Гиббереллины
Еще одну группу важных ростовых гормонов растений, открытую благодаря ряду случайных обстоятельств и тонких наблюдений, составляют гиббереллины. В последнем десятилетии прошлого века японские рисоводы обнаружили на своих чеках появление чрезвычайно вытянутых проростков. Они стали (внимательно наблюдать за этими проростками, так как хороший фермер обычно рассматривает любое крупное растение как возможный материал для селекции сорта с целью улучшения его общей продуктивности. Однако эти высокие проростки никогда не доживали до созревания и лишь изредка зацветали. Болезнь была названа баканаэ (болезнь "дурных проростков"). В 1926 г. японский ботаник Куросава обнаружил, что эти проростки были заражены грибом, названным позднее Gibberella fujikuroi (представитель кл. Ascomycetes, или сумчатых грибов). Если споры гриба перенести с зараженного проростка на здоровое растение, то последнее заболевает и его рост становится аномальным. При выращивании гриба в колбе на искусственной среде в ней накапливается какое-то вещество, которое, будучи перенесено на растение-рецептор, вызывает у него чрезмерный рост - один из симптомов, типичных для болезни "дурных проростков". Это вещество было названо гиббереллином (производное от Gibberella).
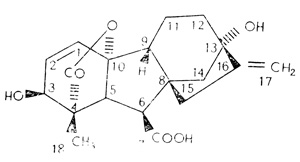
Рис. 9.19. Структурная формула гибберелловой кислоты (ГК3). Имеется свыше 50 различных гиббереллинов, различающихся главным образом по числу и местоположению дополнительных групп, таких, как -OH. Для того чтобы описать эти различия, каждому атому углерода в основном скелете был присвоен номер, причем каждая группа получила номер того атома углерода, к которому она присоединена. Наконечниками стрелок обозначены связи, выступающие над плоскостью листа; короткие штрихи - это связи, находящиеся ниже плоскости листа, а сплошные линии - связи в плоскости листа
В 30-х годах японским физиологам и химикам удалось выделить из среды, на которой выращивали гриб Gibberella, несколько веществ, ингибирующих и стимулирующих рост. Предложенная ими в итоге структурная формула стимулятора роста гиббереллина была не совсем верна. Тем не менее эти ученые правильно определили общую природу вещества и получили кристаллы, при нанесении которых на испытуемые растения наблюдались типичные для болезни баканаэ симптомы очень сильного удлинения стебля. Эта информация была опубликована в Японии во многих статьях еще до 1939 г., но, к сожалению, вторая мировая война прервала начатую работу и отвлекла внимание большинства ученых на исследования для военных целей. Волнующая история открытия гиббереллина оставалась неизвестной на Западе примерно до 1950 г., когда несколько групп исследователей в Англии и США обнаружили старые статьи о гиббереллине и вновь приступили к решению этой проблемы.
К 1955 г. английские ученые подтвердили первоначальное наблюдение, сделанное в Японии, а также выделили вещество, которое они назвали гибберелловой кислотой (рис. 9.19). Она несколько отличается от материала, выделенного японцами. Вскоре много других соединений с такой же основной структурой было обнаружено как в грибах, так и в незараженных тканях высших растений. Стало ясно, что гиббереллины представляют собой целое семейство молекул, насчитывающее в настоящее время свыше 50 соединений. Все они обладают основным тетрациклическим гиббереллановым скелетом, но у каждого из них имеются различные модификации и химические группы (например, -OH) в разных частях молекулы. Гибберелловая кислота при нанесении на некоторые растения вызывает чрезмерное удлинение стебля, а в ряде случаев и уменьшение листовой поверхности. Самое яркое проявление ее действия - это, пожалуй, быстрая стимуляция удлинения цветоножки (стрелкование) и во многих случаях - стимуляция цветения длиннодневных растений (см. гл. 11 и 12; рис. 9.20). Гиббереллин способствует выбрасыванию стрелки, увеличивая как число клеточных делений в определенных зонах, так и растяжение клеток, образующихся путем таких делений (рис. 9.21). В тех случаях, когда наносятся лишь небольшие количества гиббереллина, выход в стрелку происходит, но зачатки цветков могут и не дифференцироваться. Более высокие дозы гиббереллина обычно вызывают не только стрелкование, но и цветение. Есть некоторые данные в пользу утверждения, что у длиннодневных растений индукция длинным днем стимулирует образование гиббереллина, что в свою очередь вызывает морфогенетические реакции. В других случаях активность гиббереллина в апексах растений снижается во время их выхода в стрелку, что свидетельствует о возможном использовании гиббереллина в этот период. Гиббереллин не способен вызывать цветение у короткодневных растений и фактически оказывает, по-видимому, противоположный эффект. Пока точно не известно, какие гормоны контролируют цветение у короткодневных растений или у тех длиннодневных растений, которые вытягиваются, но не зацветают в ответ на воздействие гибберелловой кислотой. Возможно, что в последнем случае в индукцию цветения вовлечена не гибберелловая кислота, а какой-то другой гиббереллин.

