Порядок хетофоровые (Chaetophorales)
Хетофоровые по разнообразию представителей занимают одно из первых мест среди зеленых водорослей. Слоевище их, как и у улотриксовых, слагается из однорядных нитей, но нити эти ветвятся, и все слоевище дифференцировано на систему стелющихся и вертикальных разветвленных нитей.
Разнонитчатая структура составляет основное отличие хетофоровых от улотриксовых. Однако в пределах порядка стелющаяся и вертикальная структуры не всегда одинаково хорошо выражены, часто та или другая редуцируется или недоразвивается. Это создает большое морфологическое разнообразие. Многие хетофоровые обнаруживают черты высокой специализации в строении вегетативной и репродуктивной систем.
Подавляющее большинство водорослей порядка имеет на слоевище так называемые волоски или щетинки (благодаря этим образованиям они и получили свое название). Волоски у хетофоровых бывают по крайней мере двух типов. У одних они представляют конечные клетки веточек, сильно вытянутые и лишенные содержимого. У других - это выросты оболочки. Несмотря на то что волоски присущи почти всем хетофоровым, их, по-видимому, все же нельзя считать принципиальным признаком порядка. Интересно, что бурые и красные водоросли разнонитчатого строения также несут волоски.
Подавляющее большинство хетофоровых - пресноводные организмы, и только небольшое число вторично-упрощенных форм проникает в море.
Из пяти семейств порядка наибольший интерес представляют три: хетофоровых (Chaetophoraceae), трентеполиевых (Trentepohliaceae) и колеохетовых (Coleochaetaceae).
В наиболее типичной форме разнонитчатое строение проявляется у рода стигеоклониум (Stigeoclonium) в семействе хетофоровых. Большинство видов этого рода имеет хорошо развитые стелющуюся и вертикальную системы нитей (табл. 30, 1). Неправильно разветвленные вертикальные нити заметно сужаются у вершин и заканчиваются длинным волоском, состоящим из нескольких бесцветных клеток. Клетки слоевища имеют один пристенный хлоропласт, расположенный пояском, с одним или несколькими пиреноидами. Строение стелющейся части проследить труднее, так как оно во многом зависит от неровностей субстрата. Она может быть представлена рыхлоразветвленными нитями, состоящими нередко всего из нескольких клеток (рис. 220, 1), или густо разветвленными и плотно расположенными короткими нитями, складывающимися в псевдопаренхимный однослойный диск (рис. 220, 2). В какой-то мере степени развития вертикальной и стелющейся частей находятся в обратной зависимости. При сильном развитии стелющейся части вертикальная может редуцироваться до нескольких веточек или быть представленной только волосками. При незначительном развитии стелющихся нитей в основании растений развиваются ризоиды (рис. 220, 5). В отличие от прямостоячих нитей, для которых характерен диффузный рост, у стелющихся нитей рост апикальный и более короткие клетки. Обычно у стигеоклониума вертикальные ветви не от-личаются по толщине, но у некоторых видов можно выделить главную ось, клетки которой толще, длиннее и беднее содержимым, чем клетки веточек.
Такое строение можно рассматривать как переходное между стигеоклониумом и драпарналъдией (Draparnaldia) - другим родом семейства (табл. 30, 4). У драпарнальдии стелющаяся структура практически отсутствует, тогда как вертикальная развита значительно и достигает высокой специализации. Она дифференцируется на длинные и короткие ветви. Длинные состоят из крупных клеток с зубчатым хлоропластом, занимающим положение пояска по экватору клетки (рис. 220, 4). Короткие веточки густо ветвятся и располагаются мутовками вокруг главной оси. Слагающие их клетки содержат цельный хлоропласт, занимающий большую часть видимой поверхности клетки. Вершины этих веточек заканчиваются длинным волоском. Очевидно, короткие веточки несут основную ассимиляционную функцию, тогда как главные ветви играют прежде всего механическую, опорную роль. Прикрепляется драпарнальдия редуцированной стелющейся структурой и дополнительными многоклеточными ризоидами (рис. 220, 5).
У еще более специализированного рода драпарналъдиопсис (Draparnaldiopsis) ризоиды развиваются по всему слоевищу и настолько сильно, что образуют плотную обвертку вокруг главной оси (рис. 220, 6).
