1.3. Факторы внешней среды. Специфические и неспецифические ответные реакции
Растения в процессе роста и развития испытывают воздействие неблагоприятных факторов среды, к которым относятся температурные колебания, засуха, избыточное увлажнение, засоленность почв и т. д. Если данные факторы действуют на растения в пределах толерантной зоны и данное воздействие непродолжительное, то не наблюдается существенных нарушений структуры и физиологических функций растений, что обусловлено способностью организмов сохранять относительно стабильное состояние при изменяющихся условиях, то есть поддерживать гомеостаз. Если изменения внешних факторов достаточно велики (выходят за пределы толерантной зоны), возникают достаточно быстро и продолжаются достаточно долго, то данные факторы являются раздражителями. Раздражитель - это любое внешнее воздействие, достигшее пороговой силы. Способность живых структур отвечать на действие раздражителей носит название раздражимости. Наличие свойства раздражимости позволяет клеткам приспосабливаться к среде и тем самым защищать и сохранять свою жизнь. Именно поэтому К. Бернар назвал раздражимость "первым двигателем жизненных функций живого" [7].
В естественной среде обитания растения находятся в условиях постоянно изменяющихся факторов: биологических (вирусы, бактерии, грибы, конкуренция с другими растениями, влияние животных и др.); химических (вода, элементы питания, гормоны, газы, гербициды, инсектициды, фунгициды и др.); физических (освещенность, температура, излучение, механические факторы и др.) Одной из отличительных особенностей среды, в которой развивается растение, является ее непостоянство. Развитие растения приспособлено не к какому-либо одному фактору внешней среды, а к определенному сочетанию, комплексу условий.
Следует иметь в виду, что в ряде случаев повреждение организма, вызванное факторами физической природы, опосредуется химическими агентами, возникающими в растении при действии на него физического фактора. С медиаторами химической природы связано действие ионизирующей радиации, высокой температуры и ряд других физических факторов. По физиологическому значению факторы среды делят на адекватные и неадекватные. Адекватные - это естественные факторы, сопутствующие виду в процессе его эволюции, к восприятию которых он приспособлен и чувствительность к которым у данных организмов очень высока. Неадекватные - это искусственные факторы, которые не могли принимать участия в становлении вида и для восприятия которых клетки специально не приспособлены. В связи с этим реакции на неадекватные факторы, даже если они действуют в небольших дозах, могут привести к повреждениям клеток и тканей.
Действие фактора может быть длительным (например, атмосферная засуха, продолжительное нахождение растений в условиях засоления и т. д.) либо резкое повышение напряженности неблагоприятных факторов осуществляется за сравнительно короткий промежуток времени (например, суховей, резкое понижение температуры и т. д.). Ответные реакции на хроническое действие фактора и на стрессовые условия различны.
Клетка, чтобы жить и нормально функционировать, должна четко реагировать на сигналы внешней среды. Способность организмов реагировать соответствующим образом на внешние раздражители, на сигналы извне, следует рассматривать как необходимое условие приспособления клеток к окружающей среде. Для восприятия внешних сигналов клетка обладает набором необходимых рецепторов, в большинстве случаев вмонтированных в плазматическую мембрану или находящихся в протоплазме. Сигналы, которые имеют физическую, химическую и биологическую природу, клетки воспринимают со стороны внешней среды или от соседних клеток и преобразуют их в различные внутриклеточные биохимические процессы. Способность клеточных структур воспринимать определенные сигналы и их объемы и реагировать на них в значительной степени зависит от компетенции клетки.
Компетентность клетки - способность ее реагировать определенным образом на внешний индуктор - определяется наличием рецепторных молекул и их соответствием факторам среды. Кроме того, компетентная клетка обладает определенным потенциалом ответа на различные внешние воздействия. Компетентность устойчивых клеток определяется соответствием их внутренней структуры и комбинации внешних условий. При изменении напряженности факторов среды происходят изменения структурной организации и метаболических процессов в клетке с определенной скоростью и направленностью, соответствующие данным условиям.
В многоклеточном организме клетки разных типов в разные моменты времени достигают состояния компетенции для ответа на определенные факторы среды. После того как клетка становится компетентной и отвечает на определенный стимул, она изменяет свое состояние и начинает проявлять новую компетенцию (либо воспринимает другие сигналы, либо те же сигналы, но в другом объеме). Временные механизмы компетенции основываются на колебательном поведении регуляторных систем и на пластичности внутриклеточных обменов. Следовательно, компетентность клетки определяется количеством, локализацией, структурой рецепторов и потенциалом ответа на индуцирующее воздействие. Рецепторами называют специфические структуры клетки белковой или небелковой природы (лектины, фоторецепторы, хеморецепторы, механорецепторы, гормональные рецепторы).
