1.4. Адаптивные защитно-приспособительные реакции
Растительные организмы - это открытые системы, постоянно обменивающиеся веществом и энергией с окружающей средой. В клетке постоянно протекает множество разнообразных химических реакций, которые согласованы между собой по времени, скорости и месту протекания. Эта согласованность достигается благодаря разнообразным внутри- и внеклеточным системам регуляции. Неизменность среды организма (гомеостаз) поддерживается за счет притока в клетку веществ и энергии. Гомеостатические свойства целостного организма являются результатом одновременного действия многочисленных и сложноорганизованных регуляторных механизмов, обеспечивающих постоянство уровней веществ и энергии в организме. Генеральная линия любой клетки - сохранить свой внутриклеточный состав, несмотря на воздействия экстремальных факторов среды. Определенное состояние внутриклеточной среды обеспечивается синтезом белков и других соединений, внутриклеточным транспортом веществ, микрокомпартментальностью, градиентами значений pH, величинами мембранных, осмотических и водных потенциалов. Сохранение клеточного гомеостаза при постоянных изменениях условий окружающей среды определяется адаптивными возможностями данных клеток.
Сущность любой адаптации как процесса сводится к поддержанию структурной и функциональной устойчивости живой системы, обеспечивающей онтогенез и образование репродуктивных органов в изменяющихся условиях среды. Адаптация как фундаментальное свойство биологических объектов наряду с гомеостазом относится к числу центральных понятий биологии. Процессы биохимической адаптации выполняют в клетке следующие основные функции:
1. Поддержание структурной целостности макромолекул (белков, нуклеиновых кислот и др.) при их функционировании в специфических условиях.
2. Достаточное снабжение клетки: а) "энергетической валютой" - аденозинтрифосфатом (АТФ), б) восстановительными эквивалентами, необходимыми для протекания процессов биосинтеза, в) предшественниками, используемыми для синтеза нуклеиновых кислот и белков.
3. Поддержание систем, регулирующих скорости и направления метаболических процессов в соответствии с потребностями организма и их изменениями при изменении условий среды [52].
Эти фундаментальные функции необходимы всем живым системам.
Сущность адаптации - это подгонка структуры и свойств растительных организмов к условиям их произрастания. Основными направлениями стратегии адаптации организма к изменяющейся среде являются: 1) изменение количеств или типов макромолекул, имеющихся в клетке; 2) регулирование функциональной активности макромолекул; 3) такое изменение микросреды для макромолекул, которое защищало бы их от воздействия внешних неблагоприятных факторов. С помощью одного или комбинации этих механизмов организм достигает векторного гомеостаза метаболической функции, т. е. в процессе адаптации к внешней среде скорость и направление метаболических реакций "настраиваются" таким образом, чтобы организм непрерывно получал необходимые ему продукты и энергию.
Адаптация и гомеостаз - это взаимосвязанные и дополняющие друг друга процессы. Конечный результат многих стратегий адаптаций состоит в поддержании гомеостаза. Во многих случаях поддерживается не состояние определенных структур, а их функции. Вязкость (текучесть) биомембран изменяется при действии температур таким образом, чтобы могли нормально функционировать мембранно-связанные ферменты, транспортные системы, рецепторные молекулы и др. Результат адаптации в этих случаях не гомеостаз (постоянное состояние), а скорее энантиостаз (поддержание функции).
Временные параметры биохимической адаптации варьируют в широких пределах - от длительных периодов, необходимых для эволюционного изменения аминокислотных последовательностей, до долей секунды, за которые может измениться активность уже присутствующих в клетке ферментов. Чем больше времени предоставляется для адаптивных изменений, тем больше выбор возможных стратегий.
По длительности различают три типа адаптивных процессов:
1. Эволюционная (генетическая) адаптация. Это наиболее длительный процесс приспособления к среде, который зависит от приобретения новой генетической информации, детерминирующей новые адаптивные фенотипические признаки. Процесс эволюционной адаптации требует многих поколений. Медленные адаптационные изменения связаны с накоплением соответствующих генетических изменений: изменением плоидности, т. е. числа хромосом (геномные мутации); изменением структуры хромосом (хромосомные мутации); изменением в отдельных генах (генные мутации); мутациями во внеядерном генетическом материале (митохондрий, пластид). Мутации могут возникать спонтанно, и их можно вызвать воздействием экстремальных факторов. При эволюционной адаптации могут возникнуть совершенно новые молекулы, придающие организму способность к освоению новых местообитаний.
Кроме того, отдельные гены, группы генов или же отдельные нуклеотидные последовательности ДНК способны к движению, т. е. изменению своего места в геноме или же приобретению другого места в чужом геноме. Обмен генетическим материалом обнаружен внутри ДНК органелл, между ДНК разных органелл и между ДНК органелл и ядра. Такие типы обмена могут происходить только в эволюции или же составлять непрерывный процесс. Окружающая среда также изменяет геном растений. Вирусы, бактерии, взаимодействуя с растениями, приводят к изменению их наследственных свойств. В частности, заражение растений Agrobacteriurn tumefaciens, индуцирующее образование корончатых галлов, вызывает перенос бактериальных ДНК в геном растений.
Наряду с наследственными изменениями в ответ на действие окружающей среды могут происходить и ненаследственные изменения признаков (модификации). Более быстрые процессы модификации расширяют возможность вида успешно произрастать в разнообразных условиях местообитаний. К такому типу адаптации относятся:
2. Акклимация и акклиматизация. Во многих случаях адаптация осуществляется в течение жизни индивидума и требует для своего завершения от нескольких часов до нескольких месяцев. Примером служат многообразные сезонные изменения, наблюдаемые у организмов. Когда эти фенотипические сдвиги происходят в лабораторных условиях в ответ на экспериментальное варьирование какого-либо одного параметра среды (например, температуры или фотопериода), их называют акклимацией. Когда же фенотипические изменения наблюдаются в естественных условиях, адаптивные процессы называют акклиматизацией. В последнем случае могут одновременно изменяться несколько факторов среды и поэтому трудно установить, какой из них является сигналом к адаптации. Для приспособления организма к изменениям среды в этом случае может использоваться только та информация, которая уже содержалась в его геноме с самых первых дней жизни.
3. Немедленная адаптация. Адаптация происходит настолько быстро, что не может быть связана с изменениями в экспрессии генов или со значительной перестройкой клеточных структур в результате биохимических процессов. Она осуществляется путем изменения активности имеющихся ферментов. Быстрая "подгонка" активности ферментов представляет лишь первую линию защиты организма от неблагоприятных воздействий окружающей среды. Со временем на смену этой реакции приходят изменения в экспрессии генов или - в ряду поколений - изменения на генетическом уровне. Классическим примером немедленной адаптации могут служить таксисы - т. е. направленные перемещения к источнику действия (положительные таксисы) и от него (отрицательные таксисы). Например, отрицательный фототаксис хлоропластов к яркому свету и положительный - к слабому.
