Корень (А. Л. Тахтаджян)
О строении и функциях корня и о различных его модификациях мы знаем гораздо меньше, чем о стебле и листе. Одной из причин этого являются определенные технические трудности, связанные с изучением подземных органов вообще. Однако за последние десятилетия корни, их строение и функции, вызывают большой интерес, и теперь мы знаем о них гораздо больше, чем было известно ботаникам в прошлом столетии или даже в первой половине нашего века.
Зачаток корня, или корешок (зародышевый корень), имеется уже в семени. При прорастании семени корешок превращается в главный, или первичный, корень. По бокам главного корня развиваются боковые корни, зачатки которых появляются ближе к его верхушке. Большинство двудольных характеризуется преобладающим ростом главного корня (он развивается сильнее, чем боковые корни) и корневая система называется поэтому стержневой. У однодольных растений, а также у многих травянистых двудольных (например, у лютика и подорожника) главный корень рано отмирает или развивается слабо и корневая система образуется из так называемых придаточных корней, формирующихся у основания воздушного стебля или на видоизмененных подземных стеблях. Эти более или менее одинаково развитые придаточные корни и их боковые корни образуют так называемую мочковатую корневую систему. У многих растений придаточные корни могут образовываться также на воздушных стеблях (воздушные корни), а иногда даже на листьях.
Корни исполняют иные функции, чем стебли, и поэтому неудивительно, что и строение их во многом совершенно иное. Корню не нужна жесткая структура стебля, но в то же время он должен быть достаточно прочным и гибким^ чтобы надежно удерживать растение в почве, особенно при сильном ветре. Поэтому ксилема вместе с механической тканью сосредоточены в центре корня, где они придают ему максимальную прочность на разрыв, а также обеспечивают предельную гибкость.
В отличие от воздушных стеблей корням приходится прокладывать путь в твердой, нередко в очень твердой почве. Поэтому нежная точка роста корня, ее апикальная меристема, надежно защищена от повреждений при трении о твердые частицы почвы специальным образованием, называемым корневым чехликом. Корневой чехлик небольшой, длиной обычно меньше 1 мм. Он состоит из рыхло расположенных тонкостенных клеток. При прохождении корня через почву эти клетки постепенно стираются и заменяются новыми, так что кончик корня все время остается защищенным. Однако было бы неправильно думать, что функция корневого чехлика ограничивается только защитой точки роста. Читатель, конечно, знает, что для корня характерен положительный геотропизм. Как показали специальные исследования, у корней зона, воспринимающая геотропическое раздражение (т. е. влияние силы тяжести), находится именно в корневом чехлике. В этом очень легко убедиться: растения, у которых корневой чехлик нетрудно удалить (например, злаки), геотропическую реакцию утрачивают, но после регенерации чехлика она у них вновь восстанавливается. Положительный геотропизм корня - явление довольно сложное, и многое в нем нам еще неизвестно. Но уже давно было высказано предположение, что восприятие геотропического раздражения связано с перемещением в клетках корневого чехлика особых тяжелых частиц, чаще всего крупных крахмальных зерен. В зависимости от ориентации корня эти частицы оказывают давление на различные участки эктоплазмы, вызывая там соответствующее возбуждение. Однако характер этого возбуждения пока неясен.
Корень не только исполняет чисто механическую, якорную функцию, но и служит также для активного всасывания воды, ионов минеральных солей, некоторых продуктов жизнедеятельности почвенных микроорганизмов и корневых выделений других растений. Ионы калия, кальция, магния и фосфора и органические молекулы соединений азота и серы в результате действия корневого давления и транспирации передвигаются по ксилеме в стебли и листья. Огромную роль в процессе всасывания играют крошечные корневыеволоски, расположенные вблизи кончика корня. Каждый корневой волосок представляет собой вырост поверхностной клетки корня. Полость корневого волоска является продолжением полости клетки, причем ядро клетки переходит в корневой волосок. Благодаря наличию корневых волосков всасывающая поверхность корня увеличивается примерно в 18 раз. Наружные слои очень нежной и тонкой стенки корневого волоска слизистые, они сливаются с почвенными коллоидами, что в сильной степени способствует всасывающей деятельности.
Корни выполняют и другие функции. В корнях синтезируются гормоны роста, алкалоиды и другие физиологически активные вещества.
Архитектура корневой системы, степень ее развития и глубина проникновения в почву очень различны у представителей разных систематических групп и разных жизненных форм. Она в сильной степени зависит также от климатических и почвенных условий, в первую очередь от распределения воды в почвенных горизонтах. Наиболее слабо развита корневая система у однолетних растений. Но она расположена очень неглубоко также у ряда многолетних растений, в частности у большинства кактусов, злаков и других растений. В то же время у многих ксерофитов корни проникают на очень большую глубину, особенно там, где вода залегает далеко от поверхности. У верблюжьей колючки (виды рода Alhagi, семейство бобовых) корни столь длинные, что достигают грунтовых вод. В растительных сообществах у растений разных ярусов корневые системы располагаются на разной глубине, что дает возможность более полного использования почвы и более мирного сосуществования разных жизненных форм.
