Порядок полиподиевые (Polypodiales)
Семейство глейхениевые (Gleicheniaceae) (Т. Н. Попова)
Около 160 видов глейхениевых распространены во влажных, нередко горных районах тропиков и субтропиков, а также в умеренных
широтах южного цолушария. Северная граница их распространения проходит через Японию в восточном полушарии и Мексику - в западном. Южная граница продвинута гораздо дальше от экватора - до Магелланова пролива. Особенно много глейхениевых в Малезийской флористической области и в тропической Америке. Отсутствуют глейхениевые ныне в Северной Африке, Европе, Западной и Центральной Азии и в Северной Америке (исключая Мексику), но были обильны на этой территории в прошлые геологические времена. Подчеркивая контраст между нынешним и былым распространением глейхениевых, английский палеоботаник А. Сьюорд сообщал, что он собирал листья глейхении дважды: на опушке тропического малайского леса и в меловых отложениях Гренландии с ее ныне суровым климатом.
Представители семейства - светолюбивые травянистые растения, наземные, иногда вьющиеся. Характерным признаком глейхениевых является наличие довольно крупных продольно раскрывающихся спорангиев с косым кольцом. Тип спорангия, простая структура корневищ, наличие волосков (а не чешуй) у многих видов свидетельствуют о примитивности семейства. Как и у схизейных, спорангии не имеют покрывальца, но на ранних стадиях обычно защищены волосками или чешуями. Расположены спорангии на нижней поверхности перышек, образуя сорусы по жилкам в их средней части, реже на их концах. Число спорангиев в сорусе чаще всего невелико: 2-5 у глейхении (Gleichenia), 8-15 или более, иногда группирующихся в виде полушария, у дикраноптериса (Dicranopteris) (рис. 113, 114). Все спорангии в одном сорусе развиваются одновременно. Споры трилетные.
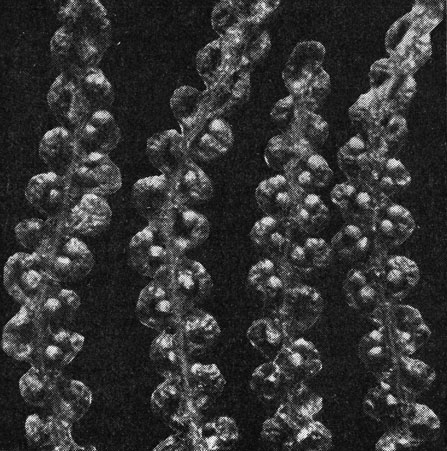
Рис. 113. Спорангии на нижней поверхности сегментов листа глейхении двуплодной (Gleichenia dicarpa)
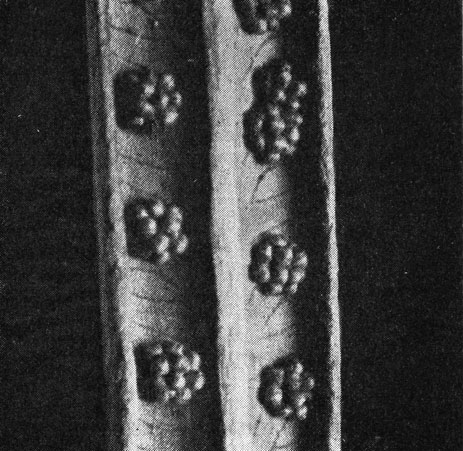
Рис. 114. Сорусы на нижней поверхности листа дикраноптериса линейного (Dicranopteris linearis)
До середины нынешнего столетия в это семейство (и даже в род глейхения) включали своеобразный ксерофильный папоротник платизому мелколистную (Platyzoma microphyllа), эндемик Квинсленда (северо-восток Австралии). В настоящее время принято выделять его в монотипное семейство платизомовых (Рlаtyzomataceae) по наличию ряда специфических признаков: длинные многоклеточные простые волоски на корневищах, тесно сближенные листья двух типов тонкие нитевидные и перистые, короткие волоски или сосочки по краю перьев, своеобразные одиночные спорангии на концах жилок, диморфизм спор (каждый спорангий содержит либо 16 крупных спор, либо 32 споры вдвое меньшего размера).
Семейство делят на два подсемейства: собственно глейхениевые (Gleichenioideae), включающие 2 рода - глейхения со 150 видами и дикраноптерис с 10 видами, и строматоптерисовые (Stromatopteridoidaeae) с единственным новокаледонским родом строматоптерис (Stromatopteris). Общим для всех их является наличие сорусов, лишенных покрывалец, с небольшим числом спорангиев, раскрывающихся продольно. Некоторые современные птеридологи рассматривают эти подсемейства как самостоятельные семейства.
У представителей подсемейства глейхениевых ползучие разветвленные тонкие корневища дорсивентрального типа достигают длины нескольких метров. Листья также обычно большие, дважды или трижды перистые.
Папоротники подсемейства глейхениевых имеют очень характерные ложнодихотомически разветвленные листья с неограниченным ростом в длину. Роды и подроды подсемейства различают по характеру ветвления листьев и разнообразию эпидермальных придатков. Глейхендям свойственны бахромчатые щитовидные чешуи и звездчатые волоски, а дикраноптерис характеризуется разветвленными волосками различной формы; чешуй нет (рис. 115).
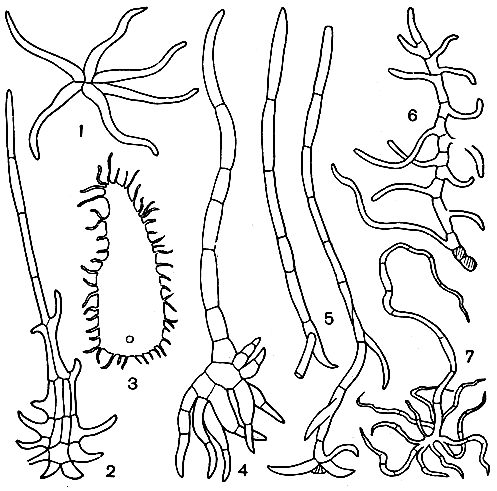
Рис. 115. Волоски и чешуи глейхениевых: 1 - звездчатый волосок глейхении длинной (Gleichenia longissima); 2 - чешуя рахиса глейхении щетинистой (G. hispida); 8 - чешуя верхушки рахиса глейхении усеченной (G.truncata); 4 - волосок верхушки главного рахиса дикраноптериса Куррана (Dicranopteris curranii); 5 - волоски нижней поверхности листа дикраноптериса опушенного (D. pubigera); 6, 7 - то же дикраноптериса линейного (D. linearis)
Гаметофит у глейхении и дикраноптериса зеленый наземный, талломного типа (сердцевидная, позже лентовидная пластинка).
Род глейхения подразделяют главным образом по типу ветвления листа на подроды: глейхения (Gleichenia), мертензия (Mertensia) и диплоптеригиум (Diplopterygium). Некоторые исследователи рассматривают их как самостоятельные роды.