Таким образом, организм располагает способами биохимической адаптации разной степени сложности, которые позволяют ему успешно приспосабливаться к изменениям окружающей среды. Приспособление организмов к различным факторам среды вовлекает все уровни организации живого, начиная с молекулярного и кончая уровнем биоценоза. Многие растения избегают перегрева и избыточного освещения под пологом других видов растений. В этом случае приспособление осуществляется на уровне биоценоза. Охлаждение животных организмов с помощью секреции потовых желез, а растительных - путем транспирации является приспособлением на уровне организма. Переход хлоропластов от беспорядочного расположения к пристеночному при сильном освещении следует отнести к адаптации на клеточном уровне, а высокую термоустойчивость белков у термофильных организмов можно рассматривать как адаптацию на молекулярном уровне.
Благодаря всем существующим приспособительным реакциям биологическая эволюция породила виды животных, растений, микроорганизмов с совершенно различными требованиями к температуре, воде, элементам питания, свету и т. д. Приспособление растений к неблагоприятным условиям среды привело в ряде случаев к тому, что данные факторы стали для них необходимыми условиями существования. Примером служат некоторые галофиты. Солерос, представитель галофитов, развивается на засоленных почвах (до 0,1% по хлору) лучше, чем на пресном субстрате. С другой стороны, из работ Б. А. Келлера известно, что семена солероса лучше произрастают на пресном субстрате, чем на засоленных почвах. Это объясняется тем, что верхние слои солончаковых почв содержат крайне незначительное количество солей благодаря их вымыванию зимними осадками и прорастение семян солероса происходит в условиях пониженного засоления почв. По мере роста солероса засоленность почвы постепенно возрастает в связи с испарением воды и концентрацией солей в верхних слоях почвы. Таким образом, у большинства галофитов, в частности у солероса, семена произрастают при пониженной концентрации солей, затем происходит постепенное приспособление проростков к засолению, а для взрослых растений высокое содержание солей в почве является необходимым условием их развития.
В крайних условиях существования растительных организмов обнаруживаются различные приспособительные реакции, которые позволяют им выжить и дать потомство в неблагоприятных условиях. На Севере из-за низких температур и короткого периода вегетации половое размножение растений затруднено. В этих условиях преобладают способы вегетативного размножения. Они занимают и удерживают соответствующие участки ценозов посредством образования корневищ, обширной дернины. Однако при повышенной способности растений полярной зоны к вегетативному размножению функция генеративного развития у них не угасает. Возникли жизненные формы, у которых цветение завершается не в один, а в два года: цветковые почки в разные периоды развития могут перезимовывать, и растения дают плоды на втором году жизни. У многих злаковых растений тундры инициация цветения происходит за год до самого цветения. Кроме того, обнаруживается и иной тип репродуктивной адаптации. У тимофеевки альпийской побеги двух типов, каждый из которых образует генеративные органы в разные сезоны года. Поэтому резкое ухудшение условий в какой-либо из сезонов года не может полностью подавить генеративное размножение. Подобные типы адаптации растений к экстремальным условиям сочетаются с повышенной продуктивностью семян.
Примеры различных адаптивных признаков у высших растений почти неисчерпаемы. Строение отдельных органов растений (листьев, корней, стеблей) приспособлены к выполнению функций в условиях их местообитания. В засушливых условиях корни глубоко уходят в почву к водоносным слоям, а для более полного использования питательных элементов почвы растения образуют мочковатую разветвленную корневую систему, которая может использовать значительные объемы почв. У мангровых растений в условиях затопления образуются воздушные корни, через которые ткани обеспечиваются кислородом. Условия жизни растений определяют и форму листьев. Во влажных затененных местах форма листьев иная, чем в засушливых условиях. У растений засушливых мест обитания (ксерофиты) листья редуцированы в колючки (кактусы) или превращены в мясистые образования (суккуленты), либо очень утоньшены и имеют большое количество устьиц (полынь).
Если адаптация захватывает сферу физиологических функций или метаболических процессов, то следует говорить о физиологической адаптации растительных организмов. Различают два ее типа: 1) медленная, связанная с отбором и закреплением полезных генетических изменений; 2) быстрая, ненаследуемая, связанная с изменением метаболических процессов и физиологических функций.
Наследуемые физиолого-биохимические изменения связаны с усовершенствованием анатомо-морфологической структуры отдельных органов и изменением физиологических функций, в частности основного физиологического процесса - фотосинтеза. Эволюция растительных организмов осуществлялась путем совершенствования механизмов утилизации солнечной энергии и углекислого газа в соответствии с изменением на нашей планете условий радиационного и газового режима. Появились разнообразные био- и генотипы растений с фотосинтетическим аппаратом, приспособленным для эффективной работы в различных экологических условиях [22; 19; 35]. Ярким примером адаптации подобного рода являются различные пути первичной фиксации углекислого газа у C3-, C4- и САМ растений. Для эволюционно более молодых C4-растений характерна кранц-анатомия (дифференцировка тканей листа на клетки мезофилла и клетки обкладки). Они первично фиксируют CO2 через ФЕП-карбоксилазу и до взаимодействия с РБФ-карбоксилазой CO2 концентрируется в листе. Этот процесс происходит в клетках мезофилла листа путем карбоксилирования фосфоенолпирувата с образованием малата или аснартата (C4-кислот) и их транспорта в клетки обкладки листа (рис. 2). В них из поступивших органических кислот (C4-кислот) благодаря декарбоксилированию вновь освобождается CO2, которая взаимодействует с рибулезо-1,5-бисфосфатом (РБФ). Клетки обкладки листа C4-растений, как и клетки мезофилла листа C3-растений, имеют энзиматический аппарат, катализирующий отдельные реакции как восстановительного, так и окислительного превращения рибулезо-1,5-бисфосфата. Однако у C4-растений дополнительные реакции (карбоксилирование фосфоенолпирувата и декарбоксилирование органических C4-кислот) способствуют повышению концентрации CO2 в клетках обкладки листа. А это создает благоприятную ситуацию для реакции карбоксилирования рибулезо-1,5-бисфосфата, а следовательно, для восстановительного цикла фотосинтеза, а не окислительного цикла фотодыхания. Следовательно, у C4-растений происходит редукция фотодыхания. У C3-растений при сильном освещении и повышенной концентрации O2 интенсивность фотодыхания может многократно превосходить интенсивность обычного темнового дыхания и составить до 50 - 75% скорости фотосинтеза.