Описанные водоросли характеризуются также тем, что их слоевище покрыто тонким слоем слизи. В отличие от них у хетофоры (Chaetophora) все нити погружены в плотную слизистую обвертку, которая имеет шаровидную или листовидную форму (рис. 220, 7, 8). Базальная часть хетофоры состоит из отдельных шаровидных клеток, от которых радиально расходятся вер

Рис. 220. Строение хетофоровых: 1-3 - Stigeoclonium (1, 2 - строение стелющейся части слоевища, 3 - ризоиды); 4-5 - Draparnaldia (4 - клетка главной оси, 5 - ризоиды); 6 - часть слоевища Draparnaldiopsis; 7-9 - Chaetophora (7, 8 - внешний вид растений, 9 - часть слоевища) тикальные нити, густо разветвленные у вершины (рис. 220, 9)
К этому же семейству относится интересная водоросль фритчиелла (Fritschiella), единственный вид которой приспособился к существованию в почве (рис. 221). Под поверхностью почвы простираются стелющиеся нити, от которых вниз отходят ризоиды, а вверх - вертикальные нити. Последние, достигая поверхности почвы, густо ветвятся, образуя пучки ветвей.
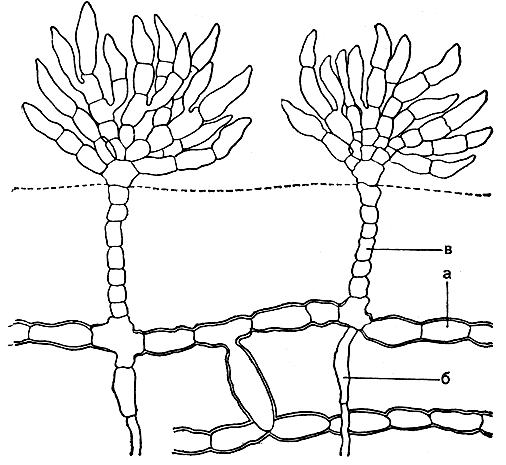
Рис. 221. Строение хетофоровых: Fritschiella tuberosa. а - стелющиеся нити; б - ризоиды; в - вертикальные нити
Тенденция к редукции вертикальной системы приводит к образованию большого числа стелющихся форм (табл. 30, 6). Все стелющиеся представители семейства, как правило, эпифиты или эндофиты, растущие на водных цветковых растениях и водорослях, некоторые из них поселяются на гидроидах и мшанках. Слоевище этих водорослей устроено по-разному. У одних форм на стелющихся нитях развиваются очень короткие вертикальные нити, оканчивающиеся волоском (рис. 222, 1); у других волоски образуются непосредственно на клетках стелющихся нитей, как, например, у водоросли афанохете (Aphanochaete, рис. 222, 2). У близкого рода болбоколеон (Bolbocoleon) волоски берут начало от специальных более мелких клеток (рис. 222, 3). Полная потеря вертикальных нитей и волосков наблюдается у эндодермы (Endoderma), растущей внутри оболочек морских водорослей и мшанок (рис. 222, 4). У морских хетофоровых стелющиеся нити часто соединяются в компактную дисковидную пластину. Она остается однослойной или становится многослойной в центре в результате того, что здесь образуются короткие вертикальные нити, плотно соединенные между собой. У водоросли прингсхеймиелла (Pringsheimiella, табл. 30, 6) вегетативные клетки однослойного диска уплощены и радиально вытянуты, тогда как зрелые спорангии в центре диска вытягиваются вверх и принимают клиновидную форму. Волоски у дисковидных форм обычно отсутствуют.