Рис. 9.20. Один из главных эффектов, проявляемых гиббереллином, заключается в индуцировании растяжения стебля, особенно цветоножки, у растений, для удлинения и цветения которых необходимы длинные дни. За таким стрелкованием иногда, но не всегда следует цветение. Растения капусты справа были обработаны гибберелловой кислотой, тогда как такие же растения слева не обработаны. (С любезного разрешения S. H. Wittwer и M. J. Bukovac, Michigan State Umversity.)
Однако не все растения реагируют на гиббереллин. Анализируя результаты многочисленных тестов, физиологи обнаружили корреляцию между исходным размером растения и степенью его реактивности в отношении гиббереллина. Если сравнивать, например, карликовый горох или карликовую кукурузу с их высокорослыми формами, то выясняется, что нанесение гиббереллина на карликовые растения вызывает их усиленный рост, тогда как при аналогичной обработке высокорослых форм эффект незначителен или его нет совсем (рис. 9.22). Поскольку во многих случаях различие между карликовостью и высокорослостью определяется только одним геном, предложена привлекательная гипотеза, согласно которой карликовость фактически обусловлена неспособностью растения продуцировать достаточное количество гиббереллина для удовлетворения своих основных потребностей. Следовательно, нанесение гиббереллина на некоторые генетические карлики дает высокорослые формы. Такие карликовые растения, ставшие высокими благодаря обработке гиббереллином, конечно, все равно будут иметь карликовые генотипы и при размножении будут давать карликовое потомство. Организмы, фенотипы которых изменены с помощью химической или физической обработки так, что они становятся похожими на организмы с другим генотипом, называются фенокопиями.
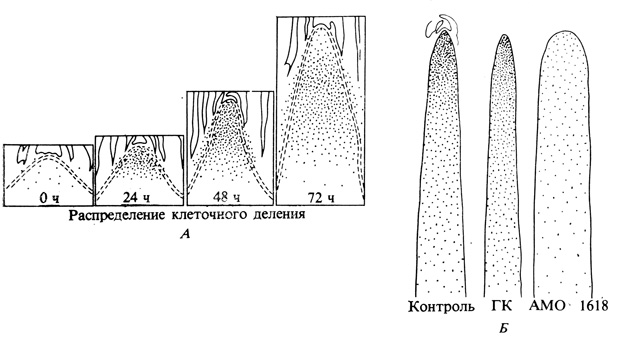
Рис. 9.21. Обработка гиббереллином розеточных растений вызывает усиление как деления, так и растяжения клеток. Это легко можно видеть на продольных срезах через центральный цилиндр Samolus parviflorus (А), в котором деление клеток усиливается после внесения ГК. (Каждая точка соответствует одной митотической фигуре на срезе толщиной 64 мк.) Используя АМО 1618, антигиббереллиновый ретардант роста, можно показать, что меристематическая активность верхушек стебля у Chrysanthemum morifolium непосредственно обусловлена действием ГК. Сплошные линии показывают границы сосудов, в пределах которых каждая точка представляет одну митотическую фигуру на срезе толщиной 60 мк. (Sachs. 1965. Ann. Rev. PI. Phys., 16, 73-96; Sachs et al. 1959. Amer. J. Bot.,46, 376-384.)
