Открытие фитохрома
История открытия фитохрома в начале 50-х годов группой исследователей министерства сельского хозяйства США составляет одну из самых волнующих глав в истории изучения растений. Эта история началась примерно на 30 лет раньше с работы двух экспериментаторов, имевших дело с табаком. У. Гарнер и Х. Аллард пытались размножить мутантный тип крупнолистного табака - Мэриленд Маммот, который случайно вырос в единственном экземпляре среди других табачных растений. В определенное время исходный тип обильно зацвел, а Мэриленд Маммот - нет. Желая получить семена этого ценного нового типа и боясь, что растение может не зацвести до осенних холодов, ученые решили перенести его в теплицу. Однако, несмотря на все принятые меры, растение упорно оставалось в вегетативном состоянии примерно до середины декабря, когда на нем появились зачатки цветков - через несколько месяцев после того, как у нормальных растений успешно завершилось образование семян.
Анализ разнообразных факторов, которые могли бы быть ответственны за это необычное поведение, привел Гарнера и Алларда к неизбежному выводу, что растение зацвело под влиянием очень малой длины дня в северном полушарии перед рождеством. Они установили, что цветение можно вызвать переносом растений Мэриленд Маммот в специальные камеры с искусственно сокращенным световым днем (рис. 11.1). Табак Мэриленд Маммот, зацветающий лишь в том случае, если длина дня меньше некоторой критической величины, стали называть растением короткого дня. К этой категории относятся и многие другие растения, в том числе некоторые формы сои, хризантем и пуансеттии. Напротив, шпинат и некоторые зерновые зацветают только тогда, когда длина дня превышает определенную критическую величину; это растения длинного дня. Наконец, имеется группа растений (томаты и др.), цветение которых не зависит от длины дня (нейтральные растения). Реакция растений на длину дня называется фотопериодизмом.

Рис. 11.1. Растения табака Мэриленд Маммот, выращенные в условиях короткого дня {слева) и длинного дня (справа). (A. E. Murneek, R. O. Whyte. 1948. Vernalization and Photoperiodism, Waltham, Mass.: Chronica Botanica Co.)
Критический фотопериод для разных видов и даже сортов растений как короткого, так и длинного дня значительно варьирует. Он составляет 14 ч для Билокси - короткодневного сорта сои, растущего на широте 35°, тогда как сорт Баторавка, обычно выращиваемый на широте 45° и выше, будет цвести даже при непрерывном освещении. Эти различия в критическом фото- периоде играют важную роль в распределении растений на земном шаре.
В годы, последовавшие за открытием фотопериодизма, стало ясно, что многие растения реагируют на продолжительность .непрерывной темноты, а не на длину светлого периода. Иными словами, так называемое растение короткого дня - это на самом деле растение "длинной ночи". Для заложения цветочных бугорков ему необходима определенная минимальная продолжительность темноты, не прерываемой светом. Аналогичным образом растение длинного дня может быть в действительности растением "короткой ночи": оно будет цвести лишь в том случае, если ночной период не длиннее некоторого максимума.
Эффективный период темноты для короткодневного растения можно сделать неэффективным путем простой уловки - достаточно немного укоротить его (всего на несколько минут) или прервать в середине вспышкой слабого света. Это говорит о том, что растение может "измерять" продолжительность темноты с точностью до нескольких минут и что фотопериодизм связан с работой необычайно чувствительной световоспринимающей системы. Короткодневное растение дурнишник цветет при режиме 15 ч света+ 9 ч темноты, но не будет цвести, если темный период составляет 8,5 ч или же 9-часовой темный период прерывается коротким световым периодом (рис. 11.2). Всего лишь один подходящий период темноты может привести к цветению, даже если последующие темные периоды недостаточно длинны. Этот феномен известен как фотопериодическая индукция. У многих растений длинного дня имеет место аналогичный, но обратно направленный феномен: прерывание слишком длинного темного периода вспышкой света приводит к индукции и к заложению цветков. Таким образом, растения длинного и короткого дня, видимо, обладают сходным фотопериодическим механизмом, но последний каким-то образом действует в разных направлениях.