Мембрана с помощью своих рецепторов "анализирует" и "качественно оценивает" химические и физические факторы среды и перекодирует сигналы внешней среды на язык, понятный внутриклеточным процессам Связывание раздражителя с рецептором сопровождается конформационными изменениями рецепторных молекул, которые передают сигнал следующей инстанции языком конформационных перестроек. Последующие превращения сигналов зависят от природы клеток и от свойств раздражителя.
Стандартной реакцией мембран на внешние раздражители является деполяризация - потеря заряда или изменение знака заряда, в результате чего возникает потенциал действия и изменяются свойства мембранных компонентов. Высокоамплитудный потенциал действия может быть вызван действием температуры, света, электрической стимуляции и некоторыми химическими соединениями [17; 38]. В отсутствие раздражителей растительная клетка имеет отрицательный потенциал покоя (от - 50 до - 200 мВ), протоплазма заряжена отрицательно по отношению к наружной поверхности. Причина этого - неравномерное распределение ионов: внутри клетки находится больше, чем снаружи, ионов Cl- и K+ но меньше Ca2+. Неравномерное распределение ионов, проявляющееся в форме мембранного потенциала, обусловлено, по- видимому, действием мембранных ионных насосов (переносчиков), ионных каналов и различной подвижностью ионов в мембране. В ответ на продолжительное раздражение происходит деполяризация мембраны, а затем ее постепенная перезарядка. Возникает потенциал противоположного знака, потенциал действия, который может на время полностью компенсировать потенциал покоя или обусловить появление потенциала с обратным знаком. Потенциал действия вначале развивается с выходом Cl- из клетки и поступлением Ca2+ в клетку. Затем начинается более медленный процесс - выход из клетки ионов K+, в результате снимается потенциал действия и восстанавливается потенциал покоя, сначала с иным распределением ионов, чем до раздражения. Затем восстанавливается исходное распределение ионов при участии переносчиков (K+ и Cl- поступают внутрь клетки, а Ca2+ наружу). Независимо от природы раздражения потенциал действия имеет двухфазный характер. Однако под влиянием различных агентов могут изменяться такие параметры потенциала действия, как амплитуда, длина волны, время наступления ответной реакции. Установлено, что все растения способны при определенных условиях генерировать потенциал действия. Латентный период потенциала действия у растений - от долей до сотен секунд, а его величина может достигать 100 - 150 мВ. В многоядерной водоросли Nitella высокоамплитудный потенциал действия может быть вызван действием температуры, света и т. д. У насекомоядного растения (росянки) и мимозы механический стимул, воспринимаемый специализированными чувствительными волосками, приводит к изменению тургорного давления в клетках, и в результате в одном случае захлопывается ловушка, а в другом - спадаются листья. Потенциал действия, генерируемый в эффекторных клетках, по своим параметрам подобен наблюдаемому в нервно-мышечных системах. Распространяющиеся потенциалы действия растений и животных имеют много общего, однако у растений они протекают медленнее. Скорость распространения потенциала действия у мимозы 4 см/с, у большинства растений 0,08 - 0,5 см/с.
Электрический потенциал, по-видимому, участвует в транспорте сигналов внешней среды и запуске внутриклеточных процессов. Например, резкие изменения условий существования в зоне корней индуцируют одиночный импульс, который, достигая листьев, вызывает в них усиление газообмена и ускорение транспорта ассимилятов по проводящим пучкам. При сильном раздражении верхушек побега (0,5 M KCl, холодная вода и др.) одиночный импульс ускоряет поглощение корнями калия и фосфора. Эти данные указывают на существование у растений быстрой электрической связи между отдельными клетками и тканями.
В настоящее время делаются попытки выявить молекулярные основы восприятия сигналов и всю сопряженную последовательность событий, связанных с усилением и трансформацией данных сигналов через систему посредников.
Известно, что кальций повышает устойчивость растений к различным стрессам (высокие и низкие температуры, анаэробиоз, понижение pH, заболевание). Д. Марме с сотрудниками [64] проведены обстоятельные исследования возможного функционирования кальция как вторичного посредника в растительных клетках. Они показали, что характер распределения Ca2+ в клетках колеотилей кукурузы зависит от света: при освещении увеличивалась концентрация свободного кальция в цитозоле клетки, что сопровождалось повышением активности НАД-киназы.
Очевидно, кальций как вторичный посредник воспринимает информацию первичного сигнала (света) и регулирует таким путем биохимические процессы (в частности, активность НАД-киназы).
