Движения устьиц
Выше мы уже неоднократно упоминали о значении воды для растений. Жизнь развилась в водной среде, да и существование современных растений нельзя представить себе без присутствия воды. Чтобы убедиться в ее влиянии на жизнь растений, достаточно лишь сравнить флористическое богатство тропического дождевого леса со скудным растительным покровом пустыни. Поэтому не удивительно, что у растений выработались приспособления, позволяющие им беречь воду. Прежде всего этой цели служит развивающийся в виде покровной ткани самый наружный слой клеток (эпидермис) стеблей и листьев. Но кроме защиты глубже лежащих клеток, он должен обеспечить и газообмен с окружающей средой. Эти обе функции эпидермиса осуществляются благодаря наличию в нем способных регулировать газообмен устьиц, которые поэтому могут рассматриваться как система обратной связи. Их задача состоит в том, чтобы регулировать как поступление СО2, так и отдачу H2O. Две ограничивающие устьичную щель клетки называют замыкающими клетками, а вместе с примыкающими к ним так называемыми побочными клетками они образуют устьичный аппарат. Открывания и закрывания устьиц - это типичные настические движения, направление которых зависит от анатомического строения. Но в отличие от рассмотренных нами до сих пор изменений положения органов закрепленных в субстрате растений ответные реакции устьиц можно наблюдать только с помощью микроскопа.
Хотя площадь, занимаемая отверстиями (устьичными щелями), составляет лишь от 1 до 2% общей площади поверхности листа, количество выходящей через них в атмосферу воды может достигать 50-70% того, какое испарилось бы с открытой водной поверхности, равной по площади поверхности листа. Это объясняется высокой активностью, испарения с краевых участков малых отверстий. Она основана на том, что при открытых устьичных щелях молекулы могут выходить как под прямым углом к поверхности листа, так и в стороны, тогда как с открытой поверхности воды они поднимаются только вверх. Поскольку зеленые растения из CO2 воздуха и H2O, имеющейся в клетках, создают органические вещества, устьичные щели должны быть открытыми. Но при этом неизбежна значительная потеря воды. Механизм открывания и закрывания устьичных щелей, отвечающий рациональному поглощению CO2 и выходу H2O в тех или иных условиях, т. е. оптимально и активно регулируемый, очень сложен. Кроме имеющих преимущественное значение фото- и гидронастических движений, определенную роль играют также термо- и хемонастические ответные реакции, а равно и наличие некоторых автономных компонентов. Как мы увидим в дальнейшем, эта способность реагировать на многосторонние влияния оказывается экологически целесообразной.
В зависимости от строения и направления движения клеток при открывании щели различают многие типы устьиц. Здесь мы ограничимся рассмотрением широко распространенного у однодольных и двудольных растений Helleborus-типа (русское название рода Helleborus - морозник, или зимовник). Каждая из замыкающих клеток имеет форму семени фасоли. У такой клетки верхние и нижние участки брюшной (обращенной к щели) стенки клеточной оболочки утолщены, а спинная стенка остается тонкой. Между замыкающими и побочными клетками нередко образуются сочленения. Открывание устьичной щели, будучи обусловленным этими особенностями строения клеток, происходит как параллельно, так и перпендикулярно поверхности (рис. 37).
Что же касается механики движений, то в основе открывания и закрывания устьичных щелей лежат изменения тургорного давления. При высоком тургорном давлении устьичная щель открывается, а при низком замыкающие клетки закрывают ее. Для осуществления обоих этих процессов нужна АТФ.
Для этих ответных реакций особенно характерны изменения осмотического потенциала замыкающих клеток. Если он увеличивается, то это ведет к возникновению различия в тургоре этих клеток и прилегающих к ним клеток эпидермиса: оно, в свою очередь, приводит к открыванию щели. Напротив, закрыванию устьичной щели предшествует снижение осмотического потенциала. Вызывающие движение факторы действуют на замыкающие и побочные клетки по-разному. Эти клетки не только соседи, но они также тесно связаны и функционально.
