Цитокинины
Выше мы уже упоминали, что деление клеток в культуре растительной ткани зависит от ауксина и других факторов клеточного деления, называемых цитокининами. Такие же контрольные механизмы действуют, вероятно, при регулировании деления и роста клеток и в других частях растущего растения, например в боковых почках.
Открытие цитокининов было связано с попытками индуцировать деление определенных растительных клеток, выращиваемых в культуре. Одна из таких попыток, придпринятая в начале 40-х годов, заключалась в выделении неоплодотворенной яйцеклетки из завязи и (выращивании ее в определенной среде с целью получения гаплоидных зародышей. Эта цель не была достигнута. Однако было установлено, что рост очень молодых нормальных зародышей стимулируется при добавлении к среде молока кокосового ореха (жидкого эндосперма огромных семян кокосовой пальмы), Поскольку никакие сочетания известных тогда факторов роста не давали такого результата, было ясно, что молоко кокосового ореха содержит какое-то вещество (или вещества), ответственное за этот уникальный эффект. Поэтому несколько исследователей предприняли попытки выделить из молока кокосового ореха и охарактеризовать гипотетические факторы клеточного деления.
К открытию цитокининов привели опыты Скуга и его сотрудников из Висконсинского университета, в которых они пытались индуцировать деление зрелых сердцевинных клеток табака. В интактных растениях эти клетки увеличиваются в размерах, но никогда не делятся. Однако при пересаживании на среду, содержащую ауксин и молоко кокосового ореха, они начинают делиться. Испытывая разные соединения, группа Скуга обнаружила, что аденин в присутствии ауксина обладает некоторой стимулирующей клеточное деление активностью. Впоследствии были проверены также нуклеиновые кислоты, в состав которых входит аденин. В итоге было установлено, что ДНК, содержащаяся в одном старом образце спермы сельди, дала результаты, количественно превосходящие те, которые были получены с аденином. Позже выяснилось, что любой образец ДНК или даже одного аденозина можно превратить в чрезвычайно активный материал, если автоклавировать его в соответствующих условиях. Новое вещество, образовавшееся в таких необычных условиях, было выделено и идентифицировано как 6-фурфуриладенин. Из-за его высокой активности при индуцировании клеточного деления (цитокинеза) в сердцевине табака (в присутствии ауксина) это вещество было названо кинетином.
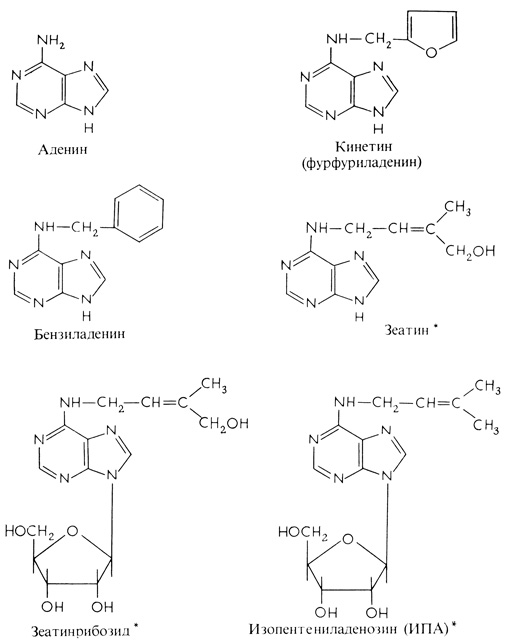
Рис. 9.27. Структурные формулы различных производных аденина с активностью, характерной для цитокининов. Звездочкой помечены природные цитокинины
После установления природы кинетина исследователи смогли заняться поисками в растениях других подобных материалов. Оказалось, что незрелые плоды и развивающиеся зерна злаков являются богатыми источниками активности, стимулирующей клеточное деление. В конечном счете из развивающихся зерен кукурузы было выделено вещество, названное зеатином (рис. 9.27), которое по своей структуре напоминало кинетин. Это вещество и близкородственное соединение изопентениладенин обнаружены сейчас во всех проверенных растениях. Такие соединения, получившие общее название цитокинины, обычно связаны с рибозой (рибозиды) или с рибозой и фосфатом (риботиды). Установлено, что стимулирующая активность молока кокосового ореха обусловлена действием смеси многих соединений, среди которых цитокинин был идентифицирован как зеатин- рибозид. В научных исследованиях широко используются синтетические цитокинины, такие, как кинетин и бензиладенин, так как чем активнее природные цитокинины, тем труднее их получать.
