Порядок норичниковые (Scrophulariales)
Семейство пасленовые (Solanaceae) (Г. А. Денисова)
В семействе около 90 родов и не менее 2500 видов, широко распространенных в тропических, субтропических и умеренных областях, главным образом в Центральной и Южной Америке.
Представители семейства - травы, кустарники или небольшие деревья с очередными (иногда в области соцветия супротивными), простыми листьями. Цветки обычно в пазушных верхоцветных соцветиях, обоеполые, актиноморфные или реже слегка зигоморфные. Чашечка обычно 5-лопастная или 5-раздельная, остающаяся, часто при плодах увеличенная. Венчик от колесовидного до трубчатого, 5-лопастный, редко двугубый. Тычинок обычно 5 или в зигоморфных цветках меньше (4-2); пыльники вскрываются продольно или верхушечными порами. Нектарный диск обычно развит. Гинецей обычно из 2 плодолистиков, редко из 5 плодолистиков, обычно с верхушечным простым столбиком с двулопастным рыльцем; завязь обычно двугнездная (иногда ложно-3 или 5-гнездная) или редко 5-гнездная, обычно с многочисленными семязачатками. Плод - ягода или септицидная коробочка, редко плод распадающийся (рис. 218). Семена с эндоспермом.
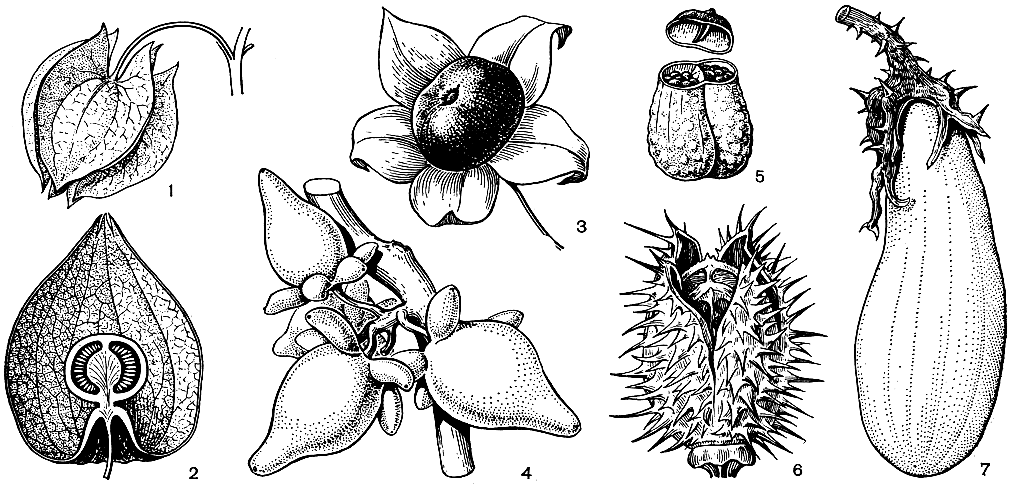
Рис. 218. Типы плодов пасленовых: 1 - никандра физалисовидная (Nicandra physalodes), ягода в пузыревидной крылатой чашечке; 2 - физалис обыкновенный (Physalis alkekengi), продольный разрез ягоды в пузыревидной некрылатой чашечке; 3 - красавка обыкновенная (Atropa belladonna), ягода; 4 - паслен сосочковый (Solanum mammosum), ягода с выростами; 5 - белена черная (Hyoscyamus niger), коробочка с открывающейся крышечкой; 6 - дурман обыкновенный (Datura stramonium), коробочка, раскрывающаяся створками; 7 - баклажан (Solanum melongena), ягода
Цветки пасленовых опыляются различными насекомыми, а в тропических странах также птицами, а иногда даже млекопитающими.
Семейство пасленовых подразделяют на 2 подсемейства - нолановые (Nolanoideae) и собственно пасленовые (Solanoideae). Первое нередко рассматривают как самостоятельное семейство нолановые (Nolanaceae). Подсемейство нолановых относительно более примитивно, чем пасленовые. Оно включает 2 близких рода - нолана (Nolana, около 75 видов, распространенных от Перу до Патагонии и на Галапагосских островах) и алона (Alona, 5-6 видов в Чили). Это травы или маленькие кустарнички с очередными, цельными, более или менее суккулентными листьями, произрастающие главным образом вдоль морских побережий. Гинецей у них из 5 плодолистиков.
Все остальные роды семейства пасленовых входят в обширное подсемейство пасленовых. Оно, в свою очередь, подразделяется на 5 триб. Самой примитивной трибой считается триба никандровые (Nicandreae), состоящая из одного монотипного рода никандра (Nicandra), обитающего в Перу и Боливии. Это однолетняя трава с грубыми ребристыми жестко опушенными стеблями, зубчатыми или лопастными листьями, одиночными цветками, 3-5-гнездной завязью и довольно многосемянной ягодой, заключенной в сильно разрастающуюся при плодах чашечку. Никандра физалисовидная (N. physalodes) широко распространилась как сорное растение во многих районах нашей страны на бахчах, виноградниках, по огородам и у дорог, проникнув на Кавказ, в оазисы Средней Азии и на Дальний Восток. Изредка ее разводят с лекарственными целями и как декоративное растение. У никандры имеется интересное приспособление против самоопыления. В эфемерных цветках этого растения рыльце увядает уже в течение часа после опыления и столбик опадает.
Самой большой трибой семейства пасленовых является триба собственно пасленовых (Solaneae), куда входит несколько десятков родов. Завязь у них 2-гнездная, редко многогнездная (например, у томата). Триба, в свою очередь, подразделяется на несколько подтриб, из которых самая примитивная - подтриба дерезовых. Сюда входят деревья, кустарники или травы. Наиболее известный и самый крупный род дереза (Lycium) заключает около 100 видов листопадных или вечнозеленых прямостоячих или вьющихся, обычно колючих кустарников, распространенных в тропических, субтропических и отчасти умеренных областях, главным образом в Южной Америке. В нашей стране 7 видов в пустынях, полупустынях и степях на юго-востоке европейской части, на Кавказе и в Средней Азии. Некоторые виды разводят для изгородей и как декоративные.
К подтрибе дерезовых относится и такой широко известный род, как красавка, или атропа (Atropa), состоящий из 4 видов, распространенных от Европы и Средиземноморья до Индии. Наиболее известна красавка обыкновенная, или белладонна (A. belladonna, табл. 56),- многолетнее травянистое растение с высоким стеблем, цветками большей частью с буро-фиолетовым или грязно-пурпуровым (иногда желтым) венчиком и многосемянной черной (иногда желтой) блестящей ягодой. Белладонна - ценное лекарственное растение, введенное в культуру. Все части растения содержат алкалоиды (атропин, гиосциамин и др.) и ядовиты. Привлекательные на вид ягоды красавки иногда являются причиной отравления детей.

Таблица 56. Пасленовые: 1 - красавка обыкновенная (Atropa bella-donna), Крым
В трибе пасленовых отдельную подтрибу составляют также роды скополия (Scopolia) и белена (Hyoscyamus). Крупные многолетние травы с утолщенным корневищем. Из 6 видов скополии, распространенных от Средней и Южной Европы до Индии, Тибета и Японии, в нашей стране в дикорастущем состоянии обитает лишь один вид - скополия карниолийская (S. carniolica) в широколиственных лесах на западе европейской части и на Кавказе. Во всех частях, но главным образом в корневище и корнях содержит алкалоиды гиосциамин и скополамин. Другие виды - скополия светло-желтая (S. lurida), обитающая в Гималаях, и скополия тангутская (S. tangutica), произрастающая в Тибете, содержат, кроме того, алкалоид атропин. Все 3 вида возделывают как ценные лекарственные растения.
В роде белена (Hyoscyamus) около 20 видов, произрастающих на Канарских островах, в Европе, Северной Африке (до центральной части), Западной и Средней Азии. В СССР - 8 видов, из которых наибольшее значение имеет белена черная (H. niger), которую в европейской части СССР заготавливают как лекарственное сырье. Введена в культуру. Белена содержит алкалоиды гиосциамин, скополамин, атропин и некоторые другие. Семена этой белены по форме и цвету сходны с семенами мака и в примеси к нему вызывают отравление. Опыление у белены перекрестное. Размеры его грязно-желтых, воронковидных венчиков, обладающих фиолетовыми пятнами, указывающими дорогу к нектару, как раз соответствуют размерам более крупных шмелей. Перекрестное опыление обеспечивается тем, что рыльце расположено выше пыльников.
Подтриба пасленовых занимает центральное положение в одноименной трибе семейства. Здесь нужно прежде всего упомянуть довольно большой род физалис (Physalis), около 100 видов которого широко распространены в тропических, субтропических и отчасти умеренных областях, преимущественно в тропической Америке. Физалисы характеризуются очень крупной пузыревидно вздутой при плодах, красной или оранжевой чашечкой. У физалиса обыкновенного (P. alkekengi) чашечка при плодах особенно велика и ярка, поэтому это растение пользуется большой популярностью как декоративное. Плоды некоторых видов, как физалиса обыкновенного и физалиса перуанского (P. peruviana), вполне съедобны и отличаются своеобразным и приятным вкусом.
К трибе пасленовых относится также и такое общеизвестное культурное растение, как овощной перец (Capsicum annuum). В ботанической литературе число видов рода капсикум колеблется от 20 до 50. К сожалению, систематика этого рода находится в таком хаотическом состоянии, что число его видов невозможно определить. Виды капсикума (в литературе обычно называемого "овощным перцем", хотя далеко не все его виды овощные растения) дико произрастают в Центральной и Южной Америке и на Галапагосских островах, а также на полуострове Флорида. Это небольшие кустарники, полукустарники или многолетние травы (в культуре обычно используемые как однолетние) с многосемянными красными, оранжевыми или желтыми, иногда беловатыми или коричневатыми плодами различной формы. В тропических странах широко культивируют несколько видов капсикума, из которых обыкновенный овощной перец, или паприка (C. annuum), выращивают как однолетнее растение в странах с умеренным и субтропическим климатом; в нашей стране- на Украине, в Молдавии, на Кавказе и в Средней Азии. Его исходная дикорастущая многолетняя разновидность распространена на полуострове Флорида, в Вест-Индии, Мексике, Центральной Америке и Колумбии. Острота вкуса плодов рода капсикум зависит от фенольного летучего вещества капсаицина. Особенно высоким является содержание капсаицина в плодах капсикума кустарникового (C. frutescens), точное место происхождения которого остается неизвестным. Приправа, получаемая из плодов этого вида, известна под названием соуса табаско. Капсикум кустарниковый возделывают в Центральной Америке, Мексике и в южных областях США. Плоды видов рода капсикум широко использовали в Америке еще ацтеки, которые повседневно сдабривали пищу мелко истолченными сухими плодами. Колумб после первого же путешествия привез капсикум из Вест-Индии. В России первое упоминание об овощном перце содержится в рукописи "Благопрохладный цветок, или Травник" (1616).
Но, безусловно, самым важным для человека является самый большой в семействе род паслен (Solanum), насчитывающий около 1700 видов, т. е. больше половины видового состава всего семейства. Он широко распространен в тропических, субтропических и умеренных областях обоих полушарий, но главным образом в Южной Америке. Это многолетние, реже однолетние травы, полукустарники с прямостоячими или вьющимися стеблями, иногда небольшие деревья. Плод 2-гнездная многосемянная ягода. В нашей стране около 20 дикорастущих видов паслена. В их числе паслен сладко-горький (S. dulcamara) - лазящий полукустарник, большей частью с лиловыми цветками и ярко-красными ягодами. Встречается почти по всей европейской части и на юге Западной Сибири. Другой, еще более распространенный у нас вид,- это паслен черный (S. nigrum) - однолетник с белыми цветками и черными (редко зелеными) ягодами, который растет как сорняк в огородах и садах и на сорных местах. Оба вида содержат алкалоид соланин и поэтому ядовиты и вызывают отравление у человека и домашних животных. К роду паслен относится ряд важнейших для человека культурных растений. Первое место среди них занимает картофель (от немецкого слова Kartoffel). В культуре известно в основном 2 близких вида - картофель андийский (S. andigena), издавна возделываемый на территории Колумбии, Эквадора, Перу, Боливии и Северо-Западной Аргентины, и наш обыкновенный картофель клубненосный (S. tuberosum), называемый также чилийским. Родина его - Средний Чили и прилегающие острова (включая остров Чилоэ). Этот вид получил очень широкое распространение в странах с умеренным климатом. Местное население горных районов Южной Америки выращивает также и некоторые другие виды. Все виды картофеля относятся к секции туберариум (Tuberarium) рода паслен, насчитывающей вместе с дикими клубненосными видами около 200 видов, произрастающих преимущественно в Южной и Центральной Америке. Культурные виды картофеля размножаются клубнями (в селекционной работе также семенами). Введение картофеля в культуру (сначала путем эксплуатации диких зарослей) началось примерно 14 тыс. лет назад индейцами Южной Америки. В Европу (Испанию) картофель впервые был ввезен около 1565 г., откуда распространился по другим странам. В Россию картофель впервые попал в XVII в., но начало широкой культуре картофеля положил указ Сената в 1765 г. и завоз из-за границы партии семенного картофеля, разосланного по стране.
Интересен процесс опыления у картофеля. Пять его тычинок, сложенных вместе в конус, плотно прилегают к торчащему выше их столбику, головчатое рыльце которого несколько наклонено вниз. Пыльники при сотрясении высыпают небольшое количество пыльцы. При посещении цветка насекомые сначала задевают наклоненное вниз рыльце, и если на них уже была пыльца от другого цветка, то опыляют его. Но так как цветки картофеля посещают лишь сравнительно немногие насекомые, то обычно происходит самоопыление. Самоопыление это совершается благодаря тому, что рыльце выпрямляется и становится как раз на ту линию, по которой опадает пыльца.
Другим очень важным культурным представителем рода паслен является баклажан, или бадриджан (S. melongena). Это многолетнее травянистое растение с высоким стеблем, крупными листьями, фиолетовыми цветками и более или менее округлыми, грушевидными или цилиндрическими плодами. Плоды желтые, с коричневыми полосами, белые, зеленые или фиолетовые. Плоды баклажана жарят, тушат, маринуют, из них готовят баклажанную икру, соте и пр. В диком виде баклажан произрастает в Индии и Бирме. Баклажан впервые был введен в культуру в Индии, откуда его культура распространилась в другие страны, в частности в Китай. Еще за 500 лет до нашей эры в Китае культивировалась мелкоплодная форма баклажана.
Из культивируемых пасленов следует упомянуть еще наранхиллу (naranjilla), или лало (Solanum quitoense), "золотой плод Анд". Специалисты считают, что наранхилла имеет большую будущность, хотя в настоящее время мало кто знает об этом замечательном растении за пределами Колумбии и Эквадора. Это исключительно вкусный десертный плод, который используют также для приготовления желе, джема и п других целей. Свежевыжатый сок этих плодов используют в Эквадоре и Колумбии для приготовления "сорбета" (sorbete) - зеленого, пенящегося напитка с привлекательным кисло-сладким вкусом ананаса и клубники. В Панаме, Гватемале и Коста-Рике, где это растение было интродуцировано, свежий сок превращают в замороженный концентрат. Растение представляет собой кустарник высотой 1-2 м, с опушенными листьями и округлыми, желто-оранжевыми плодами, покрытыми легко удаляемыми белыми волосками. В благоприятных условиях растение плодоносит в течение всего года.
Среди видов обширного рода паслен есть целый ряд других культурных представителей.
К роду паслен очень близко стоит и многими ботаниками объединяется с ним томат, или помидор (Lycopersicon esculentum), культивируемый во множестве сортов во всех частях света. В роде томат около 7 видов, обитающих на Тихоокеанском побережье Южной Америки (Колумбия, Эквадор, Перу и Чили) и на Галапагосских островах. На местном языке науатль это растение называется томати (tomati), но при его интродукции в XVI в. в Испанию, Португалию его стали называть "золотым яблоком" (porno d'oro - отсюда "помидор").
В обширную трибу пасленовых, но в качестве отдельной подтрибы входят южноамериканский род цифомандра (Cyphomandra) и средиземноморско-азиатский род мандрагора (Mandragora). В роде мандрагора около 6 видов, распространенных от Пиренейского полуострова до Восточных Гималаев и Тибета. Большинство видов мандрагоры представляют собой многолетние травы, почти всегда бесстебельные, с очень крупными листьями в розетке, достигающей в диаметре 1-2 м и более. Мясистые, богатые крахмалом корни мандрагор обладают своеобразным ветвлением: подчас корень дает два вертикальных отростка и несколько напоминает фигуру человека. Из-за этой своей особенности мандрагора с древнейших времен овеяна легендами, приписывающими ей магическую силу. Кроме того, она содержит алкалоид гиосциамин и в средние века считалась одним из ценнейших лекарственных снадобий.
В Западном Копетдаге, в Туркменской ССР, был открыт новый вид - мандрагора туркменская (M. turcomanica). Здесь вегетация этого растения начинается с осени, с началом сезона дождей, когда оно развивает розетку крупных листьев. Цветение в условиях теплых зим Западного Копетдага происходит в начале ноября и продолжается до середины апреля. Опыляются цветки различными насекомыми. Созревание плодов наблюдается с мая до конца июня. С наступлением жаркого сухого периода растение как бы замирает и сбрасывает засохшие листья. Зрелые плоды мандрагоры туркменской крупные (диаметром до 5-6 см), оранжевые, ароматные и вполне съедобные.
В отличие от бесстебельных средиземноморских видов мандрагоры гималайско-тибетская мандрагора стеблевая (M. caulescens) имеет развитый стебель и более мелкие листья. В то же время ближайший родственный ей вид - мандрагора тибетская (M. tibetica) - представляет собой мелкое плотнорозеточное растение. Как можно предположить, в данном случае приспособительная эволюция шла по пути фиксации ювенильной фазы вегетативной сферы.
Далее в системе подсемейства пасленовых следует небольшая триба дурмановых (Datureae), характеризующаяся тем, что в результате развития двух ложных перегородок, разделяющих каждую из двух плацент первоначально двугнездной завязи, завязь становится как быче-тырехгнездной. Плод у дурмановых - коробочка или ягода. Наиболее известным представителем этой трибы является род дурман (Datura), насчитывающий около 10 видов многолетних или однолетних трав, обитающих в тропических и тепло-умеренных странах, главным образом в тропической Америке. Цветки у видов дурмана крупные, с белым воронковидным венчиком длиной от 6 до 20 см и более. Все они отличаются крупными длиннотрубчатыми цветками с дурманящим запахом и опыляются преимущественно длиннохоботковыми ночными бабочками, которые отовсюду летят на запах дурмана, пренебрегая другими растениями. В процессе эволюции удлинение трубки венчика у многих представителей рода шло параллельно с увеличением длины хоботка некоторых бабочек бражников (Sphingidae). В результате возникли виды дурмана, которые могут опыляться лишь определенными видами бражников, попадая в полную от них зависимость. Плоды дурманов - оригинальные шиповатые коробочки, раскрывающиеся четырьмя створками. Эти ядовитые растения содержат ряд алкалоидов и находят применение в фармакопее, а как анестезирующие были известны еще древним перуанцам. Местами виды дурмана культивируют в качестве декоративных. У нас в стране наиболее широко известен однолетний дурман обыкновенный, или вонючий (D. stramonium), встречающийся как рудеральное растение на пустырях, огородах, у изгородей и строений почти по всей европейской части СССР. Возделывается как лекарственное растение на юге Украины и в Краснодарском крае. Все части растения содержат ядовитые алкалоиды (гиосциамин, скополамин, атропин и др.). В южных районах СССР культивируются дурман безвредный (D. innoxia) родом из Америки и дурман индийский (D. metel) родом из Юго-Западного Китая, незрелые плоды которых содержат скополамин.
К дурману очень близок и нередко объединяется с ним южноамериканский род бругмансия (Brugmansia, табл. 56), состоящий из 5 видов кустарников или небольших деревьев с очень крупными длиной до 30 см кожистыми листьями. Огромные яркие трубчатые цветки, висящие на длинных цветоножках, утром раскрываются, а днем, в жару, снова захлопываются, что в полной мере используют для себя виды дрозофилы. Мушки собираются в венчике сразу после его раскрывания. Несмотря на то что дрозофилы находятся в цветке поблизости от пыльников, они, по-видимому, участия в опылении не принимают. По истечении нескольких дней венчик цветка, населенного мушками, опадает и вместе с ним падают на землю подросшие личинки дрозофил, продолжая питаться оставшейся в венчике пыльцой. Вероятно, плодовые мушки завершают свой жизненный цикл вместе с цветком приютившей их особи. Г. Карсон, который наблюдал определенные виды дрозофил, поселяющиеся в цветках бругмансии белоснежной (B. Candida), считает, что эти виды плодовой мушки могут существовать только в сожительстве с бругмансией. Опыляются цветки более крупными насекомыми, способными проникать в цветок снизу вверх. Удлиненные мягкие или несколько деревянистые нераскрывающиеся плоды бругмансии содержат крупные клиновидные семена. В качестве декоративного растения широко известен "древовидный дурман", представляющий собой гибрид между бругмансией белоснежной и бругмансией древесной (B. arborea, табл. 56).