Установлено, что некоторые высокорослые растения действительно содержат больше общего количества гиббереллина, чем карликовые растения, хотя это и не всегда так. Однако при хроматографическом разделении гиббереллины высокорослых и карликовых растений часто обнаруживают некоторые качественные различия. В таких случаях карликовость может быть обусловлена наличием гиббереллинов, менее активных в стимуляции роста. Но ясно, что карликовость не всегда связана с аномалиями в гиббереллинах.

Рис. 9.22. А. Влияние гибберелловой кислоты на нормальную и карликовую кукурузу. Слева направо представлены нормальное контрольное растение, нормальное растение, обработанное гиббереллином, карликовое контрольное растение и карликовое растение, обработанное гиббереллином. (В. О. Phinney, С. A. West. Gibberellins and the Growth of Flowering Plants в кн. Developing Cell Systems and Their Control. D. Rudnick, 1960. The Ronald Press.) Б. Карликовый горох, обработанный водой (слева) и ГК (справа). (С любезного разрешения A. Lang, Michigan State University.)
Гиббереллин может также индуцировать образование партенокарпических плодов, действуя самостоятельно или совместно с ауксином. Примером может служить яблоко. Многочисленные попытки добиться его партенокарпического развития с помощью одного ауксина не увенчались успехом. Сейчас опрыскивание растений смесью ауксина и гиббереллина приносит желаемые результаты. Повышение содержания природного гиббереллина в семенах яблока коррелирует также с периодом их максимального роста, что свидетельствует о возможной регуляции гиббереллином развития семени и стенки завязи. Последний аспект функционирования гиббереллина нашел применение в сельском хозяйстве при выращивании бескосточкового винограда.
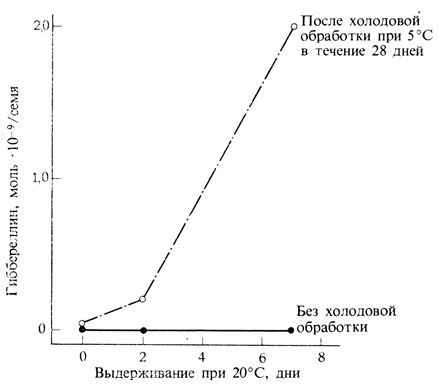
Рис. 9.23. Некоторым семенам для прорастания необходима холодовая обработка. Предварительное воздействие низкой температуры на семена лещины (Corylus avellana) включает механизм образования гиббереллина после того, как семена вновь оказываются при температуре прорастания. Образовавшийся гиббереллин способствует их прорастанию. (Ross, Bradbeer. 1968. Nature, 220, 85-86.)
Гиббереллины играют определенную роль, вероятно, не только в развитии семян, но также и в их прорастании. При этом они действуют двумя путями. Во-первых, они, по-видимому, способствуют выходу семян из состояния покоя, что можно легко продемонстрировать, нанеся гибберелловую кислоту на покоящиеся семена, которые после этого прорастают. Гиббереллин заменяет также свет или низкие температуры в тех случаях, когда для прерывания покоя семян требуются эти раздражители. В природе покой, вероятно, прерывается при повышении содержания естественного гиббереллина (рис. 9.23). Во-вторых, в семенах злаков гиббереллин является тем веществом, которое контролирует мобилизацию запасного питательного материала в эндосперме. Семена многих злаков, например ячменя, содержат запасной крахмал, способный быстро гидролизоваться в начале прорастания. После замачивания зерен ячменя, содержащих зародыши, начинается быстрый гидролиз крахмала. Если же. перед замачиванием удалить зародыши, то в семенах гидролиза крахмала не происходит. При нанесении гиббереллина на такие лишенные зародышей семена крахмал начинает гидролизоваться (рис. 9.24). Таким образом, в зародыше вскоре после набухания семян в норме, очевидно, образуются гиббереллины, активизирующие процесс гидролиза крахмала с помощью особого механизма, который будет рассмотрен более подробно в следующем разделе. Два описанных эффекта гиббереллина, проявляющихся при прорастании семян, полностью различаются между собой в том отношении, что прерывание покоя происходит в зародыше до мобилизации запасных питательных веществ. Последний процесс начинается в результате действия гиббереллина на алейроновый слой, окружающий эндосперм (рис. 9.25.)