Лист у рода глейхения перистый, однако выглядит, как дихотомически разветвленный, так как верхушечный рост листа периодически прекращается. Стержень заканчивается спящей почкой и дальнейшее развитие листа происходит благодаря удлинению перьев, стержни которых также заканчиваются спящими почками и т. д. Степень рассеченности конечных долей перьев, относительная длина стержней первого и конечного порядков, дополнительные перья у основания ветвей второго порядка, а также другие признаки создают значительное разнообразие формы листьев, длиной у одних до нескольких метров, а у других - до 30 см.
Представители наиболее обширного подрода мертензия имеют небольшие размеры. Обычно это горные растения, встречающиеся среди кустарников и в низкорослых горных лесах, на открытых травянистых или скалистых местах как в тропиках, так и в южных умеренных районах. Многие виды встречаются в саваннах Юго-Восточной Африки и Южной Америки, на скалах и вересковых пустошах в Андах и на гористых островах Тихого океана. В горах на высотах от 750 до 2250 м в поясе медленно растущих альпийских кустарников глейхении из этого подрода образуют низкие заросли или встречаются поодиночке на сфагновых участках и на скалах.
Многие глейхении из подрода диплоптеригиум обычны вдоль дорог и на кокосовых плантациях. Если образуется низкорослый вторичный лес, они могут оставаться в жизнеспособном состоянии, и их листья достигают значительной длины (до 6 м). Опираясь на кустарники и небольшие деревья, папоротники поднимаются на высоту до 3-5 м.
Представитель второго рода подсемейства дикраноптерис буро-желтый (Dicranopteris fulva) образует на острове Ямайка столь густые и высокие заросли, что людям порой приходится прорубать в них своего рода туннели. Местами переплетенные листья папоротника покрывают почву пружинящим настилом метровой толщины.
Хотя дикраноптерисы и светолюбивы, их корневища нуждаются в защите от солнца и сухости. Только благодаря быстрому росту они могут укрепляться в качестве растений-пионеров на голой земле, очень сильно прогреваемой солнцем в тропиках. Обычнейший тропический вид дикраноптерис линейный (D. linearis) имеет такие длинные листья, что они не могут сохранять прямостоячее положение и образуют труднопроходимые запутанные заросли по берегам речек, на склонах оврагов. Другим растениям практически невозможно проникнуть на территории, занятые этим папоротником (рис. 116). Его сравнивают в этом отношении с орляком (Pteridium). Заросли дикраноптериса могут существовать продолжительно, но если среди них удастся выжить сеянцам деревьев, то со временем они смогут затенить светолюбивый пацрротцик, он погибнет и произойдет следа одного, растительного сообщества другим. В особо влажных условиях на островах Адмиралтейства вьющийся дикраноптерис линейный был обнаружен на верхушках высоких деревьев 50-метровой высоты.
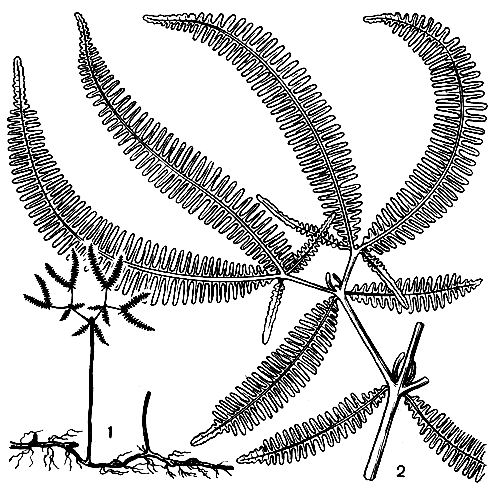
Рис. 116. Дикраноптерис линейный (Dicranopteris linearis): 1 - общий вид; 2 - часть листа
Стержни листьев некоторых видов дикраноптерисов применяются на полуострове Малакка в рыболовстве для изготовления ловушек: они стойко выдерживают двухлетнее пребывание в морской воде. Кроме того, гибкие и крепкие части стержня используются для изготовления плетеных изделий. Перистые, гребенчато надрезанные листья глейхении служат местному населению тихоокеанских островов как украшения.
Относящийся к подсемейству строматоптерисовых единственный вид - строматоптерис четкообразный (Stromatopteris moniliformis) встречается в Новой Каледонии как на сухих освещенных участках, так и в тенистых дождевых лесах на различных субстратах от листового перегноя до почти чистой глины.
Подземная часть спорофита представлена многократно дихотомически ветвящимися вертикальными и несущими листья горизонтальными корневищами. Те и другие покрыты щитовидными чешуями, от тех и других отходят ризриды и редкие корни. Интересной особенностью корней является отсутствие корневых волосков и наличие покрытой кутикулой ризодермы из толстостенных клеток. В корневищах поселяется эндофитный гриб, отсутствующий в корнях.
Надземную часть спорофита строматоптериса составляют ксероморфные листья длиной 15-30 см, первоначально улиткообразно закрученные. В отличие от собственно глейхениевых, они однажды перистые (рис. 117), прямостоячие. Перья полуэллиптические, длиной 0,5-3 см, с завернутыми краями. На абаксиальной стороне располагаются сорусы со спорангиями, их от 6 до 33 (обычно их 25-30). Споры монолетные.
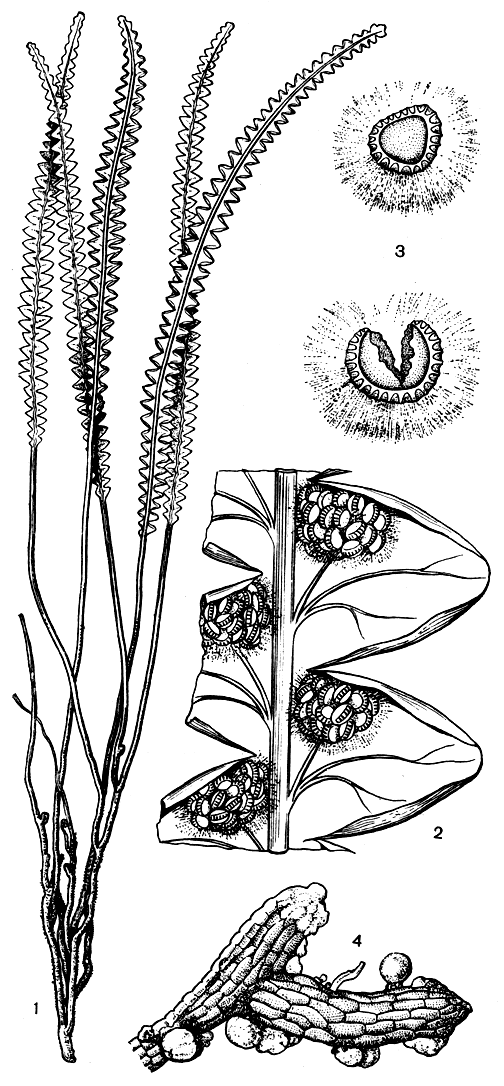
Рис. 117. Строматоптерис четкообразный (Stromatopteris moniliformis): 1 - общий вид; 2 - часть нижней поверхности листа с сорусами; 3 - сорусы; 4 - гаметофит (часть)
Гаметофит строматоптериса подземный, бесцветный, длиной около 2 см. Гаметангии и ризоиды на его поверхности распределены равномерно.