Одним из интереснейших явлений в биологии корня является более или менее взаимовыгодное сожительство с грибами. Оно носит специальное название микоризы (что буквально означает "грибокорень"), и ему посвящена большая литература. За исключением водных и паразитных растений, ряда однолетних, гречишных, крестоцветных, осоковых и немногих, других микориза характерна для подавляющего большинства цветковых растений (вероятно, не менее 90%). Многие растения, такие, как орхидные, а особенно растения, полностью перешедшие к сапрофитному образу жизни, настолько тесно связаны с грибами, что не могут развиваться без заражения микоризным грибом. Другие растения, например многие лесные деревья и кустарники, могут расти и без микоризы, но при заражении грибом развиваются значительно лучше. Наконец, ряд растений, таких, как береза, липа, многие кустарники, нередко имеющие микоризу, в благоприятных условиях питания хорошо развиваются и без заражения грибом.
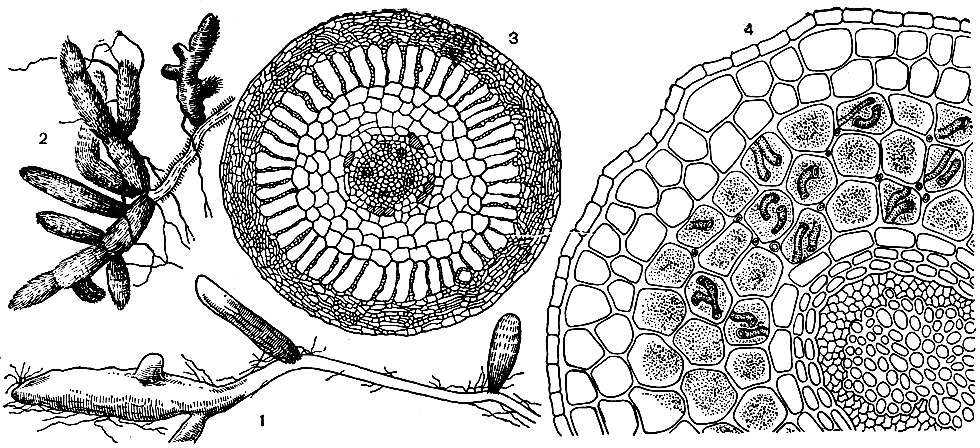
Рис. 6. Типы микориз. Эктомикоризы: 1,2 - у дуба (Quercus sp.); 3 - поперечный разрез эктомикоризы бука (Fagus sp.). Эндомикориза: 4 - поперечный разрез эндомикоризы ясеня ланцетного (Fraxinus lanceolata)
Микориза бывает двух разных типов (рис. 6). К первому типу относится эктомикориза (наружная микориза). Плотные сплетения гиф гриба оплетают корни растений толстым и плотным чехлом (гифовой мантией) и проникают, кроме того, в межклетники (но не в клетки) одного или нескольких наружных слоев коры. Под влиянием гормонов, выделяемых грибом, молодые корни обычно ветвятся и окончания их утолщаются. Корневые волоски при этом отмирают. Эктомикориза характерна для многих деревьев умеренной зоны, в том числе для дуба, березы и ивы, а также для некоторых кустарников и трав. Хотя гриб и оттягивает из корня углеводы, аминокислоты, а также, вероятно, другие органические вещества, в то же время он снабжает его неорганическими питательными веществами, которые ему легче поглощать из почвы, чем лишенному корневых волосков корню. Кроме того, будучи сапрофагом, гриб способен расщеплять некоторые недоступные растению органические соединения почвы. Очень важно также, что между грибом и корнем происходит обмен аминокислотами и физиологически активными веществами. Эктомикоризу образуют почти исключительно базидиомицеты (чаще гименомицеты, реже гастеромицеты), а также некоторые аскомицеты. Нередко в образовании микоризы участвует несколько видов грибов.
