Насекомоядные растения
Многие насекомоядные растения встречаются в болотистых районах с низким содержанием азота в почве. Эти растения удовлетворяют часть своей потребности в азоте, улавливая насекомых листьями и переваривая затем добычу. Эта инверсия более обычных отношений между животными и растениями интриговала многих исследователей, в том числе Чарлза Дарвина. Проведенные им простые опыты показали, как благотворна для насекомоядных растений мясная диета (табл. 13.1). Дарвин исследовал по существу все аспекты поведения этих удивительных растений, начиная от их методов захвата добычи до химических особенностей процесса ее переваривания. Его остроумные опыты, тщательно собранные данные и блестящие умозаключения должны восхищать всех биологов. Некоторые из его книг указаны в библиографии в конце этой главы.
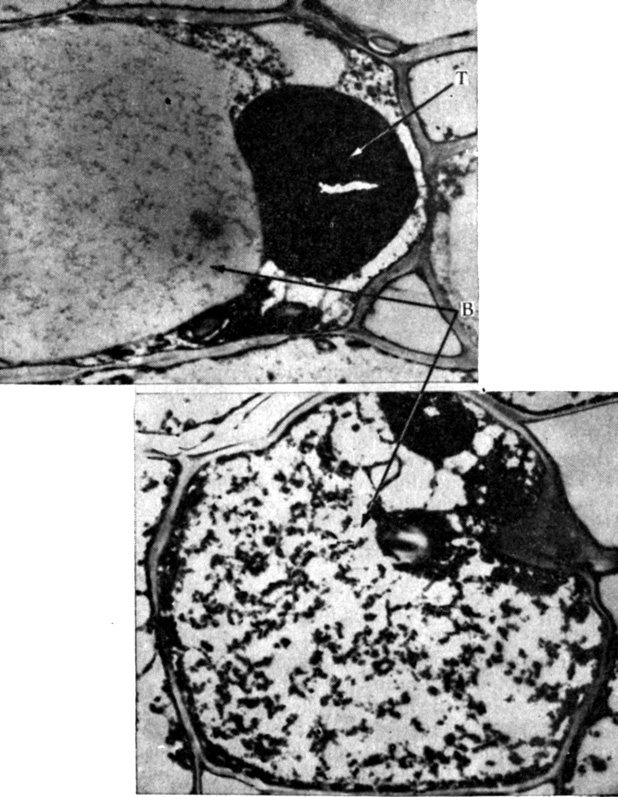
Рис. 13.4. Моторная клетка в первичной листовой подушечке Mimosa pudica до раздражения (вверху) и после раздражения (внизу). B - центральная вакуоль; T - таннины. Обратите внимание на то, что после раздражения крупная масса таннинов дробится на более мелкие единицы; следует также отметить изменение тонко диспергированного электроноплотного материала в вакуоли. X8000. Микрофотографии сделаны с клеточного материала, фиксированного OsO4. (Toriyama. 1971. Cytologia, 36, 359-395.)

Таблица 13.1. Различия между группой голодающих растений Drosera rotundifolia и группой растений, подкармливаемых жареным мясом (относительные показатели)
(Чарлз Дарвин собирал растения в поле и выращивал в сосудах, разделенных на две части. Растения в одной половине каждой культуры подкармливали жареным мясом, а другая половина служила контролем. Через три месяца после начала опыта экспериментальные растения превосходили контроль во всех отношениях, особенно по репродуктивным показателям. (Ch. Darvin, Insectivorous Plants, revised by F. Darvin, London, John Murray, 1908.))
Насекомоядные растения, обнаруженные среди грибов и в шести семействах покрытосеменных, могут использовать пассивный (наподобие липкой бумаги) или активный механизм ловли насекомых (рис. 13.5). Пассивные ловушки обычно чашеобразные и глубокие. Насекомому, которое влетело или заползло в одну из них, трудно выбраться так быстро, чтобы избежать повреждения кислым секретом и не быть убитым содержащимися в нем пищеварительными ферментами. В таких ловушках поверхность листа покрыта слизистым веществом, обездвиживающим насекомое. Такие вещества, главным образом углеводы, синтезируются железистыми клетками на поверхности листа и выводятся из клетки секреторными пузырьками диктиосом. Некоторые насекомоядные растения, например росянка (Drosera), используют как "липкую бумагу", так и активный механизм ловли: насекомые, попавшие в клейкий секрет, бьют ножками и крыльями, раздражая таким образом сенсорные клетки находящихся поблизости волосков. Эти волоски охватывают насекомое и приводят его в контакт с железистыми клетками, выделяющими сильные кислоты и гидролитические ферменты. Движения ловушки сравнительно медленны у растений с комбинированным пассивно-активным механизмом ловли насекомых и значительно быстрее у таких растений, как венерина мухоловка (Dionaea), которая при ловле насекомых полагается исключительно на движения ловушки.