Концентрация свободного Ca2+ в цитоплазме растительных и животных клеток низка (10-8 - 10-6M). Во внутриклеточных структурах (митохондриях, эндоплазматическом ретикулуме) концентрация свободных ионов Ca2+ превышает 10-3M. В животных клетках такая разница в концентрации кальция поддерживается мембранными Ca2+-АТФазами, системой Na+/Ca2+ обмена и, возможно, Ca2+-транспортирующей системой митохондрий. В растительных клетках при возникновении потенциала действия или при деполяризации мембран Ca2+ поступает в клетку извне и (или) освобождается из внутриклеточных резервуаров (цистерн ЭПР, митохондрий, вакуолей). В работах ряда исследователей показано, что Ca2+-АТФаза, локализованная в плазматических мембранах, осуществляет обмен Ca2+ на протоны (Ca2+/H+ -антипорт). В плазмалемме клеток имеются потенциалзависимые кальциевые каналы, которые открываются при деполяризации мембраны. В ЭПР также имеются кальциевые каналы, сходные с каналами плазматических мембран, причем движение Ca2+ в них направлено из цистерн ЭПР в цитозоль. Кроме того, в мембранах ЭПР растительных клеток обнаружена Ca2+-АТФаза, транспортирующая кальций из цитозоли во внутриклеточное депо (цистерны ЭПР). Концентрация свободного кальция в строме хлоропластов низкая, но она увеличивается при освещении. Значительная часть Ca2+ содержится в клеточных стенках растений (в форме нектатов, карбонатов, сульфатов) и в вакуолях (в форме оксалата).
Изменение в концентрации Ca2+ в цитозоле клеток играет существенную роль в механизмах движения протоплазмы, делении клеток, секреторной активности некоторых растительных тканей.
Таким образом, кальций, поступая из внешней среды или освобождаясь из внутриклеточных комиартментов, выступает в роли внутриклеточного медиатора, индуцирующего ряд физиологических процессов.
Кальций растительных клеток может связываться с кальмодулином и другими Ca2+ - связывающими белками. Кальмодулин - низкомолекулярный белок (Мм 16700) с высоким содержанием кислых аминокислот. Он имеет четыре участка, обладающих высоким сродством к Ca2+. Кальмодулин обнаружен в митохондриях, хлоропластах, микросомах и клеточных стенках. В цитозольной фракции имеется значительное количество этого белка (90%). Активируясь Ca2+ (10-6 M), кальмодулин регулирует активность Ca2+-АТФазы, НАД-киназы, НАД-оксидоредуктазы, протеинкиназ, липаз.
Многие реакции, индуцируемые фотохромом дальним красным (Ф730) также контролируются ионами кальция. C. Po (Ronx) предполагает следующую последовательность событий после поглощения клетками растений квантов красного света: образование Ф730 из Ф660→возрастание концентрации ионов кальция в цитоплазме клеток→связывание ионов кальция кальмодулином и непосредственное действие повышенных концентраций Ca2+ на функции клеток→связывание активированного кальмодулина с зависимыми от него ферментами и активация данных белков [26].
Следовательно, растительные клетки располагают механизмами для поддержания определенного уровня свободных ионов кальция в цитозоле и функционирования Ca2+ в качестве вторичного посредника в регуляции метаболизма.
В качестве другой сигнальной системы рассматривается ц-АМФ. У животных организмов циклические нуклетиды (ц-АМФ, ц-ГМФ) играют весьма существенную роль в системе внутриклеточной регуляции. Аденилатциклазная ферментная система отвечает за синтез сравнительно простого нуклеотида - циклического аденозинмонофосфата (ц-АМФ), способного активировать многие внутриклеточные ферменты. По своему строению ц-АМФ близок к АТФ. Он образуется из АТФ путем отделения двух фосфатных групп и последующего замыкания оставшейся фосфатной группировки в кольцо (отсюда и название - циклическая АМФ). Эта реакция катализируется аденилатциклазой, которая расположена на внутренней поверхности мембран и работает в присутствии фосфолипидов и ионов магния.
Действие экзогенных факторов может проявляться через циклические нулеотиды. В частности, Г. Мор с сотрудниками показал, что активация фитохрома красным светом сопровождается повышением в этиолированных проростках белой горчицы уровня ц-АМФ в два раза [26]. Воздействие факторов среды направлено при этом на мембрану. Начинает функционировать аденилатциклазная система (рис. 1), синтезируются циклические нуклеотиды, которые изменяют структурно-функциональное состояние хроматина, матричную активность ДНК, интенсивность новообразования белков-ферментов. В 1971 г. Т. Ленген (T. Langan) показал возможную связь ц-АМФ с регуляцией активности генома. Было показано, что ц-АМФ стимулирует фосфорилирование гистонов препаратами гистон-киназ, что приводит к активации синтеза РНК на матрице ДНК. Кроме того, ц-АМФ действует как аллостерический эффектор по отношению к протеинкиназам, которые катализируют такие реакции модификации, как фосфорилирование ядерных, цитоплазматических и мембранно-связанных белков [26]. В настоящее время выделен и очищен белок, проявляющий сродство как к ц-АМФ, так и к цитокининам. В связи с этим полагают, что существует определенная связь между циклическими нуклеотидами и фитогормонами.
