C4-Фотосинтез
В течение ряда лет считалось, что первым акцептором углерода у всех растений служит RuBP и что первыми стабильными продуктами фотосинтеза являются трехуглеродные соединения. Представление это подверглось пересмотру, после того как выяснилось, что в опытах с 14С02 некоторые растения, например кукуруза, сахарный тростник и родственные им тропические злаки, ведут себя необычно: метка включается у них в четырехуглеродные органические кислоты (щавелево-уксусную, яблочную и аспарагиновую) гораздо быстрее, чем в ФГК. Соответственно такие растения стали называть С4-растениями.
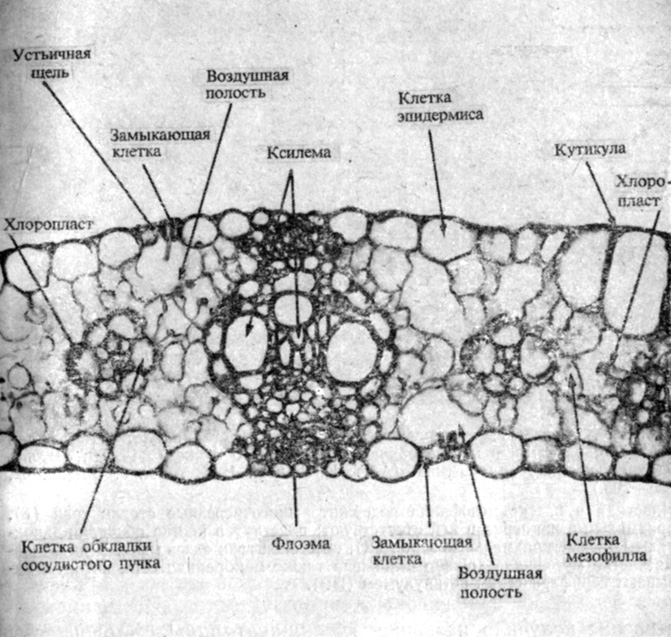
Рис. 4.18. Поперечный срез через лист кукурузы, растения C4-типа. (С любезного разрешения G. Berlin, Yale University.) Обратите внимание на кольцо плотно упакованных клеток обкладки, окружающих каждую жилку, и на рыхло упакованные клетки мезофилла, среди которых в разных местах видны крупные межклетники
Листья многих C4-растений по своей анатомии отличны от листьев C3-растений. Каждый сосудистый пучок окружен у них слоем крупных паренхимных клеток, или так называемой обкладкой сосудистого пучка, которая в свою очередь окружена слоем более мелких клеток мезофилла (рис. 4.18). Подобное строение получило название "кранц-анатомии"*. Хлоропласты в обоих этих типах клеток морфологически различны: в клетках обкладки сосудистого пучка они содержат очень большие крахмальные зерна и часто лишены гран, тогда как в клетках мезофилла в них хорошо выражены граны, но крахмала накапливается очень мало (рис. 4.19). Если в эксперименте разделить эти два типа клеток и измерить в них активность ферментов, та обнаружатся весьма четкие различия. В клетках мезофилла выше активность фосфоенолпируват(ФЕП)-карбоксилазы, катализирующей присоединение С02 к ФЕП с образованием щавелевоуксусной кислоты, а клетки обкладки превосходят клетки мезофилла по активности RuBP-карбоксилазы и других ферментов, принимающих участие в цикле Кальвина - Бенсона. Складывается впечатление, что обе эти группы клеток в С4-растениях кооперативно осуществляют превращение С02 в гексозу и по^ следующее превращение гексозы в крахмал.
* (От нем. Kranz - корона. - Прим. ред.)