Таблица 56. Пасленовые: 2 - бругмансия древесная (Brugmansia arborea), Батумский ботанический сад
Из других представителей трибы следует отметить чрезвычайно интересный род соландра (Solandra), около 10 видов которого распространены в тропической Америке. Большинство соландр представляет собой длинно- и толстостебельные лианы, обитающие в тропических горных лесах и обвивающие крупные деревья. Взбираясь по их стволам на большую высотулиана распластывает на верхушке кроны дерева простые цельнокрайние кожистые листья и подставляет солнцу крупные красивые, слегка зигоморфные колокольчатые цветки на толстых цветоножках. Многосемянные ягоды растений, свободно охваченные разросшейся яркой чашечкой, хорошо заметны сверху на поверхности кроны дерева-хозяина и поэтому охотно поедаются птицами, которые и разносят на дальние расстояния их почковидные мелкие семена.
Некоторые особи соландры в преклонном возрасте почти или совершенно теряют связь с почвой, продолжая функционировать как полуэпифиты или как настоящие эпифиты. Наиболее красиво цветущие виды рода культивируют в тропических садах и оранжереях как декоративные. Особенно популярна соландра крупноцветковая (S. grandiflora), оплетающая живым ковром заборы и стены домов.
Интересно, что опыление некоторых представителей трибы дурмановых осуществляется не только различными насекомыми, но также и летучими мышами. Это прослежено для рода трианея (Trianaea), 3 вида которого распространены в горных лесах Северных Анд. У этих растений довольно крупные цветки располагаются на концах длинных висячих ветвей и выделяют обильный нектар. Цветки раскрываются вечером, испуская резкий неприятный запах, привлекающий летучих мышей.
Триба цестровых (Cestreae) получила наименование по наиболее заметному роду цеструм (Cestrum, табл. 56), объединяющему около 150 видов, обитающих в тропических и субтропических областях Америки. Это кустарники или небольшие деревья с цельными, большей частью узкими, часто опушенными листьями. Их цветки, собранные в верхушечные или пазушные соцветия с маленькой чашечкой и длинным воронковидным или трубчатым венчиком, открываются в основном по ночам, издавая сильный запах. Особенно отличается этим цеструм ночной (C. nocturnum), названный у себя на родине "ночным жасмином". Этот кустарник, повсеместно выращиваемый в тропических садах, образует огромное количество мелких зелёновато-белых до кремовых цветков, которые по ночам испускают очень приятный и сильный аромат, привлекая к себе ночных насекомых-опылителей. Л. Оверланд (1960), изучая механизм открывания и закрывания цветков "ночного жасмина", пришел к выводу, что вещества, обусловливающие их сильный ночной запах, локализованы в паренхимных клетках верхушки лепестка. Возникновение запаха является результатом не наступления темноты, как можно было бы предполагать, а следствием внутреннего ритма растения, не зависящего от внешних условий. Открывание и закрывание молодых цветков в течение суток синхронизируется с циклом появления и исчезновения запаха: широко открытые (по ночам) цветки пахнут, закрытые (днем) - не пахнут. Со старением цветка цикличность постепенно сбивается; совсем же старые, уже оплодотворенные цветки и не пахнут, и не закрываются. Некоторые виды цеструма, как цеструм полевой (C. campestre), опыляются колибри. Плоды цеструмов - продолговатые ягоды.

Таблица 56. Пасленовые: 3 - цеструм изящный (Cestrum elegans), Батумский ботанический сад
К данной трибе принадлежит также и род табак, или никотиана (Nicotiana), насчитывающий 66 видов, из которых 45 обитают во вне-тропических частях Америки, а 21 вид приурочен к Австралии и Полинезии.
Это в основном однолетние травы, изредка многолетние травянистые растения, но встречаются также кустарники. Цветки с трубчатым воронковидным или колокольчатым крупным венчиком. В опылении табака в Америке участвуют колибри. Плод - яйцевидная, 2-4-створчатая коробочка, наполненная многочисленными мелкими сетчато-точечными семенами. Растения с тяжелым неприятным запахом. Многие виды рода содержат никотин и другие токсичные алкалоиды. Во всех странах,где обитают дикорастущие табаки, неоднократно наблюдались случаи отравления их листьями и молодыми побегами домашних животных.
В культуре наиболее известны табак настоящий (N. tabacum) и табак махорка (N. rustica). Листья махорки используют не только для курения, но они также являются основным сырьем для получения лимонной кислоты, никотина и никотиновых медицинских препаратов - никотиновой кислоты (витамин РР), никотин-сульфата, который эффективно применяют для борьбы с сельскохозяйственными вредителями. Табак выращивали в Америке задолго до открытия ее европейцами. В Европу табак завезли в конце XV и в первой половине XVI в. и сначала выращивали как декоративное и лекарственное растение.
К табакам близок род петуния (Petunia), представленный около 30 южноамериканскими видами; некоторые из них повсеместно культивируются как декоративные. У нас наиболее известна петуния гибридная (P. hybrida).
Крайне своеобразен относящийся к цестровым род маркея (Markea), 18 видов которого обитают по преимуществу в лесах тропической Америки - от бассейна реки Амазонки до Мексики. Некоторые виды маркеи представляют собой лианы с длинными изящными стеблями и простыми цельнокрайними листьями, собранными в пучки на концах веточек. Обвиваясь вокруг опоры, стебли маркеи вползают по деревьям все выше и выше, образуя в верхней части кроны дерева-хозяина терминальные соцветия с бархатистыми короткотрубчатыми актиноморфными, часто пурпурно-зелеными цветками. С. Фогель в 1958 г. проследил, что цветки некоторых видов маркеи, такие, как маркея дресслери (M. dressleri), закрыты в дневное время и открываются только по ночам, привлекая своим запахом местных лесных крыс, которые и являются основными опылителями этих растений. Другие виды рода ведут эпифитный образ жизни, что особенно характерно для маркеи курчавой (М. ulei). Этот мелкий кустарник с малозаметными невзрачными цветками встречается в густых зрелых лесах от Панамы до Перу. Его особи способны поселяться как в тенистой кроне дерева, так и на верху кроны, под открытым палящим солнцем. Тонкие воздушные корни эпифитных растений, покрытые рыхлой оболочкой, а также водопроницаемые молодые стебли и листья интенсивно впитывают в себя из воздуха и влагу, и кислород, и углекислый газ, и даже минеральные вещества, которые попадают в воздух вместе с мельчайшими частицами почвы. Эпифиты усиленно развиваются в сезоны дождей, накапливая в своих рыхлых тканях воду, которую используют затем в засушливое время года. Этими своеобразными сосудами влаги и питательных веществ испокон веков пользуются муравьи, устраивая внутри стеблей некоторых видов маркеи долгосрочные убежища. Симбиоз с муравьями выгоден и кустарнику, поскольку муравьи не только защищают свой живой дом от насекомых-вредителей и других внешних врагов, но также привносят на субстрат вместе с экскрементами питательные вещества для растения.
Очень близок к роду маркея как в систематическом отношении, так и по образу жизни род хуануллоа (Juanulloa). Растения, принадлежащие к этому роду, отличаются от видов рода маркея своими узкими трубчатыми опушенными цветками. Среди 12 видов хуануллоа, распространенных от Мексики до Боливии, имеются не только эпифиты, но даже и полупаразиты, живущие на деревьях. Обладая вполне развитыми и нормально функционирующими зелеными листьями, полупаразитные растения образуют гаустории, которые постепенно проникают в древесину молодых ветвей дерева-хозяина, высасывая из сосудов воду с растворенными в ней минеральными солями.
Наиболее подвинутой трибой семейства является триба сальпиглоссовых (Salpiglossideae). Растения, принадлежащие к ней, часто обладают зигоморфными цветками с 2-4 фертильными тычинками и септицидными раскрывающимися плодами типа коробочки с одревесневшей наружной эпидермой.
Одним из примечательных родов трибы является схизантус (Schizanthus) - эндемичный чилийский род, насчитывающий около 10 видов. От остальных пасленовых он отличается ясно выраженной двугубостью крайне зигоморфного венчика. Из 5 тычинок только 2 фертильные, 2 превращены в стаминодии, а пятая очень редуцирована. Интересен механизм опыления цветков: две фертильные тычинки располагаются внутри губы, образованной двумя нижними лопастями венчика. Когда пчела или бабочка опускается на эту губу, пыльники энергично взрываются, выстреливая пыльцу в воздух. Некоторые из пыльцевых зерен обычно попадают на насекомое, которое переносит их к другому цветку. Некоторые виды этого рода издавна культивируют как декоративные. Особенно популярен схизантус перистый (S. pinnatus, рис. 219) - многолетнее железисто опушенное растение с ажурными листьями и многочисленными пестроокрашенными цветками, называемый "цветок-бабочка" или "орхидея бедняков". В культуре выведен целый ряд форм, отличающихся окраской лепестков.
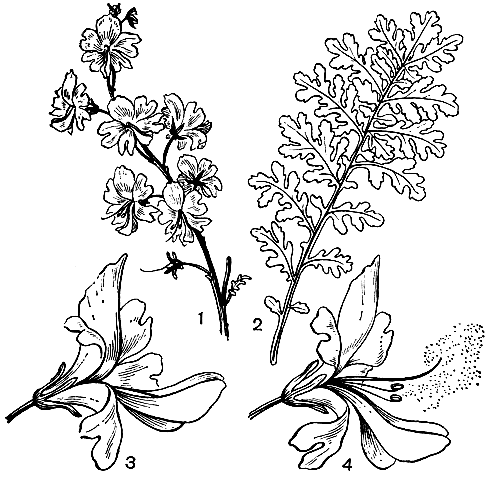
Рис. 219. Схизантус перистый (Schizanthus pinnatus): 1 - соцветие; 2 - лист; 3 - цветок; 4 - момент рассеивания пыльцы
Другой, заслуживающий внимания род сальпиглоссовых - брунфелъсия (Brunfelsia) - состоит из 40 видов, произрастающих в тропической Америке. Это широко распространенные в тропической зоне декоративные деревья или кустарники, цветущие в сезон дождей и названные поэтому "дождевыми деревьями". Листья брунфельсии цельнокрайние, кожистые, расположенные чаще на концах веточек, иногда собранные в пучки. Цветки слегка зигоморфные, синие, пурпурные или белые, с 4 фертильными тычинками. С возрастом цветки меняют окраску, как у бурачниковых. Плоды большинства видов - сочные или кожистые ягоды, окруженные слегка разросшейся чашечкой и содержащие крупные призматические семена с мясистым эндоспермом. Некоторые из видов весьма декоративны.
Род салъпиглоссис (Salpiglossis), давший название трибе, в количестве 5 видов обитает в Южной Америке. Салъпиглоссис выемчатый (S. sinuata) встречается как декоративное в культуре.
Семейство будлеевые (Buddlejaceae) (А. Е. Бородина)
Семейство будлеевых (10 родов и около 170 видов) распространено в тропических и субтропических странах обоих полушарий. Наиболее крупный род будлея (Buddleja), названный в честь английского ботаника Адама Будла (1660-1715), включает около 100 видов, распространенных в Азии, Африке и Америке. Остальные роды, иногда включаемые в будлею, олиготипны и монотипны и ограничены в распространении.
Обычно будлеевые входят в состав кустарниковых зарослей по берегам рек и на опушках лесов. Многие будлеи растут в мезофильных лесах, иногда поднимаются в горы до высоты 2000-3500 м, но некоторые виды встречаются в пустынных и полупустынных районах, например на юге Африки, в Мексике и на юго-западе США. Здесь будлеи маленькие кустарники, в высоту не достигающие 1 м и часто размножающиеся побегами от корневища. Большинство других будлеевых - более крупные кустарники или небольшие деревья, реже высокие деревья, достигающие 20-30 м, крепкие деревянистые лианы, редко встречаются и полукустарники (например, некоторые виды будлеи). Листья супротивные, мутовчатые, редко очередные, цельные, зубчатые или лопастные. Для будлеевых характерно опушение из звездчатых (плоских или на ножке) или канделябровидных волосков, густо покрывающих нижнюю сторону листа, побеги, чашечки цветков, а иногда и наружную сторону венчика. Цветки одиночные или в различного рода верхоцветных или бокоцветных соцветиях, обоеполые, актиноморфные или слабозигоморфные: чашечка 4-лопастная, венчик трубчатый, 4-лопастный, редко 5-лопастный (сананго - Sanango), обычно черепитчатый, тычинок 4, прикрепленных к трубке венчика; гинецей из 2 плодолистиков, с головчатым или коротко-2-лопастным рыльцем; завязь верхняя или редко полунижняя, 2-гнездная, очень редко 4-гнездная (аденоплея - Adenoplea) с многочисленными семязачатками на толстых плацентах. Плод обычно коробочка, редко костянка или ягода (никодемия - Nicodemia). Семена многочисленные, часто крылатые.
Будлеевые - насекомоопыляемые растения. Насекомых привлекает нектар, выделяемый на дне их ярко окрашенного венчика. У будлей цветки распускаются летом на побегах этого года, но некоторые цветут весной до появления листьев (будлея тибетская - В. tibetica), и лишь немногие, в их числе будлея Делавэ (В. delavayi), цветут с ранней весны до поздней осени. В солнечные дни на цветках будлей можно встретить множество сидящих бабочек, и поэтому во многих странах они известны под названием "куст бабочек". Бабочек привлекает резкий сладкий запах, исходящий из цветка, и обильный нектар на дне длинной и узкой трубки, доступный лишь их хоботку. Хилиантус древовидный (С. arboreus), имеющий короткую трубку венчика, опыляется жуками и мухами.
Ягоды и костянки будлеевых расселяются птицами и животными. Семена, освобождающиеся из сухих плодов, распространяются воздушными потоками. У будлей коробочка растрескивается по средней жилке, обнажая центральную плаценту и освобождая множество очень мелких шаровидных или удлиненных семян. Часто семена имеют тонкие прозрачные крылья-летучки, постепенно суженные к концам, так что семя в целом имеет веретеновидную форму и легко подхватывается ветром.
Применение будлеевых разнообразно. Виды будлей в цветущем состоянии очень декоративны благодаря множеству мелких желтых, оранжевых, белых или пурпуровых цветков, собранных в кистевидные или головчатые соцветия, поэтому большинство будлей являются прекрасными оранжерейными и садовыми растениями. В СССР на Южном берегу Крыма и на Черноморском побережье Кавказа культивируют около 10 видов будлей.
Семейство норичниковые (Scrophulabiaceae) (Н. Н. Цвелев)
Норичниковые занимают как бы центральное положение в порядке того же названия (Scrophulariales) не только потому, что это семейство крупнейшее по числу видов, но и потому, что оно показывает, настолько близкое родство со многими другими семействами порядка, что границу между ними и отдельными группами норичниковых установить часто очень трудно. Как и во многих других, преимущественно внетропических семействах цветковых растений, среди норичниковых значительно преобладают многолетние и однолетние травы, а кустарников и кустарничков в нем сравнительно мало.
Вьющиеся стебли имеет мексиканская травянистая лиана родохитон темно-красный (Rhodochiton atrosanguineus). Черешки листьев и цветоножки мексиканских лиан из рода маурандия (Maurandya) обладают способностью, подобно усикам, закручиваться вокруг ветвей соседних деревьев и кустарников (рис. 220).
Кустарниковые норичниковые редко достигают большой высоты и обычно имеют облик кустарничков. Среди последних имеются верескоподобные (эрикоидные) жизненные формы, ветви которых покрыты очень мелкими прижатыми к ним листьями, иногда напоминая ветви кипариса или подобных ему хвойных, например новозеландская хебе кипарисовидная (Hebe cupressoides). У коровяка колючего (Verbascum spinosum) с острова Крит конечные веточки растопыренно-разветвленных ветвей превращены в колючки.
Травянистые норичниковые отличаются особенно большим разнообразием жизненных форм. Однолетники составляют около 30% всех видов, причем некоторые из них, например многие виды очанки (Euphrasia), имеют высоту в несколько сантиметров. Из многолетников-ксерофитов можно отметить распространенную в степях и полупустынях Евразии додартию восточную (Dodartia orientaJis). Ее листья быстро опадают, и во время цветения все растение имеет своеобразный прутьевидный облик. Имеется несколько гидрофильных видов, к которым принадлежит, в частности, разводимая в аквариумах лимнофила разнолистная (Limnophila heterophylla). Этому виду свойственна характерная для многих водных растений разнолистность: подводные листья рассечены на линейные или нитевидные доли, а возвышающиеся над водой - цельные, зубчатые. Карликовое бесстебельное норичниковое лужайник водяной (Limosella aquatica) может развиваться как на дне неглубоких водоемов под водой, так и на влажных местах вне воды (рис. 221).
Крайне своеобразно другое "земноводное" норичниковое - хамегигас неустрашимый (Chamaegigas intrepidus), обитающий в небольших западинках с песчаным или каменистым дном на пологих склонах гранитных гор в Юго-Западной Африке. В сухое время года (часто более 10 месяцев) эти места прогреваются более чем до +50 °С. В это время от особей хамегигаса остаются лишь совершенно высохшие розетки видоизмененных листьев пирамидальной формы (см. рис. 220). После выпадения дождей, когда вода заполняет западинки, эти листья быстро набухают, а затем в течение немногих дней из розеток вырастают удлиненные побеги с розетками широкоэллиптических плавающих листьев и цветками в их пазухах. Но вот временный водоем начинает пересыхать, и от хамегигаса вновь остаются лишь видоизмененные прикорневые листья с запасами питательных веществ для быстрого развития в следующий дожливый сезон.
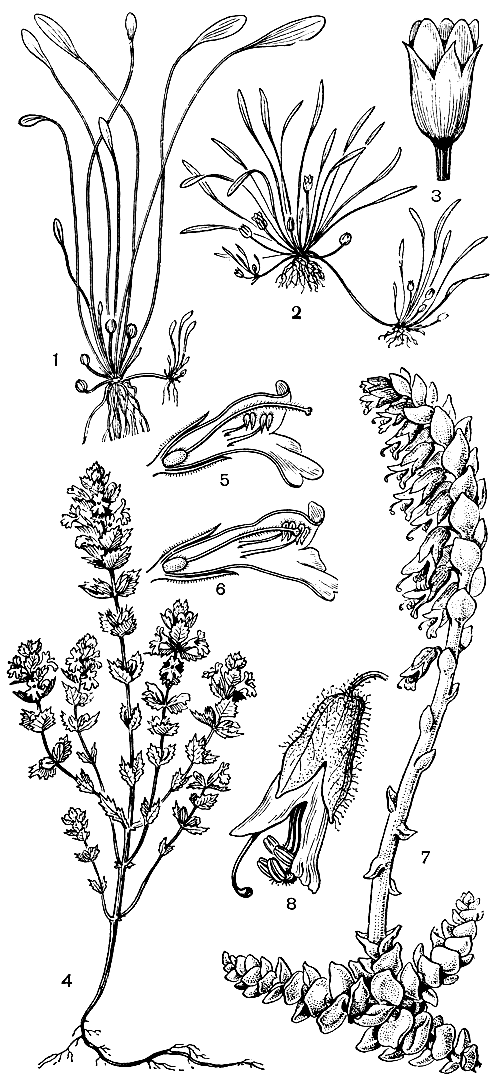
Рис. 220. Норичниковые. Цимбалария постенная (Cymbalaria muralis): 1 - общий вид растения; 2 - цветок в разрезе. Маурандия вечноцветущая (Maurandya semperflorens): 3 - общий вид растения. Хамегигас неустрашимый (Chamaegigas intrepidus): 4 - общий вид растения во время цветения; 5 - общий вид растения в засушливое время года
Многие норичниковые имеют приспособления для вегетативного размножения: длинные разветвленные корневища (например, у аврана лекарственного - Gratiola officinalis), стелющиеся и укореняющиеся в узлах надземные побеги (у цимбаларии постенной - Cymbalaria muralis), корневые отпрыски (у льнянки обыкновенной - Linaria vulgaris, табл. 57). У лужайника из пазух листьев розетки образуются длинные побеги, подобные усам земляники и несущие на верхушке молодые розетки листьев.