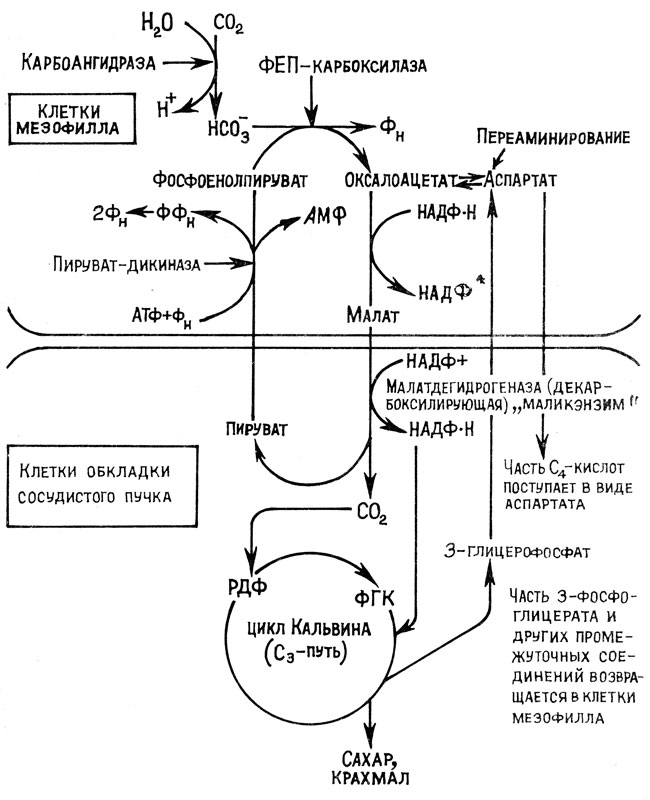
Рис. 2
Как предполагают, C4-растения имеют ряд преимуществ по сравнению с C3-видами. Они лучше адаптированы к жаркому сухому климату, имеют более интенсивный фотосинтез, более экономно расходуют воду и т. п. В связи с этими особенностями, а также потому, что у них отсутствует фотодыхание, считается, что они значительно более продуктивны, чем C3-растения.
CAM-растения (например, кактусы) в условиях очень сильного недостатка влаги фиксируют CO2 в ночные часы через ФЕП-карбоксилазу с образованием малата, когда у них открыты устьица и идет транспирация (рис. 3). Днем, во время сильной засухи, устьица этих растений закрываются, практически прекращается транспирация и поглощение CO2 из воздуха. Накопленный ночью малат декарбоксилируется, а образующаяся при этом эндогенная углекислота используется для образования метаболитов цикла Кальвина и крахмала.
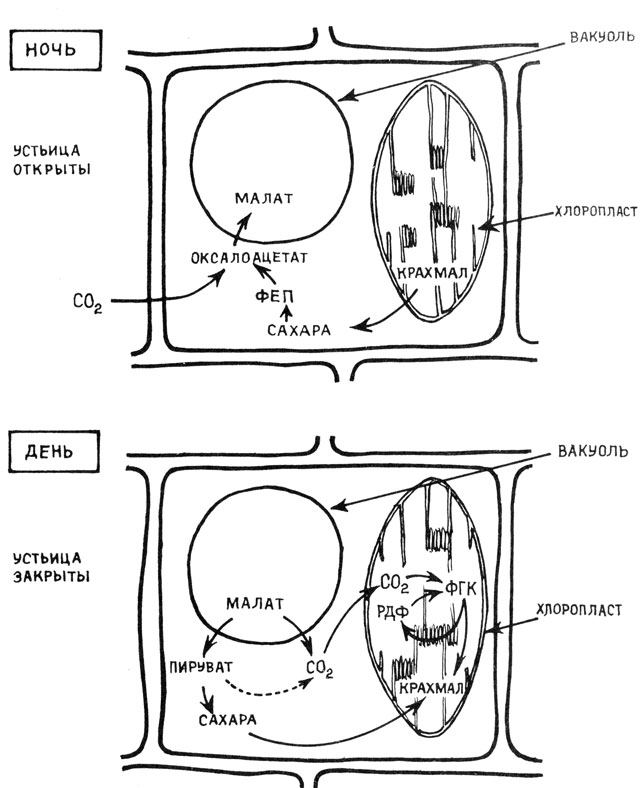
Рис. 3. Метаболизм органических кислот по типу толстянковых (САМ-метаболизм)
C3-, C4-растения, а также суккуленты (CAM-растения) различаются по ряду признаков: по анатомическому строению, интенсивности фотосинтеза, фотодыхания, активности карбоксилирующих ферментов и др.
В большинстве районов земного шара растительный мир широко представлен видами с различными морфофизиологическими свойствами и путями первичной фиксации CO2.
По анатомическому строению ассимилирующих органов растения Каракумов можно разделить на три группы: 1) виды с обычным строением мезофилла (C3-растения); 2) своеобразная группа видов, например саксаул, у которых функцию листа выполняют суккулентные ассимилирующие побеги; под эпидермисом у них располагаются два слоя ассимилирующих клеток; 3) виды, имеющие специфические клетки обкладок, характерные для C4-растений. В Каракумах, где температура воздуха колеблется от +10°С утром до +30 - 35°С днем, C3- и C4-растения представлены многолетниками и однолетниками. Среди однолетников C3-растений в основном встречаются эфемеры с коротким жизненным циклом, приуроченным к умеренному по температуре и влаге периоду.
Из растений, успешно произрастающих в Каракумах, наибольший интерес представляет саксаул с хорошо развитой корневой системой, которая достигает грунтовых вод. В связи с этим развитие его не лимитируется водой. Кроме того, саксаул является галофитом, засоление почвы не мешает ему интенсивно накапливать биомассу. Он хорошо сочетает свойства C4-растений с интенсивной транспирацией, что и обеспечивает данному растению максимальную продуктивность. Саксаул является хорошим примером сбалансированного существования, моделью гомеостаза.
В основном флора Каракумов представлена C3- и САМ-растениями, и встречается лишь несколько Севидов. В связи с этим нельзя считать доказанным, что во всех случаях C4-растения доминируют в жарких и засушливых условиях, т. е. что они лучше, чем C3-виды, адаптированы к этим условиям. По показателям, характеризующим фотосинтез, также нельзя сказать, что C4-растения имеют определенные преимущества но сравнению с C3-видами [19]. Максимальная наблюдаемая интенсивность фотосинтеза некоторых C3-видов в Каракумах равна таковой C4-растений или несколько выше. Кроме того, C3-, C4-растения мало различаются и по положению верхних температурных границ фотосинтеза. В связи с этим нельзя сказать, что C4-растения лучше адаптированы к повышенной температуре.