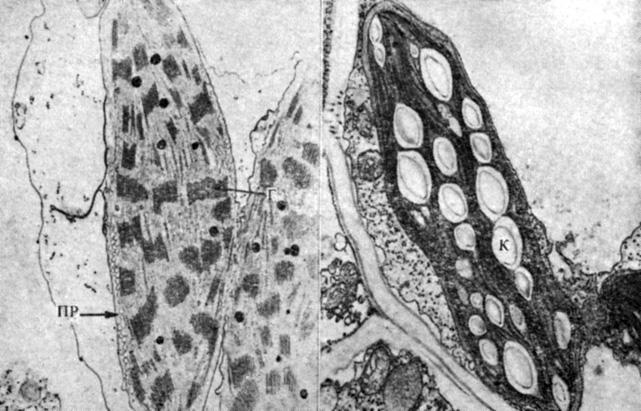
Рис. 4.19. Электронные микрофотографии хлоропластов в клетке мезофилла (X15 500; слева) и в клетке обкладки сосудистого пучка (X14 500; справа) у кукурузы, растения C4-типа. (A. Y. Kirchanski. 1975. Amer. J. Botany, 62, 695-705.) Пластида в клетке мезофилла содержит многочисленные стопки гран (Г), крахмальные же зерна в ней отсутствуют; пластиду в клетке обкладки заполняют крупные крахмальные зерна (К), но стопок гран здесь мало. В цитоплазме клетки мезофилла хорошо выражена также мембранная сеть, которую называют периферическим ретикулумом (ПР)
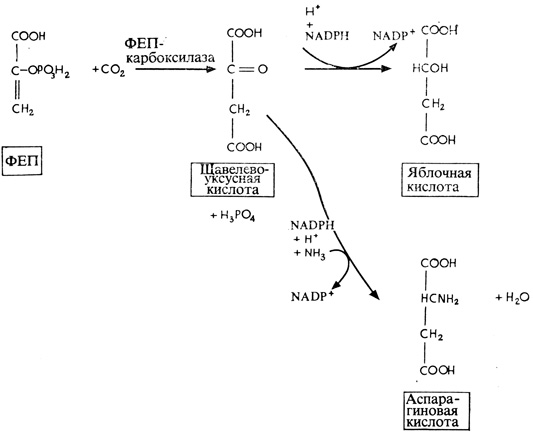
Первая из этих реакций протекает в клетках мезофилла. Здесь ФЕП присоединяет поступающую из атмосферы CO2, в результате чего образуется щавелевоуксусная кислота, которая у одних растений превращается в яблочную кислоту, а у других - в аспарагиновую. Согласно одной из гипотез, яблочная или аспарагиновая кислота диффундирует затем из клеток мезофилла в клетки обкладки и здесь декарбоксилируется с образованием CO2 и трехуглеродного соединения. Последнее снова диффундирует в мезофилл, где происходит регенерация ФЕП, после чего цикл карбоксилирования повторяется с участием новой молекулы CO2, поступившей из атмосферы. Одновременно CO2, высвободившаяся в клетках обкладки, вступает в цикл Кальвина - Бенсона, т. е. реагирует с iRuBP, что приводит к образованию ФГК и других промежуточных продуктов, свойственных C3-растениям, и в конце концов - к гексозофосфатам. Общая схема этих превращений представлена на рис. 4.20. Ясно таким образом, что даже и у C4-растений усвоение углерода в основном происходит при участии RuBP-карбоксилазы; конечная же реакция, приводящая к превращению гексозы в крахмал, протекает у них в клетках обкладки сосудистых пучков.