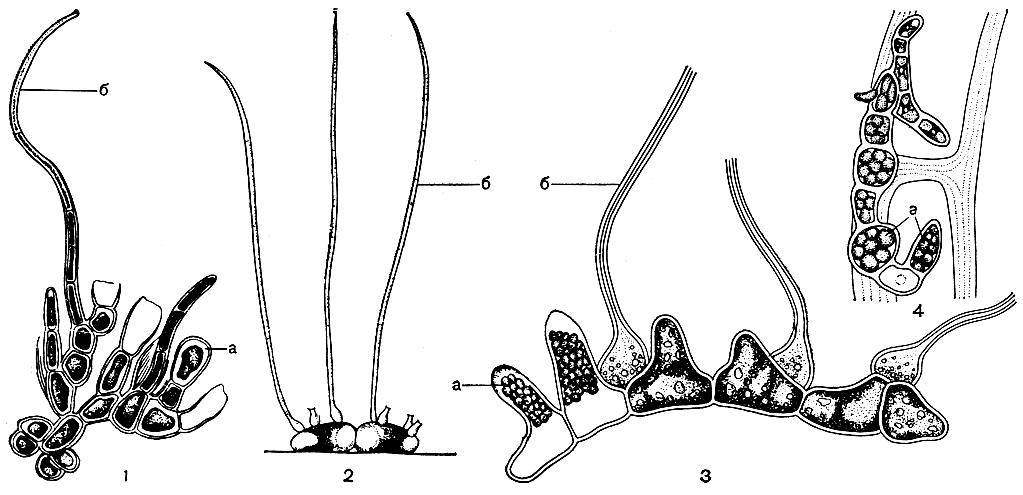
Рис. 222. Строение стелющихся хетофоровых: 1 - Pilinia maritima; 2 - Aphanochaete polychaete; 3 - Bolbocoleon piliferum; 4 - Endoderma wittrockii. a - спорангий, б - волосок
Размножение водорослей семейства хетофоровых во многом напоминает размножение улотрикса. Нередко, кроме четырехжгутиковых макрозооспор, служащих для бесполого размножения, можно встретить микрозооспоры. У большинства изученных хетофоровых половой процесс изогамный, а смена форм развития, особенно у морских форм, изоморфная. Только некоторым из них свойственна оогамия. Наличие оогамных форм среди стелющихся форм хетофоровых наводит на мысль о том, что в какой-то мере это искусственная группа.
Семейство трентеполиевых (Trentepohliaсеае) объединяет более специализированные по сравнению с хетофоровыми водоросли. Большинство родов семейства имеет достаточно хорошо развитую как вертикальную, так и стелющуюся часть и лишено волосков. Спорангии трентеполиевых - дифференцированные клетки, заметно отличающиеся по форме от вегетативных клеток.
Наиболее интересными особенностями обладает род трентеполия (Trentepohlia). Эта широко распространенная водоросль образует порошковатые скопления оранжевого или бурого цвета на камнях, пнях, стволах деревьев, деревянных постройках. Характерная окраска трентеполии обусловлена присутствием в ее клетках гематохрома - пигмента из группы каротиноидов.
Многие виды трентеполии являются водорослевым компонентом лишайников. Особенно обильно эта водоросль растет в условиях влаж-ного тропического климата.
Как правило, слоевище трентеполии имеет одинаково развитые вертикальную и стелющуюся части, однако нередко преимущественное развитие получает только одна из них. Все нити обильно ветвятся, причем место отхождения ветвей непостоянно, они возникают на вершине материнской клетки или в разных местах ее боковой стенки (рис. 223, 1). Рост нитей апикальный в результате деятельности верхушечной клетки. Цилиндрические или бочонковидные клетки одеты толстой целлюлозной оболочкой, которая нередко имеет слоистое строение. Поперечные перегородки между клетками всегда имеют посередине пору, через которую проходит цитоплазматический тяж. Особым строением обладают оболочки апикальных клеток. У ряда видов на вершине апикальной клетки образуется слоистый колпачок (рис. 223, 2-4). Клетки трентеполии, в отличие от хетофоровых, содержат несколько хлоропластов дисковидной или лентовидной формы, без пиреноидов (рис. 223, 5-8). В старых клетках всегда множество ядер.
Большую роль в жизни этой водоросли играет вегетативное размножение. Клетки нитей легко отделяются друг от друга и распыляются ветром. В виде отдельных богатых гематохромом клеток водоросль переживает неблагоприятный сухой период. Во влажное время года происходит бесполое и половое размножение трентеполии. Дву- и четырехжгутиковые зооспоры образуются в особых спорангиях на ножках (рис. 223, 9). Последние возникают на вертикальных нитях как один или несколько трубчатых выростов верхушечной клетки, отделяющихся от нее поперечной перегородкой. Эти выросты имеют характерную коленчатоизогнутую форму (рис. 223, 10, 11). Верхняя часть выростов раздувается и, превращаясь в спорангии, отделяется перегородкой от клетки-ножки.
