Порядок троходендровые (Trochodendrales) (А. Л. Тахтаджян)
Семейство троходендровые (Trochodendraceae)
Единственный представитель этого семейства - троходендрон аралиевидный (Trochodendron aralioides, рис. 110) относится к числу наиболее замечательных "живых ископаемых" Восточно-Азиатской флористической области. Это растение давно привлекает внимание ботаников, и ему посвящен ряд важных исследований, начиная с работ А. Эйхлера (1864) и Ф. Ван Тигема (1900) и кончая работами И. Бейли и III. Наст (1945), Н. В. Первухиной и М. Д. Иоффе (1962).
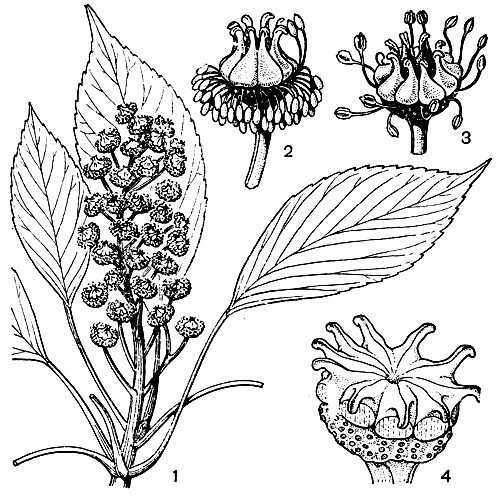
Рис. 110. Троходендрон аралневидный (Trochodendron aralioides): 1 - цветущая ветка; 2 и 3 - цветки на разных стадиях развития; 4 - плод
Троходендрон аралиевидный был впервые описан в 1835 г. во "Флоре Японии" Ф. Зибольда и Й. Цуккарини. В Японии он произрастает в горах на островах Хонсю, Сикоку и Кюсю, где приурочен к буковым или смешанным хвойно-широколиственным лесам. Позднее он был обнаружен также в южной части Корейского полуострова и на островах Рюкю и Тайвань. На Тайване он встречается в высокогорьях (на высоте от 2000 до 3000 м над уровнем моря) центральных хребтов, где иногда образует чистые насаждения, а также вдоль горячих серных источников на высоте 600 - 800 м над уровнем моря в северной части острова.
Троходендрон - вечнозеленое дерево высотой от 5 до 25 м, с очередными, простыми, перистонервными, кожистыми листьями, снабженными длинными черешками и лишенными прилистников (рис. 110). Почки крупные, с многими черепитчатыми чешуями. Устьица довольно своеобразного строения: по обе стороны от замыкающих клеток и параллельно их продольной оси находится от 2 до 4 побочных клеток, а к устьичным полюсам примыкают одна или две обычные клетки эпидермы. Но если устьичный аппарат троходендрона относительно специализирован, то его проводящая система чрезвычайно примитивна. Ксилема лишена сосудов и водопроводящие элементы представлены исключительно трахеидами. Кроме того, трахеиды гораздо более примитивные, чем у большинства современных голосеменных. Это очень длинные трахеиды, которые в ранней древесине имеют лестничные окаймленные поры, а в более узких трахеидах поздней древесины принимают более или менее округлые очертания. Все другие элементы проводящей системы также очень примитивные. В строении репродуктивных органов уже много признаков специализации.
Цветки небольшие, в коротких, верхушечных, прямостоячих, многолучевых, кистевидных верхоцветниках (кистевидных плейохазиях), обоеполые, лишенные околоцветника, с 2 довольно сильно редуцированными прицветничками, приросшими к цветоножке и цветоложу. Отсутствие околоцветника компенсируется многочисленными (обычно около 70) и расположенными по спирали тычинками с линейными нитями, которые своими основаниями приросли к нижней и отчасти средней части плодолистиков. Плодолистиков большей частью 5 - 11, чаще всего 7 - 9 (у верхушечных цветков иногда до 15 - 17), сросшихся между собой боковыми поверхностями, причем срастание проявляется в слиянии не только паренхимных тканей, но и боковых проводящих пучков соседних плодолистиков; столбики короткие, отогнутые наружу, с низбегающим двухгребневым рыльцем (примитивный признак); в результате срастания тычиночных нитей с нижней частью плодолистиков завязь троходендрона можно рассматривать как полунижнюю. Спинная часть плодолистиков выпуклая и играет роль нектарников. В каждом гнезде завязи закладывается по 25 - 30 висячих семязачатков, но значительная их часть остается стерильной. Форма семязачатков крайне своеобразна. Они образуют длинные субхалазальные выросты, удлиняющиеся в процессе формирования семени и сохраняющиеся до его созревания. По мнению Н. В. Первухиной и М. Д. Иоффе (1962), развитие этого субхалазального выроста, вероятно, связано с тем, что к моменту цветения полость завязи заполнена слизистым веществом, содержащим белки и танины. Авторы высказывают предположение о гаусториальной функции субхалазального выроста.
Отсутствие околоцветника в цветках троходендрона давало повод к предположению, что он опыляется ветром. В действительности троходендрон - растение насекомоопыляемое. Его крупные соцветия хорошо заметны в период цветения благодаря ярко-желтой пыльце на раскрытых пыльниках, а приятный запах цветков разносится довольно далеко, особенно утром в солнечную погоду. Цветки посещаются пчелами, бабочками и другими насекомыми. Кроме пыльцы, они находят в цветках троходендрона нектар, обильно выделяющийся на выпуклых спинных частях плодолистиков. Как показали наблюдения китайского ботаника X. Кенга (1959), в окрестностях города Тайбэй (остров Тайвань) около половины деревьев протандричны, остальные протогиничны. У протогиничных цветков как тычинки, так и гинецей хорошо развиты, и, следовательно, они функционально обоеполые. Что же касается протандричных цветков, то столбики у них первое время плотно прижаты друг к другу и расходятся лишь после раскрывания пыльников.
По наблюдению Кенга, плоды у протандричных деревьев не завязываются, но это нуждается в подтверждении наблюдениями в других частях ареала троходендрона.
Плод троходендрона представляет собой синкарпную многолистовку. Семена с маленьким зародышем и обильным эндоспермом. Они снабжены хорошо развитым субхалазальным придатком, который способствует распространению их ветром.
Троходендрон высоко ценится как декоративное растение.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'