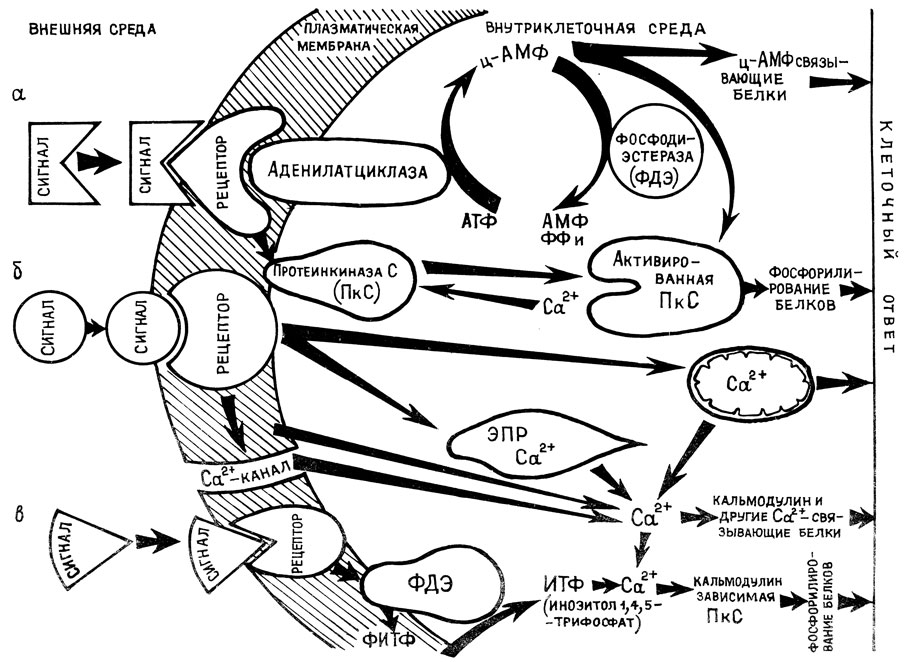
Рис. 1. Некоторые возможности пути передачи сигналов внешней среды и запуска метаболических процессов. Регуляция метаболических процессов в клетках может осуществляться за счет изменения: а - содержания ц-АМФ и активности протеинкиназного белка; б - внутриклеточной концентрации ионов кальция; в - уровня инозитолов в клетках
Таким образом, ц-АМФ является, по-видимому, "вторичным мессанджером" в цепи событий от рецепции сигналов внешней среды до изменения активности гормонального, ферментативного и генетического аппарата клетки. Связь фитохрома с синтезом ц-АМФ объясняет многостороннее влияние этого пигмента на разные звенья метаболизма, включая синтез РНК и белка.
Японские исследователи показали, что клетки культуры моркови синтезируют фитоалексины в ответ на грибную инфекцию [59]. Как полагают авторы, этот ответ опосредован другой сигнальной системой - фосфатидилинозитольной, включающей кальмодулинзависимые процессы. В растительных клетках установлено наличие системы фосфорилированных инозиголов. Инозитол-1,4,5-трифосфат (ИТФ) вызывает выход Ca2+ из внутриклеточных компартментов. ИТФ вместе с кальцием участвует в передаче сигналов извне внутрь клетки (рис. 1). Внешний сигнал связывается с рецептором, который через ряд промежуточных соединений активирует фосфодиэстеразу (ФДЭ). Данный фермент расщепляет фосфатидилинозитол-1,4,5-трифосфат (ФИТФ), в результате чего образуется инозитол-1,4,5-трифосфат и диацилглицерин. ИТФ растворим в воде, поэтому он диффундирует в цитоплазму и вызывает освобождение кальция из ЭПР, митохондрий и других компартментов. Высвобожденный Ca2+ активирует кальмодулинзависимую протеинкиназу, которая фосфорилирует внутриклеточные белки и вызывает изменение скорости и направленности метаболических процессов.
В общем виде сигнальная система клеток состоит из рецепторов, воспринимающих сигнал и функционально связанных с рецепторами вторичных посредников (Ca2+, кальмодулин, ц-АМФ, ИТФ, протеинкиназа). Эти внутриклеточные посредники служат для усиления и передачи воспринимаемого сигнала и запуска метаболических процессов.
Протеинкиназная активность обнаружена практически во всех клетках и тканях животных организмов. Ферменты, подобные по ряду свойств протеинкиназам. С животных организмов, обнаружены в клетках пшеницы и тыквы и в проростках щирицы. В последние годы в литературе появились сведения о наличии в растительных клетках Ca2+, фосфолипидзависимых протеинкиназ. Они обнаружены во фракции плазматических мембран клеток корней гороха и в цитозольной фракции, полученной из гипокотелей и стеблей тыквы [56; 68]. Протеинкиназы - ферменты, осуществляющие фосфорилирование белков по строго определенным группам серина, треонина и тирозина. Присоединение фосфата приводит к изменению структуры белковой молекулы и ее функциональной активности. Фосфорилированию подлежат структурные, транспортные и регуляторные белки. Протеинкиназа активируется кальцием (10-6-3.10-7M), фосфолипидами (фосфатидилсерин) и диацилглицерином (табл. 1).