Гораздо более широкое, почти универсальное распространение имеет эндомикориза (внутренняя микориза). Ее можно наблюдать, например, у яблони, груши, земляники, томата, злаков, орхидных и многих других видов. Она характерна для большинства цветковых растений. При эндомикоризе грибной чехол вокруг корня не образуется, корневые волоски не отмирают, но гифы проникают гораздо глубже в ткани корня и внедряются в клетки коровой паренхимы. Существует несколько различных типов эндомикоризы, причем некоторые из них сильно отличаются друг от друга. У представителей большого семейства вересковых, например у вереска, арктоуса, грушанки, черники, клюквы и др., образуется особого типа эндомикориза, занимающая в некоторых отношениях промежуточное положение между типичной эктомикоризой лесных деревьев и высокоспециализированной эндомикоризой орхидных. Гифы проникают у них в клетки коры, образуя в них плотную массу, но в то же время оплетают корни рыхлым чехлом. При этом клубки гиф в клетках коры впоследствии перевариваются растением-хозяином, что вообще характерно для эндомикоризы. С другой стороны, микоризный гриб при соответствующих условиях может, в свою очередь, стать настоящим паразитом. В образовании микоризы типа вересковых участвуют оомицеты и зигомицеты. Наиболее специализированный тип эндомикоризы можно наблюдать у орхидных. В отличие от эктомикоризы и микоризы вересковых у орхидных нет гифовой мантии вокруг корней и мицелий почти целиком находится внутри корня. Гифы гриба проникают из почвы в клетки коры корня, где образуют клубни, которые впоследствии перевариваются клетками растения-хозяина. В отличие от эктомикоризных грибов грибы, образующие микоризу орхидных, способны разлагать сложные органические вещества и снабжать корни продуктами их разложения, что имеет большое значение в сапрофитном питании орхидных. Микоризы типа орхидных имеются и у многих других растений, в частности у триурисовых и бурманниевых. Грибной компонент эндомикоризы почти всегда представлен оомицетами.
Очень велика роль микоризы в тропических дождевых лесах, где поглощение азота и других неорганических веществ происходит с участием микоризного гриба, который питается сапротрофно на опавших листьях, стеблях, плодах, семенах и пр. Основным источником минеральных веществ является здесь не сама почва, а почвенные грибы. Минеральные вещества поступают в корень непосредственно из гиф микоризных грибов. Таким путем обеспечивается более полное использование минеральных веществ и более полный их круговорот. Именно этим объясняется, что большая часть корневой системы растений дождевых лесов находится в поверхностнохм слое почвы на глубине около 0,3 м.

Рис. 7. Пневматофоры у соннератии (Soimeratia sp.) на острове Ява
Другим в биологическом отношении чрезвычайно важным, хотя гораздо менее распространенным явлением, чем микориза, служит сожительство корней с бактериями, о чем уже было подробно рассказано в 1-ом томе "Жизни растений" (глава "Азотфиксирующие бактерии"). Корневые клубеньки особенно характерны для корней большинства бобовых, но они известны также для других семейств (березовые, казуариновые, мириковые, лоховые, кориариевые, крушиновые, некоторые парнолистниковые, злаки и др.). Сожительство бактерий и корней имеет огромное практическое значение, а селекция и "генная инженерия" открывают в этой области почти сказочные перспективы.
Наше знакомство с жизнью и строением корня было бы неполным, если бы мы не остановились на различных его структурных и функциональных видоизменениях.
Обычно в корнях откладывается то или иное количество запасных питательных веществ, преимущественно углеводы, особенно крахмал и сахара. У многих растений их утолщенные и мясистые корни (часто вместе с утолщенным гипокотилем) специально приспособлены для отложения питательных веществ. Всем известными примерами являются свекла, редька, брюква, турнепс, морковь, петрушка, сельдерей, пастернак, женьшень, цикорий. У многих из них, например у свеклы и моркови, мясистая запасающая структура имеет как бы двойную морфологическую природу: верхняя ее часть состоит из гипокотиля (подсемядольного колена), а нижняя - из собственно корня, но для установления границы между ними требуются специальные анатомические исследования. Утолщение очень часто встречается также у растений с мочковатой корневой системой, например у всем известной георгины, или далии (Dahlia), относящейся к семейству сложноцветных.
Во многих случаях корни служат для вегетативного размножения. У целого ряда многолетних растений, как травянистых, так и древесных, из придаточных почек, образующихся на корнях (главных и боковых), развиваются надземные побеги, так называемые корневые отпрыски. Такие отпрыски (корневую поросль) можно наблюдать у барбариса, осины, вишни, сливы, сирени, вьюнка, бодяка, осота и многих других растений. У ряда злостных сорняков разрезанные при обработке почвы небольшие кусочки корней дают начало новым растениям, что сильно затрудняет борьбу с ними.
У некоторых лазящих растений, например у плюща, придаточные корни, образующиеся на стороне стебля, обращенной к дереву, скале или стене, проникают в трещины или в щели и крепко и надежно удерживают растение. Эти придаточные корниприцепки характеризуются отрицательным фототропизмом, что довольно необычно для корней.