Семейство матониевые (Matoniaceae) (В. Н. Гладкова)
"Мы пришли на "Паданг-Бату", или каменное поле... Мы нашли здесь крутой склон плоских утесов... Местами склон этот был совершенно обнажен, но местами, там, где утесы растрескались и торчали зубцами, развилась роскошная растительность... Здесь встретились нам впервые шишконосные из рода Dacrydium, а в лесу над самыми утесами нам встретилась роща из великолепных папоротников, Dipteris horsfieldii и Matonia pectinata с широкими листьями в виде опахала, на тонких стволах, вышиной от 6 до 8 футов. Самый высокий и красивый из них - Matonia - известен только на этих горах и не появлялся до сих пор в наших теплицах",- так описывал свои впечатления от этих папоротников известный натуралист, современник Дарвина Альфред Уоллес, совершивший восхождение на гору Офир в центре полуострова Малакка (Малайский архипелаг, 1872, в русском переводе под редакцией А. Бекетова).
И в наше время почти не удается выращивать в теплицах матониевые, так как условия их обитания, как и сами эти папоротники, очень своеобразны.
Матониевые появились на Земле в позднем триасе и были широко распространены в мезозойскую эру. Самым древним из них являлся род флебоптерис (Phlebopteris). Ископаемые остатки (поздний триас - ранний мел) его видов находят почти на всех континентах (кроме Южной Америки) от 70 ° с. ш. в Гренландии до 30° ю. ш. в Восточной Австралии, причем наибольшего развития этот род достигал в Европе. После его вымирания ареал матониевых значительно уменьшился и распался на 2 части: западную и восточную. В восточной части ареала матониевые сохранились до наших дней.
В настоящее время матониевые представлены двумя родами и четырьмя видами в горах Юго-Восточной Азии - на полуострове Малакка, на Суматре, Калимантане, Молуккских островах (Амбоина) и в Новой Гвинее. Местонахождения отдельных видов удалены друг от друга иногда на тысячи километров, Изолированные горные вершины названных островов являются последними убежищами этой вымирающей группы растений.
Два близких вида, относящиеся к роду матония,- крупные папоротники с длинными, ползучими, ветвящимися корневищами, покрытыми блестящими коричневыми волосками. Проводящая система корневищ представлена полициклической амфифлойной сифоностелой, состоящей у старых корневищ из двух или трех концентрических цилиндров, связанных листовыми следами (рис. 119). Молодые корневища имеют протостелу, которая превращается в сифоностелу по мере развития корневищ.
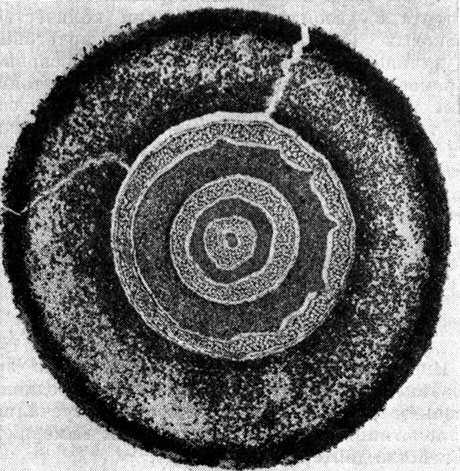
Рис. 119. Поперечный срез корневища матонии гребневидной (Matonia pectinata); видна полициклическая сифоностела
Листья матоний располагаются на корневище на большом расстоянии друг от друга; они крупные, кожистые, на длинных черешках. Пластинка листа разделена на 2 половины, каждая из которых состоит из множества гребенчато расположенных перистых сегментов. Такая структура листа возникает при последовательном ветвлении по типу завитка стержней сегментов, в результате чего внутрь каждый раз отчленяются новые сегменты, а снаружи продолжается ось. Жилки на сегментах листа свободные, анастомозируют обычно только в местах расположения сорусов, образующих один или несколько рядов по обеим сторонам средней жилки.
Матония гребневидная (Matonia pectinata) - наиболее широко распространенный представитель современных матониевых. Гораздо более редкими являются фанеросорусы (Phanerosorus). Два вида фанеросоруса растут на севере острова Калимантан и в Новой Гвинее, где они обычно селятся на известняках. Эти папоротники отличаются повисающими длинными листьями, ветвление которых характеризуется образованием покоящихся почек на некоторых ветвях и очень напоминает ветвление листьев глейхении. Сходство с глейхениевыми проявляется у матониевых не только в ветвлении листьев, но и в строении сердцевидного гаметофита с ризоидами и большим количеством боковых крыльев, в строении архегониев с длинной искривленной шейкой, в основном хромосомном числе х = 13. Похожи матониевые на глейхениевые и по строению сорусов, состоящих из немногочисленных, крупных, почти сидячих спорангиев с косым кольцом, в отличие от глейхениевых, снабженных зонтиковидным покрывальцем (но некоторые ископаемые матониевые покрывальца не имели). Такое сходство говорит о родстве матониевых и глейхениевых, произошедших, вероятно, от общего предка.
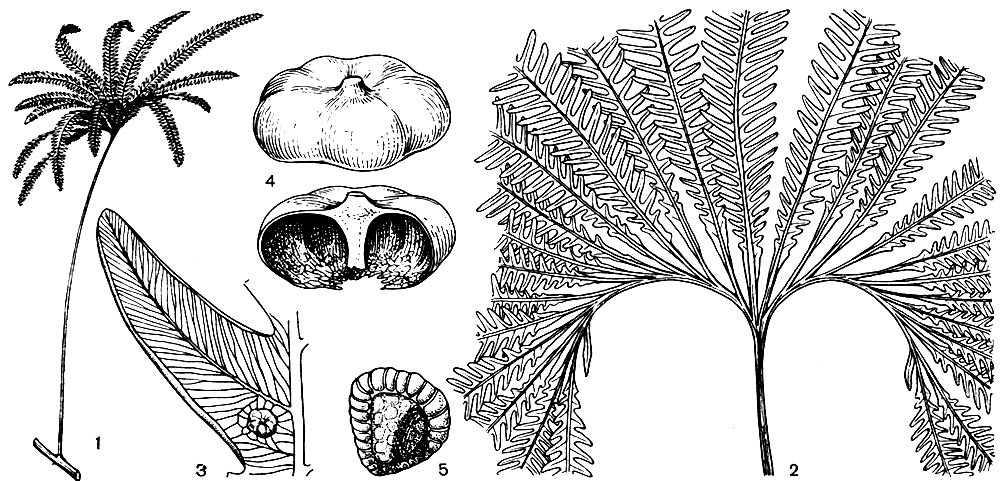
Рис. 118. Матония гребневидная (Matonia pectinata): 1 - общий вид; 2 - основание пластинки листа; 3 - сегмент с сорусами; 4 - покрывальце; 5 - спорангий
Семейство диптерисовые (Dipteridaceae) (В. Н. Гладкова)
Вместе с матониевыми в мезозойскую эру (начиная с триаса, а по некоторым предположениям даже с перми) широко встречались папоротники другого, сейчас почти вымершего семейства диптерисовых.