Таблица 1. Протеинкиназа C-подобные белки растений
Регуляция протеинкиназной активности может быть различной в зависимости от качества воспринимаемого сигнала и функциональных особенностей тканей. Она может зависеть или не зависеть от циклических нуклеотидов, быть чувствительной или нечувствительной к кальмодулину и кальцию. Активированная протеинкиназа переносит фосфатную группу с АТФ на белки, которые в свою очередь активируют другие ферменты. Биологический смысл этого каскада активации ферментов состоит в том, что он подобно каскадным усилителям, применяемым в радиотехнике, многократно усиливает первоначальный сигнал, который индуцирует целый комплекс защитно-приспособительных реакций. В результате включается синтез адаптивных белков (например, стрессовых), протекторных соединений (пролин, полиамины, олиго- и полисахариды и др.), обнаруживаются изменения на уровне мембранных структур (меняется их липидный и белковый комплекс), возникают защитные системы на структурно-метаболическом уровне, а затем следуют морфоструктурные изменения.
Например, чтобы свет оказал свое физиологическое действие на растение, он должен быть поглощен рецептором (фитохромом или другими пигментами). Одной из реакций, находящейся под контролем фитохрома, является свертывание листьев мимозы с наступлением темноты. Весь процесс завершается через 5 мин - это время слишком мало, чтобы мог осуществляться контроль на уровне транскрипции. Данный факт, а также то обстоятельство, что какое-то количество фитохрома оказывается прочно связанным с мембранами, привели к предположению, что первичное действие фитохрома сводится к изменению свойств мембраны. Молекула пигмента, поглотившего квант света, переходит в возбужденное состояние, взаимодействует с мембраной клетки и вызывает изменение ее конформации. Изменение состояния мембраны в одном месте может распространиться и на другие ее участки. В результате изменится проницаемость мембраны, ее заряд, активность связанных с ней ферментов. Все это, в свою очередь, может быть причиной изменения путей общего метаболизма клетки. Медленные реакции в ответ на изменение состояния фитохрома могут быть связаны с процессом транскрипции генов. Пигменты, участвующие в фоторегуляции морфогенеза растений, оказывают при возбуждении их светом непосредственное действие на генный аппарат растений, превращая потенциально активные гены в активные и тем самым способствуя образованию новых информационных РНК и биосинтезу до того времени "запрещенных" белков.
Рецепторный аппарат клетки представляет собой динамическую и, невидимому, высокоселективную систему, обеспечивающую как связь клеток с внешней средой, так и регуляцию их функциональной активности. Специфичность рецепторных систем в соответствии с клеточной специализацией определяет возможность осуществления характерного для данного тина клеток ответа на действие различных факторов внешней среды.
Действие любого неблагоприятного экстремального фактора вызывает ряд ответных защитно-приспособительных реакций. Характер ответных реакций в значительной степени зависит от интенсивности действующего фактора. При малой его интенсивности наблюдается нормальная ответная реакция (т. е. усиление или ослабление внутриклеточных физиологических процессов). При значительной интенсивности действующего фактора организм начинает защищаться от неблагоприятных воздействий и для этого мобилизует все имеющиеся у него потенции. При этом в организме могут возникнуть и новые свойства, отсутствующие до действия данного фактора.
Еще в 1900 г. индийский физик и физиолог растений Джегдиш Чандра Бос пришел к выводу об общности ответных реакций у животных и растений. Представления об однотипности ответных реакций организмов на окружающие условия получили развитие в работах Д. Н. Насонова и В. Я. Александрова. Было постулировано, что реакция протоплазмы клетки на окружающие условия монотонна. Она выражается в том, что в ответ на воздействия в протоплазме клеток растений и животных происходят всегда одни и те же изменения в следующей последовательности: 1) уменьшается степень дисперсности протоплазмы; 2) повышается проницаемость протоплазмы; 3) денатурируют белки; 4) происходят паранекротические изменения ядра; 5) коагулирует протоплазма [37].