Поскольку открывание устьиц наиболее сильно подвержено влиянию света, сперва мы рассмотрим их фотонастические движения. Уже слабое освещение может вызвать открывание устьиц. Особенно сильно влияет на процесс их открывания синий и красный свет, что указывает на причастность к этому процессу и процесса фотосинтеза. Решающее значение для обусловленных освещением фотонастических движений устьичных клеток имеет содержание CO2 в листе. Его уровень зависит от процессов фотосинтеза, происходящих в основном в других клетках. Значение концентрации CO2 можно показать, например, на растениях, которые способны связывать CO2 в темноте, то есть ночью, и сохранять его для обеспечения процессов фотосинтеза, происходящих днем. При этом снижение содержания CO2 в клетках приводит к открыванию устьиц и в темноте. А при высокой концентрации CO2 можно наблюдать закрывание устьиц и при освещении.
Согласно классическому представлению, существовавшему до последних лет, снижение содержания CO2 вызывает возрастание значения рН. В результате этого при рН от 6,4 до 7,3 сильно зависящий от рН фермент фосфорилаза должен активизироваться и разложить осмотически неактивный крахмал на сахара. Хлоропласты, в которых осуществляется фотосинтез, находятся в замыкающих клетках устьиц, тогда как в прочих клетках эпидермиса их обычно нет. Но, как нам теперь известно, в повышении осмотического потенциала клеток участвует не сахар, а прежде всего калий. Он извлекается из побочных клеток, служащих хранилищами калия. Как хорошо вbдно на рис. 37, побочные клетки значительно крупнее замыкающих клеток. По электрохимическим причинам в замыкающие клетки наряду с калием в эквивалентном количестве должны попадать и анионы. У некоторых растений, таких, как лук (Аllium сера) и других, содержание ионов К+ уравновешивается содержанием Сl-. Однако в большинстве случаев в качестве электрохимического эквивалента выступает яблочная кислота - известный и важный промежуточный продукт цикла лимонной кислоты. Возникшие при диссоциации яблочной кислоты протоны (Н+), вероятно, с помощью протонного насоса выводятся. А взамен при содействии специфического транспортирующего калий насоса в замыкающие клетки поступает К+. Следовательно, функция яблочной кислоты как бы двойственна, а именно обеспечение и анионами, и катионами. Препятствующим действию ионных насосов оказывается фитогормон абсцизовая кислота. О процессах, происходящих при замыкании устьичной щели, известно относительно мало. Вероятно, при этом упомянутые выше обратимые процессы протекают в обратном порядке. АТФ, необходимая для активного транспорта ионов, может образоваться, во-первых, при дыхании и, во-вторых, при фотосинтезе. Поэтому-то устьица способны открываться и на свету, и в темноте. При хорошем обеспечении растения водой его открытые устьица могут терять восприимчивость к колебаниям содержания СО. Но при недостатке воды она восстанавливается.
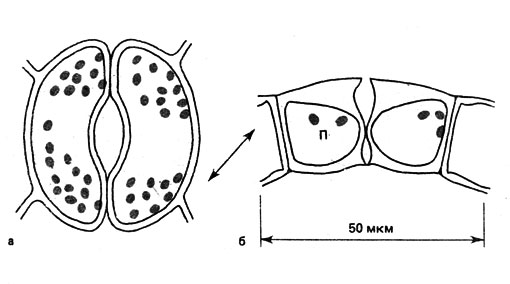
Рис. 37. Устьице (Helleborus =тип) y Helleborus niger: а - вид сверху, б - в поперечном сечении; стрелка указывает направление движения клеток при открывании устьичной щели; П - полость клетки, имеющей хлоропласта
На движение устьичных клеток, кроме света, сильно влияет и вода. Если исходить из того, что при засухе устьица, в общем-то, должны быть закрыты, их способность к гидронастическим движениям экологически необходима. Наверное, вы помните жаркое лето 1976 г., когда в течение почти шести недель не было ни одного дождя. И хотя потери урожая были значительными, все-таки большая часть растений осталась в основном неповрежденной. Несомненно, без регулирующих водообмен устьиц это не было бы возможным.