Совершенно иную функцию несут так называемые контрактильные (сокращающиеся) или втягивающиеся корни,, характерные для многих корневищных, луковичных и клубне луковичных растений. Хорошим примером растения с контрактильными корнями может служить хотя бы крокус. У крокуса, как и у многих других геофитов, кроме обычных корней развиваются более длинные контрактильные корни, которые при сокращении втягивают клубнелуковицу, погружая ее глубже в землю. При этом контрактильные корни становятся поперечно морщинистыми, благодаря чему их легко отличить от обычных корней.

Рис. 8. Ходульные корни у пандануса (Pandanus sp.) в Богорском ботаническом саду на острове Ява
Корни могут служить также для запасания воды, что особенно хорошо видно на примере некоторых тропических эпифитных орхидных. Наружная часть коры свисающих вниз придаточных воздушных корней этих растений состоит из крупных и пустых клеток, которые могут впитывать воду подобно губке. Во время дождя эти клетки наполняются водой, которая в них и хранится и по мере необходимости используется растением.
У некоторых паразитных растений, например у представителей семейства гидноровых, корни изменили функцию и превратились в присоски (гаустории), внедряющиеся в проводящие ткани растения-хозяина и оттягивающие оттуда питательные вещества.

Рис. 9. Досковидные корни у канариума огромного (Canarmm decumanum) в Богорском ботаническом саду на острове Ява
У ряда тропических древесных растений, живущих на бедных кислородом почвах, главным образом у мангровых деревьев (например, у авиценнии), а также у деревьев, произрастающих в лесах на пресноводных тропических болотах, имеются специальные вентиляционные, или дыхательные, корни, называемые пневматофорами (рис. 7). Они развиваются из подземных боковых корней и растут вертикально вверх, поднимаясь над водой или почвой. Для пневматофоров характерен, таким образом, отрицательный геотропизм. У некоторых видов пневматофоры имеют форму "колен" или петлевидных выростов, образованных небольшими отрезками поверхностных боковых корней. Их значение заключается прежде всего в снабжении подземных частей воздухом, чему способствуют постоянно слущивающаяся тонкая кора, многочисленные чечевички и сильно развитая система воздухоносных межклетников. Но, кроме того, близ поверхности почвы на дыхательных корнях образуются тонкие корешки, которые составляют главную всасывающую силу корневой системы мангровых растений. При каждом последовательном повышении уровня почвы на дыхательных корнях образуются новые группы корешков.
У ряда мангровых растений (у ризофоры и некоторых других) на стволах на высоте прилива развиваются воздушные корни, которые растут вниз и, укрепившись в почве, прочно удерживают растение в мягком иле. Это ходульные корни (рис. 8). Они нередко встречаются также у некоторых обычно небольших или среднего размера деревьев тропического дождевого леса, особенно часто у деревьев, произрастающих на болотах. Они имеются также у некоторых пальм, панданусов, некоторых трав тропического дождевого леса и даже у кукурузы. Но наиболее эффектны ходульные корни некоторых видов фикуса, особенно знаменитого баньяна (Ficus benghalensis). Многочисленные придаточные корни баньяна растут вниз, укореняются и развивают собственную корневую систему. Благодаря этому одно дерево баньяна разрастается в целую рощу, которая может занимать площадь в несколько сотен квадратных метров.
Не менее впечатляют опорные досковидные корни, наиболее характерные для крупных деревьев тропического дождевого леса, особенно для деревьев самого высокого яруса. В отличие от ходульных корней досковидные корни (рис. 9) представляют собой боковые корни. Проходящие у самой поверхности почвы или как раз над нею боковые корни развивают более или менее плоские треугольные и прилегающие к стволу вертикальные надземные выросты, напоминающие собой доски, прислоненные к дереву. Вначале эти досковидные корни округлые в сечении, но спустя некоторое время на их верхней стороне происходит сильный односторонний вторичный рост. В тропическом дождевом лесу высота досковидных корней нередко значительно превосходит человеческий рост. Иногда слабо выраженные досковидные корни встречаются и у некоторых деревьев умеренной зоны, например у бука, вяза и тополя. В тропическом лесу встречаются образования промежуточного типа между ходульными и досковидными корнями, а в некоторых случаях у одного и того же растения могут быть и ходульные и досковидные корни. Досковидные корни встречаются гораздо чаще, чем ходульные. Они имеют и более важное биологическое значение. Интересно, что у большинства деревьев, имеющих досковидные корни, нет стержневого корня и корневая система целиком состоит из поверхностных боковых корней с маленькими, растущими вниз ответвлениями. При этом наибольшая глубина проникновения корней в почву обычно не превышает 0,5 м. К сожалению, несмотря на существование ряда гипотез, в настоящее время нет еще достаточно удовлетворительной теории, объясняющей биологическое значение досковидных корней для деревьев тропического дождевого леса.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'