Эти однотипные, монотонные, появляющиеся при любом повреждении изменения могут полностью исчезнуть после устранения альтерирующего агента, если действие его не зашло слишком далеко. Неспецифичность этих признаков выражается в том, что они сопутствуют разным способам повреждения и наблюдаются у любых тканевых клеток и одноклеточных организмов. Этот комплекс неспецифических физико-химических признаков повреждения был назван паранекротическим, а состояние клеток, при котором у них развивается комплекс паранекротических изменений, - паранекрозом (паранекроз - "вблизи" или "около" смерти). Смысл этого названия заключается в том, что реакции, возникающие в клетке при раздражении и при повреждении, сходны. В дальнейшем идеи Д. Н. Насонова и В. Я. Александрова были развиты в трудах канадского физиолога Г. Селье [41]. Он ввел понятие стресса в область медицины, но оно стало широко использоваться и в физиологии растений. Г. Селье дает следующее определение этому понятию: "Стресс есть неспецифический ответ организма на любое предъявляемое ему требование". Стресс в понимании фитофизиологов есть некоторое нарушение, вызываемое неблагоприятными условиями [42].
Изменение проницаемости клеточных мембран, по-видимому, является первичным звеном в ответных реакциях. Проницаемость - способность клеток и тканей поглощать или обмениваться веществами с окружающей средой. Проницаемость мембран может меняться как под действием внутренних условий (в процессах прорастания семян, роста растений и старения клеток и тканей), так и под воздействием различных факторов среды (фитопатогенов, температурных и световых условий, анаэробиоза, избытка тяжелых металлов и т. д.). Значительные изменения проницаемости мембран растительных клеток обнаруживаются при действии абиотических факторов среды. У 4 - 5-дневных проростков пшеницы, фасоли и хлопчатника, погруженных в растворы солей хлористого, сернокислого и углекислого натрия, отмечается значительное увеличение проницаемости мембран корней и происходит усиленное выделение в наружный раствор аминокислот, органических кислот и неорганических ионов. Проницаемость растительных тканей резко меняется и при повышенной температуре среды (45°С). В литературе имеются многочисленные данные, прямо или косвенно свидетельствующие о наличии определенной связи между проницаемостью мембран растительных клеток и морозо- и холодоустойчивостью растений. По данным П. Нобела, проницаемость мембран хлоропластов неустойчивых к холоду растений (томаты, фасоль) при пониженных температурах резко возрастала, в то время как устойчивых (горох, шпинат) - не изменялась [65]. Вышеизложенное позволяет считать, что изменение проницаемости клеточных мембран является общим, первичным звеном неспецифических механизмов ответной реакции растительного организма на внешние воздействия. В настоящее время доказано, что проницаемость растительных тканей может быть использована как показатель устойчивости растений к неблагоприятным условиям внешней среды.
Возникает вопрос: определяется ли устойчивость растений к засухе, морозу, засолению одним общим механизмом или эти механизмы специфичны в каждом случае? На любое воздействие растительный организм отвечает целым комплексом защитно-приспособительных реакций, состоящих как из общих (неспецифических), так и специфических процессов. В работе Б. П. Строгонова показано, что процесс приспособления растений к сульфатному и хлоридному засолению идет неодинаково. Например, на сульфатном засолении транспирация у растений возрастает, а на хлоридном - снижается [44].
Некоторые исследователи считают, что в основе устойчивости к различным экстремальным факторам лежат неспецифические (однотипные) реакции (В. Я. Александров, Г. В. Удовенко). В. Я. Александров трактует свой большой материал по влиянию температуры на животные и растительные организмы с позиций неспецифической реакции организмов на действие повышенных температур [1; 2]. Другие связывают устойчивость с реакциями специфического характера (Н. А. Максимов, П. А. Генкель)). П. А. Генкель считает, что ответная реакция растения на неблагоприятные условия имеет сложный характер. При адаптационном процессе развертываются защитно-приспособительные реакции как неспецифического, так и специфического характера [12].
Ю. А. Урманцев следующим образом трактует вопрос о специфичности и неспецифичности ответных реакций растений. "Ответные реакции растений на действие разных неблагоприятных условий, по крайней мере в ряде случаев, могут предстать в виде специфических реализаций одной и той же закономерности". В частности, кривые, описывающие зависимость тех или иных функций растения от действия того или иного неблагоприятного фактора, как правило, имеют одну и ту же форму "колокола". Однако при анализе этих кривых отмечают, что эти формы достоверно отличаются своими амплитудами, высотами. Если исходить из концепции о единой устойчивости растений, то для всех функций растения и всех неблагоприятных условий исследователи получали бы один и тот же "колокол" с одинаковыми параметрами (амплитудами, высотами). По-видимому, специфичность ответных реакций проявляется как составная часть общих, однотипных защитно-приспособительных реакций. Специфичность ответных реакций - особенность проявления общего [47].