Таблица 57. Норичниковые: 1 - тетранема розовая (Tetranema roseum), оранжереи Ботанического института АН СССР в Ленинграде

Таблица 57. Норичниковые: 5 - льнянка обыкновенная (Linaria vulgaris), Московская область
Очень интересно, что в семействе норичниковых можно видеть переход от обычных, автотрофных растений к полупаразитам, а затем и к высокоспециализированным полным паразитам. К паразитирующим на корнях других растений, по-видимому, принадлежат все представители триб жерардиевых (Gerardieae) и погремковых (Rhinanth&ae). Значительное большинство их - полупаразиты, имеющие зеленые листья и обладающие способностью к фотосинтезу. Из корней других растений они получают лишь воду и растворенные в ней вещества с помощью корневых присосок - гаусториев. Некоторые из них, например виды зубчатки (Odontites), имеют на корнях корневые волоски и способны развиваться при отсутствии других растений, хотя в этом случае развиваются лишь очень слабые особи. Проростки видов из многих других родов полупаразитных норичниковых при отсутствии растений-хозяев дальше не развиваются и погибают. Как правило, хозяевами полупаразитов могут быть виды из самых различных семейств, хотя некоторых видов они явно избегают, например калужницу болотную (Caltha palustris). Довольно часто гаустории образуются при соприкосновении корней разных особей одного и того же полупаразита и даже разных корней одной и той же особи.
К более высокоспециализированным полупаразитам принадлежит распространенная на горных лугах Европы тоцция (Tozzia). Дальнейшее развитие ее зародыша имеет место только после соприкосновения зародышевого корня с корнем растения-хозяина и образования на нем гаустория. Сначала тоцция ведет себя как полный паразит, формируя под землей довольно толстое корневище, покрытое мясистыми чешуями - видоизмененными листьями, накапливающими в себе запасы питательных веществ. Затем образуется надземный побег с развитыми фотосинтезирующими листьями и цветками.
Тоцция представляет как бы переход от полупаразитов к полным корневым паразитам, подобным петрову кресту (Lathraea squamaria). Виды этого последнего рода паразитируют на корнях деревьев и кустарников (обычно орешника, ольхи и граба), причем их развитие начинается, как и у тоцции, с образованием толстого корневища, покрытого мясистыми чешуевидными листьями. Обычно петров крест 10- 15 лет ведет подземный образ жизни: его корневище сильно разрастается и ветвится (часто крестообразно), образуя все новые и новые гаустории. Только после этого на поверхности почвы появляются окрашенные в красноватый цвет односторонние соцветия на толстых ножках, лишенных зеленых листьев (рис. 221, 7, 8). Период основного развития (в том числе и цветение) петрова креста приходится на весну - время максимального восходящего сокодвижения у его "хозяев", когда в соке особенно много органических веществ, используемых паразитом.
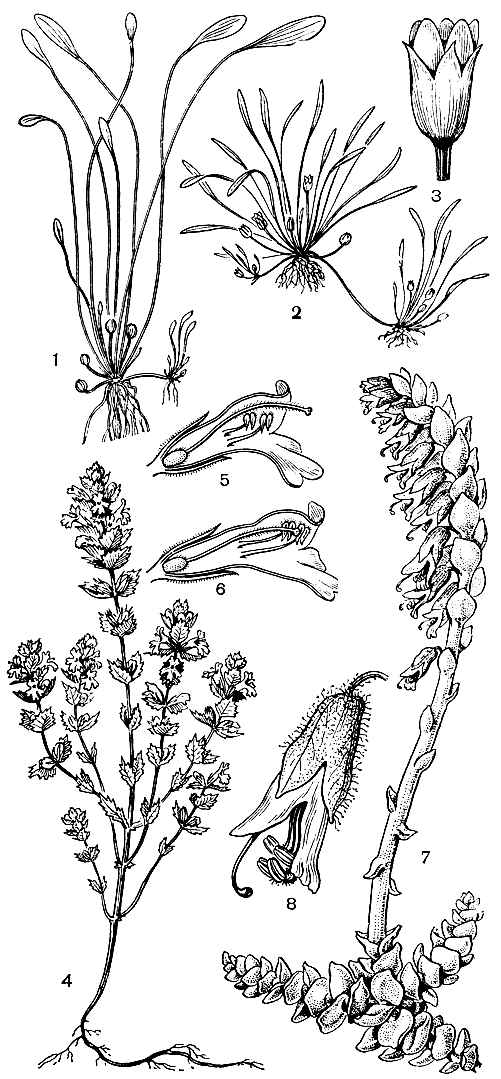
Рис. 221. Норичниковые. Лужайник водяной (Limosella aquatica): 1 - подводная форма растения с клейстогамными цветками; 2 - наземная форма растения с хазмогамными цветками; 3 - хазмогамный цветок. Очанка Ростковиуса (Euphrasia rostkoviana): 4 - общий вид растения; 5 - цветок в начале цветения; 6 - цветок в конце цветения. Петров крест (Lathraea squamaria): 7 - общий вид растения; 8 - цветок
Норичниковые-паразиты и полупаразиты всасывают много воды из корней растений-хозяев, и для ее выделения на нижней поверхности листьев обычно имеются специальные выделяющие воду органы - водяные устьица, или гидатоды. Подземные чешуевидные листья тоцции и петрова креста также имеют большое количество гидатод на своей нижней стороне, но в особых полостях, образованных завернутыми на нижнюю сторону краями чешуи и обычно содержащих много бактерий и мелких насекомых. Прежде эти полости даже принимали за ловушки для насекомых и других обитающих в почве мелких животных, но в настоящее время доказано, что ни насекомоядность, ни сожительство с бактериями в этом случае не имеют места.
Листья норичниковых не имеют прилистников и обычно расположены супротивно или очередно, реже мутовчато. Форма их может быть очень различной, но преобладают листья с черешками и цельными пластинками. У леукокарпуса пронзеннолистного (Leucocarpus perfoliate) из Южной Америки листья полустеблеобъемлющие, а у некоторых видов коровяка (Verbascum) стеблевые листья сидячие и крыловидно низбегающие на стебель.
Почти всегда обоеполые и обычно пятичленные цветки норичниковых могут быть собраны в верхушечные или пазушные соцветия с прицветничками или без них, но довольно часто располагаются и по одному в пазухах не видоизмененных в прицветники листьев. Совершенно правильных (актиноморфных) цветков среди них нет, и вообще в этом семействе значительно преобладают сильно зигоморфные двусторонне-симметричные цветки. Изредка образующиеся на верхушке кистевидных или колосовидных соцветий актиноморфные цветки (например, у льнянки обыкновенной цветки с 5 одинаково развитыми лопастями венчика и 5 шпорами) являются результатом ненормального развития одного цветка из зачатков нескольких цветков. Способность к образованию таких цветков - пелорий - была отмечена еще К. Линнеем именно у представителей семейства норичниковых. Обычно имеющиеся в соцветиях прицветники в одних случаях могут совсем исчезать, в других они являются дополнительным средством привлечения насекомых-опылителей. Так, у многих видов марьянника (Melampyrum) синие или фиолетовые прицветники хорошо контрастируют с желтыми венчиками цветков (отсюда народное название этих видов иван-да-марья).
Чашечка в пределах семейства варьирует от раздельнолистной, как у наперстянки (Digitalis), до длиннотрубчатой двугубой или покрывалообразной (глубоко вырезанной с одной стороны). Первично пятичленная чашечка часто переходит в четырехчленную или за счет редукции ее верхней лопасти, или за счет срастания 2 нижних лопастей. У погремка (Rhinanthus) немного вздутые чашечки значительно увеличиваются в размерах после цветения, а у мексиканского родохитона очень крупные колокольчатые красные чашечки служат дополнительным средством привлечения опылителей - колибри.
Подобно чашечке, венчик норичниковых может иметь блюдцевидную форму при очень короткой трубке (например, у коровяка), но гораздо чаще он бывает длиннотрубчатым, что связано с приспособлением к определенным опылителям. При двугубом венчике верхняя губа обычно образована 2 лопастями, а нижняя - 3, но у немезии (Nemesia) верхняя губа составлена 4 лопастями, а нижняя - одной. У алонсоа (Alonsoa) цветоножки перекручиваются на 180° и нижняя губа становится как бы верхней. Довольно часто в цветках норичниковых развиваются дополнительные выросты, выпуклины или вогнутости. Так, в нижней части трубки венчика близ ее основания часто имеется выпуклость, переходящая в мешкообразный вырост (у львиного зева - Antirrhinum) или шпору (у льнянки, рис. 222). У некоторых видов южноафриканской диасции (Diascia) - 2 рядом расположенные шпоры. Нередко развивается выпуклина на нижней губе, прикрывающая собой зев цветка, например у львиного зева и льнянки. Шмели и крупные пчелы, опыляющие такие цветки, легко их открывают, в то время как для других, более мелких насекомых путь к нектару закрыт. Подобное же маскообразное строение венчика достигается у коллинсии (Collinsia) и губастика (Mimulus) за счет выпуклины на верхней губе. Очень сложное строение имеют венчики видов обширного рода мытник (Pedicularis, табл. 57), у которых верхняя губа часто оттянута в обращенный вниз клювик, а нижняя губа нередко сдвинута на одну из боковых сторон (см. рис. 222). У видов американского рода калъцеолария (Calceolaria) вся нижняя губа, очень сильно развитая, превратилась в направленное вниз пузыревидное выпячивание (см. рис. 222), совершенно сходное с "башмачкообразной" нижней губой орхидеи венерина башмачка (Cypripedium).

Таблица 57. Норичниковые: 6 - мытник головчатый (Pedicularis capitata), Дальний Восток
Нектарники у норичниковых обычно имеют вид округлого или лопастного диска, расположенного вокруг завязи, реже только с одной из ее сторон. У коровяка цветки совсем лишены нектара, и насекомые посещают их только ради пыльцы. Тычинки обычно чередуются с долями венчика и почти всегда прикреплены к его трубке. Только немногие роды (например, коровяк) имеют 5 тычинок. У других родов пятая (верхняя) тычинка превращена в стаминодий, очень крупный и варьирующий по строению в родах норичник (Scrophularia, рис. 222, 3) и пенстемон (Penstemon, табл. 57). Остающиеся две пары тычинок обычно развиты неодинаково, и одна из них может редуцироваться, например у вероники (Veronica, рис. 222, 2, табл. 57).

Таблица 57. Норичниковые: 2 - пенстемон пильчатолистный (Penstemon serrulatus), парк Ботанического института АН СССР в Ленинграде

Таблица 57. Норичниковые: 4 - вероника длиннолистная(V.longifolia), Московская область
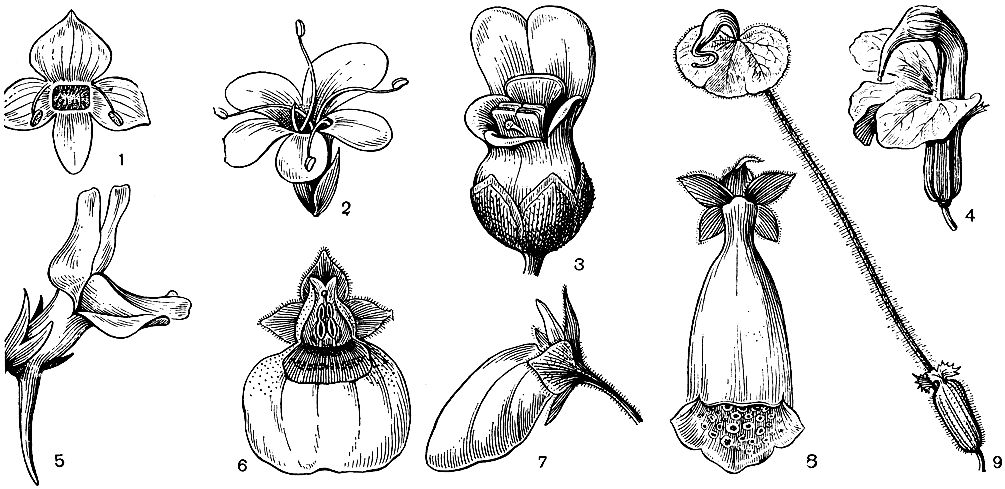
Рис. 222. Цветки норичниковых: 1 - вероника лекарственная (Veronica officinalis); 2 - селаго щитковидный (Selago corymbosa); 3 - норичник водяной (Scrop hularia aquatica); 4 - мытник мясо-красный (Pedicularis incarnata); 5 - льнянка альпийская (Linaria alpina); 6 и 7 - кальцеолария подорожниковая (Calceolaria plantaginea), вид цветка спереди и сбоку; 8 - наперстянка пурпурная (Digitalis purpurea); 9 - мытник вооруженный (Pedicularis armata)
Очень разнообразно строение пыльников. Иногда (у киксии - Kickxia) они попарно срастаются, часто на них имеются направленные вниз острия. Нередко теки их расходятся нижними концами на 180°, а вершинами срастаются друг с другом. В других случаях, напротив, теки отделены друг от друга сильно разросшимся связником (у аврана - Gratiola). Наконец, одна из тек может редуцироваться. Имеющие различное строение пыльцевые зерна часто склеиваются в комочки, но у видов трибы по-гремковых они всегда разделены друг от друга и пыльца пылевидная.
Гинецей норичниковых образован 2 плодолистиками, паракарпный или вторичносинкарпный. Цельное или двулопастное рыльце обычно расположено на столбике, отходящем от верхушки верхней завязи, лишь у эринуса (Erinus) оно сидячее.
Плоды большинства норичниковых имеют форму коробочки, вскрывающейся различными способами: 2 створками по швам плодолистиков или их средней жилке, 4 створками, 1 или 2 отверстиями с крышечкой. У торении Фурнъе (Torenia fournieri) такая крышечка занимает большую часть плода. Невскрывающиеся многосемянные ягодообразные плоды имеют африканские кустарники из рода халлерия (Halleria) и американский леукокарпус. Также невскрывающиеся, но односемянные и нередко костянкообразные плоды характерны для очень обособленной и преимущественно южноафриканской трибы селаговых (Selagineae).
Семена норичниковых имеют мясистый эндосперм и прямой или немного согнутый зародыш. Крылатые семена, напоминающие семена бигнониевых, встречаются у льнянки, погремка и немезии. У видов марьянника и некоторых других родов семена несут особые, богатые жирами придатки - элайосомы, служащие пищей для муравьев.
К семейству норичниковых относится, вероятно, не менее 250 родов и не менее 3000 видов, распространенных по всей суше земного шара, но особенно многочисленных в горных районах умеренно теплых и субтропических областей. Лишь триба жерардиевых распространена преимущественно в тропиках. Вполне определенную географическую приуроченность показывают многие крупные роды. Крупнейший род семейства - мытник (около 600 видов) - наиболее богат видами в Гималаях (здесь около 300 эндемичных видов). Роды кальцеолярия (около 300 видов), пенстемон (около 250 видов), губастик (около 120 видов) почти целиком приурочены к Америке. В Гималаях и горных районах Западной Азии и Средиземноморья широко представлен род норичник (около 300 видов).
Несмотря на обилие видов, норичниковые обычно не играют существенной роли в растительных группировках. Некоторые виды марьянника в изобилии развиваются в травянистом ярусе лесов, а виды погремка и очанки (Euphrasia) - на лугах, но они, будучи однолетниками, быстро отцветают и теряют свое значение. Более существенна роль некоторых видов хебе (Hebe), которые вместе с некоторыми сложноцветными образуют очень характерную кустарничковую группировку в горах Новой Зеландии на высоте 1200-1600 м над уровнем моря. Среди норичниковых вообще очень много петрофильных (камнелюбивых) растений, обитающих на каменистых склонах, скалах и осыпях.
Значительное большинство норичниковых имеет обоеполые перекрестно опыляющиеся цветки. Двудомность известна только у некоторых видов наперстянки. Самоопылению во многих случаях препятствует протогиния, а в более редких случаях - протандрия. При одновременном созревании рыльца и пыльников большее значение имеет разная длина тычинок и столбика. Вползая в цветок, насекомое сначала касается рыльца, на которое таким образом попадает пыльца с других цветков, а потом уже пыльников. У многих видов губастика имеется замечательное приспособление, препятствующее самоопылению при выползании насекомого из цветка. При первом же прикосновении насекомого к лопастям рыльца они в течение нескольких секунд смыкаются друг с другом и остаются в таком положении, когда насекомое выползает из цветка. У глоссостигмы (Glossostigma) лопаткообразное рыльце после первого прикосновения к нему быстро закручивается кверху над пыльниками, и насекомое на обратном пути его уже не задевает. Однако многие норичниковые способны самоопыляться, что гарантирует образование плодов, если перекрестного опыления не произошло. У одних видов рыльце постепенно завертывается книзу и в конце цветения соприкасается с пыльниками, у других рыльце приходит в соприкосновение с пыльниками при опадении венчика.
У многих видов очанки (рис. 221, 4-6) рыльце сначала находится далеко впереди пыльников, но затем основание венчика начинает удлиняться и прикрепленные к нему тычинки, выдвигаясь вперед, к концу цветения соприкасаются с рыльцем. Широко распространенное "земноводное" растение лужайник водяной обычно самоопыляется. При обитании на дне водоемов он образует совершенно закрытые, клейстогамные цветки (см. рис. 221).
Опылителями цветков у норичниковых являются насекомые, а в тропиках и субтропиках Америки также колибри. Цветки коровяка и вероники с широко открытыми цветками с короткой трубкой не имеют специализированных опылителей и опыляются различными насекомыми. Длиннотрубчатые цветки обычно приспособлены к очень ограниченному числу опылителей, нередко к одному или немногим видам пчел или шмелей. Таковы маскообразные закрытые венчики видов льнянки и львиного зева. У многих видов губастика, также опыляемых крупными перепончатокрылыми, зев венчика закрыт выпуклиной верхней губы, однако опыляющийся с помощью колибри губастик пурпурный (M. cardinalis) имеет широко раскрытые цветки. Пузыревидно вздутая нижняя губа кальцеоларий, как и у орхидеи - венерина башмачка, служит как бы ловчей ямой для насекомых. Последние, попадая в нее, могут выбраться лишь после соприкосновения сначала с рыльцем, а затем с пыльниками. Некоторые виды мытника с очень длинной трубкой венчика (см. рис. 222) опыляются длиннохоботковыми бабочками. У норичника и коллинсии тычинки и рыльце занимают нижнее положение в цветке, и насекомые (преимущественно осы) задевают за них не спинкой, а брюшком. В цветках из трибы погремковых на пыльниках имеются направленные вниз придатки в виде острий. Попеременно задевая за них спинкой, опылитель-насекомое получает по порции пылевидной пыльцы из раскрывающейся при ударе об острие щели в пыльниковом гнезде.
В распространении семян норичниковых основная роль принадлежит ветру. Во многих случаях ветер просто качает плодоносящие стебли, рассеивая семена из раскрывшихся коробочек. Семена, имеющие крыловидную кайму, рассеиваются на более далекие расстояния. В некоторых родах (коровяк, норичник и др.) имеются виды с растопыренно разветвленными и обламывающимися у своего основания стеблями, рассеивающими семена по типу перекати-поле. Семена многих видов могут распространяться также на ногах различных животных. В Южной Африке имеются зоохорные виды с крючковидно загнутыми прицветниками (у хебенштретии - Hebenstretia) или зубцами чашечки (у манулеи крючковатой - Manulea uncinata). У европейской цимбал арии постенной и некоторых стелющихся вероник цветоножки при плодах обладают отрицательным фототропизмом и загибаются вниз, вследствие чего коробочки могут зарываться в землю (геокарпия) или попадать в трещины скал и там раскрываться (см. рис. 220).
Видам марьянника свойствен мирмекохорный (с помощью муравьев) способ распространения. Их гладкие эллипсоидальные семена с поедаемым муравьями придатком из богатых маслами паренхиматических клеток часто по внешнему облику похожи на муравьиных куколок, что, по-видимому, также привлекает муравьев. Кроме того, многие марьянники имеют внецветковые нектарники, привлекающие муравьев, и не исключено, что муравьи специально распространяют такие полезные для них растения близ муравейников.
Семейство норичниковых довольно естественно подразделяется на 3 подсемейства - собственно норичниковые (Scrophulaiioideae), погремковые (Rhinanthoideae) и шаровницевые (Globularioideae). К подсемейству норичниковых относится более половины всех родов семейства, в том числе авран, губастик, лужайник, коровяк, трапелла (Trapella), левкофиллум, пенстемон, норичник, селаго (Selago), кальцеолария, льнянка, цимбалария, львиный зев. Подсемейство погремковых, насчитывающее около 85 родов, включает наперстянку, веронику, хебе, мытник, очанку, зубчатку, погремок, марьянник, петров крест и др. У подсемейства норичниковых в бутоне две верхние (задние) доли венчика кроют края боковых долей, у погремковых две верхние доли прикрыты краями одной или обеих боковых долей. По расположению долей венчика в бутоне к погремковым близко небольшое подсемейство шаровницевые, которое отличается главным образом псевдомономерным гинецеем и односемянными орехообразными плодами. В это подсемейство входят 2 рода - шаровница (Globularia, около 25 видов, распространенных от островов Зеленого Мыса до Западной Азии) и поскея (Poskea, 2 вида в Сомали и на острове Сокотра). Многие ботаники выделяют эти два рода в отдельное семейство шаровницевые (Globulariaceae), но, как это отметил еще Ханс Халлир (1903, 1912), а позднее Р. Торн (1968, 1976) и А. Л. Тахтаджян (1980), для этого нет достаточных оснований.
Значение норичниковых в жизни человека относительно невелико. Некоторые виды наперстянки дают ценное лекарство, регулирующее деятельность сердца и повышающее кровяное давление. Венчики цветков, а иногда и листья некоторых видов коровяка используют как мягчительное и отхаркивающее средство. В гомеопатии для лечения глазных болезней применяют виды очанки (Euphrasia). Паразитирующая на корнях витекса (Vitex) алектра паразитическая (Alectra parasitica) дает широко используемое в Индии средство против проказы.
Очень многие норичниковые культивируют в качестве красиво цветущих декоративных растений. Особенно широко известен происходящий из Средиземноморья львиный зев (Antirrhinum majus), многочисленные сорта которого разводят почти во всех внетропических странах. Несколько реже культивируют виды наперстянки, особенно наперстянку крупноцветковую (D. grandiflora), губастика, кальцеоларии, пенстемона, немезии, льнянки и некоторых других родов. В Западной Европе в альпинариях довольно часто культивируют новозеландские кустарнички из рода хебе (Hebe), а некоторые водные виды лимнофилы принадлежат к обычным аквариумным растениям.

