Фотодыхание
Растения, использующие только путь Кальвина - Бенсона, принято называть C3-растениями, поскольку первый стабильный продукт фотосинтеза представлен у них трехуглеродным соединением - ФГК. У C3-растений значительная часть фиксированного при фотосинтезе углерода тут же теряется вследствие распада продуктов фиксации и выделения CO2 в реакциях, идущих с потреблением кислорода. Процесс этот происходит только на свету, и потому он был назван фотодыханием. Фотодыхание открыто сравнительно недавно. Объясняется это позднее его открытие тем, что выделение CO2 при дыхании на свету маскируется поглощением CO2 в процессе фотосинтеза. Первоначально предполагалось, что и в количественном отношении, и в смысле используемого пути световое дыхание идентично дыханию в темноте, однако затем выяснилось, что на свету выделяется больше CO2. Это удалось установить в результате тщательных измерений газообмена непосредственно вслед за включением или выключением света. Наблюдающееся на свету дополнительное выделение CO2 объясняется, как выяснилось, не усилением нормального процесса дыхания, а добавлением в этих условиях совершенно иного пути - фото дыхания.
Фотодыхание обусловлено тем, что в присутствии кислорода действующий в цикле Кальвина фермент RuBP-карбоксилаза может присоединять к RuBP не только CO2 , но и O2, выполняя таким образом роль RuBP-оксигеназы. Присоединение кислорода к молекуле RuBP приводит к такому ее расщеплению, при котором (вместо двух молекул ФГК, содержащих по три атома углерода, образуется одна молекула фосфогликолевой кислоты (содержащей два атома углерода) и одна молекула ФГК. Таким образом, в оксигеназной реакции не происходит никакой фиксации CO2. Фосфогликолат позднее дефосфорилируется и превращается в гликолат, который поступает из хлоропласта в другую органеллу, также окруженную мембраной, - в пероксисому (рис. 4.17). В пероксисоме гликолат вступает в реакцию с кислородом, в результате чего образуются глиоксилат и перекись водорода. Перекись тут же распадается на воду и кислород, а глиоксилат превращается в аминокислоту глицин. Затем уже вне пероксисомы, а именно в митохондриях, из глицина образуется аминокислота серин (которая может использоваться непосредственно в белковом синтезе или претерпевать дальнейшие превращения, ведущие к образованию глюкозы). При этой реакции из двух молекул глицина образуется одна молекула серина и одновременно выделяется CO2. Таким образом, какая-то часть углерода, фиксированного в цикле Кальвина - Бенсона, теряется без того, чтобы растение могло хоть как-то этот углерод использовать. Смысл фотодыхания нам пока не ясен, но, может быть, его полезная функция (если таковая существует) связана с тем, что оно играет необходимую роль в метаболизме азотистых соединений или в их переносе из одной органеллы в другую, обеспечивая превращение гликолата в глицин. Возможно также, что фотодыхание возникло на ранних этапах существования Земли с развитием фотосинтеза. В то время в земной атмосфере, очевидно, не было кислорода, поэтому фосфогликолат не мог образовываться под действием RuBP-карбоксилазы. Однако, когда кислород, выделявшийся в процессе фотосинтеза, начал накапливаться в атмосфере, в растениях, возможно, началось накопление фосфогликолата, и, может быть, фотодыхание возникло в процессе эволюции как средство, позволявшее ограничить это накопление.
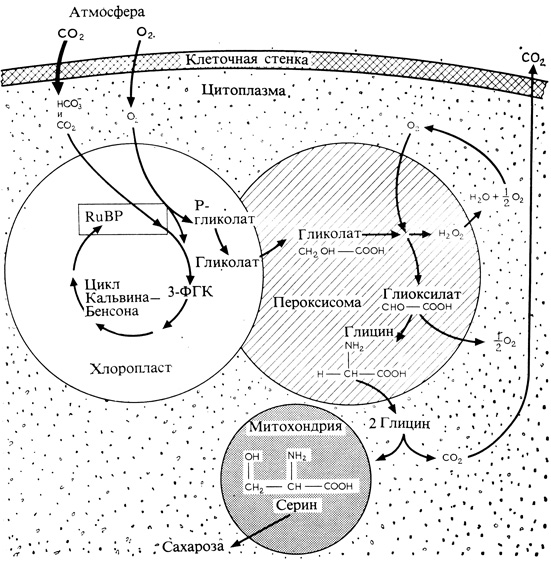
Рис. 4.17. Схема, иллюстрирующая обмен углерода в процессе фото дыхания
Не у всех растений фотодыхание в равной мере интенсивно. Заметно колеблется также и эффективность, с которой разные виды растений в процессе фотосинтеза фиксируют CO2. Интенсивность фотосинтеза у субтропических злаков, например у кукурузы, сахарного тростника и сорго (табл. 4.1), вдвое с лишним выше, чем у шпината, пшеницы, риса и бобов. Растения, более эффективно осуществляющие этот процесс (их называют C4-растениями; о них мы еще будем говорить ниже), используют иной путь усвоения CO2 в обкладках сосудистых пучков листа (так называемый C4-метаболизм), и мы здесь этот путь вкратце обсудим. К менее эффективной группе принадлежат все C3-растения; они могут терять при световом дыхании до половины всего углерода, ассимилированного в процессе фотосинтеза.

Таблица 4.1. Средняя урожайность некоторых травянистых культура
(С изменениями по I. Zelitch. 1971. Photosynthesis, Photorespiration and Plant Productivity, New York, Academic Press.)
Возможность регулировать фотодыхание представляет большой интерес для физиологов растений, поскольку урожаи некоторых культур можно было бы, очевидно, удвоить, если бы только удалось каким-то образом уменьшить эти потери потенциальных резервов растения. Попытки такого рода ведутся в разных направлениях. Исследуется, например, зависимость фотодыхания от экспериментальных условий с тем, чтобы, изменяя эти условия там, где это возможно, сводить таким путем фотодыхание к минимуму. Высокая концентрация CO2, низкая концентрация кислорода и низкая интенсивность света ограничивают фотодыхание, чем, кстати, отчасти и объясняется тот факт" что "удобрение углекислотой" повышает скорость роста у многих растений. Испытываются также химические ингибиторы фотодыхания, которые не должны быть токсичными ни для самих растений, ни для питающихся ими животных. Селекционеры разыскивают варианты или мутанты растений с низкой интенсивностью фотодыхания и пытаются включить этот признак в генотип создаваемых сортов. Следует, однако, помнить, что подавление фотодыхания может у некоторых растений иметь вредные последствия. Недавно, например, в опытах по изучению роста растений сои, отличающихся высоким уровнем фотодыхания, выяснилось, что хотя при низких концентрациях кислорода фотодыхание снижается и растения вегетативно развиваются лучше, чем в обычной атмосфере, они не образуют зрелых семян, если концентрация 02 оказывается ниже 5%. Отсюда можно заключить, что у некоторых растений фотодыхание или какие-то сопутствующие ему реакции необходимы для того, чтобы жизненный цикл протекал нормально. Действительно, трудно представить себе, как фотодыхание могло бы возникнуть в процессе эволюции и сохраниться на всем ее протяжении, если бы оно было лишено какой бы то ни было приспособительной ценности.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'