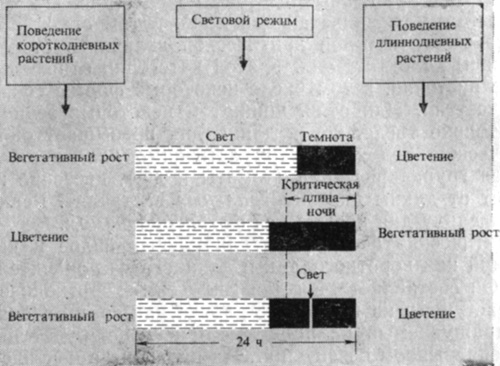
Рис. 11.2. Влияние прерывания темнового периода пучком света на цветение растений короткого и длинного дня
Короткодневный сорт сои Билокси так чувствителен к свету, что индуктивный эффект длительных темновых периодов можно снять даже минутным облучением с помощью ламп накаливания (без фильтра) в середине ночи. По этой причине Х. Бортвик и С. Хендрикс с сотрудниками пришли к выводу, что это растение было бы идеальным объектом для выяснения вопроса о том, какие длины волн наиболее эффективно предотвращают инициацию цветения; а эти сведения в свою очередь могли бы помочь в идентификации фоторецепторного пигмента, участвующего в контроле цветения. Поэтому они определили спектр действия для данного процесса, используя большой спектрограф для одновременного облучения групп растений светом с разной длиной волны (рис. 11.3). Полученные спектры действия для ингибирования цветения короткодневных растений сои и дурнишника и для активации цветения длиннодневных растений Hordeum (ячмень) и Hyoscyamus (белена) оказались поразительно сходными (рис. 11.4). Во всех случаях был обнаружен максимум активности в красной области спектра (около 660 нм) при почти полной неэффективности других областей. Сходство спектров позволяло считать вероятным, что зацветание растений как короткого, так и длинного дня контролируется одним и тем же пигментом. Анализ спектра действия привел к предположению, что поглощающий пигмент сходен с пигментом водорослей фикоцианином, который родствен желчным пигментам животных. К сожалению, такого рода пигментов авторы в своих подопытных растениях не нашли и поэтому стали вести иоиски в другом направлении.
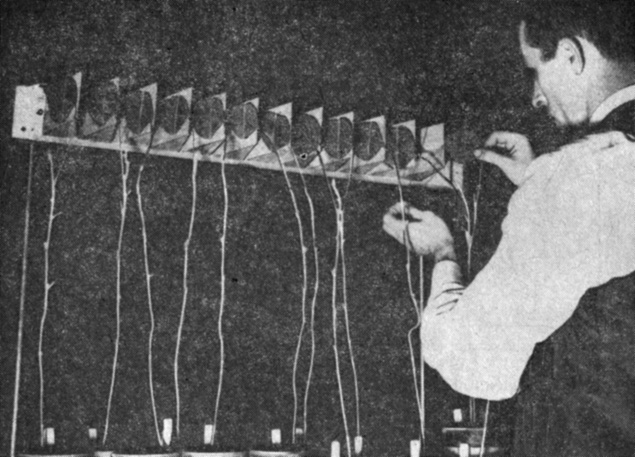
Рис. 11.3. Метод, использованный Бортвиком, Хендриксом и их сотрудниками для удерживания одиночных листьев (в данном случае листочков сои) в плоскости изображения спектрографа с целью последующего облучения светом с различной длиной волны. (Hendricks, Borthwick. 1964. Proc. First Int. Photobiol. Congr., 23-25.)
Давно было известно, что свет сильно влияет на прорастание некоторых семян. Например, увлажненные семена салата сорта Гранд Рапиде прорастают плохо в полной темноте, но быстро и хорошо при экспонировании на свету в течение нескольких минут. Оказалось, что спектр действия для этого эффекта сходен со спектром действия для цветения! Аналогично этому проросток гороха, выращенный в полной темноте, имеет очень длинный, тонкий непигментированный стебель, искривленный апикальный изгиб и почти не распустившиеся листья. Если такой этиолированный проросток выставить лишь на короткое время на свет, листья после этого разворачиваются, изгиб начинает распрямляться и удлинение стебля замедляется (рис. 11.5). Спектр действия и здесь фактически идентичен спектру для цветения. Приходится заключить, что такие разнородные реакции, как прорастание семян, изменение этиолированного проростка (его "деэтиоляция") и индукция цветения, регулируются одним и тем же пигментом-рецептором.