Таблица 57. Норичниковые: 3 - вероника нитевидная(Veronica filiformis), парк Ботанического института АН СССР в Ленинграде
Семейство бигнониевые (Bignoniaceae) (Н. Н. Цвелев)
В противоположность близкому семейству норичниковых бигнониевые в основном тропическое семейство. На обширной территории СССР только близ ее южных границ встречаются в диком состоянии немногие виды одного из родов этого семейства - инкарвиллеи (Incarvillea), включая в него прежде принимавшийся за самостоятельный и эндемичный для гор бассейна рек Чу и Или род недзвецкия (Niedzwedzkia). Более известны в нашей стране культивируемые в садах и парках ее южных районов деревья этого семейства: катальпа (Catalpa, табл. 58) и павловния (Paulownia).

Таблица 58. Бигнониевые: 1 - катальпа бигнониевидная (Catalpa bignonioides), Сухуми

Таблица 58. Бигнониевые: 2 - катальпа яицевиднолистная (С. ovata), Никитский ботанический сад
Из жизненных форм для бигнониевых наиболее характерны деревянистые лианы, которых особенно много в Южной и Центральной Америке. Лианы могут быть как вьющимися, так и лазящими с помощью различных приспособлений. Чаще всего последними служат простые или разветвленные усики, происходящие из одного или нескольких листочков сложных листьев. Они могут заканчиваться округлыми присосками, как у питекоктениума фасолевидного (Pithecoctenium phaseoloides), или одревесневшими когтевидными крючками (у макфадиены - Macfadyena, табл. 58), но обычно просто закручиваются вокруг ветвей соседних деревьев подобно усикам винограда. У некоторых бигнониевых способностью загибаться вокруг опоры обладают также черешки, черешочки и междоузлия сложных листьев. В других случаях роль усиков выполняют придаточные корни, также нередко снабженные присосками. Такие воздушные корни, образующиеся на затененной (обращенной к опоре) части стебля, имеет, например, культивируемая на юге СССР кампсис укореняющаяся (Campsis radicans, табл. 58). Некоторые бигнониевые-лианы имеют лентовидные стебли.

Таблица 58. Бигнониевые: 5 - кампсис укореняющаяся (Campsis radicans)
Бигнониевые-деревья обычно не отличаются большой высотой. К наиболее высоким среди них принадлежат некоторые южноамериканские виды жакаранды (Jacaranda), дающие ценное палисандровое дерево. Многие небольшие деревья и своеобразные "полудеревья" из этого семейства обитают в бразильских саваннах, известных под названием кампос (или кампус). Такие деревья обычно имеют толстую и сильно одревесневшую нижнюю часть ствола, переходящую выше в почти травянистую часть, отличающуюся от стеблей многолетних трав лишь способностью образовывать вторичную древесину. Среди довольно многочисленных кустарниковых бигнониевых можно отметить сильно колючие прямостоячие кустарники из родов катофрактес (Catophractes) и ригозум (Rhigozum), обитающие в пустынях Южной Африки. Полукустарник бразильского кампоса анемопегма смолоносная (Anemopaegma mirandum, рис. 223, 2), имеющий почти эрикоидный облик (линейные с завернутыми на нижнюю сторону краями листочков сидячие листья), горит ярким пламенем из-за обилия смол во всех его частях. Настоящих трав среди бигнониевых немного, и это почти исключительно горные растения. К ним принадлежат многие виды инкарвиллеи, некоторые из них двулетники или даже однолетники.
Листья бигнониевых обычно супротивные, реже очередные или мутовчатые и всегда лишены прилистников, хотя у некоторых видов у основания черешков имеется пара прилистникоподобных листочков. В семействе значительно преобладают сложные, часто дважды или трижды перистые листья, иногда достигающие в длину более метра. У бигнониевых лиан конечный листочек или несколько листочков часто видоизменяются в усики. Длинночерешковые листья с 3-9 пальчаторасположенными перисторассеченными листочками имеют оригинальный во многих других отношениях чилийский травянистый вид аргилия лучистая (Argylia radiata). Очень интересны листья филлартрона мадагаскарского (Phyllarthron madagascariense, см. рис. 223), состоящие из листовидно расширенного черешка сложного листа, сочлененного на верхушке с единственным листочком (возможно, листовидно расширенным черешочком редуцированного листочка). Простые, цельные и целыюкрайние листья имеются лишь у немногих бигнониевых. Мексиканские кустарники хилопсис линейный (Chilopsis linearis, см. рис. 223) и астиантус лозный (Astianthus viminalis) имеют очень узкие, ланцетные или линейно-ланцетные листья, напоминая по облику иву-лозу (Salix viminalis).
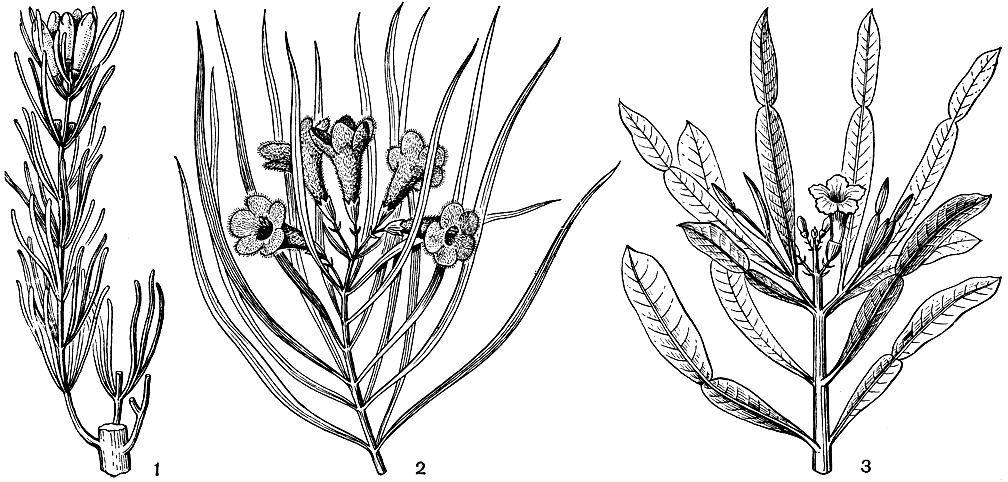
Рис. 223. Цветущие ветви бигнониевых: 1 - анемопегма смолоносная (Anemopaegma mirandum); 2 - хилопсис линейный (Chilopsis linearis); 3 - филлартрон мадагаскарский - (Phyllarthron madagascariense)
Бигнониевые могут быть совершенно голыми или иметь покров из трихом различного строения. Особенно часто встречаются очень мелкие чешуйковидные трихомы. Существенное систематическое значение имеют внецветковые нектарники, располагающиеся в узлах стебля, в углах боковых жилок листочков, на черешках, в нижней части трубки чашечки и на ее лопастях. Сахаристые вещества, выделяемые такими нектарниками, служат средством для привлечения муравьев, защищающих растение от вредителей. Очень мелкие блюдцевидные нектарники имеются даже на плодах некоторых бигнониевых. Особенно их много на молодых плодах "колебасового дерева" (Crescentia cujete), надежная защита которых от преждевременного использования в пищу насекомыми и другими животными также обеспечивается муравьями.
Способ привлечения муравьев выделениями внецветковых нектарников можно считать результатом более высокой специализации по сравнению с также широко распространенным среди тропических бигнониевых способом предоставления муравьям дарового жилища. В последнем случае ветви деревьев или кустарников имеют в своей сердцевине полости, заселяемые особыми видами муравьев. Особенно характерен этот способ для многих видов американского рода табебуйя (Tabebuia), само название которого в переводе с языка местных индейцев означает "муравьиное дерево".
Среди бигнониевых распространено и другое средство защиты от насекомых-вредителей - наличие неприятного для них запаха. Так, виды рода пахиптера (Pachyptera) имеют сильный запах лука или чеснока, а виды танециума (Tanaecium, рис. 224) сильно пахнут миндалем, что обусловлено присутствием во всем растении ядовитых цианистых соединений.
Всегда обоеполые, пятичленные и зигоморфные цветки бигнониевых обычно собраны в рыхлые конечные (редко пазушные) верхоцветные соцветия, состоящие из дихазиев. Однако имеются и исключения. Так, у макфадиены когтеносной (Macfadyena unguiscati, табл. 58) цветки располагаются по одному в пазухах обычных листьев. У аргилии лучистой расположенные очередно по спирали цветки образуют простое кистевидное (но с очень короткими цветоножками) соцветие. Каулифлория хорошо выражена у "свечного дерева" (Parmentiera cereifera, рис. 224) из Панамы и аденокалимы крупнолистной (Adenocalymma macrophyllum) из Бразилии. Аденокалима прицветниковая (A. bracteatum) имеет в соцветии очень крупные прицветники челнообразной формы, в то время как у некоторых других бигнониевых они едва заметны или даже отсутствуют.