Концепция о том, что ответные реакции растений на неблагоприятные окружающие условия протекают однотипно, сложилась в основном при изучении повреждений и гибели растений. Мнение, что ответная реакция сложнее и состоит как из неспецифических, так и из специфических реакций, возникло при изучении адаптивных изменений, где специфические ответы растения выступают на первый план. При определенной (небольшой) дозе воздействия неблагоприятного фактора, когда возможны адаптивные изменения, наблюдаются наряду с неснецифическими и специфические реакции. При усилении меры воздействия (фактор×время) организм начинает защищаться от неблагоприятного воздействия и мобилизует все имеющиеся у него средства. В последнем случае мы можем не обнаружить специфики в ответных реакциях. И. Н. Андреева и Г. М. Гринева изучали действие повышенной температуры и анаэробиоза на субмикроскопическую структуру митохондрий. Субмикроскопические картины, наблюдавшиеся в результате воздействия этих факторов, резко отличались друг от друга. При действии высокой температуры (45°С) на корни кукурузы происходит набухание митохондрий, просветление матрикса, везикуляция и уменьшение числа крист. При действии анаэробиоза обнаруживаются лентовидные и закрученные кристы, увеличивается их объем, они уплотняются, наблюдается их везикуляция и увеличение числа. При увеличении меры воздействия (в конце воздействия) морфологические картины повреждений сближаются: наблюдается высокая степень набухания митохондрий, полное отсутствие матрикса, сохраняется небольшое число крист-везикул. При действии обоих факторов митохондрии в конце концов разрушались. Фосфорилирующая активность митохондрий сохранялась при малых дозах воздействия температуры и анаэробиоза, а при сильных повреждениях наблюдалось полное разобщение окисления и фосфорилирования [4].
Соотношение специфических и неспецифических ответных реакций в значительной степени зависит от длительности действующего фактора. При кратковременном действии фактора в высокой дозе наблюдаются в основном неспецифические ответные реакции. Например, мы сходным жестом отдергиваем руку, прикоснувшись к горячему, холодному, колкому предметам. При длительном воздействии стрессового фактора срабатывает большее число метаболических звеньев, некоторые из которых обладают чертами специфичности для данного организма. Постепенное, пролонгированное действие стрессора приводит к включению процессов специализированной адаптации, которые обеспечивают систему надежности функционирования внутриклеточных процессов в экстремальных условиях.
Характер специфической реакции на стрессовые воздействия указывает на природу повреждающего фактора, а при неспецифической - природу действующего сигналу угадать трудно. Неспецифические реакции наблюдаются чаще, чем специфические. Пример специфической реакции - признаки острой недостаточности (или избыточности) элементов питания растений.
Важно отметить, что ответные реакции растений на разные факторы носят колебательный характер. Так, данные, полученные П. С. Беликовым, показывают, что при действии высокой температуры вязкость цитоплазмы вначале уменьшается, а затем увеличивается. Скорость движения цитоплазмы и выход веществ из клетки также меняются волнообразно: вначале наблюдается усиление этих процессов, затем скорость их замедляется [5]. В зависимости от силы повреждающего воздействия меняется характер этих колебаний: амплитуда, длина волны, время наступления пусковой ответной реакции. По мнению В. Я. Александрова, колебательный характер физиологических процессов в клетках при действии раздражителей отражает сложную природу ответных реакций, которые имеют разную направленность. Одни из этих реакций имеют деструктивный характер, другие направлены на сохранение внутриклеточных структур и процессов.
Можно предположить, что специфическое реагирование на действие экстремальных факторов контролируется генетическими механизмами через работу белоксинтезирующего аппарата. Неспецифическое реагирование, по-видимому, не связано с генетическим контролем и в его основе лежит физиологическая пластичность организма (пластичность мембранных компонентов, изменение структуры и активности внутриклеточных белков и пр.). Соотношение специфичности и неспецифичности в устойчивости может варьировать в зависимости от биологических особенностей объекта. В качестве примера рассмотрим два биологических объекта. Огурец как вид сформировался в условиях тропиков; ареал его естественного распространения включает отдельные районы Центральной Азии, характеризующиеся незначительными колебаниями температуры и других факторов среды. При действии экстремальных температур (низких) для сохранения жизнеспособности данных растений в основном срабатывают специфические ответные реакции, которые определяются генетическими потенциями вида. Стабильные факторы в районах Центральной Азии не обеспечили формирования пластичности обмена у данного растительного организма.
В противоположность огурцу становление рода Triticum шло на фоне заметного колебания температуры среды и других факторов. Ареал распространения пшеницы включает огромные территории от Северного полярного круга до южных пределов Австралии, Америки и Африки. Пшеница хорошо приспособлена и к горным условиям и растет на высоте 4 тыс. м над уровнем моря. Можно предположить, что для пшеницы залогом широкого распространения является хорошо развитая система специфического реагирования, подкрепленная механизмами неспецифической устойчивости. Эволюция у пшеницы шла по типу выработки механизмов лабильности мембранных компонентов, пластичности регуляторных механизмов, подвижности структуры и функции внутриклеточных белков, что позволяет пшенице иметь широкий ареал распространения.