До наших дней дожил только один род этого семейства - диптерис (Dipteris) с восемью видами, область распространения которых простирается от Северо-Восточной Индии, Южного Китая и Тайваня через Малайский архипелаг до Новой Гвинеи, Новой Каледонии и Полинезии. Диптерисы - крупные наземные папоротники с длинными ползучими волосистыми корневищами и огромными листьями, пластинка которых подобно пластинке листа матонии дихотомически рассечена на две симметричные половины, а они, в свою очередь, разделены на лопасти (рис. 120). Главные жилки в каждой половинке листа дихотомически повторно ветвятся, а ветви их связаны сетью более мелких жилок, отходящих под прямым углом от них и друг от друга. В местах соединения самых тонких из этих жилок лежат мелкие округлые сорусы, лишенные покрывальца. Иногда зрелые сорусы почти сливаются, и перемешанные с парафизами спорангии заполняют большую часть поверхности листа. Спорангии имеют ножку из четырех рядов клеток и снабжены почти вертикальным полным или неполным кольцом без выраженного стомия.

Рис. 120. Диптерис Валлиха (Dipteris wallichii)
Черешки листьев диптериса двойчатосложного (D. conjugata), самого распространенного из видов диптериса, бывают длиной более 1 м, причем достигают почти полной своей длины до того, как лист полностью развернется. При разворачивании листа стержень каждой полупластинки у основания перекручивается на 90° обеспечивая свободное горизонтальное положение ее в пространстве и оптимальное использование света.
Диптерис двойчатосложный, как и другие диптерисы, растет на открытых местах в горах, реже на равнинах, на лесных полянах, вдоль горных тропинок, иногда на прибрежных скалах. Он встречается по берегам рек, достаточно широких, чтобы образовать просвет в лесном пологе. Иногда места его произрастания подвергаются затоплению при внезапном быстром разливе рек, папоротники тогда оказываются полностью погруженными в воду.
Некоторые роды ископаемых диптерисовых очень напоминают современные виды диптериса. Например, виды рода хаусмания (Hausmannia) имели неглубоко-лопастные листья, почти не отличавшиеся от листьев современного диптериса новогвинейского (Dipteris novoguineensis). Другие ископаемые роды характеризовались более сильной рассеченностью листьев. Оригинальное устройство листа было свойственно камптоптерису спиральному (Camptopteris spiralis), крупные листья которого были вильчато разделены на две направленные вверх ветви. От каждой из них по спирали расходились жестко-кожистые, длинные (до 50 см) сегменты.
По общему плану строения листьев современные и ископаемые диптерисовые близки к матониевым. Сходство с последними диптерисовые имеют и в строении проводящей системы корневищ (сифоностела). С другой стороны, по характеру жилкования, строению монолетных спор и сорусов современные диптерисовые близки к полиподиевым, отличаясь от них основным хромосомным числом (х = 33), более примитивными спорангием и гаметофитом.
Семейство полиподиевые, или многоножковые (Polypodiaceae) (А. Е. Бобров)
Семейство полиподиевые является одним из наиболее богатых среди папоротников: оно объединяет около 50 родов и приблизительно 1500 видов. Листья расположены двурядно на верхней стороне сочных, мясистых корневищ, покрытых чешуями и волосками (рис. 121). По характеру жилкования пластинки, степени ее рассеченности, морфологии чешуй и волосков внутри семейства установлен ряд подсемейств. Это дринариевые (Drynarioideae), микросориевые (Microsorioideae), полиподиевые, или многоножковые (Polypodioideae), платицериевые (Platycerioideae) и плеопельтисовые (Pleopeltoideae).
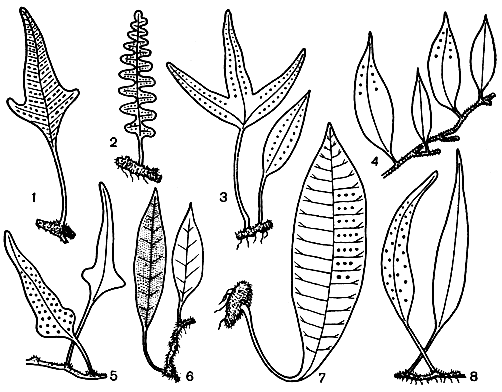
Рис. 121. Представители полиподиевых: 1 - фиматодес Левелье (Phymatodes levellei); 2 - полиподиум обыкновенный (Polypodium vulgare); 3 - крип синус копьевидный (Crypsinus hastatus); 4 - леммафиллум грушеобразный (Lemmaphyllum pyriforme); 5 - неохейроптерис полукопьевидный (Neocheiropteris subhastatus); 6 - пиррозия языкообразная (Pyrrosia lingua); 7 - колизис (Colysis); 8 - леписорус уссурийский (Lepisorus ussuriensis)
Характерным признаком семейства является расположение эллиптических или округлых сорусов (без покрывалец) на нижней стороне пластинки листа. У ряда видов полиподиума (Polypodium) наблюдаются и абортивные спорангии, получившие название парафиз. Сорусы по периферии окружены клиновидными и радиальными чешуями и волосками. У некоторых родов плеопельтисовых стенки чешуй сильно утолщены и напоминают решетку, а у платицериевых отмечены звездчатые волоски.
Спорангии полиподиевых имеют меридионально расположенное кольцо раскрывания, состоящее из 13-14 толстостенных клеток и прерывающееся щелью, отграниченной двумя клетками. Через щель (стомий) выбрасываются споры. Ножка спорангия состоит из двух рядов клеток.
Ископаемые представители семейства известны уже из отложений позднетриасового периода. В настоящее время это семейство распространено по всему земному шару, но в основном в тропиках Старого Света. Чаще всего это эпифитные растения, поселяющиеся на стволах, а также на толстых ветвях и даже на верхушках деревьев. Некоторые из них являются облигатными (обязательными) эпифитами, тогда как другие могут произрастать как на скалах и на почве, так и на стволах деревьев, т. е. являются факультативными (необязательными) эпифитами.
Как правило, не наблюдается строгой приуроченности эпифитов из полиподиевых к определенным видам растений-хозяев: они встречаются на деревьях различной систематической принадлежности, обладающих в большинстве случаев толстой шероховатой корой.
Наблюдения в природе показали, что в результате произрастания эпифитов (в том числе и папоротников) на поверхности коры дерева повышается влажность. Соответственно на этих участках увеличивается количество бактерий, грибов и насекомых, и это, в свою очередь, отражается на жизни эпифитов, вызывает появление у них различных и подчас поразительных приспособлений к совместному существованию.
Из насекомых наиболее часто на различных видах тропических полиподиевых встречаются муравьи, из родов ацтека (Azteca), фиодола (Phiodola), соленопсис (Solenopsis), кампонотус (Camponotus) и др. Их взаимоотношения с эпифитами (и наземными представителями) из полиподиевых, такими, как соленоптерис (Solenopteris), леканоптерис (Lecanopteris) и полиподиум (Polypodium), сложны и многообразны. Но то, что в корневищах этих папоротников поселяются муравьи, по крайней мере оберегает их от уничтожения крупными животными. Муравьи также извлекают пользу: они получают пристанище и богатый крахмалом, сахарами и другими питательными веществами корм.