В крайне жестких условиях пустыни некоторые растения совмещают в пределах ассимиляционного аппарата хлорофиллоносные ткани с различным типом углеродного метаболизма, которые могут обеспечивать круглосуточную фиксацию углекислого газа [12]. Все возникшие эволюционные пути превращения углерода являются модификацией цикла Кальвина (C3-фотосинтез), доминирующего у растений и сохраняющегося как основной путь ассимиляции CO2.
От устойчивости белоксинтезирующего аппарата в стрессовых условиях зависит интенсивность белкового синтеза, а следовательно, и жизнеспособность клеток в данных условиях. При адаптации у ряда устойчивых растений происходит накопление растворимых белков в их тканях. Кроме того, обнаруживается синтез ряда новых белков, возможно играющих протекторную роль.
Для физиологической адаптации большое значение имеет способность макромолекул выполнять свои функции в широком интервале интенсивностей тех или иных факторов. Такая способность обеспечивается гетерогенностью белковых молекул и молекулярным отбором определенных форм, которые проявляют максимальную активность в неблагоприятных условиях среды, к которым адаптируется растительный организм.
В 1962 г. интальянским цитогенетиком Ф. Ритосса было показано, что при нагревании личинок дрозофилы до 37°С изменяется положение пуффов в гигантских хромосомах слюнных желез. При этом тепловой шок индуцирует образование новых и ингибирует существовавшие ранее пуффы. Пуффинг свидетельствует об активности генов. Следовательно, можно полагать, что в условиях гипертермии происходят изменения генной активности. Позднее было обнаружено, что тепловой шок индуцирует у дрозофилы синтез ряда полипептидов, получивших название белков теплового шока (БТШ), и ингибирует синтез тех белков, которые образуются при "нормальных" физиологических температурах [8].
Ответные реакции генома на тепловой шок включают несколько этапов. На первом из них физический фактор (температура) оказывает влияние на структуру внутриклеточного рецептора. Возникающие при этом сигналы изменяют экспрессию генов, а это, в свою очередь, индуцирует или репрессирует синтез специфических мРНК или их трансляцию. Результатом всех этих изменений является синтез специфических полипептидов.
Работы последних лет показали, что в ответ на действие неоптимального внешнего фактора в растительной клетке может синтезироваться определенный набор стрессовых или шоковых белков. Наиболее изучен этот процесс у бактерий, птиц, рыб, дрожжей, дрозофилы, а также у высших растений (табака, кукурузы, сои и т. д.). Показано, что повышенные температуры, значение которых выходит за пределы толерантной зоны, вызывают изменение экспрессии генома, в результате чего полностью (у животных) или частично (у растений) подавляется общий синтез белка, характерный для нормальной температуры (в пределах толерантной зоны), и синтезируются белки теплового шока.
Известно, что основная масса БТШ кодируется ядерным геномом, однако имеются белки теплового шока митохондриального происхождения.
Действие низкой температуры вызывает у растений замедление (или даже остановку) синтеза большей части белков, что связано с нарушениями либо в белоксинтезирующем аппарате, либо в регуляции генной активности, и появление новых белков. Образование их обусловлено, вероятно, синтезом de novo и в основном на 80 S рибосомах.
Ответ генома на стресс (тепловой, холодовой) является быстрым и обнаруживается в течение первых 20 - 60 мин воздействия. Максимальный синтез стрессовых белков в растительных клетках наблюдается, как правило, в течение 1 - 3 часов действия температур. При увеличении действия экстремальных температур синтез стрессовых полипептидов в клетках уменьшается и восстанавливается синтез белков, происходящий в нормальных температурных условиях.
Такие растения, как соя, рис, куколь, реагируют на действие низких (4 - 10°С) и высоких (40 - 50°С) температур очень быстро. При этом изменяется скорость экспрессии одних и тех же или родственных генов, т. е. отдельные генные локусы активируются (или возможно ингибируются) как при охлаждении, так и при нагревании. При этом неспецифические реакции организма на температурный фактор проявляются не только в изменении одних и тех же генов, но и в синтезе одних и тех же клеточных белков. Например, под влиянием как низкой (10°С), так и высокой (30°С) температур в листьях проростков риса синтезируется de novo полипептид с Мм 19 кД [25].
При высоком уровне солей в среде в клетках появляются белки осмотического шока (БОШ). Полагают, что они регулируют метаболизм клетки и поддерживают его на достаточном уровне, т. к. БОШ устойчивы к высоким концентрациям солей. При снижении уровня солей белки осмотического шока исчезают и усиливается синтез белков, характерных для нормальных солевых условий.
Стрессовые белки обнаруживаются у растений при действии других экстремальных факторов (аноксии, химических агентов, заражении).
В настоящее время большое внимание уделяется исследованию роли стрессовых белков, в частности ВТШ, в механизмах ответных реакций клеток на действие экстремальных факторов среды [57; 66; 67; 58; 25].
В 1984 г. Дж. Ки с сотрудниками удалось идентифицировать гены низкомолекулярных белков, активность которых возрастала в ответ на тепловой и осмотический шок, ионы тяжелых металлов, ингибиторы окислительного фосфорилирования и другие неблагоприятные воздействия. Все БТШ - кислые белки и по размеру условно делятся на высоко-(Мм 68 - 100 кД) и низкомолекулярные (Мм 15 - 30 кД).
В растительных клетках тепловой шок вызывает формирование специфических структур - гранул теплового шока. Изучение их состава показало, что они содержат БТШ 70 и 17 кД, а также мРНК. Установлено, что подобно тепловому шоку другие стрессы (осмотический, солевой) также вызывают формирование стрессовых гранул (СГ). По-видимому, образование СГ является универсальным ответом растительных клеток на шок. При чем стрессовые гранулы, обнаруженные при солевом и осмотическом шоках, содержат белки, имеющие общие иммунодетерминанты с белками теплового шока.
Показано, что в ответ на тепловой шок происходит образование перихроматиновых гранул в ядре, цитоплазме, а также в пластидах и митохондриях. Вероятно, функциональная роль БТШ в составе гранул теплового шока заключается в защите временно нетранслируемых в период теплового шока мРНК, необходимых для синтеза нормальных белков, от повреждающего действия гипертермии [51; 67]. Предполагается, что гранулы теплового шока предохраняют внутриклеточные биополимеры от разрушения. Так, они могут защищать РНК от действия нуклеаз, активность которых при высокой температуре возрастает.
В ядре и ядрышке БТШ образуют гранулы, которые связывают матрицы хроматина, необходимые для нормального метаболизма. По окончании стресса эти матрицы освобождаются и начинают функционировать.
При стрессовых температурах (41°С) низкомолекулярные БТШ ассоциируются с митохондриями. Как показал Войников с сотрудниками (1988), БТШ участвуют в регуляции функциональной активности митохондрий кукурузы, способствуя увеличению сопряженности дыхания и фосфорилирования, а также интенсивности окислительных процессов [9; 10].