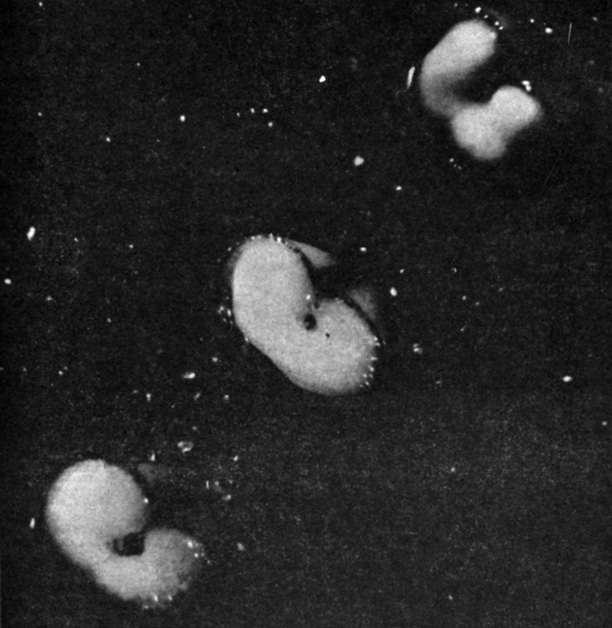
Рис. 9.24. Три стерильные половинки зерен ячменя без зародыша. На открытую поверхность каждой из них было нанесено (слева направо) 0,5 мкл воды, ГК в концентрации 1 ч. на млрд. и 100 ч. на млрд. Фотография, сделанная через 48 ч, показывает, что в зернах, обработанных гиббереллином, уже идет переваривание заполненной крахмалом запасающей ткани. Гормон гиббереллин активирует образование и выделение ферментов, вызывающих гидролиз запасных веществ. (С любезного разрешения J. E. Varner, Washington University.)
По-видимому, имеются два пути контроля гиббереллином специфических физиологических процессов. Первый путь - это просто синтез гиббереллина растением и последующая инициация гиббереллин-зависимого процесса. Второй путь более сложен. Вспомните, что существуют около 50 гиббереллинов, различающихся по своей относительной активности в зависимости от того процесса, на который они оказывают влияние. Поскольку все гиббереллины имеют сходную молекулярную структуру, любой из них можно превратить в другой, лишь слегка модифицировав его молекулу, например присоединив к ней -OH- группу в одно или более положений из нескольких возможных. Таким образом, растение может управлять тем или иным внутренним процессом, превращая в ходе метаболизма неактивный гиббереллин в активный или наоборот.
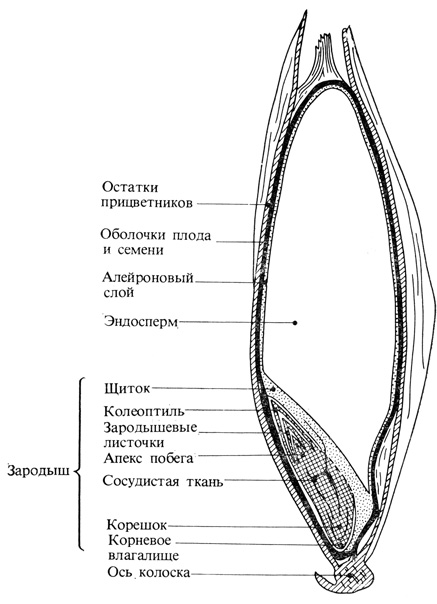
Рис. 9.25. Продольный разрез зерна ячменя
Подобно каротиноидным пигментам, стероидам и каучуку, гиббереллины представляют собой изопреноидные соединения образующиеся из ацетил-СоА придыхательном метаболизме (см гл. 5). Метаболический путь биосинтеза гиббереллинов, выяс ненный с помощью меченных 14С соединений, отображает следующая схема:
Идентификация дитерпенов в качестве промежуточных соединений в биосинтезе гиббереллинов подтверждается еще и тем фактом, что ретардант роста АМО-1618, действующий как антагонист гиббереллинов, препятствует образованию дитерпена из его предшественника.