Таблица 58. Бигнониевые: 3 - макфадиена когтеносная (Macfadyena unguiscati), Батумский ботанический сад
Сростнолистная чашечка у разных родов бигнониевых может иметь очень различное строение. Нередко она почти правильная с 5 зубцами или без них, но чаще двугубая или с одной стороны глубоко вырезанная, покрывалообразная. У африканского дерева спатодеи колокольчатой (Spathodea campanulata, см. рис. 224) крупный красный венчик выходит как бы сбоку покрывалообразной чашечки. Очень широкая красная чашечка южноамериканской лианы каллихламис (Callichlamys) служит дополнительным средством привлечения опылителей, вероятно колибри. У другой американской лианы - турретии (Tourretia) - сильно разросшаяся и направленная вперед нижняя лопасть также ярко окрашенной чашечки служит посадочной площадкой для опылителей, заменяя неразвитую нижнюю губу венчика.
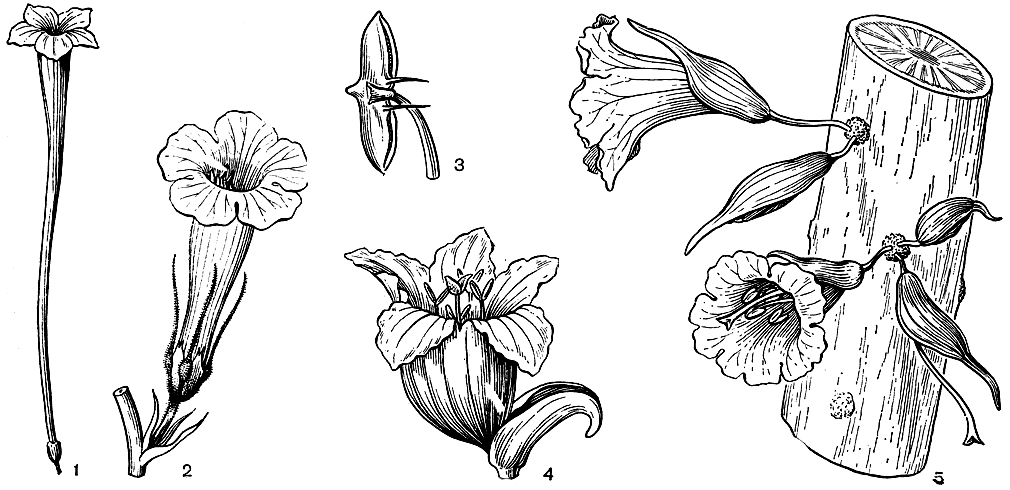
Рис. 224. Бигнониевые. Танециум длинновенчиковы и (Tanaecium praelongum): 1 - цветок. Инкарвиллея китайская (Incarvillea sinensis): 2 - цветок; 3 - пыльник. Спатодея колокольчатая (Spathodea campanulata): 4 - цветок. Парментьера свеченосная (Parmentiera cereifera): 5 - цветки на старой ветви
Все бигнониевые имеют сростнолепестные, колокольчатые, трубчатые или воронковидные венчики, в окраске которых преобладают красный и розовый цвета. Довольно часто встречаются сильно зигоморфные, двугубые венчики, похожие на венчики видов семейства губоцветных. Венчики некоторых опыляющихся длиннохоботковыми ночными бабочками бигнониевых имеют узкую, но необыкновенно длинную трубку, достигающую у рода танециум (Tanaecium) в длину 25 см. Самые мелкие, длиной 4-6 мм, венчики известны у американских лиан из рода тиннантус (Tynnanthus). Венчики древесной американской лианы пахиптеры многоцветной (Pachyptera hymenaea) в течение трех дней цветения изменяют свою окраску от темно-лиловой до светло-лиловой и белой. На третий день цветки пахиптеры уже не имеют ни пыльцы, ни нектара, но играют заметную роль в привлечении насекомых-опылителей к растению.
Большинство бигнониевых имеет 4 вполне развитые тычинки. Пятая тычинка обычно видоизменена в стаминодий, а у немногих родов, где эта тычинка имеется, она обычно недоразвита. У видов катальпы только 2 передние тычинки вполне развиты. Пыльники вскрываются продольной щелью и нередко имеют приспособления для более совершенного опыления в виде шпорцевидных или мешковидных придатков (см. рис. 224). Строение оболочки пыльцевых зерен у бигнониевых очень варьирует и является важным систематическим признаком.
Все бигнониевые имеют ценокарпный гинецей из 2 плодолистиков с двугнездной, реже одногнездной или четырехгнездной завязью и конечным двулопастным столбиком. Семязачатки почти всегда многочисленные, анатропные. Строению плодов придается основное значение при разделении семейства на трибы. Наиболее обычный тип плода - коробочка, раскрывающаяся двумя створками вдоль швов или вдоль средней жилки плодолистиков. Более редки нераскрывающиеся сочные плоды, характерные для трибы кресцентиевых (Crescentieae). Коробочки бигнониевых нередко достигают больших размеров и придают всему растению очень своеобразный облик. В этом отношении особенно замечательна кигелия перистая (Kigelia pinnata, табл. 58), иногда называемая "колбасным деревом". Крупные (длиной 30- 60 см и толщиной 7-9 см), напоминающие по форме и цвету колбасы, плоды ее свешиваются с дерева почти до земли на сильно удлиняющихся ко времени цветения цветоножках. Не менее оригинально "свечное дерево", прямостоячие цилиндрические плоды которого длиной до 1 м, окрашенные в желтый цвет, действительно создают впечатление гигантских свечей. Некоторые роды бигнониевых имеют крылатые или покрытые шипами коробочки.
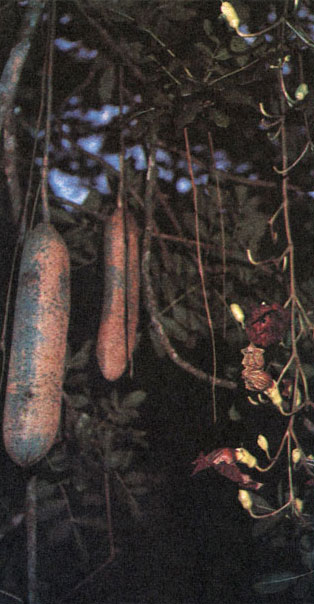
Таблица 58. Бигнониевые: 4 - кигелия перистая (Kigelia pinnata), остров Куба
Бигнониевые насчитывают свыше 120 родов и 800-900 видов, распространенных в тропиках, а отчасти и в субтропиках обоих полушарий. Лишь немногие виды заходят в южные районы умеренно теплой зоны. Особенно богата видами бигнониевых Южная. Америка, где они представлены многочисленными лианами, деревьями и кустарниками, играющими заметную роль в тропических лесах и саваннах. Хотя среди бигнониевых много нагорных видов, лишь немногие из них поднимаются в высокогорья. Это виды аргилии в Андах, амфикома (Amphicome) и инкарвиллея в Гималаях.
Все бигнониевые - перекрестноопыляющиеся растения, причем перекрестному опылению обычно способствует протандрия. Опылителями их могут быть не только насекомые (главным образом длиннохоботковые пчелы и бабочки), но также птицы (в Южной Америке - колибри) и летучие мыши. Приспособления для более надежного перекрестного опыления у бигнониевых еще недостаточно изучены. Наиболее известен механизм опыления у инкарвиллей, имеющих на почти горизонтально расположенных гнездах пыльников вниз направленные шпорцевидные придатки (см. рис. 224).
Насекомое, вползающее в трубку венчика за нектаром, задевает спинкой за эти придатки и при этом получает по порции пыльцы, высыпающейся из раскрывающейся при ударе о придаток щели в каждом гнезде. Выбираясь обратно, насекомое задевает за придатки противоположных гнезд и вновь получает по порции пыльцы из каждого пыльника. У американского рода амфилофиум (Amphilophium) сильно утолщенные двугубые венчики всегда закрыты, что защищает обильно выделяемый нектар и пыльцу от непрошеных гостей. Только крупные пчелы, опыляющие цветки этого рода, могут раскрыть зев таких венчиков и получить доступ к нектару. Очень длинные и узкие венчики упомянутого выше танециума посещаются крупными бабочками из семейства бражников. Цветки видов этого рода распускаются вечером и обладают сильным ароматом. Цветки бигнониевых, опыляемые колибри и другими мелкими птицами, обычно лишены запаха и имеют красные или фиолетовые венчики с расширенной голой внутри трубкой. Орнитофильным видом является, например, африканская спатодея колокольчатая (см. рис. 224). Летучими мышами опыляются цветки многих родов трибы кресцентиевых, в том числе "колебасовое" и "свечное" деревья. Для них характерны каулифлория, ночное цветение, обычно белая окраска венчиков, своеобразный мускусный запах цветков, выделение большого количества нектара.
Сильно сплюснутые и имеющие перепончатое крыло семена большинства бигнониевых распространяются с помощью ветра (анемохория). Парусность их обычно настолько велика, что даже в безветренную погоду или в густом лесу они разносятся на большие расстояния. У гималайского травянистого рода амфикома семена не только крылатые, но и длинноволосистые, что еще более увеличивает парусность. Сочные нераскрывающиеся плоды видов из трибы кресцентиевых распространяются с помощью поедающих их животных - эндозоохорно. Однако некоторые виды из этой трибы, имеющие довольно твердые нераскрывающиеся плоды, например центральноамериканский дендросикус широколистный (Dendrosicus lati-folius), распространяются гидрохорно - водными потоками. Имеются примеры, когда среди родов, большинство видов которых имеет характерные для бигнониевых крылатые семена, прибрежные виды обладают бескрылыми семенами, падающими в воду и распространяющимися гидрохорно. Так, из 9 видов палеотропического рода долихапдрона (Dolichandrone) 8 обитают вдали от водоемов и имеют крылатые семена, а один вид - долихапдрона Рида (D. rheedii) - растет на топких берегах рек и имеет семена с узкой окраиной из пробкообразной ткани.
Плоды некоторых американских видов кресленции, например кресцентии амазонской (Crescentia amazonica), могут распространяться как эндозоохорно, так и гидрохорно. У турретии распространение плодов также осуществляется двумя способами: экзозоохорно и анемохорно. Покрытые крючковидно загнутыми на верхушке шипами коробочки, похожие на плоды дурнишника (Xanthium), прицепляются к шерсти различных животных и разносятся ими. При этом из раскрывающейся на верхушке коробочки рассеиваются легкие крылатые семена, распространению которых способствует ветер.
Жителям внетропических стран бигнониевые известны в основном как красиво цветущие декоративные растения. Высокое (высотой до 20 м), цветущее до появления листьев дерево - павловния войлочная (Paulownia tomentosa), происходящая из Японии, часто культивируется в садах и парках юга СССР. Обычны в культуре и виды катальпы - листопадные деревья, происходящие отчасти из США, отчасти из Восточной Азии. В субтропиках ив оранжереях часто культивируются такие лианы, как кампсис укореняющийся и многие бигнонии, а также американская текома прямостоячая (Tecoma stans). В тропической Африке обычным декоративным растением является спатодея колокольчатая, называемая африканским тюльпанным деревом.
Среди тропических бигнониевых много и других, важных в хозяйственном отношении растений. Очень ценное палисандровое дерево дают бразильские виды жакаранды, введенные в культуру во многих других тропических странах. Ценную, долго сохраняющуюся древесину дают также другие американские бигнониевые: виды табебуйи, паратекома пероба (Paratecoma peroba). Широко культивируется в тропиках африканское "колебасовое" дерево (Crescentia cujete). Из его крупных, напоминающих лагенарию плодов с очень твердой оболочкой изготовляют посуду - колебасы различной формы, что достигается искусной шнуровкой молодых плодов, а также очень популярные в Африке музыкальные инструменты, похожие на трещотки или погремушки. Мякоть плодов и семена этого дерева используют в качестве лекарственного средства. В народной медицине применяют также плоды упомянутых выше "колбасного" и "свечного" деревьев. Огурцеобразные плоды пармантъеры съедобной (Parmentiera edulis) употребляют в пищу. Амазонский кустарник аррабидея чика (Arrabidaea chica) дает несмываемую краску, используемую индейцами для татуировки при ритуальных танцах.
Семейство педалиевые, или кунжутовые (Pedaliaceae) (А. Е. Бородина)
Семейство педалиевых (12 родов и около 90 видов) распространено по песчаным морским побережьям и в пустынях Старого Света (тропическая и Южная Африка, Мадагаскар, остров Сокотра, Южная и Юго-Восточная Азия, Новая Гвинея и Северная Австралия).Некоторые виды натурализовались и в Америке. Наиболее широкий ареал имеют роды сезам, или кунжут (Sesamum, рис. 225, около 30 видов), и педалиум (Pedalium, 2 вида), распространенные в тропической и Южной Африке и в Азии. Жозефиния (Josephinia, 5 видов) распространена в Австралии и на прилежащих островах. Большинство родов приурочено к Африке или к Африке и Мадагаскару, а род ункарина (Uncarina, 9 видов) эндемичен для Мадагаскара. Самые большие роды - сезам, птеродискус (Pterodiscus, 15 видов в Африке), ункарина и цератотека (Ceratotheca, около 10 видов в Африке).
Педалиевые обычно однолетние, реже многолетние травы. Исключение составляют ункарины, которые обычно кустарники, реже небольшие деревья, иногда достигающие в высоту 8 м. Стебли стелющиеся по земле или прямостоячие; листья супротивные, цельные или лопастные. Для педалиевых характерно наличие на всех частях растений клейких железистых волосков. Цветки обычно одиночные или же в 3-цветковых дихазиях, обоеполые, зигоморфные, большей частью с характерными желёзками - метаморфизированными цветками - у основания цветоножек. Чашелистиков обычно 5, иногда 4, у основания более или менее сросшихся; венчик обычно ярко окрашенный, широкотрубчатый, 5-лопастный и несколько двугубый; 4 тычинки фертильные, а пятая превращена в маленький шиловидный стаминодий. Гинецей из 2 плодолистиков, с длинным тонким столбиком и 2-лопастным рыльцем; завязь верхняя, 2-4-гнездная, иногда гнезда разделены ложными перегородками, с 1 - многими семязачатками на каждой плаценте. Плод - локулицидная коробочка или нераскрывающийся - заключает часто крылатые семена с маленьким прямым зародышем и мясистым эндоспермом.
Яркая окраска венчика, привлекающие пятна и полоски на нижней, наиболее крупной его лопасти и в верхней части широкой трубки, нектарный диск - все это, несомненно, свидетельствует об опылении педалиевых насекомыми.
Механизм опыления у кунжута был подробно изучен Б. М. Козо-Полянским (1947). Боковые цветки в дихазиях у кунжута недоразвиты и имеют форму шаровидных зеленых образований диаметром около 1 мм. На них иногда видны капельки жидкости, и, возможно, они функционируют как нектарники. В нормально развитых цветках нектар выделяется выростом цветоложа, имеющим вид воротничка вокруг основания завязи. Трубка венчика разделена перехватом на две неравные части, в месте перехвата она несет множество длинных мягких волосков, сходящихся в центре трубки и не пропускающих к нектару мелких насекомых. На розовом или лиловатом венчике имеется рисунок из двух вдавленных желтых пятен, соединенных полосой из темно-розовых или малиновых точек. Насекомое садится на нижнюю губу в месте первого желтого пятна, темная полоса указывает ему путь к нектару, а второе желтое углубление v-образной формы, расположенное внутри трубки на брюшной стороне, служит местом работы для крупных насекомых, добывающих нектар. Основными опылителями в наших условиях являются пчелы, они не только длительно обследуют открытые цветки, но и открывают также более зрелые бутоны.
Как запасной вариант возможна и автогамия благодаря тому, что зрелые рыльца и пыльники сближаются между собой, а венчик при малейшем толчке отваливается и соскальзывает вниз, при этом пыльца из пыльников обязательно попадает на рыльце. При самоопылении завязываются нормальные плоды.
Педалиевые замечательны разнообразием плодов. Плод кунжута - цилиндрическая коробочка, раскрывающаяся на верхушке двумя створками с острыми концами и заключающая в себе множество семян. Другие плоды не раскрываются и на своей поверхности имеют разнообразные выросты, которые способствуют их расселению. У птеродискуса плоды имеют крыловидные выросты, благодаря которым они легко перемещаются по поверхности земли потоками воздуха; шаровидные плоды жозефинии тоже перекатываются благодаря выростам на их поверхности. Некоторые плоды имеют шипы с колючками, которыми они прикрепляются к шерсти животных (см. рис. 225).
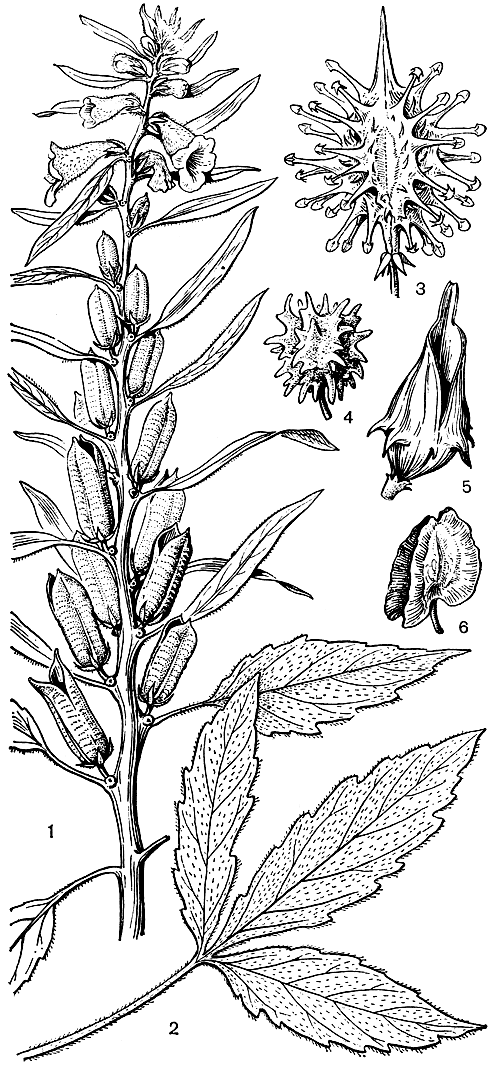
Рис. 225. Педалиевые. Кунжут индийский (Sesamum indicum): 1 - ветвь с цветками и плодами; 2 - стеблевой лист. Плоды педалиевых: 3 - ункарина звездочконосная (Uncarina stellulifera); 4 - жозефиния крупноцветковая (Josephinia grandiflora); 5 - педалиум колючий (Pedalium murex); 6 - птеродискуе прекрасный (Pterodiscus speciosus)
В практическом отношении важнейшим растением семейства является кунжут индийский (Sesamum indicum) - одно из лучших масличных растений тропиков Старого Света, культивируемое от западных берегов Африки вплоть до Китая и Японии, а также в Америке. Это довольно крупные (высотой до 1-2,5 м) быстро растущие однолетние растения с продолжительностью вегетационного периода от 2,5 до 5 месяцев. Красные, палево-розовые или белые цветки распускаются в пазухах листьев и формой и окраской напоминают наперстянку. Семена содержат от 50 до 65% масла, которое служит основой высококачественного пищевого масла, приближающегося по вкусу к оливковому маслу; кунжутовое масло также применяют в производстве маргарина и в мыловарении, высшие его сорта используют в парфюмерии. Кроме того, семена кунжута используют при производстве восточных сладостей, особенно тахинной халвы, а также для обсыпки сдобных булок вместо обычного у нас мака. Из копоти сгоревшего масла изготовляли высококачественную китайскую тушь. В диком состоянии кунжут неизвестен. Время и место происхождения культурного кунжута, как и большинства древнейших культурных растений, остаются невыясненными. Наиболее вероятно африканское происхождение кунжута, где сосредоточены почти все дикорастущие виды сезама, однако возможно, что родиной его являются Северная Индия и Пакистан. Возделывание кунжута началось задолго до нашей эры в странах Юго-Западной Азии; культура кунжута была известна в древней Греции и Риме, в Месопотамии, Аравии, Индии, а с начала нашей эры и в Китае. В настоящее время общая площадь посевов кунжута в мире 6,4 млн. га. В СССР кунжут возделывают в Средней Азии, Азербайджане и Краснодарском крае. В Центральной и в Южной Африке как масличное растение культивируют сезам лучистый (S. radiatum). Цератотеку сезамовидную (C. sesamoides) родом из Западной Африки культивируют ради маслянистых семян и листьев, которые кладут в суп. Кору мадагаскарских ункарин используют при получении слабых дубителей и стойкой краски для тканей, а их листья - как жевательный бетель.
Очень близко к педалиевым небольшое тропическое и субтропическое американское семейство мартиниевых (Martyniaceae) с 3 родами, также растущими на песчаных почвах и морских побережьях: монотипный род мартиния (Martynia), краниолария (Craniolaria, 2 вида) и пробосцидея (Proboscidea, 6 видов). От педалиевых мартиниевые отличаются одногнездной завязью и париетальной плацентацией, а также плодом с мясистым опадающим экзокарпием и деревянистым эндокарпием; столбик при плоде расщепляется на две части и превращается в жесткие, изогнутые, заостренные на конце придатки, напоминающие слоновые бивни и служащие для расселения плодов в шерсти животных. Наступая на такие плоды, пасущийся скот ранит ноги, что вызывает опасные заболевания. Как заносное растение мартиния встречается в Африке.
Семейство заразиховые (Orobanchaceae) (Н. Н. Цвелев, Э. С. Терехин)
Все виды семейства заразиховых принадлежат к числу многолетних корневых паразитов и полностью лишены хлорофилла. С необычным образом жизни тесно связаны основные особенности этого семейства: своеобразный облик растений, отсутствие настоящих корней, чешуевидные листья, очень мелкие и многочисленные семена с очень слабо редуцированным зародышем.
Семена заразиховых-легко проникают в глубь почвенного слоя с дождевой водой, где проростки вступают в контакт с корнями растений-хозяев. Вероятность успешного контакта невелика, однако его возможность обеспечивается у заразиховых двумя приспособлениями. Во-первых, обилием семян, которое дает каждое растение. Например, в одной завязи эгинетии индийской (Aeginetia indica) образуется до 70 000 семян, а одна особь заразихи (Orobanche, табл. 59) может дать до 500 000 и более семян. Во-вторых, семена заразихи, подобно семенам петрова креста и некоторых других паразитных норичниковых, прорастают только под воздействием особых веществ - стимуляторов, выделяемых корнями растений-хозяев. Эти выделения не только вызывают прорастание семян, но и ориентируют рост проростка в направлении хемостимулятора (корня растения-хозяина).
Приспособительное значение этих особенностей очевидно, и, вероятно, ими обладают и другие представители семейства. Зона действия стимуляторов прорастания распространяется в почве довольно широко, поэтому далеко не все прорастающие в этой зоне семена заразихи способны достичь корня растения-хозяина: возможности проростка очень невелики, лишь в некоторых случаях зародышевый корешок заразихи достигает нескольких миллиметров, у заразихи кумской (Orobanche cumana), например, до 0,5 см. При соприкосновении с корнем растения-хозяина верхушка зародышевого корня заразихи преобразуется в гаусторию (присоску) и активно внедряется в него при посредстве протеолитических ферментов.
Дальнейшее развитие проростка заразихи происходит лишь после контакта ее гаусториальных клеток с проводящей системой корня растения-хозяина. В месте внедрения из части проростка, примерно соответствующей гипокотилю, развивается клубневидное образование, называемое обычно клубеньком. В клубеньке запасаются питательные вещества, закладываются плодущие побеги и вторичные гаусториеобразующие органы, обеспечивающие вегетативное размножение растения-паразита. У многих видов заразихи, особенно у видов, паразитирующих на культурных, обычно однолетних растениях, из клубенька в первый же сезон развивается плодущий побег, который отмирает вместе с питающим растением. Однако у видов, паразитирующих на многолетних растениях, развитие паразита возобновляется Б следующем году из новых клубеньков, образующихся в результате повторных внедрений вторичных гаусториеобразующих органов в корни растения-хозяина. У пучкоцвета трубкоцветкового (Phacellanthus tubiflorus), паразитирующего на корнях ясеня, система вторичных гаусториеобразующих органов формирует густую сеть, из которой в отдельные годы возникает сразу до нескольких десятков побегов, принадлежащих к одной особи.
У представителей некоторых родов заразиховых клубеньки существуют в течение ряда лет я дают начало все новым и новым подземным побегам. Последние, правда, зацветают не каждый год, так как из зачатков побегов здесь развиваются обычно мощные растения. У некоторых видов цистанхи (Cistanche) стебли достигают в высоту 1-1,5 м. На развитие таких побегов требуется как минимум 2-3 года. Поэтому "клубенек" цистанхе иногда весит более 10 кг. Напротив, у маннагеттеи (Mannagettaea) стебли крайне редуцированные, подземные, на поверхности почвы бывают видны лишь соцветия этого растения.
Цветки заразиховых располагаются в пазухах прицветников (иногда еще с прицветничками), обычно сходных по форме и строению с чешуевидными листьями, образуя конечные колосовидные, кистевидные, метелковидные, реже щитковидные или головковидные соцветия бокоцветного типа. У южноазиатской эгинетии (Aeginetia) и у некоторых американских заразих очень короткие стебли заканчиваются кистью с немногими цветками на очень .длинных цветоножках, а у видов распространенного в Крыму, на Кавказе и в Передней Азии рода дифелипея (Diphelypaea) стебель несет лишь один, очень редко два крупных цветка (рис. 226, табл. 59). Чашечка очень разнообразна по строению. Так, в пределах обширного рода заразих чашечки варьируют от колокольчатых, 4-5-зубчатых до расщепленных на 2 цельных или двузубчатых сегмента. Сростнолепестный пятичленный венчик обычно имеет трубчатую, воронковидную или колокольчатую форму. У цистанхе он еще близок к актиноморфному, пятилопастный, но у большинства родов семейства он зигоморфный и обычно двугубый. У южноазиатской христисопии трубчатой (Christisonia tubulosa) голубые венчики достигают в длину 7,6 см, а у дифелипеи ярко-красные или желтые венчики могут быть длиной до 5 см.