Множественность хлоропластных БТШ, как низкомолекулярных, так и высокомолекулярных форм, предполагает выполнение ими различных функций. В частности, БТШ хлоропластов участвуют в сборке больших и малых субъединиц РВФК, образовании мультисубъединичного функционального комплекса данного фермента, в адаптивных изменениях уровня фотосинтетической активности.
В настоящее время стрессовые белки разделяют на группы, каждая из которых состоит из белковых изоформ, кодируемых сходными по структуре генами, образующими семейства генов.
БТШ 90 - в основном растворимые цитоплазматические фосфопротеины, которые синтезируются при оптимальной температуре, а в условиях теплового шока синтез их значительно активируется. Полагают, что некоторые белки данного семейства могут осуществлять временную регуляцию активности некоторых рецепторных молекул.
БТШ 70 - мобильные внутриклеточные белки; быстро перемещаются при тепловом шоке из цитоплазмы в ядро, концентрируясь в ядрышке, и возвращаются в цитоплазму при понижении температуры. Полагают, что данные белки играют значительную роль в нормализации процессов жизнедеятельности клеток в стрессовых условиях.
БТШ 00 - конститутивные компоненты митохондрий и хлоропластов. Хлоропластный БТШ 60 известен как белок, способствующий сборке РБФК.
БТШ 20 - низкомолекулярные белки, участвующие в образовании высокоупорядоченных структур (гранул теплового шока).
БТШ 8,5 (убиквитины) - высококонсервативные белки, состоящие из 75 - 76 аминокислотных остатков, которые присоединяются к N-концу полипептида и делают его доступным для протеолиза. Убиквитины осуществляют селективную деградацию денатурированных белковых молекул, которые накапливаются в стрессовых условиях.
Следовательно, быстрый и эффективный синтез БТШ в ответ на тепловой шок имеет важное термоадаптивное значение. БТШ осуществляют одну из основных стратегий защиты растений в экстремальных условиях.
Полупериод жизни стрессовых белков около 20 часов, в течение которых клетки сохраняют терморезистентность. Очевидно, эти белки необходимы на начальных этапах стресса для запуска более длительных адаптивных внутриклеточных процессов (синтеза ряда протекторных соединений, изменения структуры мембранных компонентов и др.).
Специфичность ответных реакций, связанных с синтезом стрессовых белков, заключается в том, что эти полипептиды обнаруживаются лишь после воздействия экстремальных факторов и отсутствуют в нормальных условиях. Однако эта специфичность относительна: разные факторы (тепло, холод, аноксия, химические агенты и т. д.) вызывают сходные изменения экспрессии генов и синтез одних и тех же внутриклеточных белков [8].
Физиологическая адаптация к стрессовым воздействиям обусловлена также способностью организма регулировать внутриклеточный уровень различных защитных веществ, в частности, низкомолекулярных азотсодержащих соединений: пролина, диамина (путресцина) и полиаминов (спермидина, спермина) (рис. 4).
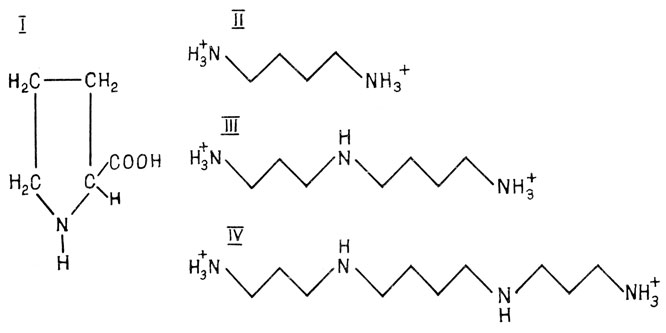
Рис. 4. Структурные формулы некоторых протекторных соединений: I - пролин, II - путресцин H+3N - (CH2)4 - NH+3, III - спермидин H+3N - (CH2)3 - NH+2 - (CH2)4 - NH+3, IV - спермин H+3N - (CH2)3 - NH+2 - (CH2)4 - NH+2 - (CH2)3 - NH+3
Первые сообщения о повышенном содержании пролина в условиях засоления и водного дефицита относится к 60-м гг. Стюарт с сотрудниками показал, что при водном стрессе происходит накопление свободного пролина в растительных клетках. Сверхпродукция пролина обнаруживается в присутствии высоких концентраций солей (170 мМ NaCl). Существуют пролинаккумулирующие виды (ячмень, шпинат, арахис, хлопчатник и др.), в которых содержание этой аминокислоты может повышаться при стрессе в 100 и более раз. Первичным триггером, запускающим синтез пролина в растениях в условиях водного дефицита и засоления, большинство исследоваталей считают потерю клеткой тургора.
Известны два пути биосинтеза пролина у высших растений - глутаматный (основной) и орнитиновый (катаболитный). Глутамат является не только предшественником, но и первым продуктом деградации пролина (рис. 5). Накопление пролина при водном и солевом стрессах обусловливается увеличением скорости его образования из глутамата и частично по катаболитному пути из орнитина.
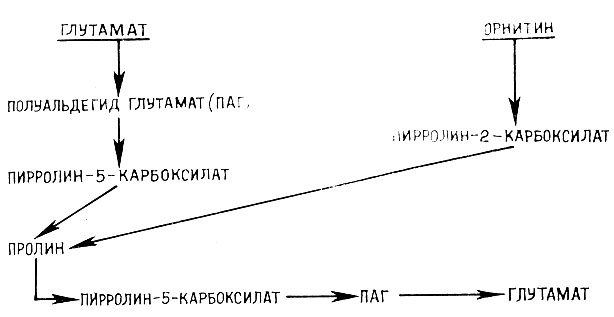
Рис. 5. Пути биосинтеза и деградации пролина в растениях
Пролин действует как осморегулятор в клетке. Он способствует удержанию в ней определенного количества воды. Пролин - инертный осмотик, в высокой концентрации он не оказывает влияния на ферментативные реакции. Важная роль пролина в солеустойчивости растений связана со способностью его снижать токсическое действие солей, в частности NaCl. Высокая растворимость пролина в сочетании с его очень низкой способностью ингибировать ферменты может увеличивать растворяющий объем клетки и тем самым снижать концентрацию солей в цитозоле.
Благодаря своим гидрофильным группам молекулы пролина способны образовывать агрегаты, которые, связываясь с белком, повышают растворимость и защищают его от денатурации. В присутствии пролина восстанавливается утраченная при дегидратации клеток (при засухе, высокой концентрации солей в среде) оводненность мембран.
