Хлоропласты и другие пластиды
Уникальной особенностью растительных клеток является присутствие в них ряда разнообразных цитоплазматических телец, так называемых пластид. Среди этих пластид важнейшую роль в зеленой растительной клетке играют хлоропласты (рис. 2.19 и 2.20) - центры фотосинтетической активности, в которых сосредоточены весь хлорофилл и все вспомогательные пигменты, связанные с фотосинтезом.
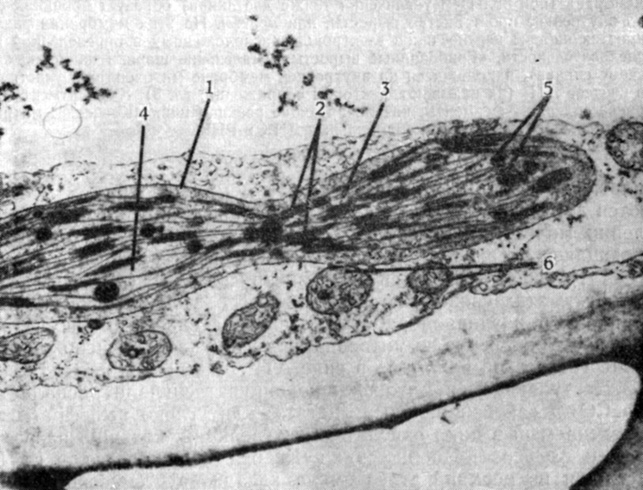
Рис. 2.19. Хлоропласт Samanea saman, находящийся, по-видимому, в процессе деления. (С любезного разрешения Morse M. J., Yale University.) Видны две мембраны, образующие оболочку (1) хлоропласта; граны (2), т. е. стопки тилакоидов; мембраны стромы (3), связывающие между собой граны; строма (4) и многочисленные жировые капельки (5). В цитоплазме, окружающей хлоропласт, видно несколько митохондрий (6). (X16 000.)
У высших растений хлоропласты по форме напоминают линзу. Диаметр их равен 5-8 мкм, а толщина - приблизительна 1 мкм. Каждый хлоропласт окружен двойной мембраной (обладающей избирательной проницаемостью) и содержит также сложную внутреннюю систему мембран. Основная структурная единица хлоропластов - тилакоид - представляет собой тонкий,, плоский мешочек, ограниченный однослойной мембраной. В нем находятся хлорофилл, вспомогательные пигменты и ферменты, принимающие участие в фотохимических реакциях фотосинтеза. Тилакоиды собраны в группы наподобие стопки монет. Эти стопки называются гранами. Все пространство между гранами заполнено бесцветной стромой, в которой содержатся многие ферменты, участвующие в фиксации CO2. В клетке имеется в среднем около 50 хлоропластов, причем каждый хлоропласт развивается, как полагают, из пропластиды. По-видимому, пропластиды способны реплицироваться путем какого-то деления (именно так и увеличивается их число в клетке); зрелые хлоропласты также иногда проявляют способность к репликации, однако это наблюдается реже. У этиолированных (выросших в темноте) растений пропластиды называются этиопластами.
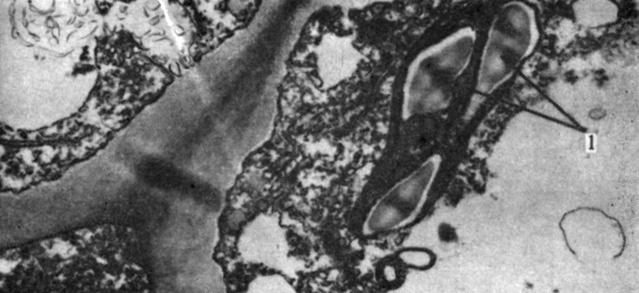
Рис. 2.20. Хлоропласт Samanea, содержащий несколько крупных крахмальных зерен (1). (X16 000.) (С любезного разрешения Morse M. J., Yale University.)
Будучи мельче хлоропластов, пропластиды не обладают и свойственной хлоропластам ламеллярной (слоистой) структурой. Вместо этого они содержат проламеллярное тело - упорядоченный, "паракристаллический" центр каналов, которые после надлежащей стимуляции светом изменяют свою ориентацию и превращаются в параллельные слои (рис. 2.21). У цветковых, или покрытосеменных, растений зрелые хлоропласты могут развиться из пропластид только при освещении, тогда как у некоторых голосеменных это превращение завершается и в полной темноте. Другое различие между большинством голосеменных и покрытосеменных связано с их относительной способностью превращать пигмент протохлорофилл в хлорофилл, т. е. осуществлять реакцию, в которой к молекуле протохлорофилла присоединяются два атома водорода и остаток фитола ("фитольный хвост") (рис. 2.22). У всех покрытосеменных для этого превращения необходим свет, а у некоторых голосеменных оно может происходить и в темноте. В тканях корня хлоропласты обычно не развиваются даже на свету, однако у некоторых растений, в частности у моркови и у ипомеи, клетки корня могут на свету зеленеть. Вопрос о том, почему пластиды в цитоплазме некоторых клеток не достигают зрелости, остается пока открытым.