Таблица 59. Заразиховые: 1 - дифелипея красная (Diphelypaea coccinea), Южный Крым
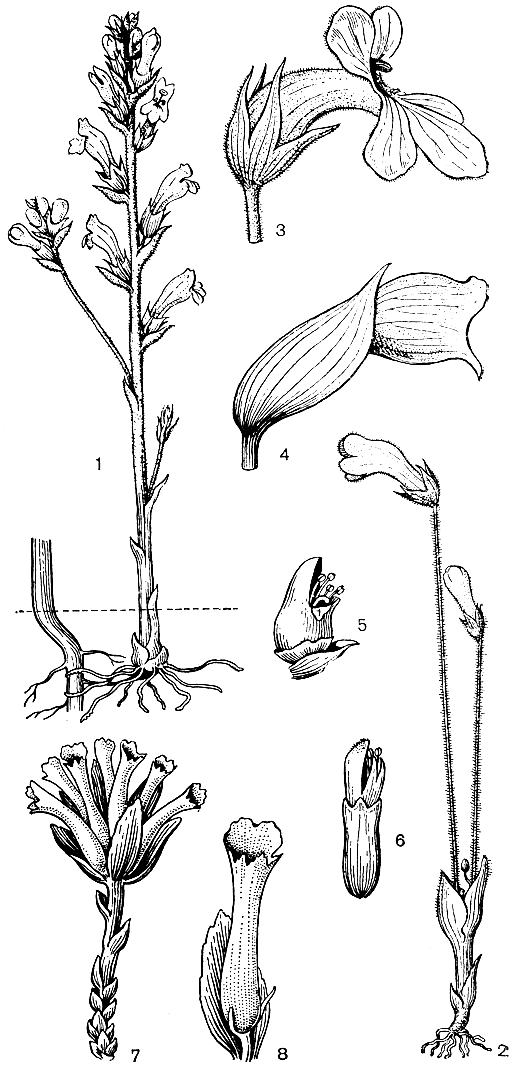
Рис. 226. Заразиховые. Заразиха ветвистая (Orobanche ramosa): 1 - общий вид растения. Заразиха одноцветковая (O. uniflora): 2 - общий вид растения. Дифелипея красная. (Diphelypaea coccinea): 3 - цветок. Эгинетияиндийская (Aeginetia indica): 4 - цветок. Бошнякия русская (Boschniakia rossica): 5 - цветок. Маннагеттея иркутская (Mannagettaea ircutensis): 6 - цветок. Пучкоцвет трубкоцветковый (Phacellanthus tubiflorus): 7 - общий вид растения; 8 - цветок
Тычинок в цветке заразиховых обычно 4, редко 3 (у тиенмуйи - Tienmuia), прикрепленных к трубке венчика близ ее середины или ниже, чередуясь с лопастями или зубцами ленчика. Пятая тычинка отсутствует, реже превращена в стаминодий, как у заразихи одноцветковой (Orobanche uniflora).
Заразиховые имеют паракарпный гинецей, состоящий из 2, реже 3 или 4 (у копсиопсиса - Kopsiopsis) плодолистиков. Столбик заканчивается головчатым, воронковидным или 2-4-лопастным рыльцем. Завязь верхняя, одногнездная (лишь у христисонии и бошнякии - Boschniakia - близ основания двухгнездная), с 2-6, но обычно с 4 париетальными плацентами, несущими многочисленные семязачатки. У большинства видов плод - коробочка, раскрывающаяся двумя, реже тремя створками, у пучкоцвета плод ягодообразный. Семена содержат богатый крахмалом и маслами эндосперм и редуцированный зародыш.
Семейство заразиховых включает 13 родов и около 200 видов, распространенных очень широко, но крайне неравномерно. В Европе, Азии и северной части Африки встречается около 90% всех видов семейства, особенно многочисленных в Евразии, от Канарских островов и Пиренейского полуострова до Гималайских гор. В южном полушарии заразиховые почти отсутствуют. Не считая заносных из Европы, здесь встречаются только немногие виды заразихи в Австралии и Южной Америке. Тропики также бедны заразиховыми. В горных районах таежной зоны Азии и Северной Америки широко распространена паразитирующая на некоторых видах ольхи бошнякия русская (Boschniakia rossica), заходящая в низкоширотные районы Арктики. В Северной Америке, кроме того, встречаются 2 эндемичные секции заразихи и 3 эндемичных рода: конофолис (Conopholis), эпифагус (Epifagus) и копсиопсис.
Многочисленные (около 140) виды заразих (Orobanche, табл. 59) обитают в различных растительных группировках, за исключением болот, лесов таежного типа и тундры. Особенно много их на каменистых и мелкоземлистых склонах гор с травянистой, кустарничковой или кустарниковой растительностью. Среди них имеются как строгие монофаги, например паразитирующая на плюще заразиха плющевая (Orobanche hederae), так и виды, имеющие растений-хозяев из многих семейств, например заразиха ветвистая (Orobanche ramosa). Из других родов семейства виды цистанхи приурочены в основном к пустыням и полупустыням, паразитируя на кустарниках или кустарничках семейства лебедовых (в том числе на саксауле), а также на тамариске и джузгуне. Их высокие толстые, стебли, несущие густые колосовидные соцветия, выглядят очень эффектно среди пустынной растительности. В полупустынях Центральной Азии, заходя и на территорию СССР, встречаются виды маннагеттеи (рис. 226, 6), паразитирующие на карагане (Caragana). В лесах обитают такие замечательные во многих отношениях роды заразиховых, как азиатские эгинетия и христисония (первый из них паразитирует на однодольных, главным образом на злаках), восточноазиатский пучкоцвет (рис. 226, 7, 8), обычно паразитирующий на ясене, североамериканские конофолис и эпифагус, паразитирующие на таких лесообразующих породах, как дубы и буки.
Система заразиховых пока еще недостаточно разработана. Кроме того, все еще остаются неясными границы между заразиховыми и близким к нему семейством норичниковых, где также имеются облигатные паразиты, лишенные хлорофилла (например, петров крест).
Цветки многих заразиховых протогиничны: гинецей созревает раньше, чем пыльники. Однако в роде заразиха преобладают гомогамные цветки и потому самоопыление вполне возможно. Опылителями заразиховых обычно являются шмели, пчелы и другие перепончатокрылые. Приспособления, способствующие опылению, еще недостаточно изучены. К ним, вероятно, относятся мешковидные или шпорцевидные придатки пыльников эгинетии и христисонии. Некоторые виды заразих имеют приятный запах - дополнительное средство привлечения насекомых-опылителей. Разветвленные колосовидные соцветия североамериканского эпифагуса обычно имеют 2 вида цветков: клейстогамные и хазмогамные. Клейстогамные цветки имеют небольшой замкнутый венчик, всегда фертильны и располагаются в нижней части соцветий (иногда еще и в верхней). Хазмогамные, цветки, располагающиеся в средней или верхней части соцветий, имеют открытый двугубый венчик, но плодоносят крайне редко. Развивающиеся под землей клейстогамные цветки отмечались еще у западносредиземно-морского вида цистанха фелипея (Cistanche phelypaea).

Таблица 59. Заразиховые: 2 - цинтанхе желтая (Cistanche flava), Туркмения, Репетек
Мелкие и легкие семена заразиховых распространяются с помощью ветра и дождевых капель. У пучкоцвета лесными животными распространяются плоды, заполненные клейкой пульпой, в которую погружена масса мельчайших семян. Засохший венчик часто долго сохраняется при плодах, препятствуя быстрому рассеиванию семян. Сухие стебли многих заразих с еще содержащими семена плодами могут переноситься ветром, рассеивая семена по способу перекати-поле.
Некоторые виды заразих принадлежат к числу злостных сорняков, значительно снижающих урожаи сельскохозяйственных культур. Это особенно относится к заразихе кумской (Orobanche cumana), в изобилии развивающейся на полях подсолнечника, заразихе египетской (O. aegyptiaca), паразитирующей главным образом на бахчевых культурах, реже на помидорах, табаке, картофеле и других культивируемых растениях, заразихе ветвистой (O. ramosa, см. рис. 226) и заразихе Мутеля (O. mutelii), поражающих капусту, помидоры, табак, коноплю и другие культуры. Кормовым культурам из семейства бобовых (люцерне, клеверу и др.) иногда вредят заразиха желтая (O. lutea), заразиха малая (O. minor) и заразиха городчатая (O. crenata). В Юго-Восточной Азии некоторый ущерб плантациям сахарного тростника иногда наносит эгинетия индийская (Aeginetia indica). Этот вид имеет крупные ярко окрашенные цветки, и в некоторых странах введен в культуру в качестве декоративного растения. Из видов флоры СССР очень декоративны виды дифелипеи, имеющие крупные ярко-красные, реже желтые цветки. Успешные попытки введения их в культуру на альпийских горках уже были. Серебристая от войлочка зелень растений-хозяев (обычно розеткообразующих васильков из секции псефеллус) очень хорошо сочетается с ярко-красными цветками дифелипеи.

Таблица 59. Заразиховые: 3 - заразиха Кочи (Orobanche kotschyi), Таджикистан, Варзобское ущелье
Семейство геснериевые (Gesnepviaceae) (Л. И. Иванина)
В семействе около 130 родов и 2000 видов. Оно имеет широкий ареал, занимающий в основном тропические и субтропические области Азии, Африки. Америки и Восточной Австралии, а также Мадагаскар и Новую Зеландию. Представители семейства встречаются и в теплоумеренных зонах Восточной Азии, Южной Америки и в Западной Европе (карта 14).
Карта 14. Ареал семейства геснериевых
По внешнему виду геснериевые очень различны: от мелких трав до небольших деревьев. Преобладают многолетние мягковолосистые травы, но много и наземных или эпифитных полукустарников, реже встречаются лианы и кустарники. Стебли геснериевых сочные, со временем одревесневающие, обычно с моноподиальным ветвлением. У тропических трав Старого Света встречаются прямостоячие стебли, утолщенные в основании и слегка сбежистые к вершине. Они не обладают хорошо развитым корнем. Укороченные стебли с прикорневой розеткой листьев характерны для геснериевых горных субтропических или умеренных районов Старого Света. Геснериевые с ползучими или вьющимися стеблями встречаются у представителей трибы трихоспоровые (Trichosporeae), а стебли с надземными столонами (усами) развиваются у многих американских эписций (Episcia, табл. 60). Для американских геснериевых характерно образование подземных клубней (гипокотильного происхождения) или чешуйчатых корневищ.
Листья геснериевых супротивные или мутовчатые, редко очередные, нерасчлененные, цельнокрайные или зубчатые, сочные и часто опушенные, от округлых до ланцетных, со слегка неравнобоким основанием, обычно с перисто-краевым жилкованием.
Цветки геснериевых (рис. 227) обоеполые, протандричные, сравнительно крупные, одиночные в пазухах листьев или в различных бокоцветных соцветиях. Они обычно 5-членные, чашечки 5 (реже 4-6)-лопастные. Венчик из 5 лепестков, зигоморфный, колокольчатый, воронковидный или трубчатый, обычно с двугубым отгибом, иногда почти актиноморфный. Андроцей состоит из 4 или 2 тычинок (у сенполий - Saintpaulia, стрептокарпусов - Streptocarpus, циртандр - Cyrtandra, рис. 228, и других, но часто в этих цветках присутствуют стаминодии). У менее продвинутых в эволюционном отношении родов, таких, как рамонда (Ramonda), конандрон (Conandron), тычинок 5. Гинецей паракарпный, из двух плодолистиков, с простым столбиком и 2-лопастным или головчатым рыльцем. Завязь большей частью одногнездная, редко двугнездная, верхняя (в подсемействе циртандровых - Cyrtandroideae), или полунижняя, или почти нижняя (у большинства представителей подсемейства геснериевых - Gesnerioideae). Плод - коробочка, вскрывающаяся локулицидно или маргиницидно, реже нераскрывающаяся. У циртандр и у некоторых американских геснериевых с верхней завязью (беслерия - Besleria, колумнея - Columnea, табл. 60, кодонанта - Codonanthe) плод - ягода или ягодообразный. Семена многочисленные, мелкие, с прямым зародышем, с эндоспермом или без него.

Таблица 60. Геснериевые. Колумнея славная (Columnea gloriosa): 1 - ветвь с цветками; 2 - плод. Эписция медная (Episcia cupreata): 3 - ветвь с цветками. Рамонда миконская (Ramonda myconi): 4 - общий вид растения; 5 - плоды
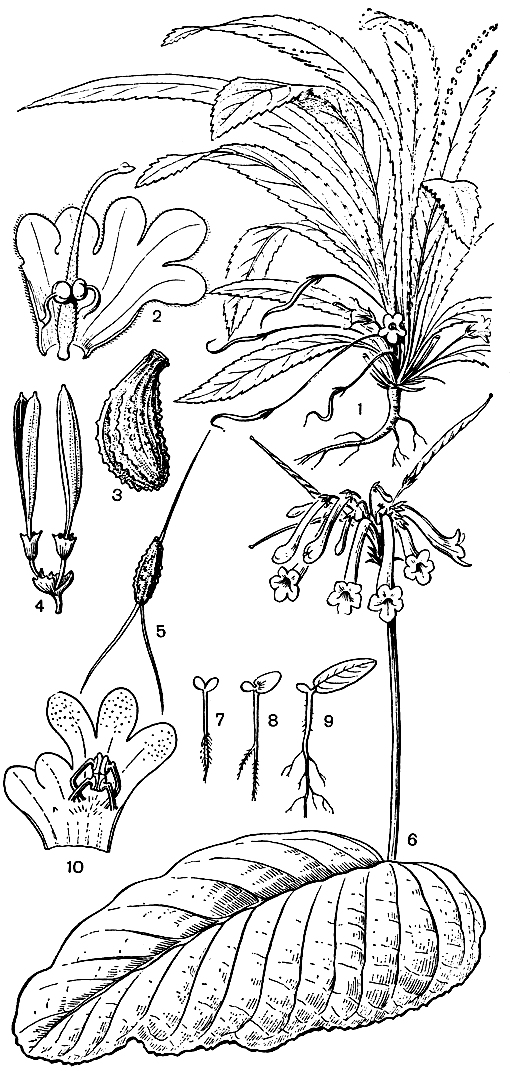
Рис. 227. Геснериевые. Дидимокарпус стройночешуйчатый (Didymocarpus gracilipes): 1 - общий вид растения; 2 - цветок в развернутом виде. Дидимокарпус крупнолистный (D. macrophylla): 3 - семя; 4 - коробочки. Эсхинантус паразитический (Aeschynanthus parasiticus): 5 - семя. Стрептокарпус трубчатоцветковый (Streptocarpus solenanthus): 6 - общий вид растения. 7, 8,9 - схема развития однолистных стрептокарпусов. Монофиллея особая (Monophyllaea singularis): 10 - цветок в развернутом виде
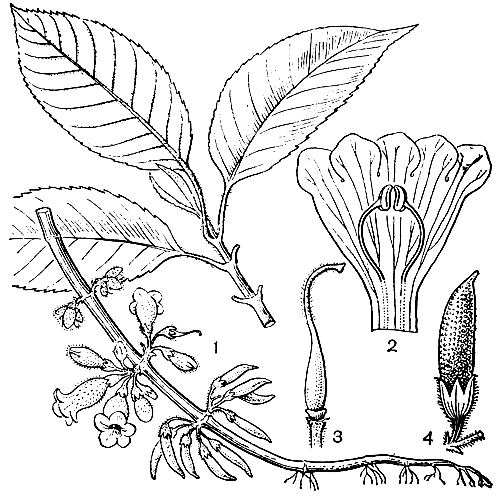
Рис. 228. Циртандра западная (Cyrtandra occidentalism): 1 - общий вид растения с цветками и плодами; 2 - цветок в развернутом виде; 3 - гинецей; 4 - плод
Цветки геснериевых обычно специализированы для перекрестного опыления при помощи различных опылителей: колибри, летучими мышами и насекомыми - пчелами, бабочками и молями. Однако у некоторых представителей семейства (особенно у стрептокарпусов) возможно самоопыление. Американские геснериевые, опыляемые колибри, обычно с ярко-красными длиннотрубчатыми цветками, с хорошо развитым нектарным диском или с крупными отдельными желёзками. Изучение вест-индских геснериевых (Л. Ског, 1976) показало тесную приспособленность разных колибри к посещению только одного вида растений, длина трубки венчика которого почти совпадает с длиной носика птиц. Так, для геснерии кубинской (G. cubensis) потенциальным опылителем является архилохус колубрис (Archilochus colubris), для геснерии бесстебелъной (G. acaulis) - антракоторакс манго (Anthracothorax mango). У растений, опыляемых летучими мышами, цветки обычно белые или кремовые, реже пурпурные, с очень обильным нектаром и пыльцой. Венчик таких растений часто с широким зевом, и, очевидно, поэтому на Кубе такие виды ритидофиллума (Rhytidophyllum, рис. 229) называют "львиным зевом". Замечено, что крупные цветки (длиной до 7 см) американской лианы гастерантуса живокостного (Gasteranthus delphinioides) опыляются в Панаме пчелами-эвглоссами. С. Фогелем (1966) был описан особый случай опыления среди двудольных растений у протандричной глоксинии многолетней (Gloxinia perennis, рис. 230). Цветок ее посещается самцами тропической пчелы эвлемы черной (Eulaema nigrita). У основания светло-сиреневого венчика глоксинии заметно темное пятно. Здесь расположены железистые клетки (осмофор), содержащие душистое, сильно пахнущее вещество. Пчела летит на цветок, привлекаемая тяжелым тминно-ментоловым запахом (сходным с запахом некоторых орхидей), и заползает в венчик с целью собрать душистое вещество, которое в дальнейшем играет важную роль при копуляции и может быть отнесено к половым аттрактантам цветка (см. Жизнь растений, т. 5, кн. I, с. 66). В тот момент, когда пчела находится в цветке, она осыпается пыльцой.
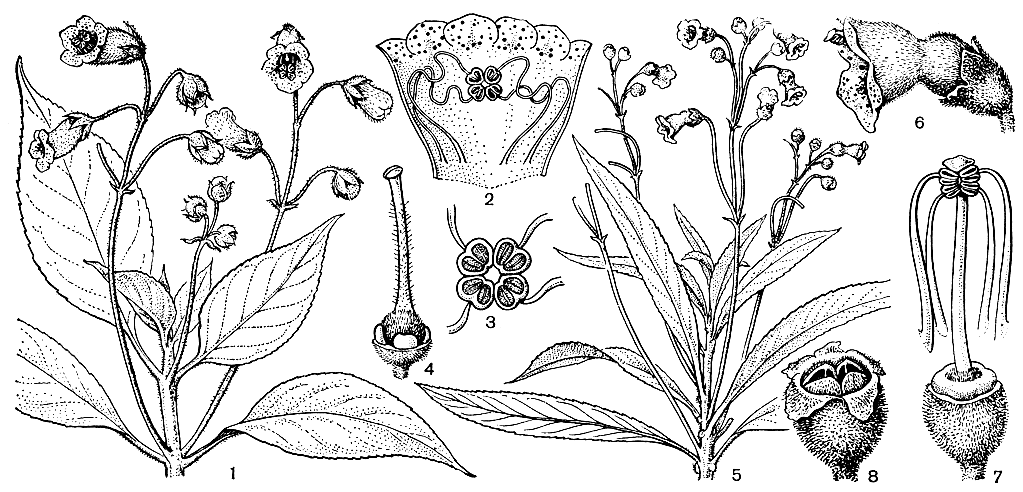
Рис. 229. Геснериевые. Капанея крупноцветковая (Capanea grandiflora): 1 - ветвь с цветками; 2 - раскрытый венчик с андроцеем; 3 - пыльники; 4 - гинецей. Ритидофиллум войлочный (Rhytidophyllum tomentosum): 5 - ветвь с цветками; 6 - цветок; 7 - гинецей и андроцей; 8 - плод

Рис. 230. Глоксиния многолетняя (Gloxinia perennis): 1 - ветвь с цветками; 2 - разрез цветка (а - осмофор); 3 - продольный разрез через железистые клетки осмофора; 4 - цветок и опылитель эвлема черная (Eulaema nigrita)
Перелетая, пчела попадает в цветок с развитым рыльцем. Здесь она задевает рыльце и происходит перекрестное опыление. Пчела же, собрав душистое вещество, выпускает его в воздухе, создавая как бы ароматное облачко, привлекающее самок, после чего происходит копуляция.
В семействе среди разнообразных способов рассеивания диаспор (в основном семян и ягод) преобладают два - анемохорный и зоохорный. Мелкие семена геснериевых легко разносятся ветром. У эпифитных полукустарников Южной и Юго-Восточной Азии (у эсхинантусов - Aeschynanthus, локсостигм - Loxostigma, лизионотусов - Lysionotus) семена приспособлены к полету благодаря волоскам, в несколько раз превышающим длину семян. Путь расселения отдельных видов на островах Вест-Индии иногда связывают с направлением сильных ветров и считают, что семена некоторых геснериевых во время ураганных ветров были перенесены с острова Пуэрто-Рико на остров Гаити, а затем на Кубу. У полукустарника Южной Азии бейки нитевидной (Boeica filiformis) створки коробочки при вскрывании плода остаются соединенными на верхушке и при основании. При раскачивании стеблей или коробочек мелкие семена легко вылетают и разносятся ветром. Здесь как бы сочетаются баллистические и анемохорные приспособления. Спиральное скручивание коробочек у стрептокарпусов, очевидно, способствует активному разбрасыванию семян. Для плодов монотипного рода каролофричии (Carolofritschia) характерна базиокарпия. Мелкие коробочки этого растения, обитающего в девственных лесах Камеруна, расположены у основания стеблей. Поедая сочные плоды геснериевых, птицы также способствуют их распространению. Возможно, этим объясняется обширный ареал рода циртандра (Cyrtandra, 600 видов), на островах Юго-Восточной Азии, Океании и в прибрежной Восточной Австралии. В семействе встречается и мирмекохория. Она замечена у тропических американских эпифитных кодонант (Соdonanthe), поселяющихся на муравейниках. Семена у кодонант крупные, с ариллусовидными придатками (Л. И. Иванина, 1967).
Геснериевые преимущественно мезофильные растения. Наибольшее видовое разнообразие их связано с тропическими вечнозелеными лесами. Они обитают в тенистых ущельях, на террасах рек и вблизи водопадов, на сырых скалах или растут на стволах деревьев в районах с частой облачностью, дождями и туманами. В Америке геснериевые встречаются преимущественно до 1200 м, реже до 2400 м (и до 3500 м) над уровнем моря. В Гималаях и в горах Китая они обычно поселяются на высоте от 2000 до 3000 м. Произрастают они и в горных листопадных лесах и кустарниковых зарослях, а в Африке (на восточных склонах Драконовых гор) и среди кустарниково-ксерофитной растительности. В Карибской и других флористических областях Америки в районах с наличием сухого и влажного сезонов года или в переменно-влажных лесах растения имеют ряд приспособлений для сохранения воды и питательных веществ. У ахименесов (Achimenes), колерий (Kohleria, табл. 61), глоксиний для этой цели служат мясистые чешуйчатые корневища, а у синнингий (Sinningia) и рехштейнерий (Rechsteineria) - подземные клубни.