Значимость пролина для осмоадаптации установлена не для всех растительных организмов. В частности, не обнаружена строгая корреляция между уровнем пролина и устойчивостью к водному стрессу у кукурузы.
В условиях стресса адаптивный потенциал растительных клеток может быть обеспечен за счет других протекторных соединений (редуцирующих сахаров, полиаминов, бетаинов, органических анионов и др.).
Диамины и полиамины (путресцин, спермин, спермидин) - соединения с широким спектром действия. Их содержание возрастает, с одной стороны, в активно пролиферирующих клетках, а с другой - при действии неблагоприятных факторов. Высокий эндогенный уровень диаминов и полиаминов у солеустойчивых клеток на среде с NaCl достигается благодаря повышенному содержанию предшественников (орнитина, метионина) и активирующему действию солей на их биосинтез; Запуск ускоренного биосинтеза данных веществ в клетках после пересадки их на среду с NaCl наблюдается уже через час действия соли.
В эукариотических клетках путресцин образуется за счет декарбоксилирования орнитина. Образование спермина происходит путем конденсации путресцина и пропиламинного остатка декарбоксилированного S-аденозилметионина при участии фермента спермидинсинтетазы (рис. 6). Аминопропилтрансфераза (сперминсинтетаза) переносит на спермин еще одну пропиламиновую группу с образованием спермина.
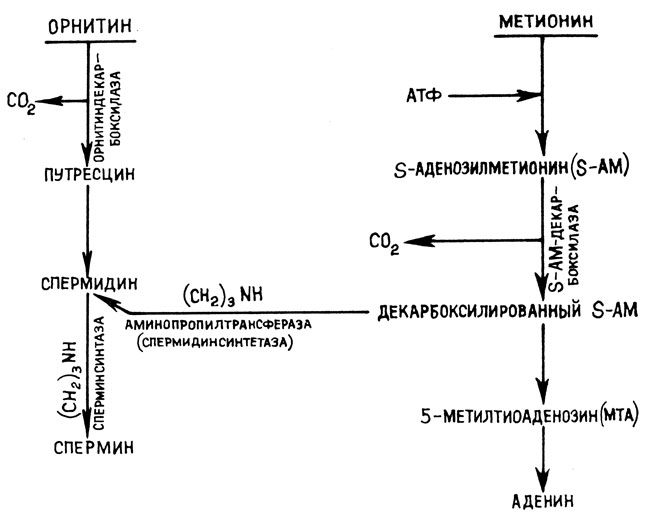
Рис. 6. Пути биосинтеза ди- и полиаминов
В физиологических концентрациях, пределы которых в растительной клетке достаточно широки (1 - 10 мкМ), ди- и полиамины действуют как ростстимулирующие вещества и стабилизаторы полимерных структур, а в высоких концентрациях они сами могут быть причиной стресса. Полиамины способны предотвращать повреждения, вызванные морозом, засухой, озоном и действием солей [55]. Механизм действия этих соединений во многом определяется их основной природой и легкостью образования ионных связей с отрицательно заряженными группами различных биополимеров. Катионные свойства полиаминов проявляются при связывании их с нуклеиновыми кислотами, фосфолипидами мембран. Благодаря образованию подобных комплексов полиамины стабилизируют нативную структуру ДНК, РНК и рибосом. Диамины и полиамины в широких концентрациях (1 - 10 мкМ) действуют на растительную клетку как ростовые вещества (повышают скорость макромолекулярных синтезов, точность считывания информации, стабильность структур, проницаемость мембран; снижают РНКазную и протеазную активность и задерживают подобно цитокининам старение). Поддержание нативной структуры нуклеиновых кислот, стабилизация мембран, торможение лизиса клеточных структур, стимуляция процессов, связанных с синтезом белка, дают основание интерпретировать роль диаминов и полиаминов в клетке как протекторную. Например, у растений, произрастающих в условиях засоления, нарушается функциональная активность клеточных и субклеточных структур и тормозятся процессы роста. В связи с этим поддержание повышенного внутриклеточного уровня ди- и полиаминов должно способствовать адаптации к стрессу.
Высокая аккумуляция путресцина, спермидина и спермина обнаружена у растений при солевом и осмотическом шоке. Солеустойчивые клетки табака, постоянно культивируемые в присутствии NaCl, содержали путресцина, спермидина и спермина в 2 раза больше, чем на среде без NaCl. Судя по высокой скорости их клеточного роста, жизнеспособности клеток, повышенный эндогенный пул этих соединений является для них нормой, обеспечивающей гомеостаз в условиях засоления. Повышение концентрации нолиаминов в физиологически допустимых пределах в растительных клетках при действии неблагоприятных факторов можно рассматривать как ответную реакцию приспособительного характера.
Существуют и другие механизмы защиты клеток в условиях водного и солевого стресса. В частности, бетаин [(CH3)3N+-CH2COO-] выполняет в растительных клетках защитную функцию при высоких концентрациях солей в среде. Бетаины - это метилированные производные аминокислот и аминов. Они обнаружены у многих растений: в пшенице, ячмене, вике, листьях табака и др. Большое количество бетаина содержится в сахарной свекле (в корнях - 0,6%, в старых листьях - 3% на сухое вещество).
Адаптация растений к солевому стрессу может быть связана за счет изменения заряда внутриклеточных биополимеров в результате их метилирования. А это может приводить к увеличению устойчивости внутриклеточных макромолекул к дегидратации и действию токсических соединений, которые образуются при увеличении ионной силы. Главными метильными донорами в растительных клетках являются бетаин, метионин, S-аденозилметионин (S-AM). При солевом и водном стрессе S-AM может быть использован на синтез дополнительных количеств полиаминов и этилена. У солеустойчивых растений уровень бетаинов, которые являются донорами метильных групп, повышается. Для реакций трансметилирования с участием бетаинов не требуются дополнительные источники энергии, так как они сами обладают большим ее запасом.
Белки, нуклеиновые кислоты, углеводы модифицируются при помощи метилтрансфераз, которые переносят метильные группы от S-AM и бетаинов к специфическим участкам полимерных молекул. Перенос метильных групп от S-AM на специфические участки ДНК - самая распространенная модификация ДНК. Метилирование изменяет функциональную активность ДНК и других внутриклеточных полимеров и повышает их устойчивость к различным воздействиям.
Механизм адаптации на уровне целых растений заключается в изменении донорно-акцепторных отношений, которые регулируются посредством изменений как скорости ростовых процессов (акцептора ассимилятов), так и интенсивности фотосинтеза (донора и ассимилятов). В работах А. Т. Мокроносова с сотрудниками обнаружена положительная корреляция между интенсивностью фотосинтеза и относительной скоростью роста, которая сохраняется в онтогенезе растений [35]. Фотосинтез в значительной степени детерминирован процессами роста и определяется всей совокупной активностью аттрагирующих систем целого растения. Это справедливо при условии, когда внешние факторы (CO2, свет и др.) не лимитируют скорость фотосинтеза.