Как действует гиббереллин
При рассмотрении гиббереллина, так же как и аукоина, мы сталкиваемся с одной и той же проблемой: как можно объяснить, что очень малые количества данного вещества способны контролировать многочисленные и разнообразные морфогенетические реакции, включающие прорастание семян, деление и растяжение клеток, закладку цветков. Лишь один феномен был проанализирован подробно - индукция гидролиза крахмала в беззародышевых семенах ячменя.
Мы теперь знаем, что контроль расщепления крахмала гиббереллином сводится к регуляции образования и высвобождения ферментов. Нанесение гиббереллина на беззародышевые семена ячменя приводит к появлению и выделению амилазы (рис. 9.26), а также других ферментов. Амилаза вызывает гидролиз крахмала (по-латыни amylum), содержащегося в эндосперме зерна ячменя. Если удалить алейроновый слой, то можно показать, что образование фермента происходит именно в этой ткани. Следовательно, алейрон продуцирует и выделяет гидролитические ферменты, расщепляющие запасные питательные вещества в эндосперме. Именно эти алейроновые клетки, являющиеся "клетками-мишенями", и реагируют на гиббереллин. Данная система может служить примером органоспецифической регуляции роста, так как гиббереллин - своего рода ключ к запасным питательным веществам - образуется в зародыше, содержащем единственные в семени способные к росту ткани.

Рис. 9.26. Процессы, происходящие в алейроновых клетках зерна ячменя после обработки его гиббереллином
Каким образом гиббереллин вызывает проявление α-амилазной активности? Во-первых, очевидно, что фермент представляет собой не просто активированную форму предварительно синтезированного неактивного запасного белка, а образуется заново из составляющих его аминокислот. Это было показано путем добавления меченых аминокислот к зернам ячменя или алейроновым слоям, инкубированным с гиббереллином. В результате происходило включение радиоактивности в белок. Это включение предотвращалось ингибиторами синтеза белка, такими, как циклогексимид. На место действия гиббереллина в процессе синтеза белка указывает тот факт, что ингибиторы ДНК-зависимого синтеза РНК (например, актиномицин D) препятствуют также и синтезу амилазы. Отсюда можно заключить, что гиббереллин должен участвовать в образовании молекул мРНК на ДНК-матрице в качестве дерепресеора генов, кодирующих гидролитические ферменты; он как бы дает разрешение на выработку этих ферментов.