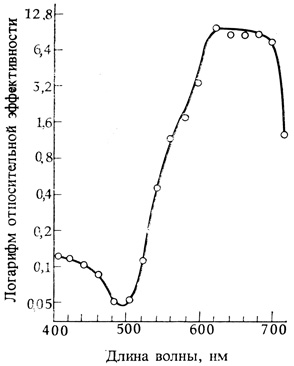
Рис. 11.4. Обобщенный спектр действия для фотопериодизма (вычислен по данным Parker et al. 1949. Amer. J. Bot., 36, 194-204)
Но что это за пигмент? Ответ был найден в результате новой интерпретации старых опытов па прорастанию семян салата, проведенных в 1935 г. Л. Флинтом и Э. Мак-Алистером. Эти исследователи показали, что прорастание семян салата Гранд Рапиде не только стимулируется красным светом, но также подавляется светом дальней красной области спектра с длиной волны около 730 нм (рис. 11.6). Это открытие привело к гипотезе, что красный свет изменяет пигмент в одном направлении, а дальний красный - в другом. Опыты с последовательным воздействием красного (К) и дальнего красного (ДК) света подтвердили эту гипотезу. В таких условиях семя реагировало лишь на последнее облучение, как будто форма пигмента, закрепленная при последней экспозиции, была единственным эффективным фактором (рис. 11.7). Даже семена, облученные 100 раз попеременно красным и дальним красным светом, прорастали так же, как и облученные тем и другим светом по одному разу. Во всех случаях реакцию прорастания определяло последнее световое воздействие, предшествующее темноте.
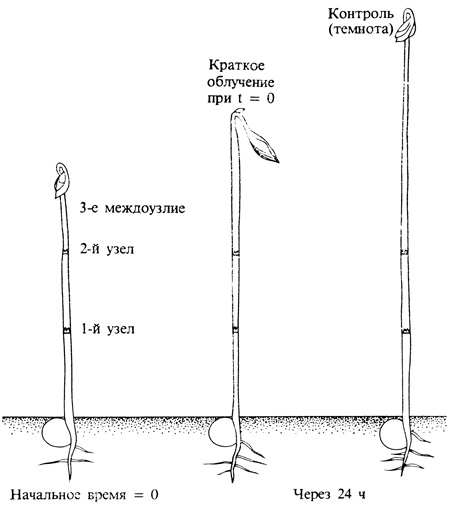
Рис. 11.5. Семидневный этиолированный проросток гороха до 5-минутного облучения красным светом и через 24 ч после него
Затем Бортвик и Хендрикс изучили влияние дальнего красного света на реакции цветения и диэтиолирования, вызываемые красным светом. И здесь была обнаружена "обратимость К - ДК" (рис. 11.8). Это позволило предсказать, что существует один активный пигмент в двух взаимно фотообратимых формах, одна из которых образуется под действием красного, а другая - дальнего красного света (рис. 11.9). Обратимые сдвиги поглощения при 660 и 730 нм вслед за облучением соответственно красным и дальним красным светом дали возможность легко выявлять пигмент и измерять его количество, особенно в этиолированных растениях, где экранирование хлорофиллом не создает затруднений (см. ниже). Вскоре пигмент был обнаружен в экстрактах растений, сконцентрирован, очищен, подвергнут анализу и частично охарактеризован. Его назвали фитохромом (от греческих слов, означающих "растение" и "краситель"). Его две формы были названы соответственно Фк (фитохром, поглощающий красный свет) и Фдк (поглощающий дальний красный свет). Фитохром синтезируется в форме Фк. Поэтому этиолированные проростки содержат Фк, а не Фдк. Облучение красным светом превращает большую часть фитохрома в. Фдк - физиологически активную форму, тогда как последующее облучение дальним красным светом превращает Фдк обратно в Фк (рис. 11.10).
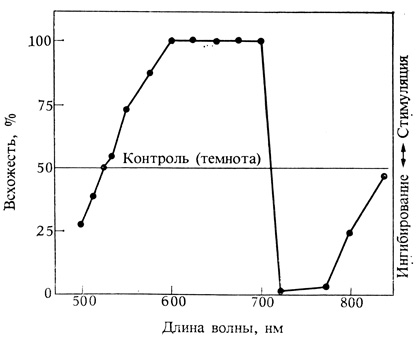
Рис. 11.6. Семена чувствительного к свету салата, у которых слабым белым светом индуцировано 50%-ное прорастание, можно стимулировать далее красным светом (600-690 нм) или ингибировать дальним красным светом (720-780 нм). (Flint, McAlister. 1937. Smithsonian Inst. Misc. Collection, 96, 1-8.)
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'