В экстремальных условиях у растений наблюдается торможение процессов клеточного деления и особенно роста клеток в фазе растяжения. Вследствие этого снижается "запрос" клеток на продукты фотосинтеза и уменьшается их отток из хлоропластов. Например, подавление роста растений на холоде сопровождается 4 - 5-кратным ингибированием транспорта продуктов фотосинтеза из хлоропластов в клетку. В свою очередь высокий уровень ассимилятов в хлоропластах может ингибировать фотосинтез.
Нормализация нарушенных стрессовыми условиями донорно-акцепторных отношений может происходить на уровне изменений первичной структуры внутриклеточных мембран. Стремясь к нормализации донорно-акцепторных отношений, растения увеличивают в стрессовых условиях объем запасающей (акцепторной) способности мембранных компонентов. При действии экстремальных температур в клетках, как правило, происходит увеличение объема мембран эндоплазматического ретикулума, изменение структурной организации митохондрий, хлоропластов и других внутриклеточных структур. В частности, при холодовом закаливании растений в хлоропластах возрастает доля тилакоидов стромы относительно тилакоидов гран. Стресс, вызванный действием низких температур, приводит также к увеличению содержания высоковосстановленных соединений, при этом отношение липиды/белки в тилакоиддых мембранах хлоропластов возрастает. Например, при холодовом закаливании ряда растений (озимых ржи, ячменя, пшеницы и шелковицы) отношение липиды/белки в хлоропластных мембранах увеличивается на 160 - 230%. Кроме того, у термоустойчивых растений увеличивается отношение ненасыщенные жирные кислоты/насыщенные формы. В результате при отрицательных температурах мембраны закаленных растений не "застывают" и сохраняют в данных условиях функциональную активность.
Таким образом, у. устойчивых растений при закаливании структурообразовательный процесс, являясь акцептором ассимилятов, участвует в регуляции фотосинтетического метаболизма и обеспечивает поддержание донорно-акцепторных отношений у растений в стрессовых условиях.
Механизмы физиологической адаптации проявляются и в процессе регуляции онтогенеза растений, проходящего в конкретных условиях существования и в зависимости от внешних экологических факторов, которые меняются в течение года и не одинаковы в разных климатических зонах. У растений с продолжительным периодом развития в регуляции онтогенеза существенную роль играет эндогенный отсчет времени, механизм "биологических часов" или "физиологических часов". В настоящее время центральную роль в биологическом хронометрировании отводят эндогенной циркадной (околосуточной) ритмичности процессов жизнедеятельности. Вместе с тем эндогенная регуляция онтогенеза должна корректироваться влиянием напряженностей внешних факторов. Сочетание эндогенного контроля онтогенеза с восприятием внешних сигналов и регулирует проявление ряда физиологических состояний, в том числе разных типов покоя семян и вегетирующих растений. Адаптационные приспособления организма, основанные на способности изменять интенсивность жизнедеятельности, несомненно имеют большое значение для формирования устойчивости растений.
Организмы в течение онтогенеза находятся в различных состояниях: активной жизнедеятельности, замедленной и ограниченной жизнедеятельности (покой, характеризующийся пониженным обменом, и состояние обратимого прекращения жизнедеятельности - анабиоз, характеризующийся отсутствием обмена). Кроме того, существуют промежуточные состояния между анабиозом, или покоем, и активной жизнедеятельностью. У многих растений в течение суток, сезона обнаруживается ритмичность ростовых и других физиологических процессов. Под ритмичностью понимают регулярно повторяющуюся смену периодов активного роста и периодов его торможения. Различают суточную, обусловленную изменением освещения днем и ночью, и сезонную (годовую) ритмичность, обусловленную и вменениями температуры, содержания воды и других факторов в течение года. Эндогенная природа биологических ритмов, в частности суточных и сезонных, убедительно показана и признается сегодня большинством исследователей. Однако между живой системой и окружающей средой имеются сложные взаимосвязи. Факторы внешней среды проявляются прежде всего в качестве датчиков времени, т. е. в установлении соответствующих фазовых и частотных соотношений функций организма с окружающей средой. Устойчивость организма определяется, по-видимому, созданием внутренних связей между скоростью, направленностью отдельных физиологических процессов и периодичностью действия среды. На ранних этапах онтогенеза ритмика физиологических процессов более пластична, поэтому организмы в молодом возрасте быстрее приспосабливаются к новым условиям окружающей среды.
Состояние растений, когда у них отсутствует видимый рост, принято называть покоем. Органы, находящиеся в покоящемся состоянии (почки деревьев, клубни и корневища многолетних травянистых растений), дышат, в них идет превращение запасных веществ, но скорость всех этих процессов мала. Покой может наступать в любое время года: зимой - у деревьев, летом во время засухи - у степных луковичных растений, осенью - у свежесобранных клубней картофеля. В начале зимы в почках и ветвях деревьев постепенно исчезает крахмал и накапливаюся сахара и липиды, что повышает устойчивость клеток и тканей к низким температурам. В период покоя у некоторых растений происходит скрытый рост, закладываются цветки. В покое могут находиться не только целые растения, но и отдельные органы. Пазушные почки на растущих побегах после короткого периода роста впадают в состояние покоя, хотя верхушечная ночка и листья продолжают интенсивный рост. Состояние покоя характерно и для тропических растений, несмотря на круглогодичные благоприятные условия.
Все многообразные виды покоя можно разделить на два основных типа: вынужденный и глубокий (или органический). Вынужденный покой - такое состояние, когда видимого роста нет из-за отсутствия необходимых условий в среде (пониженные температуры, отсутствие света, воды и кислорода могут задержать прорастание семян). Достаточно создать оптимальные условия, и вынужденный покой прекращается.
Отсутствие видимого роста, несмотря на благоприятные условия внешней среды, называют глубоким (или органическим) покоем. Это - наследственно закрепленное состояние, возникшее в процессе эволюции под влиянием неблагоприятных условий (зима, сухой сезон) и трудно прерываемое с помощью воздействия ряда факторов (тепла, воды и др.). Состояние глубокого покоя возникает до наступления неблагоприятных условий существования. Например, в степи летний покой начинается задолго до засухи. У деревьев в умеренной зоне вхождение в покой происходит задолго до зимы - осенью при положительных температурах и высокой влажности. Сигналом для перехода в состояние глубокого покоя является продолжительность дня, которую воспринимают листья. У растений умеренной зоны переход в состояние покоя начинается с наступлением коротких дней, у степных и тропических растений - длинных дней.
