоптерис (Ceratopteris) распространен в тропических и субтропических районах земного шара. Цератоптерисы - водные или болотные растения, либо свободно плавающие в водоемах вместе с салъвинией (Salvinia), пузырчаткой (Utricularia), водным гиацинтом (Eichhornia crassipes) и другими водными растениями, либо укореняющиеся в илистых почвах мелководных водоемов, в канавах, часто среди зарослей ситняга (Eleocharis), кувшинок (Nymphaea) и ряски (Lemna). Характерны они также для периодически затапливаемых почв, являясь сорняками на рисовых полях, в посадках таро (Colocasia esculenta). Несмотря на то что цератоптерис очень широко распространен, однообразие условий, в которых он существует, не благоприятствует интенсивному видообразованию, и все разнообразие форм в пределах этого рода многие птеридологи сводят к одному виду - цератоптерису василистниковидному (Ceratopteris thalictroides, рис. 112); другие же различают 4-5 видов.
Рис. 112. Цератоптерисы. Цератоптерис василистниковидный (Ceratopteris thalictroides): вверху - общий вид (видны два типа листьев); внизу слева - фрагмент спорофилла с одиночными спорангиями. Цератоптерис рогатый (С. cornuta): внизу справа - стерильный лист (вегетативное размножение)
Поднимающиеся над водой или лежащие на ее поверхности в виде розетки листья цератоптериса несколько напоминают листья некоторых зонтичных. Они тонкие и нежные, перисторассечены, с сегментами разной ширины. Фертильные и стерильные листья различны: первые более рассечены (до четырежды раздельных), с очень узкими конечными сегментами. Разнообразие формы листьев, присущее цератоптерису, складывается в процессе его развития. Первые листья мелкие, слабо рассеченные, с сетчатым жилкованием; последующие листья крупнее, длиной до 50 см. Рассеченность пластинки; с ростом; усиливается, становясь максимальной у фертильных листьев, которые продуцирует зрелое растение. Узкие сегменты фертильных листьев имеют почти параллельные жилки с небольшим количеством анастомозов. Вдоль крайних из этих жилок располагаются рядами одиночные спорангии, защищенные завернутыми краями сегментов, почти смыкающимися у молодых фертильных листьев.
Как и у многих других водных растений, в мезофилле листьев цератоптериса имеется большое количество воздушных полостей, а вдоль крупных мясистых листовых черешков проходят продольные воздушные каналы. Полны воздухом и его толстые корни, отходящие от прямого короткого корневища. Они не свисают вниз, а как бы распластаны под корневищем.
Корневища, молодые листья и черешки у их основания снабжены тонкими сидячими нерешетчатыми чешуями. У основания чешуй на черешках. Иногда имеются воздушные пузырьки, увеличивающие плавучесть растения.
Цератоптерис интересен не только водным образом жизни, то также жизненным циклом. Это однолетник, по крайней мере на затапливаемых полях, и как бы приспособлен к их циклу. Молодые растения укоренены в почве. При затоплении поля они становятся свободно плавающими и активно размножаются вегетативно. В синусах листовых сегментов, на верхушках листьев, иногда на рахисе появляются многочисленные мелкие выводковые почки, развивающиеся в новые молодые растения, долго остающиеся связанными с материнским. Освобождаются они после загнивания или обламывания листа. У основания черешка материнского листа часто развивается пучок придаточных корней, предназначенный для дальнейшего питания молодых растений. Когда поле высыхает, папоротники увядают и погибают, возобновляясь из спор на следующий сезон. Споры их прорастают на обнаженных участках почвы или под водой. По наблюдениям индийских ученых С. Пала и Н. Пала, однолетний цикл свойствен цератоптерису и в невысыхающих мелководных водоемах, в которых он ослабевает и загнивает с наступлением холодного сезона.
Благодаря своей плавучести цератоптерисы способны выживать во время сильных разливов рек, подъемов воды, образуя плавучие популяции и даже завоевывая при этом новые жизненные пространства. Единично занесенные в водоем, они способны через несколько лет широко распространиться на влажных местообитаниях окрестностей этого водоема.
Но насколько быстро в одних местах ареал цератоптериса расширяется, настолько быстро в других он может сокращаться при изменении условий среды, нарушениях местообитаний вследствие мелиорации, сокращения культуры риса и таро, вырубки деревьев и уничтожения других растений, затеняющих поверхность водоема.
Цератоптерис имеет некоторое практическое значение. Молодые экземпляры с нежными мясистыми сочными листьями употребляют в пищу как салат. Кроме того, цератоптерис часто выращивают в бассейнах и аквариумах. Благодаря своей декоративности и быстрому вегетативному размножению он приобрел большую популярность у аквариумистов. В аквариумах (по данным М. Д. Махлина) крупные экземпляры могут прожить и дольше года, иногда более трех лет, а дочерние растения легко перезимовывают при подсветке электролампами.
Положение рода в семействе адиантовых дискуссионно. Своеобразие условий, в которых он обитает, наложило отпечаток на его анатомическое и морфологическое строение. Он сохраняет в своем строении черты как адиантовых, так и общего предкового ствола, от которого разошлись цератоптерисовые и остальные адиантовые. Крупный сферический почти сидячий спорангий рода цератоптерис с прерванным вертикальным кольцом из 40-80 утолщенных клеток рассматривается иногда как упрощенный спорангий хейлантоидных папоротников, но кольцо у цератоптериса частично утратило свою функцию, оно служит только для вскрывания спорангия, но не способствует рассеиванию спор, так как не возвращается резко в первоначальное положение, как это происходит у других папоротников. Спорангий часто разрушается и без участия кольца, высыхая и высыпая крупные споры, плавающие или погружающиеся на дно. Строением трилетных, с гребневидным узором на экзине спор цератоптерис напоминает некоторые схизейные (роды Anemia, Mohria). Общие с последними черты имеет цератоптерис и в строении примитивного, часто двудомного гаметофита, который здесь сердцевидный, асимметричный на ранней стадии развития, голый, с бесцветными ризоидами и с погруженными антеридиями.
Своеобразие признаков рода побуждает некоторых птеридологов выделять цератоптерис в отдельное семейство, в пользу чего говорят и характерные для него хромосомные числа n = 39 и 40.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'