Таблица 61. Геснериевые: 2 - колерия приятная (Kohleria amabilis), оранжереи Ботанического института АН СССР в Ленинграде
В семействе выделяют обычно 2 подсемейства: циртандровые (Cyrtandroideae) и собственно геснериевые (Gesnerioideae). Растения из подсемейства циртандровые произрастают в основном в Старом Свете и отличаются биологическими особенностями роста и развития. После прорастания семян у них развиваются неравные семядоли (анизокотилия). Первичные корни нередко отмирают в ювенильной стадии, их сменяют придаточные. Особый интерес среди двудольных представляет онтогенез однолистных стрептокарпусов и монофиллей (Monophyllaea, 34 южноазиатских вида, см. рис. 227). К стрептокарпусам относится около 140 видов, обитающих в основном в горах Экваториальной и Южной Африки, на Мадагаскаре и Коморских островах, а также в Таиланде и Бирме. У мо-нокарпических стрептокарпусов и у монофиллей одна семядоля после прорастания семян не развивается и отмирает, а другая продолжает расти. В месте перехода гипокотиля в главную жилку затем появляется соцветие. Семядольный лист у стрептокарпуса Купера (S. cooperi) достигает в длину 70 см, а у монофиллей Вилъдена (M. wildeana) - 100 см.

Таблица 61. Геснериевые: 1 - аллоплектус головчатый (Alloplectus capitatus), оранжереи Главного ботанического сада в Москве
В горах Западной Европы в необычном для геснериевых климате (зимы с отрицательными температурами, а лето с засушливыми периодами) встречаются три рода этого семейства. В Пиренеях и на Балканском полуострове про израстают рамонды (Ramonda, 3 вида), на Фессалийском Олимпе обитает янкея (Jankaea, монотипный род), а в Родопских горах и в более южных горах Балканского полуострова встречаются габерлеи (Haberlea, 2 вида). Рамонды растут и в тенистых лесах, и в полностью высыхающих ущельях на известняках. Они обладают редкой для цветковых растений особенностью (пойкилогидрией): находясь в засушливый период в почти воздушно-сухом состоянии, после увлажнения вновь продолжают жизнедеятельность.
В ботаническом саду в Ленинграде рамонда Наталии (R. nathaliae) и рамонда миконская (R. myconi, табл. 60) переносят суровые зимы и обычно цветут с июня до середины августа.
Геснериевые имеют исключительно декоративное значение. Многие из них давно уже признаны ценными красиво цветущими растениями. Особенно широко известны тропические "глоксинии", которыми называют гибридные синнингии (Sinningia, табл. 61) и "африканские фиалки", или сенполии. Они входят в основной ассортимент цветочных хозяйств мира. К роду синнингия сейчас присоединяют и большинство видов рехштейнерий, и тогда среди них насчитывают 75 видов, распространенных в тропических районах Америки. Наиболее часто разводят гибриды синнингии прекрасной (S. speciosa), размножая их семенами, делением корней или черенками.

Таблица 61. Геснериевые: 5 - синнингия соразмерная(Sinningia eumorpha), оранжереи Ботанического института АН СССР в Ленинграде
Не менее излюбленными в комнатном цветоводстве являются долго цветущие сенполии. В Америке обществом любргтелей африканских фиалок зарегистрировано более 1200 сортов, относящихся к гибридам сенполии фиалко-цветковой (Saintpaulia ionantha). Все виды сенполии (около 20) произрастают в Восточной Африке в Усамбарских и Улугурских горах вблизи водопадов, на террасах рек, в окружении водяной пыли или тумана. В цветоводческих хозяйствах и ботанических садах многих стран культивируют ахименесы, колерии, колумней, эписции, кодонанту грациозную (Codonanthe gracilis), рехштейнерию пурпуровую (R. cardinalis, табл. 61), стрептокарпусы, эсхинантусы и др.

Таблица 61. Геснериевые: 3 - рехштейнерия беловолосистая (Rechsteineria leucotricha), оранжереи Ботанического института АН СССР в Ленинграде

Таблица 61. Геснериевые: 4 - рехштейнерия пурпуровая(R. Cardinalis), оранжереи Главного ботаческого сада в Москве
Семейство подорожниковые (Plantaginaceae) (А. Е. Бородина)
В семействе подорожниковые 3 рода и около 265 видов, распространенных в умеренных областях обоих полушарий с немногими видами в тропиках.
Подорожниковые обычно многолетние или однолетние травы, иногда кустарнички. Листья обычно очередные или реже супротивные. Цветки в головках или колосьях, актиноморфные, обоеполые. Чашечка 4-лопастная или 4-раздельная. Венчик обычно 4-лопастный, редко 3-лопастный. Тычинок обычно 4 или редко 1-2, прикрепленных к трубке венчика. Гинецей из 2 плодолистиков с тонким столбиком с 2-лопастным рыльцем; завязь верхняя, 2-гнездная или 1-гнездная. Эндосперм целлюлярный. Плод - перепончатая коробочка, раскрывающаяся поперечной щелью, или ореховидный, окруженный остающейся чашечкой. Семена обычно с маленьким зародышем и мясистым эндоспермом.
Подорожник (Plantago) - самый большой (260 видов) род семейства, распространенный в умеренных областях обоих полушарий и лишь отдельные виды - в тропиках. Большинство подорожников - многолетние растения, однако встречаются и однолетники, например подорожник лузитанский (P. lusitanica) или подорожник заячий (P. lagopus) из Западного Средиземноморья, некоторые подорожники Западной Азии и Южной Америки. В пустынях Северной Африки встречается своеобразный кустарниковый подорожник мавританский (P. mauritanica, рис. 231) с головчатыми соцветиями и стеблями, олиственными пучками линейных листьев.
Рис. 231. Подорожник мавританский (Plantago mauritanica): 1 - общий вид растения; 2 - цветок в начальной стадии цветения; 3 - цветок в последующей стадии цветения; 4 - плод; 5, 6 - семя
Другой род - побережник (Littorella) - распространен в Европе, на островах Атлантического океана и в умеренных районах Южной Америки (3 вида). Побережник озерный (L. lacustris) встречается по берегам прудов, озер и морским побережьям на затопляемых водой местах, в результате чего у него выработались две формы - водная и сухопутная. Сухопутная форма имеет розетку узких дорсивентральных листьев длиной до 5 см. Из пазух листьев на длинной ножке выходит один мужской цветок и два сидячих женских, которые созревают до появления тычинок. Водная форма внешне похожа на шильник, или полушник (Isoetes, см.: Жизнь растений, т. 4, с. 118), имеет розетку цилиндрических более длинных листьев и размножается только вегетативно.
Третий род семейства - монотипный род бугиера (Bougueria)- эндемичен для Анд Южного Перу, Боливии и Северной Аргентины.
Для большинства подорожниковых характерно ветроопылеыие. Сразу после растрескивания пыльников, пока еще из них не высыпалась вся пыльца, некоторые подорожники (например, подорожник ланцетный - P. lanceolata) посещают пчелы и мухи. Насекомые садятся на колоски, ползают по ним и на лапках собирают пыльцу, а при перелете с соцветия на соцветие осуществляют перекрестное опыление. Если же пыльцы в пыльниках больше нет, то насекомые на соцветия не садятся.
Подорожники находят применение в народной и официальной медицине. Семена подорожников при намокании и отваривании дают массу слизи, которую используют в медицине и в промышленности. Широко известны семена подорожника блошиного (P. psyllium), распространенного от Западного Средиземноморья до Индии, которые используют в производстве шелковых и хлопчатобумажных тканей, для глянцевания цветной бумаги, в типографском деле, для приготовления мороженого и как наркотическое средство. Этот и близкий к нему вид подорожник яйцевидный (P. ovata) из Ирана культивируют в Индии и Японии, собирая по 2 урожая в год; их семена экспортируют в западноевропейские страны и в США.
Для приготовления салатов и гарниров в Европе издавна культивировался подорожник оленерогий (P. coronopus) (его название связано с зубчатой формой края листа). При посеве ранней весной урожай этого подорожника собирают через три месяца. Другие подорожники, в частности наш подорожник ланцетный, используют в приготовлении кормовых травяных смесей.
Семейство пузырчатковые (Lentibulariaceae) (Е. А. Земская)
Все представители этого семейства, насчитывающего 4 рода и более 180 видов, распространенных во всех частях света,- плотоядные растения. Это многолетние и однолетние наземные, реже эпифитные травы с очередными или собранными в розетки листьями, растущие в воде, на болотах и влажных местах. Цветки у пузырчатковых зигоморфные, обоеполые, собранные в кистевидные или колосовидные соцветия или одиночные, как, например, у жирянки (Pinguicula). Чашечка у пузырчатки двулопастная или 4-5-лопастная, остающаяся при плодах. Двугубый венчик с короткой трубкой снабжен при основании шпорцем или мешочковидным придатком. Верхняя губа венчика двулопастная, нижняя - трехлопастная или цельная. Тычинки две, с плоскими расширенными нитями, прикрепленными при основании венчика; пыльники одно- или реже двугнездные. Гинецей из 2 плодолистиков; завязь верхняя, одногнездная, с многочисленными семязачатками; столбик очень короткий, с двулопастным рыльцем, с неравными лопастями. Плод - одногнездная коробочка шаровидной или яйцевидной формы, иногда удлиненная, раскрывающаяся двумя или четырьмя створками и разрывающаяся неправильно; редко плод односемянный, нераскрывающийся. Семена мелкие, со слабо развитым эндоспермом.
Перекрестное опыление у пузырчатковых в умеренной зоне осуществляется насекомыми (пчелами, мухами, журчалками), которых привлекает находящийся в шпорце нектар. Указателем нектара для насекомых у некоторых видов жирянки являются белые или желтые пятна на нижней губе, а у пузырчатки обыкновенной (Utricularia vulgaris, рис. 232) - оранжевые полоски на вздутии нижней губы. Так как у жирянок рыльце нависает над пыльниками, насекомое сначала касается головкой воспринимающей поверхности рыльца, оставляя на ней принесенную пыльцу, а затем пыльников, пыльца из которых переносится на другие цветки. У пузырчатки обыкновенной вздутие на нижней губе, служащей местом посадки, плотно закрывает вход в цветок. Под тяжестью насекомого нижняя губа отклоняется вниз, открывая вход в цветок, а тычинки, плотно прилегающие к верхней губе, касаются головы и спинки насекомого. На нижней лопасти рыльца имеются жесткие сосочки, которые, действуя подобно зубцам гребенки, при движении над ними насекомого снимают пыльцу с его волосков. При отсутствии насекомого может происходить самоопыление, благодаря тому что нижняя лопасть рыльца, нависающая над пыльниками, завертывается таким образом, что поверхность рыльца касается пыльников. У жирянок и пузырчаток встречаются и клейстогамные цветки.
Рис. 232. Пузырчатка обыкновенная (Utricularia vulgaris): 1 - веточка; 2 - пузырек с добычей; 3 - продольный разрез пузырька (а - антенны, б - щетинки, в - клапан, г - верхняя губа, д - нижняя губа, е - железистые волоски и желёзки)
Семена большинства представителей семейства распространяются птицами, к ножкам которых они прилипают, или разносятся течением воды. Птицы разносят также части растений и зимующие почки. У некоторых видов, как, например, у пузырчатки жесткой (U. rigida), поверхность семян покрыта клейким веществом, что препятствует смыванию их течением. Семена эпифитной пузырчатки мелкополосчатой (U. striatula) снабжены крючочками, которыми они цепляются за мох. Эпифитная пузырчатка Манна (U. mannii) образует несколько тысяч очень мелких семян, сходных по форме с семенами орхидных и распространяющихся ветром.
Самый большой род в семействе - пузырчатка - охватывает более 200 видов, распространенных в основном в восточной части тропической Южной Америки, на севере Центральной Африки, а также в Юго-Восточной Азии и тропической Австралии. Среди представителей рода встречаются наземные формы с цельнокрайними листьями, распространенные в тропической зоне, и водные с рассеченными или перистыми листьями, произрастающие преимущественно в умеренной и отчасти в тропической зоне. В Европе известны только 6 видов, а в нашей стране встречается 8 видов.
Род получил свое название благодаря наличию у него ловчих пузырьков. Пузырьки бледно-зеленоватые, косояйцевидной или шаровидной формы, диаметром от 2 до 5 мм, располагаются на листьях, реже на стеблях растений (см. рис. 232). В пузырьке имеется ротовое отверстие, по краям которого находятся длинные разветвленные волоски, названные Дарвином антеннами, и несколько жестких щетинок. Нижний край отверстия (нижняя губа) утолщен и вдается внутрь пузырька плотным валиком. От верхнего края (верхняя губа) отходит тонкий и очень эластичный клапан, наружная поверхность которого несет большое количество желёзок, выделяющих клейкое вещество и сахар, служащие приманкой для маленьких животных, главным образом личинок, реже взрослых мелких рачков, дафний, циклопов, а также личинок комаров, мелких червей и инфузорий. Иногда в ловушку случайно попадают даже мальки рыб. Клапан открывается при легчайшем прикосновении, и животное с током воды засасывается внутрь. Попав в полость, животное не может выйти обратно, так как клапан тотчас же закрывается и не может быть открыт давлением изнутри. Через некоторое время животное погибает и служит пищей для растения. Продукты пищеварения поглощаются находящимися в пузырьках всасывающими клетками. Внутренняя поверхность пузырька покрыта железистыми волосками, которые выделяют ферменты, участвующие в процессе переваривания захваченных животных. Среди водных форм пузырчатки встречаются свободно плавающие растения, на всех стадиях развития живущие под водой, не прикрепляясь ко дну водоема. К ним относится наиболее широко распространенный вид пузырчатка обыкновенная - растение с погруженными в воду крупными, многократно перистыми листьями, несущими ловчие пузырьки, и выступающим над водой цветоносом с кистью желтых цветков. Некоторые из водных видов, как, например, пузырчатка Брема (U. bremii), растут погруженными в воду, образуя бесцветные побеги, которыми растение прикрепляется к субстрату. Водные формы пузырчатки, произрастающие в умеренной зоне, обычно цветут и плодоносят спорадически, размножаясь в основном вегетативно.
Среди тропических видов встречаются эпифиты - растения с крупными листьями и яркими цветками. К ним относятся бразильские виды пузырчатка почколистная (U. reniiformis), растущая на болоте, на мхах, и пузырчатка лотосолистная (U. nelumbifolia), живущая в скоплениях воды в розетках листьев бромелиевых.
Представители другого рода - жирянка, включающего около 45 видов, распространены в Европе, Северной Азии и Северной Америке. Растут они по берегам ручьев, на сфагновых болотах, некоторые из них как эпифиты на мхах и деревьях, например жирянка ворсистая (P. villosa) и жирянка древесная (P. lignicola). В экваториальной зоне виды жирянки поднимаются в прохладные пояса высоких гор. На территории СССР произрастают 5 видов. Из них наиболее широко распространена жирянка обыкновенная (P. vulgaris, рис. 233). Жирянка - единственный род в семействе, представители которого имеют настоящие корни. Пузырьков для ловли насекомых у жирянки нет и эта функция выполняется непосредственно листьями. Листья цельные, относительно толстые, мясистые, светло-зеленые, широкоэллиптические или сравнительно длинные и узкие, образуют прикорневую розетку. На верхней стороне листа находятся многочисленные желёзки на ножках, выделяющие сахаристую слизь, и сидячие желёзки, которые выделяют слизь, содержащую ферменты, переваривающие добычу. Привлекаемые блестящими капельками слизи, насекомые опускаются на лист и прилипают к его поверхности. Раздражение насекомыми поверхности листа вызывает медленное его скручивание, и захваченное таким образом насекомое переваривается.
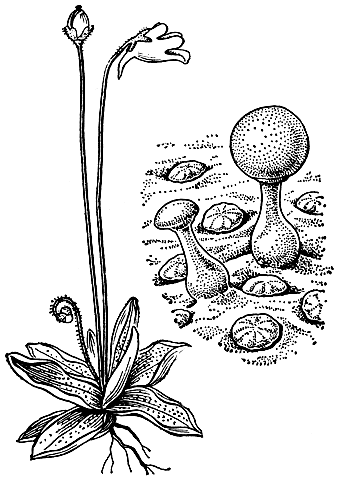
Рис. 233. Жирянка обыкновенная (Pingvicula vulgaris): справа - слизевые желёзки на ножках и пищеварительные сидячие желёзки на поверхности листа
Наиболее сложно устроена ловушка у представителей близкого к роду пузырчатка по строению цветков и плодов - рода генлисея (Genlisea), насчитывающего 15 видов, распространенных в Центральной Америке, Вест-Индии, Африке и на Мадагаскаре. Ловушки генлисеи состоят из отдельных пузырьков с длинными трубочками, оканчивающимися двумя спирально скрученными шиловидными отростками, полость которых открывается посредством длинной щели, разделенной перегородками на множество мелких воронкообразных отверстий. Внутренние стенки отростков покрыты многочисленными щетинками, направленными вниз и препятствующими выходу животных обратно.
Близким к пузырчатке по строению вегетативных органов и устройству ловушки является род полипомфоликс (Polypompholyx).
Практическое значение пузырчатковых невелико. Жирянку обыкновенную издавна применяли в народной медицине. Экстракт жирянки используют в настоящее время как эффективно действующее средство при лечении астмы. Листья жирянки содержат фермент, превращающий свежее молоко в своеобразную тягучую массу (тэтмиелк) - излюбленное блюдо в Северной Скандинавии. Некоторые эпифитные виды пузырчатки с красивыми цветками культивируют в оранжереях вместе с орхидными.
Семейство миопоровые (Myoporaceae) (Н. Н. Цвелев)
К близкородственному норичниковым семейству миопоровых (рис. 234) принадлежат кустарники, реже небольшие деревья или полукустарники с цельными очередными, реже супротивными, ланцетными или яйцевидными листьями без прилистников. Большинство миопоровых принадлежит к числу ксерофитов, и потому многие из них имеют обильное опушение из железистых или простых волосков. Нередко встречаются также точечные желёзки на листьях и чашечках, особенно у видов миопорума (Myoporum). Наиболее ксерофильные австралийские кустарники эремофила горбатолистная (Eremophila gibbifolia) и миопорум соляпколистный (M. salsoloides) имеют сильно редуцированные листья и напоминают по облику некоторые виды солянок.
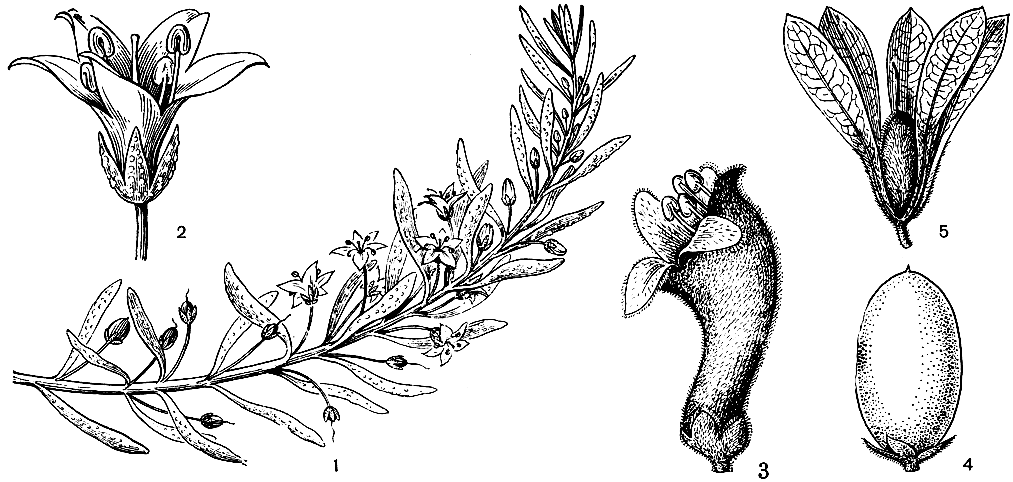
Рис. 234. Миопоровые. Миопорум низкий (Myoporum humile): 1 - цветущая ветвь; 2 - цветок. Эремофила длиннолистная (Eremophila longifolia): 3 - цветок; 4 - плод. Эремофила супротивнолистная (E. oppositifolia): 5 - плод с чашечкой
Цветки миопоровых обоеполые, пятичленные, актиноморфные (у миопорума) или зигоморфные (у эремофилы), обычно одиночные и расположенные в пазухах листьев, реже образующие небольшие верхоцветные соцветия, также расположенные в пазухах листьев. Чашечки часто почти до основания рассечены на 5 лопастей; венчики обычно короткотрубчатые или колокольчатые, нередко густоволосистые. Из 5 прикрепленных к трубке венчика тычинок одна часто видоизменена в стаминодий. Гинецей образован 2 плодолистиками, с простым столбиком и цельным или едва двулопастным рыльцем. 2 первичных гнезда верхней завязи могут делиться ложными перегородками на несколько частей, и завязь становится 3-10-гнездной с 1-8 семязачатками в каждом гнезде.
3 рода и около 140 видов миопоровых распространены преимущественно в Австралии, где они играют заметную роль в ксерофильных кустарниковых зарослях. Кроме того, немногие виды этого семейства встречаются в Новой Зеландии, на Новой Гвинее, в Восточной Азии, Южной Африке, на Маскаренских и Гавайских островах, в Вест-Индии.
Богатые нектаром цветки миопоровых нередко имеют медовый запах и опыляются насекомыми, преимущественно перепончатокрылыми.
Мясистые, костянковидные плоды обычно распространяются птицами - эндозоохорно. Однако имеются виды (например, эремофила супротивнолистная - Eremophila oppositifolia), с крупными, сильно расширенными долями остающейся при плодах чашечки и распространяющиеся главным образом с помощью ветра (анемохорно). Плоды у таких видов обычно мелкие и имеют слабомясистый экзокарпий.
Хозяйственное значение миопоровых невелико. Некоторые виды культивируются в качестве декоративных растений в садах и парках, а в умеренно теплых широтах - в оранжереях. Миопорум плоскоплодный (Myoporum platycarpum) дает смолу, близкую по возможностям использования к сургучу, а новозеландский миопорум яркий (M. laetum), миопорум гавайский (M. sandwicense) и австралийская эремофила Митчелла (E. mitchellii) доставляют ценную древесину. Некоторые виды миопорума содержат ядовитые вещества с гемолитическим действием (разрушают эритроциты крови) и могут вызвать отравление или падеж скота.
Семейство акантовые (Acanthaceae) (С. С. Морщихина)
Семейство акантовых объединяет около 2600 видов, относящихся примерно к 250 родам. В основном это многолетние травянистые растения и полукустарники, широко распространенные в тропиках и субтропиках обоих полушарий. Около 10 родов, включая большинство крупных, представлены в обоих полушариях. Преобладают в семействе роды небольшие, с ограниченным ареалом. В обширном ареале акантовых выделяются четыре центра наибольшей концентрации родов: Южная и Юго-Восточная Азия, тропическая Африка и Мадагаскар, Южная Америка и Центральная Америка с островами Карибского моря. Несколько видов произрастает на юго-востоке США, 3 вида аканта (Acanthus, табл. 62) встречаются в Южной Европе.