У центральноамериканских представителей соленоптериса, а также у леканоптериса из Юго-Восточной Азии следствием симбиоза является разрастание паренхиматических тканей корневища, образующих клубневидные утолщения. Соленоптерис получил даже название "папоротник-картофель" из-за сходства его клубнеобразно утолщенных корневищ с клубнями картофеля. Есть указания на то, что в полостях таких клубневидных утолщений размещаются кладки яиц, куколки, личинки, а также королевы муравьев. Взрослые муравьи поедают сладкие паренхимные клетки, выедая клубни изнутри до такой степени, что они становятся проницаемыми для воды. Частичная редукция сосудистого тяжа корневища сопровождается образованием адсорбирующих волосков, заполняющих полость клубней. Проницаемые для воды участки развиваются в верхней части мясистых клубнеобразных корневищ, при этом папоротник приобретает характер так называемых "цистерновых" эпифитов типа бульбофиллума (Bulbophyllum) из орхидных.
Особенности мест произрастания эпифитных полиподиевых определяют их приспособления к дефициту влаги. Лишь 10% эпифитных полиподиевых произрастает в ксерофитных тропических лесах с сухим воздухом и малым количеством влаги. Для папоротников этих типов тропического леса характерны кожистая редуцированная листовая пластинка, толстая кутикула и приспособления для гидрорегуляции в виде решетчатых чешуй, как у леписоруса, или звездчатых волосков, как у пиррозий (Pyrrosia). Большая часть видов полиподиевых (65%) встречается в мезофитных и гигрофитных тропических лесах с высокой относительной влажностью воздуха и положительной в течение всего года температурой, а также в высокогорных лесах пояса туманов. Эти папоротники обладают рассеченной и слабокожистой пластинкой.
Интересны адаптации многоножковых из различных подсемейств к сухому климату. Большинство из них уменьшает поверхность испарения в результате свертывания листовой пластинки. В одних случаях пластинка свертывается на нижнюю сторону и спорангии оказываются заключенными внутри трубки, как у леписоруса, тогда как в других случаях пластинка сворачивается кверху и спорангии выворачиваются наружу, вместе с чешуями, покрывающими нижнюю поверхность пластинки, как это наблюдается у многоножек. Кроме того, в сухое время года сами листья целиком буреют и высыхают. Вероятно, это сопровождается и разрушением хлорофилла. Но достаточно выпасть небольшому дождю, как листья разворачиваются, зеленеют, споры высыпаются из спорангиев и прорастают. Есть указания на то, что во влажных условиях споры многоножковых прорастают уже в спорангиях.
Из других биологических особенностей многоножковых следует указать на высокий процент аутополиплоидов (основное число хромосом у многих родов семейства 37), т. е. полиплоидов с увеличенным числом идентичных наборов хромосом того же вида.
Представители этого палеотропического (ареал в тропиках Старого Света) рода, насчитывающего около 20 видов, являются крупными (длиной до 1 м) эпифитами. Их листья расположены в виде широкой воронки или гнезда (гнездовые эпифиты). Листья диморфные - стерильные, похожие на лист дуба, и спороносные - перистые, прямостоячие или свешивающиеся, с хорошо выраженным твердым черешком и кожистой пластинкой (рис. 122).

Рис. 122. Дринария Мейена (Drynaria meyeniana): 1 - общий вид; 2 - спорангий; 3 - участок сегмента
Спорангии расположены рядами в верхней или средней, реже нижней части пластинки по ее краю или на нижней стороне. По расположению спорангиев и степени редукции пластинки род дринария подразделяется на несколько подродов, которым иногда придается родовой ранг. Это - псевдодринария (Pseudodrynaria), аглаоморфа (Aglaomorpha), дринариопсис (Drynariopsis) и др.
Большинство дринарий приурочено к горным лесам тропической Азии. Имеются указания на неприхотливость видов дринарий к характеру субстрата: так, дринария Лаурента (Drynaria laurentii) произрастала на протяжении ряда лет на алюминиевой крыше дома на горе Нимба (Гана). Несколько видов дринарий разводят в прохладных оранжереях ботанических садов как декоративные висячие растения.
Род полиподиум, насчитывающий 75 видов, представлен некрупными эпифитными и наземными формами со слабо кожистыми или перепончатыми перисторассеченными листьями. Толстое, ветвящееся корневище полиподиумов покрыто темно- и светло-коричневыми клиновидными чешуями; листья отходят от него рядами, с чем несомненно связано название рода - многоножка. Сорусы округлые или овальные и располагаются посредине или у края сегментов с нижней стороны пластинки (рис.123).
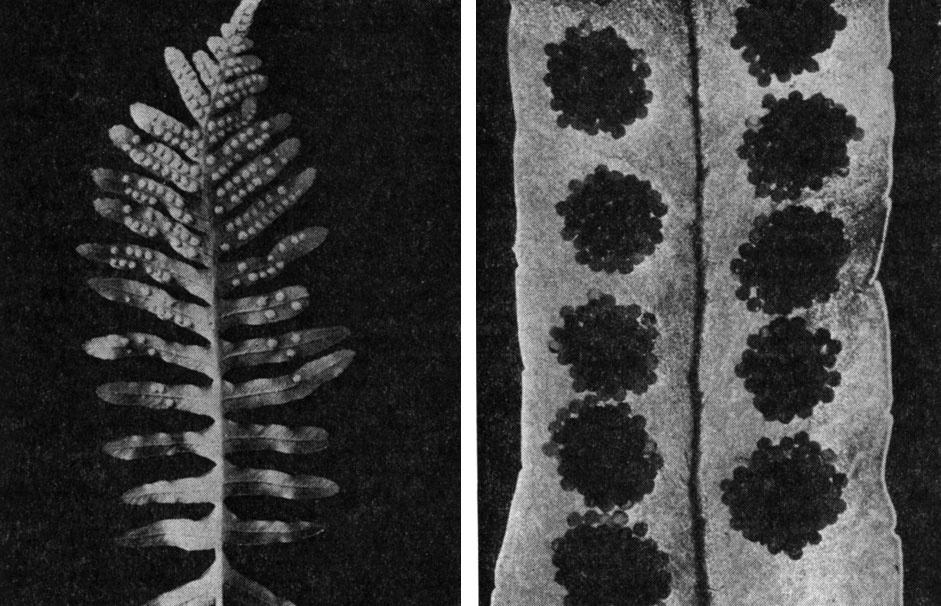
Рис. 123. Полиподиум обыкновенный (Polypodium vulgare): слева - общий вид листа; справа - сорусы
Виды рода - обитатели лесов тропической, реже субтропической и умеренной зон; они встречаются на стволах и ветвях деревьев, на скалах, а изредка на почве. В СССР распространено 6 видов.
Сочные и сладкие корневища полиподиума (по-немецки полиподиум называют "сладким корнем") содержат глюкозиды, яблочную кислоту и сапонины. Имеются указания на применение в медицине отвара листьев и корневищ многоножки.
Многочисленные садовые разновидности полиподиума обыкновенного (P. vulgare) и полиподиума южного (P. australe) нередко культивируют в оранжереях и открытом грунте.
В роде около 70 видов, распространенных в Восточной и Юго-Восточной Азии и на островах Тихого океана. Это некрупные (длиной до 30-40 см) эпифитные, реже наземные папоротники с цельными, перистыми и надрезанными листьями со слабо утолщенной срединной жилкой. Горизонтально расположенное ветвящееся корневище толщиной 0,4-0,8 см покрыто коричневыми клиновидными чешуями. Виды микросориума встречаются во влажных тропических лесах низменностей, в горных тропических лесах, у термальных источников, поднимаясь до верхней границы древесной растительности в горах тропической зоны.