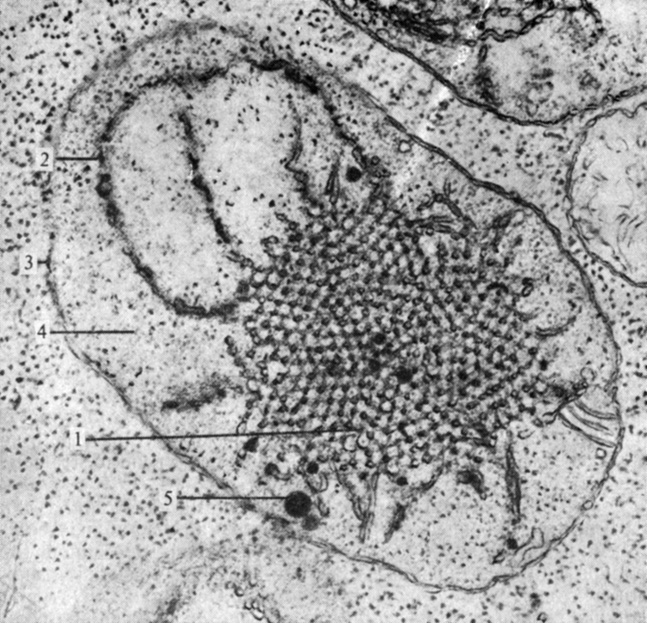
Рис. 2.21. Этиопласт (пропластида) из первичного листа 10-дневного растения гороха, выращенного в темноте. (С любезного разрешения Hurkman W. J., Purdue University.) Отметьте высокоупорядоченную структуру проламеллярного тела (1), из которого при освещении должны развиться граны. Видно, что в левом верхнем углу одна грана (2) уже начала развиваться. Видны также две мембраны оболочки (3) пластиды, строма (4) и жировая капелька (5). (X44 000.)
Хлоропласты содержат свою, специфическую, ДНК, которая отличается от ядерной ДНК и передается по наследству через пропластиды, находящиеся в цитоплазме материнской клетки (яйцеклетки); от отцовского растения (через пыльцевую клетку) хлоропластная ДНК не наследуется. Содержащиеся в хлоропласте ферменты кодируются либо ядерной, либо хлороиластной ДНК, некоторые же из них, например рибулозобисфос-фаткарбоксилаза, состоят из двух белковых субъединиц, из которых одна кодируется ядерной ДНК, а другая - ДНК пластиды. В хлоропластах содержатся также рибосомы, РНК, аминокислоты и ферменты, необходимые для синтеза белка. Все это придает хлоропластам известную автономность, т. е. делает их в какой-то мере независимыми от остальных клеточных, структур. Поэтому некоторые биологи полагают, что хлоропласты - это потомки каких-то посторонних организмов, которые когда-то случайно внедрились в незеленые клетки и тем самым сделали их автотрофными (способными самостоятельно создавать необходимые им питательные вещества путем фотосинтеза). Согласно этой гипотезе, фотосинтезирующие одноклеточные организмы, лишенные окруженных мембраной органеля (прокариоты), некогда случайно объединились с гетеротрофными организмами (неспособными к фотосинтезу и, следовательно, зависящими в своем питании от предобразованных питательных веществ). В эволюционном смысле это оказалось выгодным, и потому такой симбиотический (взаимовыгодный) союз сохранился в виде современной эукариотической растительной клетки, имеющей окруженное мембраной ядро и другие органеллы. В пользу такой точки зрения свидетельствуют результаты исследований ультраструктуры клеток. В этих исследованиях выявилось разительное сходство между хлоропластами высших растений и сине-зелеными водорослями, у которых нет ни ядра, ни окруженных мембраной хлоропластов, а имеются только пронизывающие цитоплазму ламеллы (рис. 2.23). Митохондрии также содержат специфическую ДНК и аппарат белкового синтеза, а следовательно, и они, возможно, являются потомками каких-то свободноживущих организмов. Отмечено,, что митохондрии обнаруживают известное структурное сходство с некоторыми примитивными бактериями, а их ДНК в химическом отношении родственна ДНК прокариотических организмов.