Таблица 62. Акантовые: 3 - акант падуболистный (Acanthus ilicifolius), Пуна
Наряду с крупными травами среди акантовых существуют почти бесстебельные растения с прикорневой розеткой листьев, обычны также и кустарники высотой 2-4 м. В подсемействах тунбергиевых (Thunbergioideae) и мендонкиевых (Mendoncioideae) представлены лианы с более или менее одревесневающим стеблем. Встречающаяся на Кубе оплония карликоволистная (Oplonia nannophylla) представляет собой кустарничек высотой 4-8 см, а одна из разновидностей оплонии вооруженной (O. armata) на Ямайке вырастает небольшим деревом. Кроме этого, лишь два небольших южноамериканских рода - трихантера (Trichanthera) и бравезия (Bravaisia) - представлены деревьями. Бравезия целънокрайняя (B. integerrima), растущая в болотистых лесах и на сырых лугах,- 18-метровое дерево с ходульными придаточными корнями у основания ствола.
Для многих акантовых характерно простое или железистое опушение, а также присутствие цистолитов в наземных вегетативных органах.
Листья у акантовых простые, супротивные, иногда очень крупные, как у некоторых видов аканта.
Цветки акантовых обоеполые, зигоморфные, собраны в различного рода соцветия, для которых всегда характерно наличие крупных и иногда ярких брактей. Преобладают соцветия типа тирса и различные его модификации. Вследствие недоразвития соцветий у некоторых акантовых наблюдается одиночное расположение цветков в пазухах листьев. Иногда укорочение оси соцветия ведет к появлению головчатых соцветий.
Чашечка из 5 (редко из 4 или 3) сросшихся в нижней части долей, у видов тунбергии (Thunbergia, табл. 62) редуцирована, и ее роль выполняют крупные прицветнички, полностью окутывающие бутон. Лепестки срастаются в трубку различной формы, заканчивающуюся 5-лопастным обычно неправильным отгибом, или венчик двугубый, с двухлопастной верхней и трехлопастной нижней губой. Иногда одна из них не доразвивается. Так, у видов аканта, блефариса (Blepharis) и других близких к ним родов венчик состоит только из нижней губы. Тычинок обычно 4 (редко 5), расположенных попарно; при этом одна пара часто имеет более длинные тычиночные нити. Иногда развиваются только две фертильные тычинки, и в таких цветках можно обнаружить 1-3 стаминодия. Пыльники, раскрывающиеся продольной щелью, вокруг которой нередко имеется опушение, задерживающее пыльцу. Иногда пыльники снабжены внизу шпорцевидными выростами. Связник часто расширен, и пыльники могут располагаться несимметрично, на разной высоте (рис. 235). У некоторых видов нижний пыльник меньше или совсем не развит; такие тычинки с одним пыльником имеются, например, у видов аканта. Гинецей из двух плодолистиков. Завязь окружена нектарным диском верхняя, двугнездная, с 2-10 или многочисленными анатропными или амфитропными семязачатками в каждом гнезде. Столбик простой, нитевидный, несет обычно двулопастное рыльце. Форма и размеры лопастей варьируют, нередко одна лопасть расположена выше другой, иногда одна из них не развита (рис. 235). У видов рода оплония наблюдается гетеростилия. У акантовых преобладает протандрия, и для них характерно насекомоопыление. Опылителей привлекают яркий, разнообразной окраски венчик и прицветники соцветия, обильный нектар, а иногда и экстрафлоральные нектарные желёзки на прицветничках, прицветниках, черешках листьев и на стеблях. Наибольшее значение в опылении акантовых, по-видимому, имеют виды пчел. Пыльники, обращенные друг к другу открывающейся опушенной стороной, держат щеткой волосков пыльцу, пока пчела, пробираясь за нектаром, не раздвинет тычинки. Тогда тело насекомого густо обсыпается пыльцой. Некоторые акантовые обнаруживают высокую специализацию к определенным видам опылителей. Таким примером является тунбергия крупноцветковая (Thunbergia grandiflora) из лесов Ассама (Индия) и Бирмы. По наблюдениям ван дер Пейла (1954), крупные голубые цветки ее доступны лишь пчелам-плотникам из рода ксилокопа (Xylocopa latipes). Широкие тычиночные нити смыкаются, закрывая доступ в нижнюю часть цветка. Крупное сильное насекомое, стремясь к нектару, раздвигает их, а жесткие шпорцы пыльников, прижатые спинкой насекомого, вспарывают пыльники, обсыпая пчелу пыльцой. Более мелкие виды ксилокопы не в силах раздвинуть тычинки и редко посещают эти цветки. У тунбергии прямостоячей (T. erecta) и других пчелы облегчают себе добычу нектара, прокусывая венчик снаружи. На цветках тунбергии крупноцветковой их атакуют муравьи, постоянно посещающие ыектарные желёзки на прицветничках и редуцированной чашечке, валиком окаймляющей основание венчика. Муравьи вынуждают пчел-плотников забираться внутрь цветка и таким образом способствуют их опылению.

Таблица 62. Акантовые: 1 - тунбергия крылатая (Thunbergia alata), оранжереи Главного ботанического сада в Москве

Таблица 62. Акантовые: 2 - джастисия мясокрасная (Justicia carnea), оранжереи Главного ботанического сада в Москве
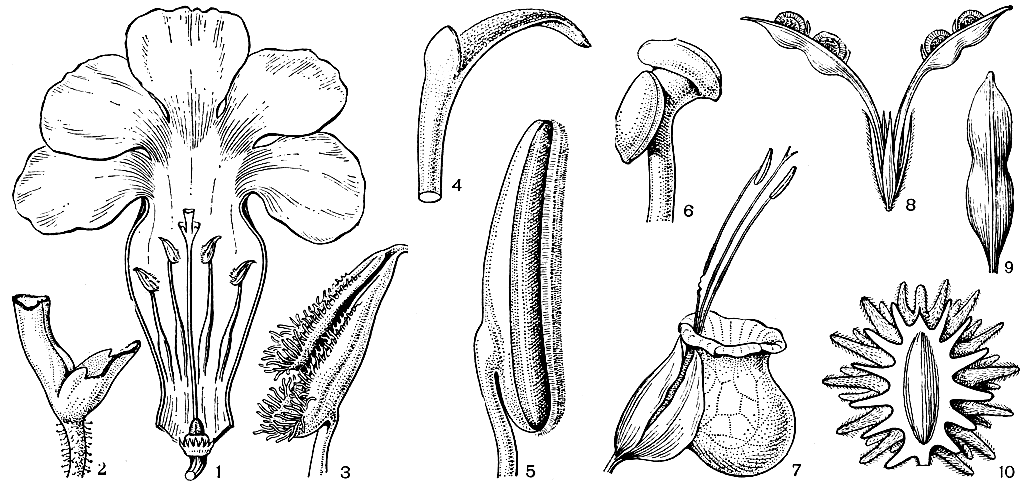
Рис. 235. Акантовые. Тунбергия родственная (Thunbergia affinis): 1 - цветок в продольном разрезе; 2 - рыльце; 3 - тычинка. Руэллия приятная (Ruellia graecizans): 4 - рыльце. Акант мягкий (Acanthus mollis): 5 - тычинка с опушенными пыльниками. Джастисия американская (Justicia americana): 6 - тычинка с асимметрично расположенными пыльниками. Лоутеридиум Доннелл-Смита (Louteridium donnellsmithii): 7 - цветок. Азистазиггангекая (Asystasia gangetica): 8 - раскрывшийся плод. Джастисия разноплодная (Justicia hetero carpa): 9 - раскрывающаяся коробочка; 10 - продольный разрез нераскрывающегося плода
У видов афеландры (Aphelandra), якобинии (Jacobinia) и санчесии (Sanchezia) из лесов тропической Америки в опылении участвуют колибри. Причудливые, с мешковидно вздутой трубкой цветки лоутеридиума Дон-нелл-Смита (Louteridium donnellsmithii) - кустарника из Центральной Америки - посещают и, видимо, опыляют летучие мыши.
У некоторых видов барлерии (Barleria), pyэллии (Ruellia, табл. 62), блехума (Blechum), стенандриума (Stenandrium) и других родов известна клейстогамия. У видов блефариса установлен апомиксис.

Таблица 62. Акантовые: 5 - руеллия приятная (Ruellia graecizans), оранжереи Ботанического института АН СССР в Ленинграде
Характерный для акантовых тип плода - двугнездная коробочка, раскрывающаяся двумя створками, в каждой из которых остается половина расщепившейся перегородки (см. рис. 235). Исключение составляют представители подсемейства мендонкиевых, у которых во время развития цветка различимы 2 гнезда завязи, но одно из них дегенерирует и образуется одногнездный нераскрывающийся костянковый плод. У джастисии разпоплодной (Justicia heterocarpa) из Восточной Африки известно наряду с обычными двугнездными коробочками образование нераскрывающихся плодов с четырьмя зубчатыми крыловидными выростами (рис. 235, 9, 10). Эти плоды могут, очевидно, и рассеиваться ветром, и цепляться за шерсть животных.
Господствующий способ распространения семян у акантовых - разбрасывание их в стороны при растрескивании коробочки, чему способствует охватывающий семя снизу крючковидный вырост семяножки, так называемый ретинакулум или якулятор. Французский ботаник Ив Сель (1967, 1969) подробно исследовал механизм раскрытия плода. Открытие коробочки происходит за счет неравномерного высыхания слоев клеток в перегородке. Наружные слои сильно ссыхаются и сокращаются, внезапно разрывая перегородку и весь плод посередине, по центральной жилке плодолистиков. Коробочка открывается толчком, прижатые концы крючковидных выростов с силой отгибаются, действуя как рычаг, и семена, подобно пущенному спортивному диску, выбрасываются в две стороны иногда на десятки метров.
У некоторых видов блефариса, барлерии и других родов, обитающих в засушливом климате, раскрытие зрелой высохшей коробочки происходит только в дождь, когда богатые пектином клетки кончика плода набухают, сцепление их ослабевает и плод так же толчком открывается, разбрасывая семена. Семена многих пустынных видов покрыты или окаймлены шелковистыми гигроскопичными волосками или чешуйками, которые при намокании pac топыриваются и набухают, становясь клейкими. Волоски ориентируют семя корешком зародыша к земле и приклеивают их к частицам почвы так, чтобы не смыло водой и не унесло ветром. Прорастают семена пустынных акантовых очень быстро. У блефариса крупного (Blepharis grossa) за ночь после намачивания вырастает корень длиной около 3 см. В сухом состоянии семена таких видов могут долго сохранять всхожесть. П. Г. Мейеру (1961) удалось прорастить в Мюнхенском ботаническом саду семена петалидиума щетинистого (Petalidium setosum), пролежавшие в гербарии 12 лет.
Благодаря разбрасыванию семян виды акантовых часто образуют сплошные заросли почти без примеси других растений и играют значительную роль в травянистом покрове влажных тропических лесов. Акантовые встречаются во всех тропических и субтропических формациях, нередки они в открытых ксерофильных лесах, саваннах и кустарниковых зарослях. Во влажных лесах на склонах Кордильер они поднимаются до 1600-1800 м над уровнем моря, а в Западных Гималаях даже до 2000 м. В то же время акантовые можно найти на морских побережьях: в мангровах восточного полушария образует колючие заросли акант падуболистный (A. ilicifolius, табл. 62). Колючие ксерофильные виды блефариса, петалидиума и других родов встречаются в Сахаре, пустынях Южной Африки и Западной Азии. Виды гигрофилы (Hygrophila) растут по берегам рек, ручьев и на болотах, частично затопляются и живут как водные растения. Некоторые виды руэллии, джастисии и других родов, занесенные за пределы своего ареала, стали сорняками во многих тропических и субтропических странах. Семейство акантовых делят на 5 подсемейств: нельсониевые (Nelsonioideae), которые, подобно норичниковым, имеют многочисленные семязачатки в гнездах завязи; тунбергиевые (Thunbergioideae), имеющие по 2 семязачатка в каждом гнезде; мендонкиевые (Mendoncioideae), у которых одно гнездо завязи не развивается и образуются костянковидные плоды; акантовые (Acanthoideae) с несколькими или двумя семязачатками в гнездах завязи и черепитчатым сложением долей венчика в бутоне; руэллиевые (Ruellioideae), для которых характерно скрученное сложение листочков околоцветника (табл. 62).

Таблица 62. Акантовые: 4 - эрантемум Уата (Eranthemum wattii), оранжереи Ботанического института АН СССР в Ленинграде
Акантовые не имеют большого практического значения. Некоторые виды используют в народной медицине и выращивают как декоративные растения. Форма листьев средиземноморских видов аканта лежит в основе архитектурных украшений коринфских и композитных капителей, орнаментации фризов, карнизов и других элементов зданий.
Семейство хвостниковые (Hippuridaceae) (А. П. Белавская)
К семейству относится только один род водяная сосенка (Hippuris), состоящий из одного очень полиморфного вида. Водяная сосенка - растение сырых мест и мелководий умеренных широт северного полушария. В СССР широко распространена водяная сосенка обыкновенная (H. vulgaris, рис. 236), которая встречается от тундры, где ее охотно поедают олени, до Дальнего Востока и Средней Азии. Это небольшое (высотой 15-20 см) стройное растение с полым неветвящимся стеблем и мутовчатыми линейными листьями. Надводная часть стебля несет мелкие жесткие листья (1-4 см), а подводные листья, тонкие и нежные, могут вытягиваться до 10 см в длину. Гетерофиллия, типичная вообще для водных растений, связана у водяной сосенки с глубиной и, следовательно, с интенсивностью освещения. Выращивая ее в разных условиях, можно получать разные типы листьев: корневищные, ювенильные водные, взрослые водные, ювенильные воздушные и взрослые воздушные. Цветки водяной сосенки упрощены до предела: вместо околоплодника - небольшой валик, тычинка одна, завязь с одним нитевидным рыльцем. Опыление совершается ветром. Плод - костянка с тонким околоплодником. Распространяются плоды водой или птицами, прилипая к ним вместе с илом, чем и объясняется широкое распространение этого растения. Водяная сосенка растет одинаково хорошо и на заболачивающихся низинах, и на мелководьях до глубины 30-40 см, хотя известны ее находки с глубин 1,5-2 м.
Рис. 236. Сланоягодниковые и хвостнпковые. Уруть колосистая (Myriophyllum spicatum): 1 - общий вид растения; 2 - женский цветок; 3 - мужской цветок. Водяная сосенка обыкновенная (Hippuris vulgaris): 4 - наземная и мелководная форма; 5 - глубоководная форма; 6 - цветок
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'