Мы уже отмечали, что C4-растения осуществляют фотосинтез более эффективно, нежели C3-растения, отчасти вследствие того, что фотодыхание выражено у них слабо и, значит, уже фиксированный углерод они впустую не расходуют. Такое поведение C4-растений, возможно, связано с особенностями их анатомии (с "кранц-анатомией"). Фотодыхание включает образование гликолата и его последующий распад в присутствии кислорода. В C4-растениях конечная фиксация CO2 в цикле Кальвина - Бенсона протекает в клетках обкладки сосудистого пучка, плотно прижатых к соседним клеткам. Считается, что в такой компактной ткани, без межклетников, кислорода очень мало и что именно этот крайне низкий уровень кислорода лимитирует фотодыхание. Ясно также, что при отсутствии межклетников вокруг клеток обкладки затруднен и доступ CO2 к хлоропластам, так что если бы эти клетки функционировали обособленно, они не могли бы эффективно осуществлять фотосинтез. Возможно, что C4-клетки рыхлого мезофилла играют роль коллекторов CO2 и концентрируют ее с помощью ФЕП-карбоксилазы в виде органических C4-кислот. Фермент ФЕП-карбоксилаза действительно способен выполнять эту задачу очень эффективно; он может фиксировать CO2, превращая ее в органические кислоты, при гораздо более низких ее концентрациях, чем это делает RuBP-карбоксилаза. Образовавшиеся таким путем органические кислоты транспортируются к хлоропластам, локализованным в: клетках обкладки. Здесь CO2 высвобождается в высокой концентрации и при сравнительно низком содержании кислорода, благодаря чему хлоропласты этих клеток могут весьма эффективно фиксировать CO2 в виде сахаров через цикл Кальвина - Бенсона. С этой точки зрения C4-фиксация представляется чем-то вроде насоса, поставляющего CO2 для C3-пути. К этому можно добавить, что само положение клеток обкладки создает условия для передачи конечных продуктов фотосинтеза (в частности, сахарозы) непосредственно в ситовидные трубки флоэмы, по которым эти продукты могут затем транспортироваться в другие части растения.
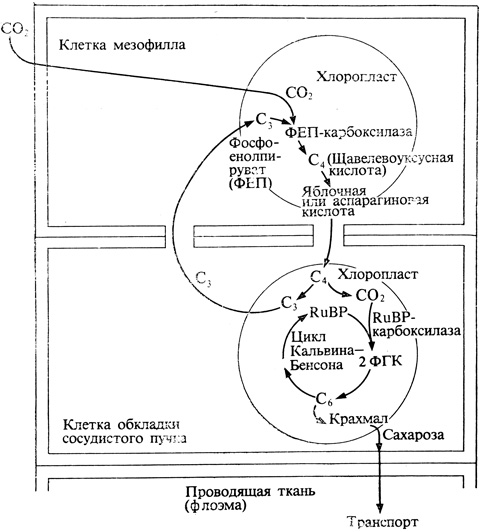
Рис. 4.20. Гипотетическая схема, позволяющая представить себе кооперативное действие клеток мезофилла и обкладочных клеток сосудистого пучка у C4-растений. (С изменениями по U. Ltittge. 1973. Stofftransport der Pflanzen, Berlin and New York, Springer-Verlag.)
Впрочем, возможно, что все это объяснение, хотя и вполне логичное, тем не менее неверно с биологической точки зрения. Как выяснилось недавно, даже и в недифференцированных культурах тканей C4-растений сохраняется низкий уровень фотодыхания. Между тем такие культуры выращиваются из одних только клеток мезофилла и, естественно, не могут обладать кранц-анатомией. Учитывая это обстоятельство, быть может, следует считать, что данное объяснение справедливо в своей химической, но не в структурной части.
Благодаря своей способности к особенно эффективной утилизации CO2 C4-растения могут фиксировать ее при более низких концентрациях, чем это делают C3-растения. Если, например, поместить по одному растению той и другой группы под стеклянный колпак, т. е. в пространство с ограниченным запасом CO2, то C3-растение погибнет от истощения, до которого его доведет C4-растение. Произойдет это потому, что вся CO2, выделяемая C3-растением в процессе фотодыхания, будет немедленно захватываться C4-растением, и C3-растение "додышится", наконец, до полного исчерпания всех своих ресурсов. В табл. 4.2 приведена сравнительная характеристика этих двух групп растений.

