Спектрофотометрическое определение фитохрома
Для определения содержания фитохрома в растительной ткани можно использовать двухволновой спектрофотометр.
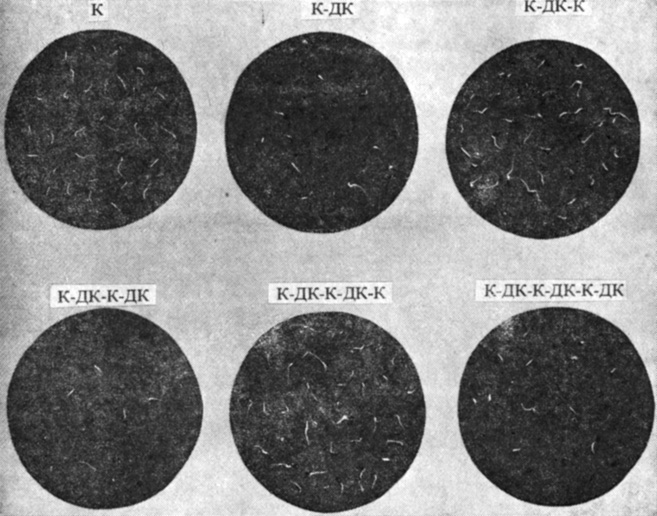
Рис. 11.7. Обратимость фотореакции при прорастании семян салата. Каждую партию семян после выдерживания в темноте облучали в указанной последовательности и затем снова помещали в темноту на 2 дня. К - красный свет; ДК - дальний красный свет
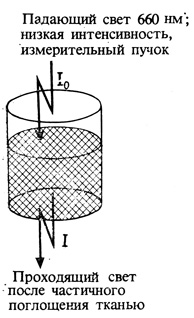
1. Кювету наполняют кусочками растительной ткани (или экстрактом из них) и облучают красным светом (660 нм) высокой интенсивности. Примерно минутного актиничного облучения достаточно для предельной степени превращения Фк в Фдк.
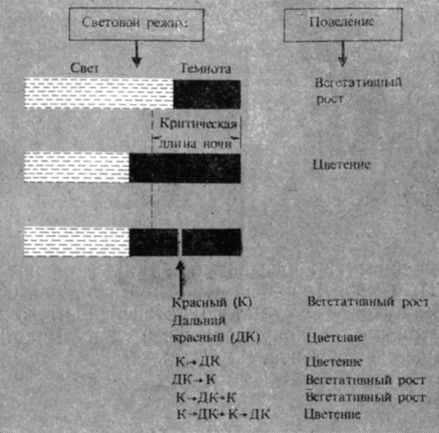
Рис. 11.8. Обратимое регулирование цветения короткодневного растения красным и дальним красным светом
2. Поглощение (П), называемое также оптической плотностью (ОП), измеряют попеременно при 660, 730, 660 нм и т. д. Измеряющий пучок имеет малую интенсивность, а каждое облучение настолько кратковременно, что не вызывает значительного превращения пигмента.
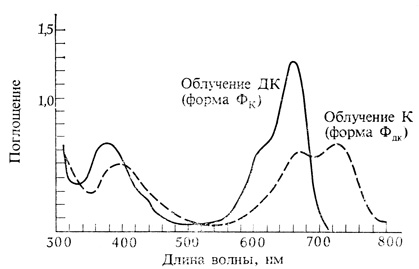
Рис. 11.9. Спектр поглощения растворов фитохрома после облучения красным (660 нм) или дальним красным (730 нм) светом. Обратите внимание на то, что спектры поглощения двух форм перекрываются: при облучении светом 660 нм образуется около 75% Фдк и 25% Фк, а при 730 нм - примерно 2% Фдк и 98% Фк. Фитохром поглощает также в синей области, вызывая эффекты, промежуточные между эффектами красного и дальнего красного света. (Siegelman, Butler. 1965. Ann. Rev. Plant Physiol., 16, 383-392.)
[ОП при 660 нм - ОП при 730 нм] после облучения актиничным светом с длиной волны 660 обозначают ΔОП660.
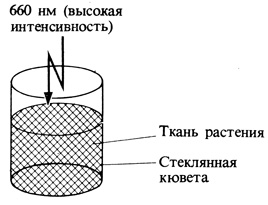
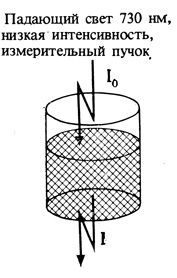
3. Затем ткань в кювете облучают дальним красным светом (730 нм) высокой интенсивности. Продолжительность актиничного облучения должна быть достаточна для предельной степени превращения Фдк в Фк.
4. Повторяют этап 2.
[ОП при 660 нм - ОП при 730 нм] после облучения актиничным светом обозначают ΔОП730.