Интересными биологическими особенностями обладает микросориум крылоножковый (Microsorium pteropus), распространенный от Индии до Индонезии, включая Южный Китай, Филиппины и Малайзию. От других видов рода он отличается трехлопастными листьями с низбегающей на черешок нижней частью пластинки листа. Этот вид произрастает нередко в горных тропических лесах, в тропических лесах низменностей, по берегам ручьев и источников, в местах, периодически затопляемых в период дождей, на камнях, гниющих стволах деревьев. Листья этого папоротника диморфные: развивающиеся под водой - цельные, а надводные - трехлопастные. На погруженных в воду листьях развиваются небольшие почки, которые позднее дают молодые особи, отделяющиеся от материнского растения.
В оранжерейных условиях микросориум крылоножковый размножают, как и другие многоножковые, делением корневища и проращиванием спор. Любители-аквариумисты разводят его в аквариумах.
Род объединяет 25 видов, распространенных в Азии. Это небольшие наземные и эпифитные формы с кожистыми и травянистыми линейными и линейно-ланцетными листьями. Сорусы эллиптические, по периферии прикрытые темно-красными решетчатыми радиальными чешуями на коротких ножках. Эти чешуи более заметны у молодых листьев, т. к. в зрелом состоянии часто отваливаются. Размеры чешуй у сорусов, на корневищах, а также степень развития склеренхимного тяжа варьируют у разных видов.
Леписорус произрастает на почве, скалах, на ветвях и стволах деревьев, поднимаясь в горы до 3000 м над уровнем моря. В СССР (на Алтае и на Дальнем Востоке) встречаются три вида леписоруса. Из них леписорус уссурийский (L. ussuriensis) на юге Приморского края обитает большей частью на замшелых скалах, а как эпифит встречается изредка и лишь на одиночных деревьях во влажных смешанных лесах. Однако был описан уникальный случай массового развития этого папоротника как эпифита в одной горной долинке в Уссурийском заповеднике ("долина эпифитов"). Здесь леписорус уссурийский встречается на большей части деревьев, нередко от основания ствола и почти до вершины.
Представители рода платицериум (известно 17 видов) произрастают в тропических лесах Старого Света. Это крупные эпифиты с диморфными листьями - стерильными и спороносными (табл. 25, 26).
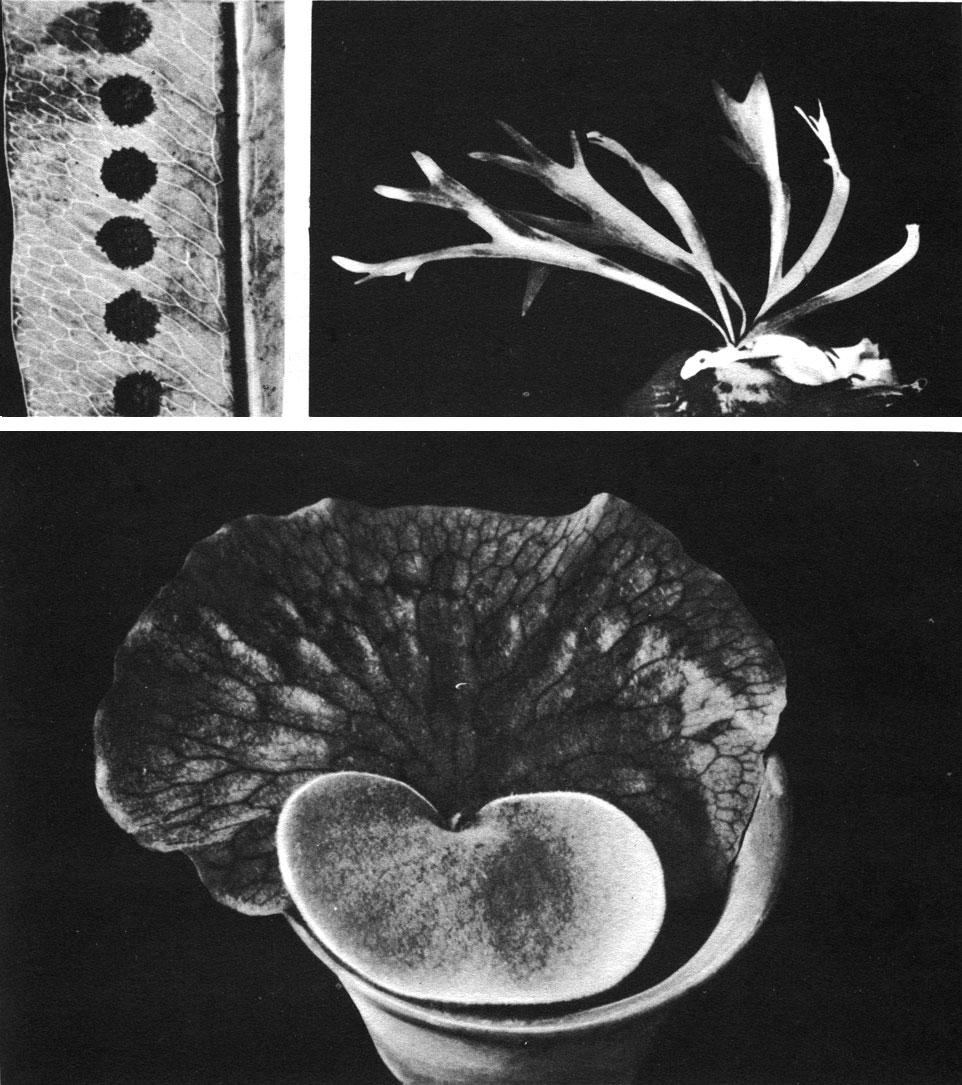
Таблица 25. Полиподиевые: вверху слева - фиматодес фиджийский (Phymatodes vitiensis), часть листа с сорусами; вверху справа - платицериум оленерогий (Platycerium bifurcatum); внизу - платицериум ангольский (P. angolense), молодое растение с лепешковидными стерильными листьями
Стерильные листья широко округлые, цельные или изрезанные наверху. Своим нижним и боковыми краями они тесно прижаты к стволу или ветви дерева, верхняя их часть отходит от опоры так, что образуется ниша, в которой постепенно накапливается перегной, пронизанный корнями этого удивительного эпифита. Перегной образуется не столько за счет попадающего в нишу опада от других растений, сколько главным образом из отмирающих старых стерильных листьев самого "оленьего рога", которые постепенно перекрываются разрастающимися новыми листьями. С возрастом ниша все более и более увеличивается и перегной в ней у наиболее мощных видов "оленьего рога" достигает иногда массы 100 кг.