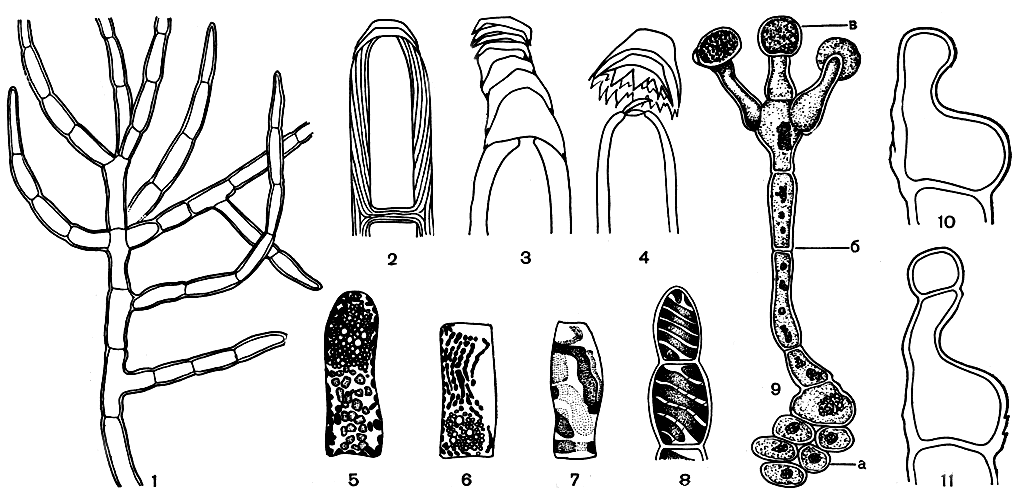
Рис. 223. Строение трентеполии (Trentepohlia): 1 - часть вертикального слоевища; 2-4 - образование колпачка на верхушечной клетке; 5-8 - клетки с хлоропластами; 9 - нить со спорангиями на ножках; 10,11 - образование спорангия (сбоку), а - стелющаяся часть, б - вертикальная нить, в - спорангий на ножке
Обычно зрелые спорангии отрываются от нити и разносятся ветром. Освобождение и прорастание зооспор происходит только при условии достаточного увлажнения.
Изогаметы развиваются в особых сидячих "спорангиях". Это шаровидные образования, занимающие разное положение на нитях: они могут быть верхушечными, боковыми, интеркалярными. Развиваются они преимущественно на стелющихся нитях. Эти сидячие "спорангии" образуются в результате увеличения размеров клеток и, в отличие от спорангиев на ножке, не отделяются от нити при созревании гамет. Не всегда развивающиеся в них двужгутиковые клетки размножения ведут себя как гаметы, нередко они прорастают без слияния или дают апланоспоры.
Трентеполиевые - высокоспециализированная группа в порядке Chaetophorales. Это особенно бросается в глаза при ее сравнении с семейством Chaetophoraceae. Очевидно, во многом специализация трентеполиевых - результат специфических наземных условий обитания.
Семейство колеохетовых (Coleochaetaceae) представлено единственным родом колеохете (Соleochaete). 14 видов этого рода растут в пресных водах как эпифиты, образуя подушковидные или дисковидные слоевища (табл. 30, 3).
Слоевища колеохетовых имеют в разной мере выраженную разнонитчатую структуру. У представителей с развитыми вертикальными нитями они обычно собраны в плотную полусферическую подушку, заключенную в слизь. У многих видов, однако, развивается только базальная часть, которая часто образована плотно расположенными нитями и имеет вид компактного округлого диска. Рост слоевища всегда апикальный. Одноядерные клетки заполнены крупным пристенным хлоропластом неправильной формы с одним или двумя четкими пиреноидами. Обычно клетки несут характерные заключенные в футляр щетинки (рис. 224, 3).
Бесполое размножение колеохетовых происходит весной и ранним летом двужгутиковыми крупными зооспорами. Они образуются по одной в обычных клетках и выходят через округлое отверстие на вершине короткого бородавчатого выроста. Зооспоры отличаются от-сутствием стигмы и тем, что единственный хлоропласт занимает в них боковое положение. При прорастании зооспоры образуются две лежащие друг над другом клетки, из которых верхняя формирует щетинку, а нижняя делится с образованием клеток диска (рис. 224, 1-3).