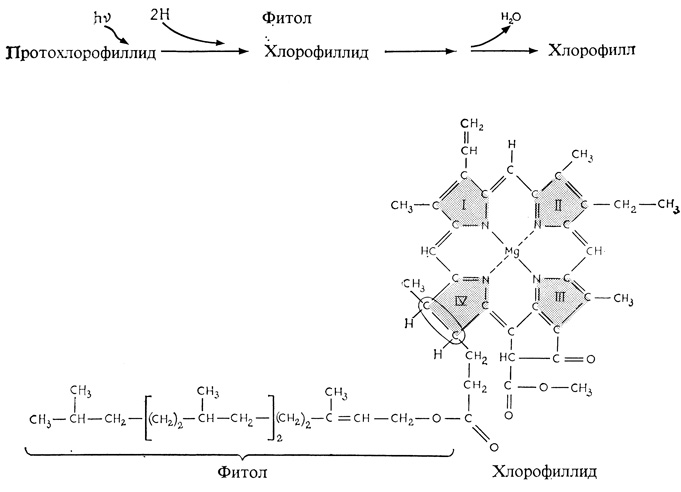
Рис. 2.22. У покрытосеменных к протохлорофиллиду под действием света присоединяются два атома водорода, в результате чего он превращается в хлорофиллид. Последующее присоединение остатка фитола переводит хлорофиллид в хлорофилл. Внизу показана структура хлорофилла a. Обратите внимание на атом магния, занимающий центральное положение, и на присоединенные к нему четыре пиррольных кольца (I-IV). Эта структура напоминает гем, у которого в центре находится атом железа, также соединенный с четырьмя пиррольными кольцами. В протохлорофилле группа -C-C- в кольце IV (отмечена овальной рамкой) заменена группой -C=C-
Сторонники гипотезы, о которой идет речь, указывают, что если, например, одноклеточное жгутиковое Eugtena выращивать на протяжении нескольких поколений при довольно высоких температурах, то размножение самих клеток будет опережать размножение их хлоропластов, так что клетки будут постепенно все больше и больше бледнеть. В конце концов в результате такого "разбавления" образуются совсем бесцветные клетки, в которых не будет уже ни одного хлоропласта и ни одной пропластиды. Эти клетки так и останутся незелеными, навсегда утратившими способность к автотрофии. Можно также вызвать исчезновение хлоропластов, воздействуя на клетки стрептомицином и некоторыми другими веществами. Иными словами, клетки можно "вылечить" от "заселивших" их хлоропластов с помощью либо тепловой терапии, либо химиотерапии.
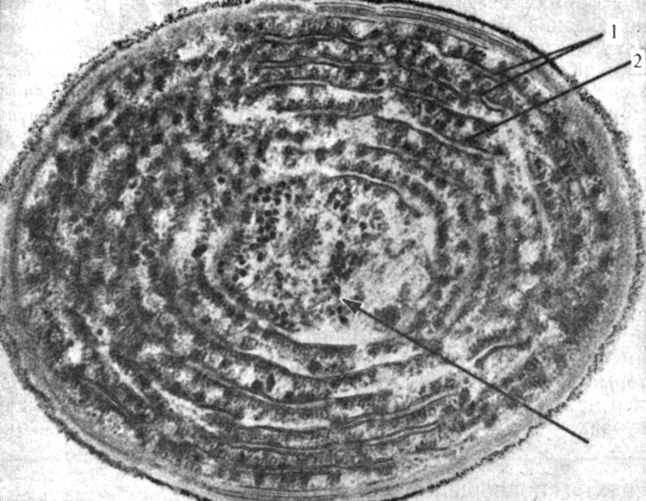
Рис. 2.23. Электронная микрофотография сине-зеленой водоросли Synechococcus lividus. (Edwards M. R. опубликована в статье Gantt E. 1975. BioScience, 25, 781-788.) Видно, что фотосинтетические ламеллы (1) и строма (2) распределены по всей клетке. На ламмелах находятся дисковидные фикобилисомы (показаны стрелкой), содержащие несколько различных пигментов. (X54 000.) Обратите внимание на сходство между этой клеткой и хлоропластом высшего растения (рис. 2.19)
Биологи научились выделять из клеток интактные хлоропласты методом дифференциального центрифугирования. Можно показать, что эти хлоропласты в течение некоторого времени сохраняют все свойства, присущие фотосинтетическому аппарату клетки. Изолированные интактные хлоропласты на свету фиксируют CO2, высвобождают O2 и генерируют высокоэнергетические фосфатные связи. Однако вне клетки хлоропласты не в состоянии ни поддерживать свое собственное существование, ни размножаться. Если они действительно присутствуют в клетке как "захватчики", некогда проникшие в нее извне, то приходится признать, что существование их теперь в большой мере зависит от остальных клеточных структур.