Следовательно, покой - важное приспособительное свойство растительного организма. В состоянии покоя клетки обладают максимальной способностью к выживанию в неблагоприятных условиях естественной среды и в условиях, с которыми растение обычно в природе не встречается (например, рентгеновское излучение).
В процессе анабиоза (криптобиоза) в организме совсем прекращаются биохимические процессы, однако жизнеспособность сохраняется, т. е. организмы, находящиеся в состоянии анабиоза, могут восстанавливать полную жизнеспособность при соответствующих условиях. В природе анабиоз встречается при определенном сочетании некоторых условий среды (например, низкой влажности и обеднения или лишения кислорода). Так, семена многих растений могут сохраняться в течение многих сотен лет в глубоких и сухих слоях почвы, обедненных кислородом и не подвергающихся резким колебаниям температуры. Состояние анабиоза характеризуется наличием только связанной воды в виде гидратных оболочек полярных групп веществ, составляющих остов протоплазменного геля [13]. Связанная вода не растворяет вещества, находящиеся в промежутках гелевой структуры, что и является одной из причин прекращения биохимических процессов при полном анабиозе. В состоянии анабиоза могут находиться низшие пойкилогидрические растения (мхи, лишайники), которые при уменьшении влажности воздуха высыхают до воздушно-сухого состояния. Например, высшие лишайники содержат 6 - 9% воды, влажность зеленого мха Pleurozinm в сухие дни понижается до 8%, кукушкина льна - до 9, сфагнума - до 7,8% абсолютно сухой массы. Оводненность тундровых лишайников в жаркие дневные часы при низкой (30 - 40%) относительной влажности воздуха составляет 6 - 8% абсолютно сухой массы. В результате такого глубокого высыхания эти растения переходят к анабиозу и фотосинтез прекращается.
Лишь немногие высшие растения способны к анабиозу при высыхании всех их частей. Среди покрытосеменных растений глубокое обезвоживание без потери жизнеспособности могут переносить только несколько видов: Bamondia mathalia (сем. геснериевых), Trilepis pilosa и Carex physodes (сем. осоковых). Эти растения произрастают обычно в условиях, характеризующихся периодами сильной засухи. Например, Carex physodes хорошо представлен в пустыне Каракумы. Эти растения могут высохнуть настолько, что их легко растереть в порошок, однако после увлажнения жизнеспособность их восстанавливается.
Высшие гомойогидрические растения, которые регулируют свой водный обмен, как правило, погибают при глубоком увядании. Однако семена этих растений являются пойкилогидрическими и способны высыхать до воздушно-сухого состояния с переходом к анабиозу. Влажность созревших семян в зависимости от вида составляет около 5 - 15% их массы. Легкость воздушно-сухих семян создала предпосылки для их расселения на большие расстояния. Это обеспечило завоевание покрытосеменными растениями огромных пространств на земле, что было бы невозможно при вегетативном размножении. Расселение шло не только в пространстве, но и "во времени", благодаря анабиозу. Семена, пребывающие в течение ряда лет в состоянии анабиоза, сохраняют всхожесть, что позволяет им прорастать в новых местах при подходящих условиях.
Адаптация может проявляться и в популяционных эффектах. Гетерогенная популяция в отношении физиологических свойств составляющих ее элементов более приспособлена к широкому диапазону меняющихся факторов среды, чем гомогенная популяция. Если популяция состоит из субпопуляций ксеро- и мезоморфных структур, то в засушливых условиях преимущества будут иметь ксероморфные (устойчивые к засухе), а во влажных - мезоморфные растения. В этом случае проявляется основной принцип биологических систем: гетерогенная система более устойчива, чем гомогенная структура. В связи с этим, по-видимому, важно использовать при посевах широкий набор семян высокопродуктивных сортов, которые могут ежегодно давать высокие урожаи.
Кроме того, в природе существуют симбиотические организмы, состоящие из различных особей, находящихся в морфологическом и физиологическом единстве. Примерами симбиотических отношений являются микоризы (симбиоз гриба и корня высших растений), клубеньки бобовых растений (симбиоз азотофиксирующих бактерий и бобовых), лишайники (симбиоз гриба и синезеленых водорослей) и др. При симбиотических отношениях организмов проявляется взаимоадаптация, в результате которой устойчивость симбионта повышается.
Классическим примером симбиоза являются лишайники, которые отличаются высокой устойчивостью к дефициту воды и высоким температурам. В данном случае симбиоз выгоден для обоих партнеров вследствие сбалансированного автотрофного и гетеротрофного типов питания. Водоросль в условиях симбиоза способна к усилению фотосинтеза. При выделении ее из лишайника способность к гиперфункции фотосинтеза исчезает. По-видимому, образование и транспорт больших количеств органических соединений клетками водоросли в лишайнике поддерживается воздействиями со стороны гетеротрофного симбионта. Следовательно, в симбиотической системе проявляется адаптивная реакция компонентов симбиоза, распространяющаяся на биохимические и морфологические явления. Взаимодействия двух систем у лишайников значительно повышают устойчивость организма. Например, накипные лишайники, растущие на камнях или горных породах, в пустынях способны без вреда для себя переносить температуру +70 ÷ +80°С. При высокой температуре слоевище лишайников теряет большую часть воды, и активная жизнедеятельность, включая фотосинтез, протекает только в утренние часы, когда сохраняется влага выпавшей росы. Взаимоадаптация двух систем у лишайников обеспечила, по-видимому, и долговечность их обитания на Земле.
Примеры адаптационных признаков у растительных организмов почти неисчерпаемы. Адаптация может проявляться на всех уровнях организации живого. Устойчивость растений обусловливается способностью организмов к морфоанатомической, физиологической и экологической адаптации. Благодаря различным типам адаптации и способности организмов к репарации многие растения занимают обширные ареалы и в определенных условиях произрастания изменяются в очень широких пределах. Для физиологической адаптации немаловажное значение приобретает способность макромолекул выполнять свои функции в широком интервале интенсивностей тех или иных факторов. Физиологическая адаптация является результатом явлений, происходящих на уровне транскрипции и трансляции генов, активных перестроек макромолекул и мембранных компонентов, создания в клетках и тканях определенного гормонального баланса.
Таким образом, ответ растительных организмов на действие экстремальных факторов среды включает стрессорные реакции и процессы специализированной адаптации. Биологическое значение стрессорных реакций заключается в срочной мобилизации всех защитных систем для сохранения жизнеспособности клеток в данных экстремальных условиях. Специализированная адаптация, определяя надежность функционирования всех молекулярных, клеточных, тканевых систем, обеспечивает сохранение высокого уровня продуктивности организмов в данных условиях.
Сохранение жизнеспособности растений в экстремальных условиях в значительной степени зависит от наличия и активности внутриклеточных репараторных систем.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'