Во всех случаях невозможно провести резкую границу между специфическими и неспецифическими реакциями. Кажущаяся неспецифичность физиолого-биохимических и других признаков повреждения не абсолютна; здесь, по-видимому, следует скорее говорить о сходстве явлений, чем об их тождестве, так как на фоне однотипных реакций обычно удается подметить специфические особенности. По-видимому, в сочетании специфического и неспецифического характера ответных реакций заложена возможность реагирования живых систем и развитие их в эволюции.
При изучении процессов устойчивости иногда наблюдаются случаи одновременной устойчивости к двум или нескольким ее видам. П. А. Генкель, анализируя ряд подобных фактов, сформулировал понятие о сопряженной устойчивости, которая может быть положительной или отрицательной. Хорошим примером сопряженной устойчивости является повышение жароустойчивости и солеустойчивости у проса сорта Кремовый, который перед посевом подвергался обработке 1/40 M CaCl2. В этом случае проявляется положительная сопряженная устойчивость. Обработки CaCl2 вызывают повышение вязкости протоплазмы и снижение интенсивности обмена, что способствует большей жаро- и солеустойчивости растений. При меры положительной и отрицательной сопряженной устойчивости приводятся в работах А. Кашлана. Выращенный в вегетационных опытах табак подвергался предпосевному закаливанию против засухи. Было обнаружено повышение засухоустойчивости и одновременно сульфатоустойчивости у растений и понижение хлоридоустойчивости. Более подробный анализ показал, что улучшение роста и продуктивности на сульфатном засолении у закаленных к засухе растений связано не с повышением сульфатоустойчивости, а с их повышенной жароустойчивостью, так как контрольные незакаленные растения сильно снижают свою жароустойчивость на сульфатном засолении почвы. На хлоридном засолении снижение хлоридоустойчивости у закаленных к засухе растений связано с их повышенным метаболизмом, большим поглощением солей и более развитой корневой системой (больший объем и поверхность поглощения корней) [11, 12].
Неоднократно отмечаемое сходство в реакциях растений на неблагоприятные факторы среды, например на холод и тепло [48; 46], и наличие положительной сопряженной устойчивости позволили сформулировать вывод, что резистентность растений к различным экстремальным условиям может контролироваться одними и теми же эндогенными факторами [1; 60; 61]. Сходство ответных реакций можно объяснить существованием широкого круга неспецифических приспособительных реакций и тем, что специфическое реагирование на такие экзогенные воздействия, как холод и тепло, сопряжено с системой индуцированного синтеза белка, т. е. осуществляется по единому типу генетической регуляции физиологических процессов. Сходство ответных реакций растений на температурный, водный и солевой стресс определяется, невидимому, тем, что в этих условиях в клетках создается водный дефицит, который может быть устранен с помощью однотипных защитно-приспособительных процессов (усиления синтеза пролива и др.).
Кроме понятия сопряженной устойчивости П. А. Генкель ввел понятие конвергентной устойчивости. Конвергенция - это наблюдаемое сходство различных организмов, вызванное одинаковыми условиями существования - одинаковым давлением отбора. Существуют два типа конвергентной устойчивости: 1) типичная конвергенция, когда устойчивость различных организмов обусловлена одинаковыми условиями существования; 2) нетипичная, когда разные условия приводят к одинаковому результату. Примером нетипичной конвергенции является высокая жароустойчивость древесных пород зимой, связанная с их обезвоживанием и накоплением липидов на поверхности протопласта [12].
Кроме того, наблюдаются случаи расходящейся нетипичной конвергентной устойчивости, когда одинаковое воздействие приводит к неодинаковому результату.
Для всех организмов, находящихся на различных уровнях организации, можно вычленить некоторые сходные характерные черты в их реакции на внешние воздействия. К ним относятся: 1) способность отвечать на действие раздражителей включением сигнальных систем, которые рецептируют сигнал, усиливают его и запускают ответные физиолого-биохимические процессы; 2) способность сочетать в ответных реакциях признаки неспецифические, в значительной степени не зависящие от природы воздействующего фактора, с признаками специфическими, характерными для данного фактора. Источником специфических ответных реакций является гетерогенная расчлененность систем, источником неспецифичности - взаимосвязанность ее частей, кооперативность их взаимоотношения. Под влиянием раздражителей возникают повреждения, выражающиеся в нарушении структуры и функции клетки. Процессы возбуждения приводят к активации процессов жизнедеятельности клеток. В результате этого действие последующих раздражителей начинает восприниматься клеткой с меньшей силой, появляется закалка. На фоне закалки происходит восстановление - репарация исходных функций и структур.
У высших наземных растений прочный контакт со средой в условиях неподвижного образа жизни вызывает необходимость развития активных приспособительных реакций, совершенствования способов их адаптации к постоянно меняющейся, гетерогенной среде обитания. Исследование защитных реакций необходимо для решения вопросов, связанных с интродукцией, селекцией растений, а также для разработки методов искусственного повышения устойчивости клеток и организмов к биотическим и абиотическим факторам среды.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'