В хлоропластах локализуются различные пигменты. Самый важный из них, хлорофилл, - единственный пигмент, непосредственно участвующий в фотосинтезе, - встречается в нескольких формах, слегка отличающихся одна от другой по спектрам поглощения. У некоторых водорослей хлоропласты особенна богаты фикобилинами - синим или красным, а в хлоропластах большинства высших растений присутствуют окрашенные в желтый, оранжевый или красный цвет каротиноиды. По-видимому, каротиноиды на свету защищают хлорофилл от разрушающего действия молекулярного кислорода. Кроме того, они повышают эффективность фотосинтеза, поглощая и передавая хлорофиллу энергию тех длин волн, которые хлорофиллом не поглощаются, благодаря чему эта энергия может все же использоваться для фотосинтеза. На протяжении почти всего вегетационного периода каротиноиды в листьях незаметны, поскольку их окраска маскируется высокой концентрацией хлорофилла; осенью же, когда концентрация хлорофилла в стареющих листьях снижается, ярко окрашенные каротиноиды выступают на первый план, так что осенняя окраска листвы зависит в основном именно от них. В какой-то мере, правда, на нее влияют и некоторые другие пигменты, находящиеся вне хлоропластов, например ярко-красные антоцианы, присутствующие в клеточных вакуолях.
Спектры поглощения и пигменты
Пигменты представляют собой соединения, способные в силу своей электронной структуры поглощать излучение определенной длины волны в видимой части спектра. График, описывающий зависимость поглощения от длины волны, носит название спектра поглощения. Спектр поглощения данного соединения может быть уникальным, т. е. он может однозначно характеризовать это соединение, но если даже это не так, то он все же позволяет нам составить хотя бы некоторое представление о структуре поглощающего вещества.
Для того чтобы исследовать спектр поглощения какого-либо соединения, требуется монохроматический свет. Его получают при помощи призмы, дифракционной решетки или же светофильтра. Затем световой пучок пропускают через раствор изучаемого пигмента и сравнивают прошедшую через раствор световую энергию с энергией падающего пучка света. Поглощение энергии зависит от коэффициента поглощения (α) пигмента, от его концентрации (c) и от длины оптического пути (l). Эту зависимость - ее часто называют законом Бэра-Ламберта - можно записать в следующем виде:
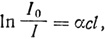
где I0 - энергия падающего света, а I - энергия света, прошедшего через раствор. Для растворов обычно пользуются десятичным логарифмом и вместо коэффициента поглощения (α) используют коэффициент экстинкции (ε). Мы можем теперь написать
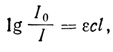
где c - концентрация в моль/л, I - оптический путь в см и ε - коэффициент молярной экстинкции в л/моль/см. Величину lgI0/I - называют поглощением (А), или оптической плотностью (OD).
В спектрофотометре осуществляются получение монохроматического светового пучка, пропускание его попеременно через раствор изучаемого вещества и через чистый растворитель, измерение I0 и I и регистрация логарифма их отношения. Типичный спектр поглощения хлорофилла b в органическом растворителе представлен на рис. 2.24. Этот пигмент окрашен в зеленый цвет, потому что он поглощает в синей (400-450 нм) и красной (600-700 нм) областях спектра, но пропускает зеленые и желтые лучи.
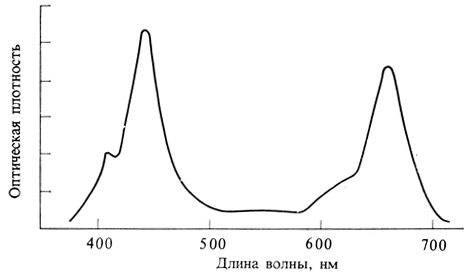
Рис. 2.24. Спектр поглощения хлорофилла b в органическом растворителе
В клетках высших растений помимо хлоропластов содержатся еще и другие типы пластид, лишенные характерных для хлоропластов ламеллярной структуры и фотосинтетического аппарата. Среди этих пластид различают бесцветные лейкопласты и окрашенные хромопласты, окраска которых определяется высокой концентрацией каротиноидных пигментов. Подобно хлоропластам, эти тельца, по-видимому, передаются по наследству через какие-то структуры типа пропластид, присутствующие в материнской цитоплазме. Лейкопласты играют в клетке роль хранилищ для запасных питательных веществ, таких, например, как крахмал, а потому, вероятно, у них имеется и ферментный аппарат, необходимый для синтеза этих веществ из более мелких молекул-предшественников. Хромопласты, по всей вероятности, способствуют опылению растений и распространению семян, так как они придают цветкам и плодам их яркую окраску, привлекающую животных. Как правило, хромопласты в клетке не развиваются, если в ней имеются хлоропласты. В некоторых зреющих плодах, например в томатах, переход от зеленой окраски к желтой, а затем к красной отражает три последовательные стадии развития: преобладание хлоропластов, убыль хлоропластов и рост числа хромопластов, нагруженных каротиноидами. Причины этого перехода не вполне ясны.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'