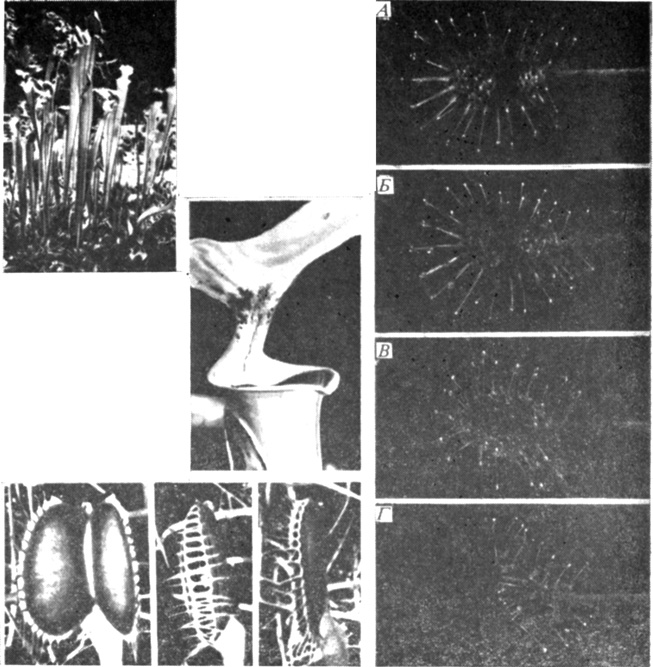
Рис. 13.5. Ловушки у насекомоядных растений. Слева вверху: Sarracenia flava - растение с кувшинчиком - ловит насекомых в свои глубокие чаши. Ниже отдельный кувшинчик представлен в увеличенном виде. (Фото предоставлено J. Mazrimas, co-editor Carnivorous Plant Newsletter.) Справа: поведение листа росянки (Drosera) во время поимки насекомого. а (через час после появления добычи) - насекомое борется с чувствительными волосками, расположенными внутри ловушки в 3 ряда; б (спустя 1,25 ч) - раздражаемые чувствительные волоски двигаются и отталкивают насекомое от края листа; в и г - то же спустя 3 и 4 ч. Чувствительные волоски, расположенные у края и не подвергавшиеся механической стимуляции, двигаются, и листовая пластинка загибается вокруг насекомого. Слева внизу: ловушка венериной мухоловки (Dionaea). Слева направо: до раздражения, через 10 с после того, как два раздражения были восприняты чувствительными волосками, и суженая ловушка через день после поимки насекомого. (Снимки Drosera и Dionaea из Williams. 1976. Proc. Amer. Phil. Soc., 120, 187-204.)
Ловушка у Dionaea состоит из листа, образующего две лопасти с рядом волосков по краю и тремя триггерными волосками, расположенными на верхней поверхности в виде треугольника (рис. 13.6). Механическое раздражение одного из триггерных волосков вызывает один-единственный потенциал действия, который распространяется по клеткам волоска и лопасти, возбуждая в результате клетки лопасти, изменение объема которых вызывает ее движение. Для закрывания ловушки необходимы два последовательных потенциала действия или даже больше, если интервал между двумя потенциалами больше 15 с. Это условие (подобно необходимости ряда последовательных дней для фотопериодической индукции) предполагает наличие примитивного типа "памяти". Растение еще "помнит" первый потенциал действия, если второй следует за первым достаточно быстро, но этот "след памяти" вскоре стирается. По-видимому, один потенциал действия приводит к увеличению количества какого-то вещества, контролирующего движение. Уровень этого вещества, которое со временем должно разрушаться, после одного раздражения все еще остается подпороговым. Но если второй потенциал действия придет раньше, чем этот уровень существенно снизится, концентрация вещества превысит порог возбуждения и произойдет реакция.