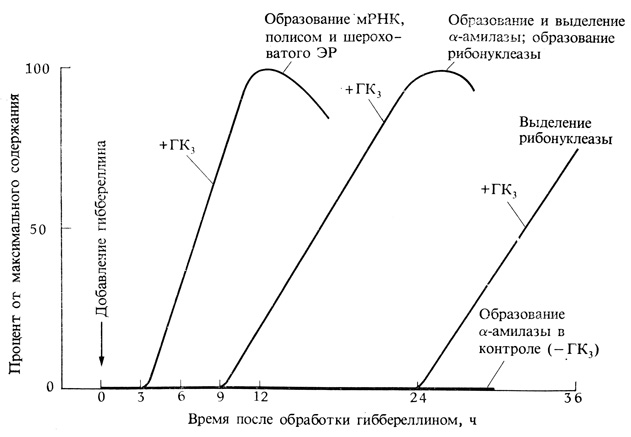
Попытки доказать существование такой специфической мРНК наталкиваются на трудности, связанные с очень малым количеством образующейся мРНК и отсутствием методов, позволяющих отличить ее от других мРНК. Последняя проблема была недавно решена благодаря обнаружению на одном конце молекул мРНК цепи адениновых остатков. Поскольку аденин соединяется водородными связями с уридином, это свойство позволяет отделять мРНК с помощью колонки со связанным полиуридином, к которому может присоединяться аденин. Информационная РНК сорбируется колонкой, тогда как другие РНК свободно проходят через нее. Затем, заменив раствор в колонке, можно элюировать и определить мРНК. Этот метод показал, что примерно через 4 ч после добавления гиббереллина меченые нуклеозиды включаются в мРНК с помощью ядер алейроновых клеток. Это происходит за несколько часов до появления α-амилазы. Кроме того, появление α-амилазы тормозится при добавлении на ранних этапах ингибитора кордицепина, который, как считается, специфически предотвращает завершение синтеза мРНК. Чем позже добавляется кордицепин, тем меньше его эффект. Если его добавить примерно через 12 ч после нанесения гиббереллина, то никакого ингибирующего влияния он уже не будет оказывать. Следовательно, индуцированный гиббереллином синтез мРНК для α-амилазы к этому времени должен был завершиться.
Специфическая природа вновь синтезированной мРНК была окончательно установлена с помощью комбинации изящных методов. После того как выделенную мРНК внесли в белок-синтезирующую систему in vitro, содержащую рибосомы, тРНК, необходимые ферменты и аминокислоты, путем сочетания иммунохимических и электрофоретических методов было показано, что образовавшийся белок идентичен настоящей α-амилазе!
Примерно в то же время, когда появляется мРНК, наблюдается и резкое увеличение числа полисом и шероховатого эндоплазматического ретикулума в алейроновых клетках. Такие изменения типичны для клеток, производящих секретируемые ферменты. Действительно, гиббереллин, по-видимому, способствует как секреции, так и синтезу ферментов. Было показано, что гиббереллин инициирует образование не только α-амилазы, но и других гидролаз, особенно протеазы и рибонуклеазы. Таким образом, один гормон, очевидно, вызывает ряд событий, приводящих к быстрому преобразованию всех запасных питательных веществ семени в вещества, доступные для молодого растения. Гиббереллин способствует также выделению всех этих ферментов из алейроновых клеток в эндосперм. Синтез и высвобождение α-амилазы начинается примерно через 9 ч после добавления гиббереллина (рис. 9.26). Рибонуклеаза синтезируется одновременно с α-амилазой, но до ее выделения из клеток должно пройти более 24 ч с момента добавления гиббереллина. Ферменты расщепляют запасные питательные вещества на растворимые продукты, которые затем транспортируют к растущим апексам растения и используются в качестве источников энергии и материалов, необходимых для образования новых клеток.
Если гиббереллин может дерепрессировать определенные гены в клетках алейронового слоя, не удивительно, что он может также оказывать влияние на деление и дифференциацию клеток в других частях растения путем "включения" других генов. Какие гены при этом включаются, почти наверняка зависит от природы клеток. Сделано не много работ, посвященных роли гиббереллина в инициации или контроле растяжения клеток. У совсем взрослых растений овса гиббереллин отвечает за значительное удлинение междоузлий стебля до цветения. Было установлено, что в отсутствие ауксина такое удлинение полностью обусловлено растяжением клеток, хотя в природных условиях при наличии определенного количества ауксина в узле происходит также и деление клеток. Первоначальные результаты показывают, что гиббереллин индуцирует растяжение клеток благодаря подкислению клеточных стенок примерно таким же образом, как было описано ранее для ауксина. Однако чувствительные к гиббереллину клетки не реагируют на ауксин. Различия между клетками этих двух типов объясняются, вероятно, наличием у них разных рецепторов гормонов.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'