5. ΔОП660-ΔОП730=Δ(ΔОП) есть мера содержания фитохрома в растительной ткани.
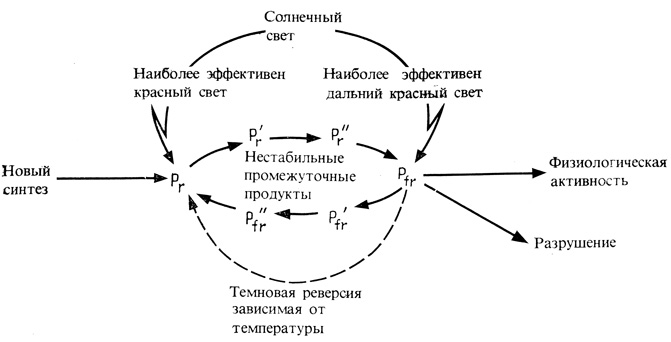
Рис. 11.10. Влияние красного и дальнего красного света на состояние фитохрома и его физиологическую активность. Фк(pr) и Фдк (pfr) в период фотоконверсии переходят через промежуточные короткоживущие формы в более стабильные формы, что показано короткими стрелками. Солнечный свет дает около 50% фитохрома в форме Фдк, 30% в форме Фк и остаток - в форме промежуточных продуктов
Из рис. 11.9 видно, что и Фк, и Фдк имеют широкие спектры поглощения, которые перекрываются в красной и дальней красной областях (660-730 нм) и в синей области (400-460 нм). Поэтому любая радиация в этих участках спектра превращает какую-то часть фитохрома в форму Фдк. Красный свет с длиной волны 660 нм наиболее эффективен и образует 75% Фдк, в то время как дальний красный свет с длиной волны 730 нм наименее эффективен и дает лишь 2% Фдк. Длины волн между 600 и 730 нм и синий свет обладают промежуточной эффективностью. Поскольку в ткани, выращенной в темноте, весь фитохром представлен в форме Фк, фактически любое облучение повышает уровень Фдк. Исключение составляет зеленый свет с длиной волны 500-550 нм, так как ни Фк, ни Фдк существенно не поглощают этих лучей. Поэтому в опытах с фитохромом зеленый свет используют как "безопасный".

Рис. 11.11. Некоторые примеры регуляции морфогенеза фитохромом. А. Проростки Sintiingia speciosci, выращиваемые при ежедневном 8-часовом периоде белого света высокой интенсивности, подвергали воздействию слабого света в течение 16 ч 15 ночей подряд. Для дополнительного освещения использовались: дальний красный свет, поддерживающий низкий уровень Фдк; (слева); холодная белая люминесценция, поддерживающая высокий уровень Фдк (в середине); контроль - темнота (справа). Обратите внимание на удлиненные вертикальные черешки и маленькие скрученные листовые пластинки у проростка, обработанного дальним красным светом. Растение, на которое воздействовали ночью белым светом, сходно с контрольным, но только его листья лучше пигментированы. (Satter, Wetherell. 1968. Plant Physiol. 43, 953-960.) Б. Оба проростка Chenopodium album выращивали в течение 21 дня под источниками излучения, обеспечивающими одинаковую плотность потока квантов в зоне ФАР (400-700 нм) и различную - в дальней красной области (>700 нм). Первое растение получило больше дальнего красного света, поэтому у него меньше фитохрома находится в форме Фдк. (Morgan Smith 1976 Nature, 262, 210-212.) В. Трехдневные проростки горчицы Sinapis alba, выращенные в темноте (слева), на свету (в середине) и росшие 42 ч в темноте, а затем 30 ч на свету (справа)
Превращения Фк⇔Фдк действуют как метаболический механизм, включающий и выключающий определенные реакции. Это переключение косвенно регулирует множество биофизических, биохимических, гистологических и морфологических процессов в растениях (рис. 11.11). Многие из наступающих изменений происходят после первого воздействия света на этиолированный проросток, когда некоторая часть его фитохрома переходит в форму Фдк. Эти изменения, обобщенно называемые деэтиоляцией, помогают растению адаптироваться к свету. При этом изменяется активность многих ферментов и содержание растительных гормонов, из этиопластов развиваются хлоропласты, происходит синтез хлорофилла, каротиноидов и антоциановых пигментов из предшественников. После позеленения этиолированных проростков система фитохромов продолжает влиять на рост и развитие растения в течение всей его жизни. Взаимопревращения Фк и Фдк не только влияют на индукцию цветения у растений как короткого, так и длинного дня, но и участвуют также в регулировании клубнеобразования, покоя, опадения листьев и старения. Однако эффект превращений фитохрома в растениях, выросших на свету, зависит также от времени воздействия света. Чувствительность таких растений к определенным формам фитохрома имеет ритмический характер. Эта интересная проблема будет рассмотрена в следующей главе.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'