Таблица 4.2. Различия между растениями с C4- и C3-циклами первичного фотосинтетического карбоксилирования
(R. G. S. Bidwell. 1974. Plant Physiology, New York, Macmillan.)
Причины появления в процессе эволюции C4-пути не вполне ясны. Среди C4-растений встречается много тропических злаков. Поскольку с повышением температуры фото дыхание резко усиливается, способность ограничивать его в этих условиях, несомненно, можно считать преимуществом. Однако, до тех пор пока сама функция фотодыхания нам не известна, трудно судить о том, почему C4-путь возник и почему он имеется у одних растений и отсутствует у других. В тропических областях и пустынях C3- и C4-растения существуют бок о бок друг с другом и ни тот, ни другой тип не обнаруживает какого-либо преобладания.
Метаболизм органических кислот у растений семейства Crassulaceae (САМ-метаболизм)
Суккуленты, произрастающие в засушливых областях, например Cactus, Kalanchoe и Sedum, также фиксируют атмосферную CO2 с образованием четырехуглеродных соединений. Однако по своему физиологическому поведению эти растения отличаются от других представителей C4-типа. Устьица у них открыты ночью и закрыты днем, обычно же картина бывает обратной: свет стимулирует открывание устьиц, а в темноте они остаются закрытыми. Естественно, что потеря воды на испарение через открытые устьица ночью гораздо меньше, чем днем, т. е. при максимальной температуре и минимальной относительной влажности (см. гл. 6), а потому такой тип поведения представляет несомненную выгоду для растений пустыни. Эти растения поглощают в ночное время атмосферную CO2, образуя в результате ее фиксации четырехуглеродные органические кислоты, главным образом яблочную. Роль первичного акцептора углерода играет у них, как и у прочих C4-растений, ФЕП (трехуглеродное соединение). Днем, когда хлорофилл активируется светом, яблочная кислота декарбоксилируется с образованием трехуглеродного соединения и CO2, из которой затем и строятся шестиуглеродные сахара в цикле Кальвина - Бенсона. Из трехуглеродного соединения регенерируется фосфоенолпируват, который может теперь вновь выступать в роли акцептора CO2. Подобное поведение, т. е. чередование на протяжении суток двух процессов - накопления кислот (в ночное время) и их распада (днем), получило название CAM-метаболизм*, по семейству Crassulaceae, к которому принадлежат многие из таких суккулентов и в котором этот феномен был впервые обнаружен. У САМ-растений первичное карбоксилирование и образование шестиуглеродных сахаров протекают в одних и тех же клетках, но в разное время, тогда как у прочих C4-растений эти процессы происходят одновременно, но могут быть приурочены к разным клеткам. Рис. 4.21 иллюстрирует эти соотношения.
* (От англ. Crassulacean acid metabolism - метаболизм кислот у Crassulaceae. - Прим. перев.)
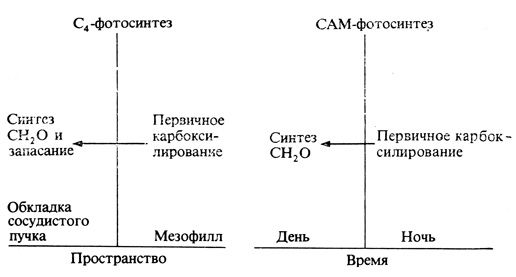
Рис. 4.21. Модель, иллюстрирующая разобщение отдельных этапов фиксации углерода в пространстве (у C4-растений) и во времени (у САМ-растений). (С изменениями по W. M. Laetsch. 1974. Annual Rev. PL Physiol, 25, 27-52.)
Конечные продукты фотосинтеза, гексозофосфаты, переводятся в крахмал для хранения и в сахарозу для транспортировки в другие части растения; кроме того, они превращаются в процессе дыхания в различные строительные блоки, необходимые растению. Все эти процессы мы рассмотрим в следующей главе.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'