В большинстве случаев вода непосредственно влияет на гидронастические движения устьичных клеток. Если воды в растении много, то устьица открываются, тогда как при недостатке воды они закрываются. При этом концентрация осмотически активных веществ повышается или понижается. В управлении движениями устьичных клеток особое значение имеют фитогормоны. При недостатке воды усиливается образование абсцизовой кислоты, что можно выявить уже через несколько минут. Очевидно, этот гормон препятствует деятельности ионных насосов, в результате чего устьица закрываются. В противоположность абсцизовой кислоте цитокинин вызывает открывание устьиц.
Но и при тургорном давлении, остающемся в замыкающих клетках на одном уровне, устьица могут закрываться и открываться. Первое происходит, например, при дождевании растений, страдавших от нехватки воды, поскольку соседние устьицам клетки эпидермиса легче поглощают воду, чем сами замыкающие клетки. Прямо противоположное происходит у срезанных побегов, у которых возникающее вскоре после срезки нарушение обеспечения водой приводит к потере воды сначала побочными клетками, а в результате - к открыванию устьичных щелей (эффект Иванова). После того как относительное содержание влаги в замыкающих и побочных клетках придет в равновесие, эта ответная реакция вскоре уступит место движениям, закрывающим устьичные щели. Как показывает изложенное выше, замыкающие клетки устьиц реагируют на концентрацию СО2 и паров H2O в межклетниках листа. Более того, они функционируют и как специфические "осведомители" о влажности окружающего растение атмосферного воздуха. Например, у листьев с одинаковым содержанием H2O сопротивляемость транспирации в сухом воздухе гораздо выше, чем во влажном. Благодаря этому при низкой влажности воздуха растение может уменьшить транспирацию. Как и движения, обусловленные геотропизмом, движения устьичных клеток можно рассматривать как кибернетическую систему. Изменение тургорного давления контролируется взаимосвязанными; закономерностями, а замыкающие клетки оказываются постоянными компонентами. Особую роль при этом играют CO2 и H2O.
О хемонастических движениях устьичных клеток здесь можно подробнее не рассказывать. Ведь при рассмотрении фото- и гидронастических ответных реакций уже было упомянуто значение абсцизовой кислоты, а также и других веществ.
Термонастические движения замыкающих клеток нередко лишь с трудом можно отличить от фотонастических, поскольку зависимость состояния устьиц от температуры соответствует зависимости его от фотосинтеза. Однако у растений, обеспеченных водой, зависимость открывания устьиц от концентрации СО2 теряется. При относительно высоких температурах такая ответная реакция рациональна. Как раз при этих обстоятельствах важно испарять воду. Благодаря происходящему при транспирации охлаждению предотвращается перегрев листьев, и их температура остается в пределах, оптимальных для осуществления фотосинтеза.
Если открывание и закрывание устьиц вызываются внутренними факторами, то речь идет об автономных движениях. Хотя они, следовательно, и не относятся к настиям, мы о них все же коротко расскажем. Например, способность реагировать на действие внешних факторов сильно зависит от времени суток. Поэтому говорят о циркадных (имеющих примерно суточную периодичность) компонентах движений устьичных клеток. Это можно показать, если внешние условия остаются неизменными. Так, в частности, Musa acuminata - растение родом из тропиков Восточной Азии, относящееся к семейству банановых, - обнаруживает правильное чередование готовности минимально и максимально открывать устьичные щели. Указателем времени для поддержания этого ритма служит наступление темноты. После рассмотрения настий мы поговорим об автономных движениях подробнее.
Найти всю информацию о печати флаеров можно на сайте Типографии Альфа.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'