Быстрые движения листьев у чувствительного растения mimosa pudica
Листья у Mimosa pudica, как и у Samanea, совершают циркадные никтинастические движения. Но они могут также чрезвычайно быстро двигаться в ответ на механические, температурные, химические и электрические раздражители. Через несколько секунд после прикосновения к листу черешки опускаются и листочки складываются (рис. 13.1). Такая быстрая реакция, требующая высокой степени координации, обусловлена преобразованием механического стимула, воспринимаемого сенсорными клетками черешка, в электрический сигнал. Этот сигнал - вероятно, деполяризация мембраны - быстро распространяется по ткани, пока не дойдет до моторных клеток листовой подушечки, которые тотчас же изменяют свой объем, и это приводит к движению листьев или листочков.

Рис. 13.1. Листья Mimosa pudica (вид сверху) до тактильного раздражения (вверху слева) и после него (внизу справа)
Распространяющийся электрический сигнал можно измерить с помощью двух стеклянных электродов, заполненных раствором соли, один из которых помещают на черешок, а другой - в почву, окружающую корни, или на стенку горшка с растением. Сигнал представляет собой кратковременный сдвиг разности потенциалов между электродами (рис. 13.2). Он передается по черенку у мимозы со скоростью около 2 см/с и примерно в 5 раз быстрее по листу насекомоядного растения Dionaea (венерина мухоловка). Скорость передачи возрастает в 3 раза при повышении температуры на каждые 10°, что указывает на участие химических реакций. При удалении флоэмы распространение сигналов замедляется; значит, они передаются по клеткам флоэмы. Сигнал не может преодолеть барьер из мертвых клеток; это означает, что он, вероятно, не связан с диффузией водорастворимых гормонов.
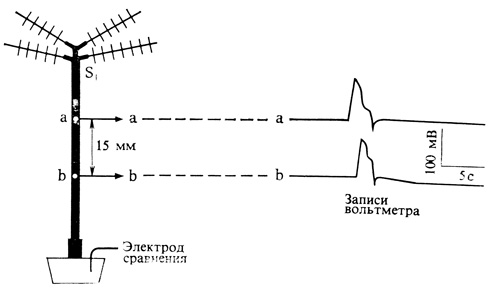
Рис. 13.2. Передача электрического сигнала, инициированного Холодовым раздражением в области S1 в черешке Mimosa pudica. Потенциалы действия (справа) одновременно регистрируются двумя электродами а и б, расположенными как показано на схеме слева. Электроды сравнения для а и б находятся в почве. (По Sibaoka. 196-8. Symp. Soc. Exp. Biol., 20, 49-73.)
Хотя наружные электроды и позволяют установить, что в растениях для связи между клетками используются электрические сигналы, они не дают никаких указаний относительно механизмов передачи или участвующих в ней клеток. С помощью внутриклеточных микроэлектродов (см. рис. 7.8) удалось показать, что определенные клетки во флоэме и протоксилеме мимозы являются "возбудимыми" (рис. 13.3). Их потенциал покоя, т. е. трансмембранный потенциал до возбуждения, значительно более отрицателен, чем у окружающих клеток. Когда в результате электрической или химической стимуляции этот потенциал (т. е. электроотрицательность внутренней стороны мембраны) снижается до определенного "порогового" уровня, клетка дает "разряд". Все стимулы, которые сильнее порогового, вызывают одинаковую реакцию ("всё или ничего"), состоящую в деполяризации до нуля или даже до положительного уровня с последующим быстрым возвратом к потенциалу покоя. Возбудимые клетки в растительной ткани всегда связаны плазмодесмами, по которым, видимо, и передаются электрические сигналы. Распространяющийся сигнал, называемый потенциалом действия, сходен с потенциалами действия в нервной ткани, с той лишь разницей, что в растениях не было найдено никакого химического передатчика. Ацетилхолин, который передает сигнал от одной клетки к другой в нервной ткани, есть и в растениях, но нет убедительных данных в пользу того, что он и здесь участвует в передаче межклеточных сигналов.
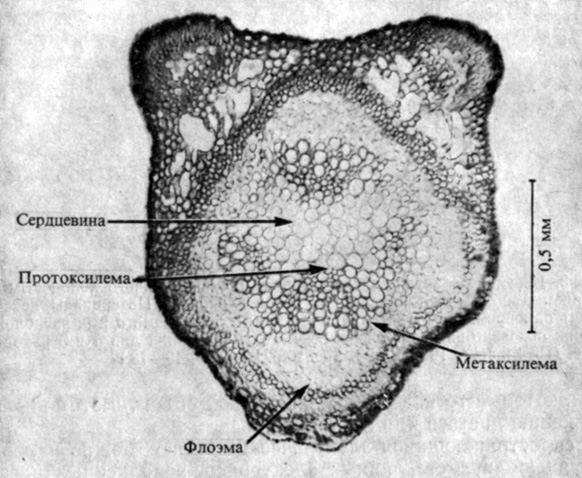
Рис. 13.3. Поперечный срез черешка Mimosa pudica. Электрические измерения показывают, что все клетки в протоксилеме 'возбудимы'. (Sibaoka. 1966. Symp. Soc. Exp. Biol., 20, 49-73.)
Раздражение черешка у мимозы легким прикосновением или каплей воды возбуждает потенциал действия, ограниченный пределами данного листа. Но если стимулировать лист ожогом или повреждением, вовлекаются и другие листья; такие стимулы вызывают не только потенциал действия, но и так называемый изменяющийся потенциал. Этот потенциал, по-видимому, передается химическим веществом, которое распространяется по ксилеме, проходит через листовую подушечку и инициирует на ее дальней стороне новый потенциал действия. Последний передается по возбудимым клеткам флоэмы и протоксилемы, пока не дойдет до другой листовой подушечки, в которой затем происходят характерные изменения тургора, ведущие к складыванию листа. В зависимости от силы первоначального раздражения изменяющийся (градуальный) потенциал может снова пройти через листовую подушечку и стимулирует еще один потенциал действия на ее дальней стороне. Таким образом, изменяющийся потенциал координирует движение различных листьев растения, возбуждая потенциалы действия, передающиеся по определенным зонам ткани.
По-видимому, у мимозы движение органов всецело определяется изменениями тургора. Уменьшению объема тех или иных моторных клеток сопутствует выделение из них ионов K+ и танниноподобных веществ во внеклеточное пространство. Такое изменение, обусловленное, как полагают, быстрым увеличением проницаемости мембран, сопровождается электрическими сигналами, которые можно выявить с помощью микроэлектродов. Внешнее сходство между этим процессом и сокращением мышц у животных позволяет предположить и наличие общих внутренних особенностей, таких, как участие сократительных белков типа актомиозина; однако пока нет веских данных, которые подтверждали бы гипотезу о подобном механизме или противоречили ей.
Некоторые исследователи подчеркивали значение таннинов для движений мимозы. Эта гетерогенная группа конденсированных фенольных соединений может образовывать комплексы как с белками, так и неорганическими ионами. Таннины денатурируют белки и могли бы повреждать клетку, но не вызывают вредных эффектов, пока они изолированы в мембранных мешочках вакуолей. Видимо, их особенно много в вакуолях растений, способных к тигмонастическим движениям (рис. 13.4). Во время таких движений танниновые агрегаты в вакуолях диссоциируют, и некоторые таннины, видимо, выделяются через мембраны моторных клеток. Затем они диффундируют в гидатоды (поры) в эпидермисе листовой подушечки. Поскольку таннины обладают вяжущим вкусом и действуют как природные репелленты, выделение таннинов при движении листа может способствовать защите растения от насекомых.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'