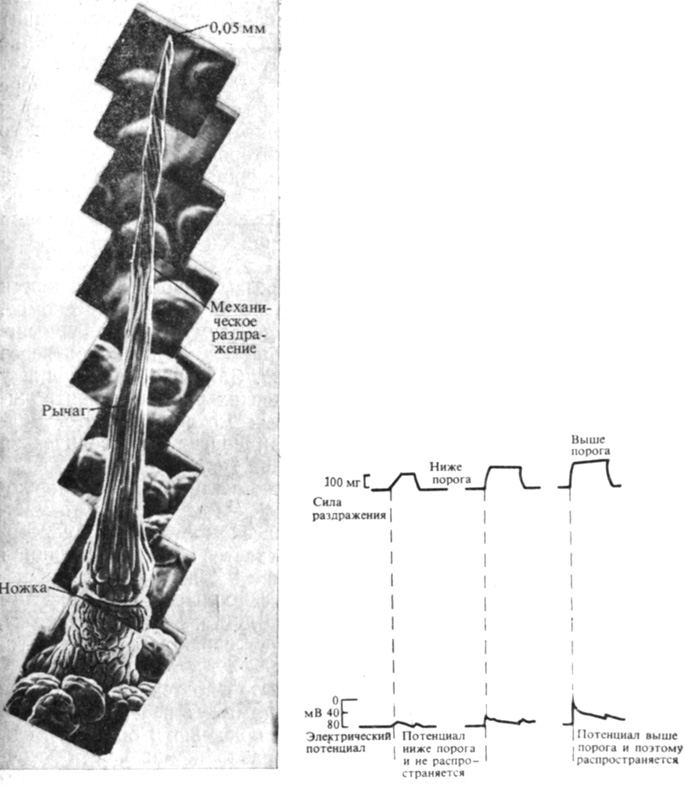
Рис. 13.6. Слева: микрофотография чувствительного волоска Dionaea, полученная с помощью сканирующего электронного микроскопа. Материал не обработан, если не считать напыления золотом слоем 10-50 нм. (Mozingo et al. 1970. Amer. J. Bot., 57, 593-598.) Справа: механическое раздражение чувствительного волоска инициирует распространяющийся потенциал действия, если оно сильнее определенного порогового уровня
Движение ловушки происходит чрезвычайно быстро, как и должно быть для того, чтобы можно было поймать насекомое. Весь процесс, включая преобразование стимула, передачу сигнала и закрывание ловушки, завершается меньше чем через 0,1 с после достаточного раздражения. Закрывание ловушки у Dionaea, так же как и у никтинастических растений, контролируется изменениями объема ключевых моторных клеток; однако не ясно, играют ли K+, Cl- или другие неорганические ионы значительную роль в регуляции их тургора.
Двукратное раздражение не ведет к полному закрытию ловушки. Поэтому очень мелкие насекомые могут выползти между волосками и спастись. Но если насекомое задевает сенсорный волосок, то каждый раз возбуждается новый потенциал действия, ловушка закрывается плотнее и стимулируется секреция кислот и пищеварительных ферментов железистыми волосками на верхней поверхности листа. Таким образом, сопротивляющееся насекомое ускоряет свою собственную гибель!

Рис. 13.7. Горох (Pisum sativum) поддерживает свой стебель с помощью усиков (слева), a Parthenocissus tricuspidata образует подушечки, приклеивающиеся к опорному субстрату (справа). (Второе фото из Reinbold et al. 1972. In: D. J. Carr, ed., Plant Growth Substances, Springer-Verlag, New York.)
Ловушка у Dionaea остается закрытой до тех пор, пока все тело насекомого, кроме его скелета, не будет переварено; если насекомое крупное, это может длиться неделю и больше. Если же ловушка закрылась в результате раздражения каким-то непищевым объектом (не насекомым) или кратковременного механического воздействия, она может опять открыться уже через несколько часов. Таким образом, как заключил Дарвин, сами продукты переваривания держат ловушку закрытой. Когда пища переварится и ловушка снова откроется, листья опять окажутся в горизонтальном положении, оптимальном для фотосинтеза. Таким образом, в отсутствие мясной пищи растение возвращается к автотрофному, фотосинтетическому питанию.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'