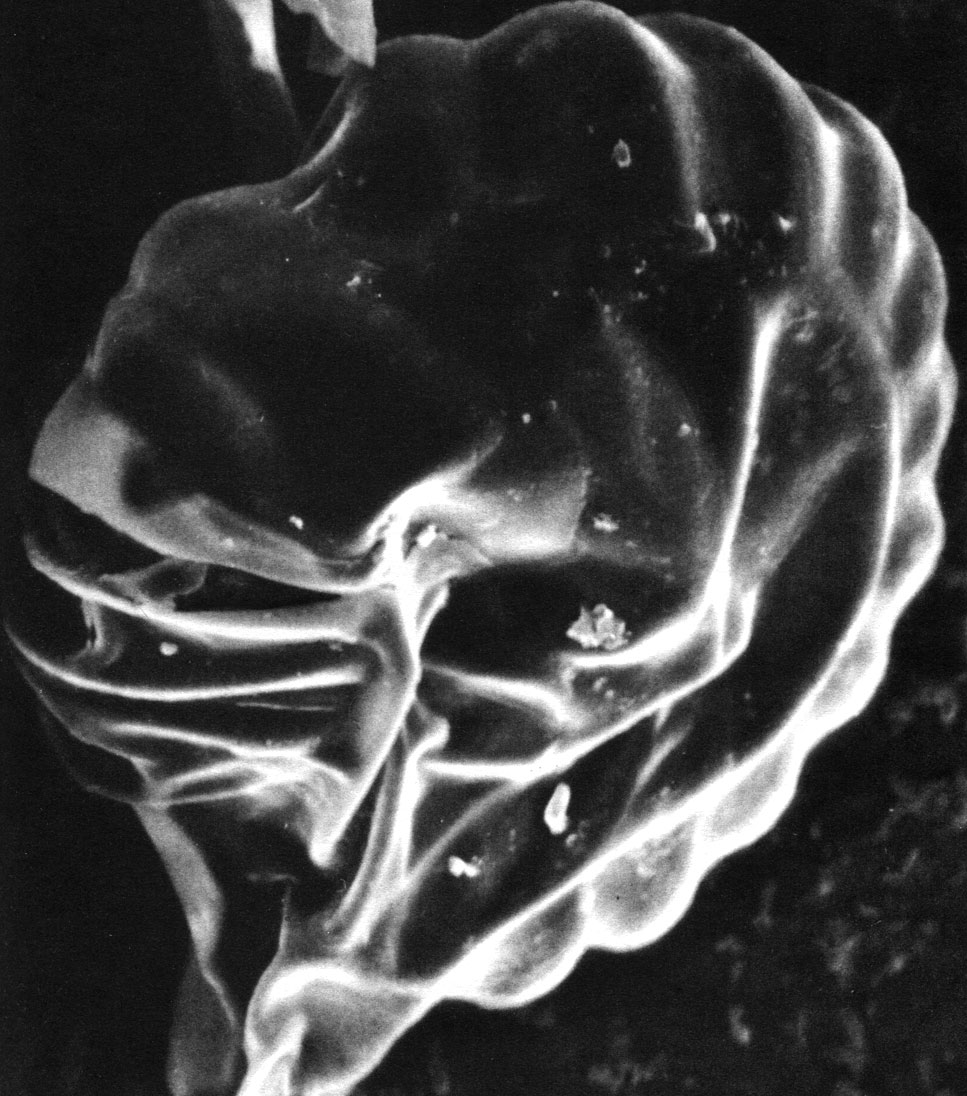
Таблица 31. Спорангий фиматодеса фиджийского (Phymatodes vitiensis) под сканирующим электронным микроскопом (увел, около 1250). Слева виден стомий
Наряду со стерильными у платицериумов развиваются листья совершенно иного строения и функционального значения. Прямостоячие или свешивающиеся, они напоминают по форме рога оленя, с чем и связано популярное название этих эпифитов "олений рог". Это ассимилирующие и спороносные листья. У папоротников, достигших зрелости, вся нижняя поверхность листа на концах его разветвлений покрывается кирпично-красным налетом из многочисленных спорангиев.
"Оленьи рога" достигают иногда огромных размеров. Так, обнаруженный на острове Страдброк (Восточная Австралия) экземпляр платицериума большого (Platycerium grande) имел в поперечнике 1,8 м. Под тяжестью таких эпифитов-гигантов, по словам Чемберлена, деревья иногда выворачиваются с корнями.
Виды рода платицериум произрастают в средней части больших ветвей, но чаще на коре стволов крупных отдельно стоящих вечнозеленых и листопадных деревьев. Нередко вместе с "оленьим рогом" встречаются и другие эпифитные папоротники, а также орхидеи и мхи.
Из-за своеобразия общего облика, а также строения спорангиев этот род выделяли в отдельное семейство, но особенности жилкования, строения гаметофитов и хромосом позволяют сближать его с родом полиподиум.
Эти красивые и оригинальные растения широко культивируют в прохладных оранжереях; молодые растения прикрепляют вместе с мхом к подвешенным под кровлей отрезкам стволов. Выведено несколько десятков садовых форм платицериумов. Виды платицериума хорошо реагируют на подкормку, а в холодное время года требуют меньшего и более редкого полива.
Семейство граммитисовые (Grammitidaceae) (Т. Д. Сурова)
Тропическое семейство граммитисовые объединяет около 500 видов, которые в течение многих лет входили в род полиподиум (Polypodium). Лишь в 1940 г. китайский ботаник Р. Цинь выделил эту группу в отдельное семейство, что было принято и большинством других птеридологов. К настоящему времени известно 12 родов этого семейства, три из которых - граммитис (Grammitis), ктеноптерис (Ctenopteris) и ксифоптерис (Xiphopteris) - объединяют 400 видов и считаются наиболее примитивными, а 9 других - производными от них.
Граммитисовые и полиподиевые различаются многими признаками как спорофита (чешуи на корневищах, жилкование, опушение, спорангии и споры), так и гаметофита (форма, опушение, особенности развития, длительность жизни). Пожалуй, наиболее характерным и постоянным является различие в строении спор: монолетных, не содержащих хлорофилл - у полиподиевых и трилетных, зеленых - у граммитисовых. Однако исследователи, не имеющие в своем распоряжении спор, могут с достаточной уверенностью отличить граммитисовые по одноклеточным тонким, но очень жестким игловидным волоскам, располагающимся на черешках, на поверхности листьев и между спорангиями. Волоски граммитисовых могут быть также многоклеточными, ветвящимися, с жесткими одноклеточными ветвями, но не мягкими или железистыми, как у большинства полиподиевых.
Граммитисовые - эпифитные или наскальные папоротники, растущие преимущественно в горных дождевых лесах на значительной высоте (1000-2000 м над уровнем моря) в зоне туманов. Они широко распространены в тропиках, а некоторые встречаются в умеренной зоне южного полушария. Граммитисовые входят в сообщества теневых эпифитов, включающие, кроме мхов и лишайников, многочисленные гименофилловые и некоторые мелкие полиподиевые. Все они живут в глубокой тени под пологом леса на стволах деревьев и древовидных папоротников, на мшистых камнях, иногда на плотном ковре из мхов и лишайников. Настоящих наземных папоротников в семействе граммитисовых нет, хотя некоторые из них от типично эпифитного обитания на стволах деревьев перешли к жизни в расщелинах скал в альпийском или субальпийском поясе гор в тропиках.
Граммитисовые - мелкие или средние по величине растения (длина листа у разных видов от 6 до 25 см), образующие благодаря очень коротким корневищам, несущим тесно расположенные листья, подобие дернины. Листья разнообразны по форме у разных родов. Степень рассеченности листа, жилкование и расположение сорусов - признаки, положенные в основу деления семейства граммитисовых на роды (рис. 124).