Половое размножение колеохете представляет собой высокоспециализированную оогамию. У колеохете подушковидного (Coleochaete pulvinata) оогонии образуются в верхней части вертикальных нитей. Они имеют характерную бутылковидную форму: нижняя, расширенная часть содержит хлоропласт, а верхняя, бесцветная вытянута наподобие трихогины красных водорослей. При созревании вершина оогония разрушается, в результате чего теряется некоторое количество цитоплазмы. В то же самое время протопласт нижней части округляется, формируя единственную яйцеклетку (рис. 244, 4-6). Антеридии развиваются по нескольку вместе на вершине вертикальных нитей, часто тех же самых, которые несут оогонии (рис. 224, 6). Каждый антеридий образует единственный антерозоид шаровидной формы с двумя длинными жгутиками на переднем конце. После оплодотворения верхняя, бесцветная часть оогония отделяется перегородкой, тогда как базальная постепенно растет. Одновременно оогоний начинает обрастать разветвленными нитями, возникающими из соседних клеток, которые, смыкаясь, в конце концов образуют плотную обвертку красного или бурого цвета (рис. 224, 7, 8). Эта обвертка позднее разрушается, и ооспора переживает период покоя в своей собственной толстой оболочке, которая у нее постепенно развивается. Весной содержимое ооспоры вновь приобретает зеленую окраску и делится с образованием 16-32 клиновидных клеток. Эти клетки, превращаясь в зооспоры, выходят наружу в результате разрыва оболочки ооспоры (рис. 224, 9).
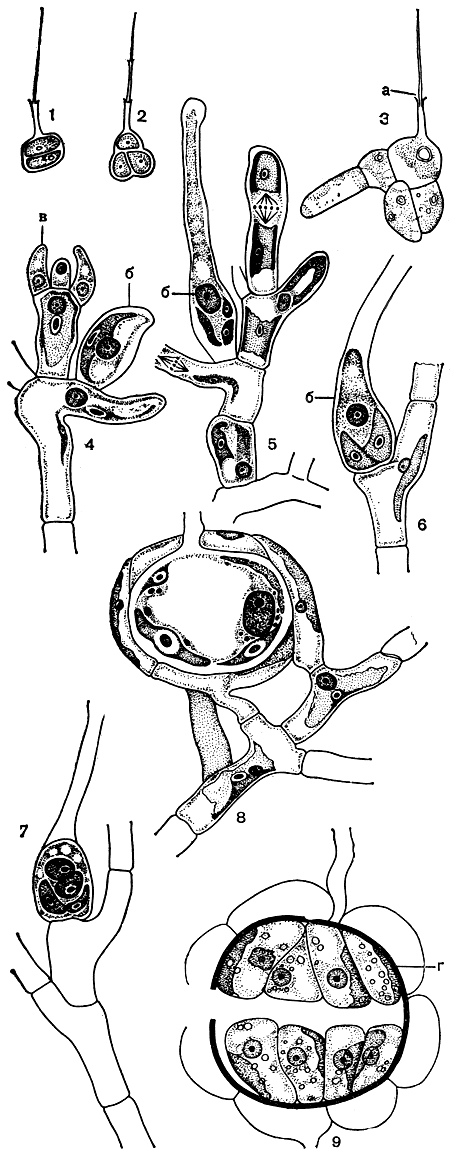
Рис. 224. Строение колеохете (Coleochaete): 1-3 - ранние стадии прорастания зооспоры; 4, 5, 6 - развитие оогония и антеридия (4); 7 - оогоний после оплодотворения; 8 - образование обвертки вокруг оплодотворенного оогония; 9 - прорастающая ооспора. а - щетинка, б - оогоний, в - антеридий, г - оболочка ооспоры
Подобно многим оогамным зеленым водорослям, колеохете занимает обособленное положение в порядке, и связи его с другими хетофоровыми, так же как и со всеми зелеными водорослями, не совсем ясны.
В настоящее время система всех нитчатых зеленых водорослей строится главным образом на морфологическом критерии. Однако нетрудно заметить, что как раз в морфологии водорослей более чем где бы то ни было проявляется параллельное и конвергентное развитие, т. е. многие заведомо далекие группы водорослей проходят одни и те же этапы морфологической эволюции. Этот факт показывает, что для построения системы водорослей, которая отражала бы эволюцию организмов и их родственные связи, одного морфологического критерия недостаточно. Требуется всестороннее изучение водорослей, в том числе их размножения и развития.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'