Взаимодействие цитокинина с ауксином
В присутствии ауксина цитокинины чрезвычайно активны при индуцировании деления растительных клеток в культуре тканей. Иногда они стимулируют также развитие почек в образовавшемся каллусе. Дифференциация в культуре тканей зависит преимущественно от относительной концентрации присутствующих ростовых веществ. Высокое молярное отношение цитокинина к ауксину приводит к образованию почек. Если концентрации цитокинина и ауксина примерно равны, то стимулируется рост недифференцированного каллуса. При высоком отношении ауксина к кинетину наблюдается тенденция к инициации роста корней (рис. 9.28). В типичных системах с клеточным растяжением, таких, как колеоптиль овса и эпикотиль гороха, кинетин обычно ингибирует стимулированный ауксином продольный рост и стимулирует поперечный.
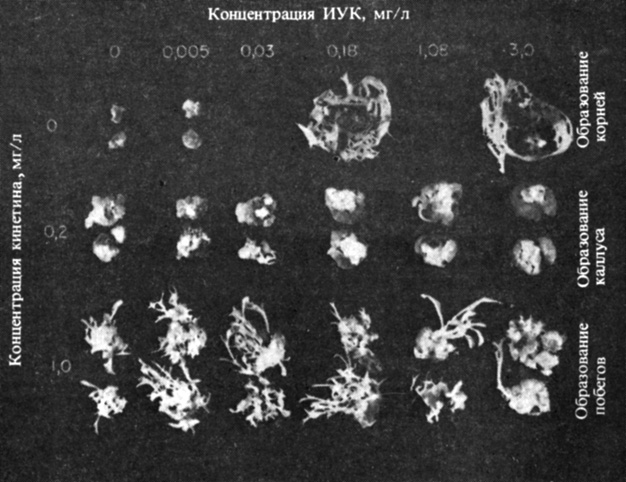
Рис. 9.28. Взаимодействия кинетина и ИУК при образовании каллуса и дифференциации тканей в культуре тканей табака. Для максимального образования тканей необходимо присутствие ауксина и кинетина. При высоком отношении ауксина к кинетину образуются корни, при высоком отношении кинетина к ауксину - побеги, а при равных концентрациях преобладает образование каллуса. (Skoog, Miller. 1957. Symp. Soc. Exp. Biol., 11, 118-131.)
Обнаружение взаимодействия кинетина с ауксином потребовало систематического изучения других физиологических систем, таких, как апикальное доминирование, на которое, как известно, влияет ауксин. При этом было установлено, что локальное нанесение кинетина на репрессированные почки устраняет ингибирование и вызывает рост этих почек (рис. 9.29). Было также показано, что ингибирование роста боковых почек ауксином в действительности обусловлено индукцией образования в них этилена (см. гл. 10). Вместе с тем внесение цитокинина ингибирует синтез этилена. Развертывание молодых листьев также стимулируется цитокининами.

Рис. 9.29. Устранение апикального доминирования с помощью кинетина. Почка на растении справа три дня назад была локально обработана кинетином в концентрации 330 ч. на млн. (Sachs, Thimann. 1967. Amer. J. Bot., 54, 136-144.)
У взрослых растений синтез цитокининов происходит главным образом в корневой системе, откуда они по ксилеме транспортируются в надземную часть растения. Чем ниже на стебле расположены боковые почки, тем дальше они от источника ауксина, находящегося в апексе стебля, и тем ближе к источнику цитокининов в корнях. Так как апекс растения, проявляющего апикальное доминирование, в процессе роста удаляется от боковых почек, последние начинают распускаться в результате того, что цитокинин преодолевает влияние ауксина. Антагонизмом ауксина и цитокинина объясняется также образование "ведьминой метлы" вследствие полного устранения нормального подавления роста покоящихся почек, отделенных от апекса лишь несколькими короткими междоузлиями (рис. 9.30). В природе это происходит тогда, когда гриб, индуцирующий образование "ведьминой метлы", проникает в растение и вырабатывает вещества с активностью, характерной для цитокининов.

Рис. 9.30. 'Ведьмина метла' на иве возникает в результате роста многочисленных боковых почек, вызванного образованием цитокинина проникшим в растение патогеном (ср. с равномерным расположением веток на дереве справа сверху). (С любезного разрешения W. A. Sinclair, Cornell University.)