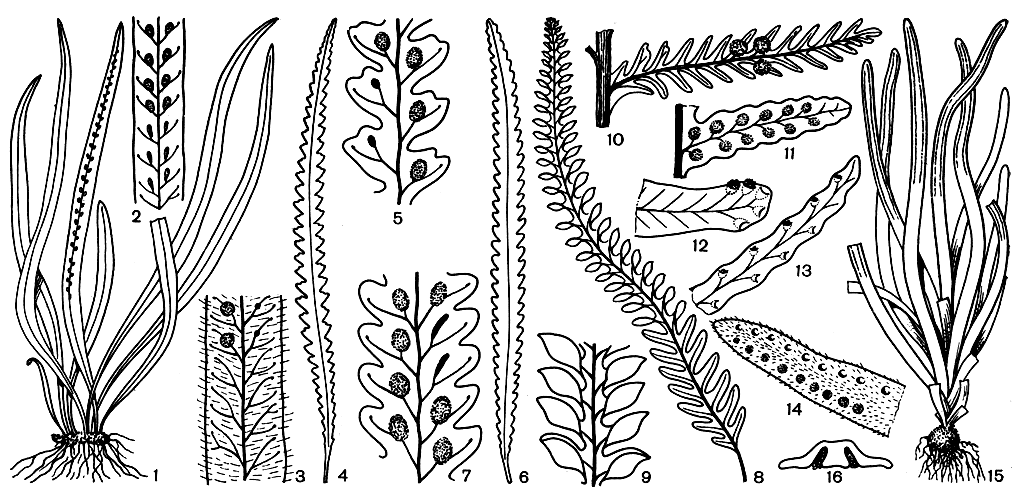
Рис. 124. Общий вид и расположение сорусов на листьях папоротников семейства граммитисовых: 1 - граммитис усыпанный (Grammitis adspersa) - общий вид; 2 - граммитис усыпанный - часть листа с сорусами (увел.); 3 - граммитис Холтума (G. holttumii) - часть листа с сорусами (увел.); 4 - ксифоптерис Иеронима (Xiphopteris hieronimusn) - лист (умен.); 5 - ксифоптерис Иеронима - часть листа с сорусами (увел.); 6 - ксифоптерис рассеянноволосистый (X. sparsipilosa) - лист (умен.); 7 - ксифоптерис рассеянноволосистый - часть листа с сорусами (увел.); 8 - калиммодон клобучковый (Calymmodon cucullatus) - лист; 9 - калиммодон клобучковый - часть спороносного листа (увел.); 10 - ктеноптерис тонкорассеченный (Ctenopteris tenuisecta) - одно перо (увел.); 11 - ктеноптерис небольшой (С. subminuta) - перо (увел.); 12 - ктеноптерис крылатый (С. alata) - одно перо (увел.); 13 - ктеноптерис Лейза (С. leysii) - одно перо (увел.); 14 - ктеноптерис кхасийский (С. khasyana) - одно перо (увел.); 15 - склероглоссум крохотный (Scleroglossum pusillum) - общий вид (увел.); 16 - склероглоссум крохотный - поперечный срез спороносного листа
Граммитис характеризуется простыми линейными или ланцетными листьями с цельной кожистой или мясистой пластинкой. Черешки и поверхность листа часто покрыты короткими или длинными, жесткими волосками красного цвета. Жилки обычно однажды вильчатые, у видов с мясистыми листьями они почти незаметные.
Сорусы граммитисовых располагаются либо на поверхности жилок, либо на их концах - по одному на каждой верхней жилке. Сорусы округлые или удлиненные, при созревании могут сливаться, образуя две сплошные полосы, параллельные срединной жилке. У граммитиса всеобщего (Grammitis universa), ширина листа которого составляет всего 2-3 мм при длине 2,5-5 см, зрелые, слившиеся сорусы занимают все пространство между срединной жилкой и краем листа.
У ксифоптериса в каждом сегменте перисторассеченных листьев имеется одна простая или вильчато разветвленная жилка, верхняя ветвь которой несет продолговатый сорус. В системе семейства этот род занимает промежуточное положение между граммитисом и ктеноптерисом. Для последнего характерны перистые листья с перистоветвящейся жилкой и одним или несколькими сорусами в каждом сегменте. Сорусы ктеноптериса иногда поверхностные, как у ктеноптериса разнолистного (Ctenopteris heterophylla), но обычно либо слегка погружены в ткань листа, либо почти утоплены в ней. Из ктеноптериса обычно выделяют род просаптия (Prosaptia) с глубоко погруженными сорусами. Спорангии просаптии сидят как бы в кармашках, открытых на краю пластинки или недалеко от края. Внешне эти сорусы так напоминают сорусы рода даваллия, что к нему неоднократно ошибочно относили виды просаптии.
Сорусы всех граммитисовых лишены покрывальца, но могут быть частично защищены тканью листа (у видов с погруженными сорусами) или загнутым краем листа - у родов акросорус (Acrosorus) и калиммодон (Calymmodon). В отличие от полиподиевых, у которых ножка спорангия из двух рядов клеток, у граммитисовых она состоит, за исключением верхушки, из одного ряда клеток. Спорангии граммитисовых с ясно выраженным стомием, глад-кие или у некоторых видов со щетинками.
Высыпающиеся из спорангия споры прорастают сразу же, без периода покоя. Очень часто, как и у гименофилловых, первые деления могут происходить в споре, находящейся в нераскрывшемся спорангии. Молодые гаметофиты граммитисовых имеют нитевидную форму, и их нетрудно принять на этой стадии за гаметофиты гименофилловых. Однако с возрастом различия между ними проявляются все более и более отчетливо.
Молодые гаметофиты граммитисовых можно отличить по четковидным или бочонковидным клеткам нити, а впоследствии - по развивающейся лопатчатой пластинке, несущей на брюшной стороне в срединной части антеридии и архегонии. Антеридии и архегонии граммитисовых более подвинутого типа, чем у гименофилловых, но более примитивные, чем у полиподиевых. Органы полового размножения закладываются у граммитисовых обычно позднее, чем у других папоротников (исключая гимено- филловые), да и все развитие их гаметофита - исключительно медленный процесс. Нитчатая стадия может длиться от нескольких месяцев до года или даже двух лет, а созревание - еще дольше. У ктеноптериса Дженмана (Ctenopteris jenmanii), изученного в лабораторных условиях, архегонии появились в возрасте 4 года. Необычно время появления антеридиев. Они формируются после появления нескольких архегониев, а не наоборот, как у большинства папоротников. Долгоживущие гаметофиты граммитисовых способны к вегетативному размножению путем распадения нити на несколько кусков или путем почкования, подобно гаметофитам гименофилловых и виттариевых. В зрелых гаметофитах, найденных в естественных условиях, в родительских клетках ризоидов всегда присутствуют гифы грибов.
Мы уже отмечали некоторые общие черты в облике и развитии гаметофитов у таких далеких семейств, как граммитисовые и гименофилловые, что не связано с их родством и объясняется одинаковыми условиями существования.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'