Действие цитокининов лежит в основе другого феномена, над которым долго ломали головы физиологи растений. Часто приходилось наблюдать, что если с растения, например табака, удалить листья, то содержание белка в них быстро снижается, а содержание растворимого азота - возрастает. Предполагалось, что этим массированным разрушением белка по крайней мере частично объясняется короткий период жизни многих срезанных растений и их частей, особенно листьев. Случайно было обнаружено, что добавление кинетина в питательный раствор, в который погружены черешки листьев, срезанных с растения Xanthium, приводит к более длительному сохранению зеленого цвета листьев. Таким образом, цитокинины задерживают старение. Впоследствии было показано, что данный эффект обусловлен тем, что цитокинины способствуют поддержанию определенных уровней белка и нуклеиновой кислоты, вероятно, путем снижения скорости их распада, а также в результате сохранения целостности клеточной мембраны. Было отмечено, что нанесение цитокинина на листья интактных растений тоже задерживает их старение (рис. 9.31). Вероятно, цитокинин должен постоянно содержаться в воде, поступающей от корней к листьям, чтобы препятствовать старению последних.

Рис. 9.31. Одной из важных функций цитокининов в растении является замедление процессов старения. Самые нижние первичные листья на растении фасоли (Phaseolus vulgaris) обычно желтеют и опадают, тогда как более молодые листья развиваются (растение слева). Нанесение цитокинина бензиладенина на листья растения, расположенного справа, замедлило старение, и листья сохранили свою зеленую окраску. (С любезного разрешения R. A. Fletcher, University of Guelph, Canada.)
Как действуют цитокинины
В отсутствие цитокинина синтез ДНК продолжается до тех пор, пока в клетке имеется ауксин, но клетки при этом не делятся. Цитокинин требуется для некоторого процесса, происходящего после завершения репликации ДНК, но до начала настоящего митоза. Этим процессом может быть образование необходимых для клеточного деления белков, вероятно, тех, которые связаны с митотическим аппаратом. Действительно, после добавления цитокининов наблюдается появление различных белков, и это происходит даже тогда, когда синтез новой мРНК блокируется ингибиторами. Поэтому вполне вероятно, что цитокинины контролируют трансляцию предобразованных мРНК, продукты которых имеют отношение к некоторым аспектам клеточного деления.
Другой ключ к механизму функционирования цитокининов может лежать в их структуре. Цитокинины родственны компонентам нуклеиновых кислот. Фактически цитокинин можно обнаружить как одно из оснований, расположенных рядом с антикодоном в молекулах нескольких тРНК, и в этом положении он, вероятно, участвует в связывании тРНК с рибосомами в процессе синтеза белка (см. рис. 2.11). Однако из этого нельзя понять, каким образом цитокинины функционируют в качестве гормонов, так как после нанесения на растительную ткань они не включаются как таковые в тРНК. Скорее, они расщепляются до аденина, который после включения в полинуклеотидную цепь модифицируется путем присоединения боковой цепи - изопентенила. Тем не менее структурное сходство цитокининов с компонентами нуклеиновых кислот, а также характер их действия на растение свидетельствуют о том, что цитокинины, возможно, играют какую-то роль в синтезе белка. Действительно, молекулы цитокинина непрочно связываются с рибосомами - тельцами, на которых в клетке синтезируются белки. Местом связи фактически служит белок, который сам соединен с рибосомами. Цитокинин и связывающий белок могут быть частью фактора инициации, участвующего в синтезе специфического белка на рибосомах. Контролируют ли цитокинины именно таким путем клеточное деление, а возможно, и другие аспекты клеточного метаболизма, остается пока неизвестным.
Так же, как и в случае других гормонов, функционирование цитокинина осуществляется, вероятно, разными путями, приводящими к различным эффектам. Рассмотрим, например, каким образом цитокинин предотвращает старение листьев. В присутствии цитокинина уровни белка в листьях выше, чем в его отсутствие. В этом случае цитокинин не ускоряет синтез белка, а ингибирует его расщепление. Как уже упоминалось, было показано, что цитокинин поддерживает целостность клеточных мембран. Как он выполняет эти функции, в настоящее время неизвестно. Но механизм станет ясным, когда мы узнаем больше о функционировании гормонов на молекулярном уровне.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'