Порядок колокольчиковые (Campanulales) (А. Л. Тахтаджян)
Семейство колокольчиковые (Campanulaceae)
В семействе более 80 родов и примерно 2300 видов. Колокольчиковые произрастают преимущественно во внетропических областях Старого и Нового Света, но некоторые роды представлены и в тропических странах, особенно в горных районах.
Большинство колокольчиковых - травы, многолетние или реже однолетние, прямостоячие или иногда вьющиеся, наземные или редко водные или эпифиты. Есть среди них также полукустарники и даже древовидные формы. У некоторых родов корни клубневидно утолщенные. Листья очередные или иногда супротивные (кодонопсис - Codonopsis, канарина - Canarina и некоторые другие), редко мутовчатые (например, у островскии - Ostrowskia), цельные, зубчатые, лопастные или редко перистые. Характерно наличие млечников во флоэме листа и стебля, отсутствующих, однако, у рода сфеноклея (Sphenoclea). У большинства родов углеводы откладываются в форме инулина. Цветки обычно довольно крупные, в верхоцветных или чаще бокоцветных соцветиях, иногда в густых головках, окруженных оберткой (букашник - Jasione и некоторые виды эдрайантуса - Edraianthus) или в густых колосьях (сфеноклея и некоторые виды кольника - Phyteuma). Нередко цветки одиночные, верхушечные или пазушные (у новогвинейского рода филлохарис - Phyllocharis - пазушные цветки эпифилльные, приросшие к средней жилке листа), обычно обоеполые, очень редко однополые (у некоторых видов колокольчика - Campanula, легузии - Legousia и лобелии - Lobelia двудомные), актиноморфные или более или менее зигоморфные, большей частью 5-члеыные, в большинстве случаев с 2 прицветничками. Лопасти чашечки большей частью свободные, обычно остающиеся. Лепестки более или менее сросшиеся, редко свободные почти до основания, как у кольника и мишоксии (Michauxia). Тычинки в одинаковом числе с лепестками, свободные или чаще прикрепленные к основанию венчика, к диску или реже к трубке венчика нити свободные или более или менее сросшиеся, у основания обычно более или менее расширенные; пыльники интрорзные, свободные (и тогда в бутоне большей частью склеены в трубку) или же довольно плотно спаяны в трубку. Гинецей из 5-2 плодолистиков, с простым или чаще лопастным столбиком, который в верхней части, включая наружные стороны лопастей рыльца (а у монотипного кавказского рода гаделлии - Gadellia - почти исключительно лопасти рыльца), обычно густо покрыт одноклеточными волосками; завязь обычно нижняя, но иногда более или менее полунижняя, как у кодонопсиса, валенбергии (Wahlenbergia), платикодона (Platycodon) и некоторых видов лобелии, или даже верхняя, как у азиатского рода цианантус (Cyananthus) и некоторых видов кодонопсиса и валенбергии; 2-5-, редко 6-10-гнездная, иногда почти 1-гнездная, обычно с многими семязачатками в каждом гнезде, чаще всего на толстых плацентах; у южноафриканского рода мерсьера (Merciera) в 1-гнездной или не полностью 2-гнездной завязи имеется всего 4 базальных семязачатка, а у другого южноафриканского рода - сифокодон (Siphocodon) - семязачатки в 3-гнездной завязи висячие с верхушки полости гнезд. У верхушки завязи под основанием столбика находится нектарный диск, который у евразиатского рода бубенчик (Adenophora) бывает цилиндрическим, чашеобразным или кольцеобразным. Плоды колокольчиковых очень разнообразного типа. Чаще всего плод - коробочка, иногда спирально скрученная (южноафриканский род призматокарпус - Prismatocarpus), вскрывающаяся верхушечными, боковыми или базальными створками или порами, или даже неправильно; реже плод - ягода, редко криночка (сфеноклея и паришелла - Parishella) или орехообразный (перакарпа - Peracarpa). Семена мелкие, обычно многочисленные, с прямым зародышем и мясистым и обычно обильным эндоспермом.
Большинство колокольчиковых мезофиты, но многие из них гигрофиты или, напротив, ксерофиты. Есть среди них и немногие водные растения, как растущая у нас в европейской части лобелия Дортманна (Lobelia dortmanna) или североамериканская монотипная хаувелия водная (Howellia aquatica). Большого разнообразия достигли у них формы роста. Хотя большинство колокольчиковых - травы, но среди них довольно много древесных, точнее, древовидных форм. Но и сами травянистые колокольчиковые достаточно разнообразны. Большинство их прямостоячие, но немало и лиан, как некоторые виды кодонопсиса (например, встречающийся у нас в Уссурийском крае кодонопсис мелковолосистый - C. pilosula) или африканского рода цифия (Cyphia). Есть среди колокольчиковых очень интересные геофиты с клубневидно утолщенным гипокотилем и главным корнем. Таковы, например, роды островския, платикодон, канарина или виды кодонопсиса и цифии. Есть крошечные, цочти бесстебельные растения, как очень редкая малезийская лобелия Арчболда (L. archboldiana, рис. 238) и некоторые другие виды этого рода. Встречаются также эпифиты (эпифитный образ жизни часто ведет африканская канарина Эмина - C. eminii). Очень разнообразны древовидные колокольчиковые. Оригинальный вид имеет эндемичное для Азорских островов канделябровидное карликовое деревце азорина Видаля (Azorina vidalii), достигающая в высоту 2 м. Гораздо более внушительных размеров достигают древовидные лобелии на Гавайских островах; некоторые из них достигают 15-метровой высоты. Причудливый вид имеет растущая на Гавайских островах древовидная брайамия замечательная (Brighamia insignis, рис. 238), невысокий мясистый стебель которой увенчан розеткой листьев. Некоторые южноафриканские роды (роэлла - Roella, ригиофиллум - Rhigiophyllum) имеют эрикоидный облик. Среди древовидных колокольчиковых есть также лианы, как некоторые виды неотропического рода центропогон (Centropogon).
Уже начиная с классической работы К. Шпренгеля (1793), опыление цветков колокольчиковых служит объектом интенсивных исследований. Но, несмотря на это, далеко не все группы колокольчиковых изучены в этом отношении достаточно хорошо и на долю будущих исследователей остается много нерешенных вопросов.
Для подавляющего большинства колокольчиковых характерно перекрестное опыление, которое обеспечивается прежде всего обычно очень сильно выраженной у них протандрией. Опылителями являются главным образом крупные пчелиные и бабочки, а также пилильщики, жуки и другие насекомые. Но немало среди колокольчиковых и орнитофильных растений, особенно в Америке. Африке и на Гавайских островах. Одной из наиболее характерных особенностей механизма опыления у колокольчиковых является подача пыльцы столбиком, а не непосредственно пыльником. Это довольно необычное для цветковых растений явление, кроме представителей порядка колокольчиковых, известно еще только для некоторых протейных, калицеровых и сложноцветных.
Семейство колокольчиковых подразделяется на 4 подсемейства - собственно колокольчиковые (Campanuloideae), цифиевые (Cyphioideae), сфеноклеевые (Sphenocleoideae) и лобелиевые (Lobelioideae). Некоторые ботаники считают эти группы трибами, т. е. придают им более низкий таксономический ранг. Другие же, наоборот, рассматривают их в ранге самостоятельных семейств.
Подсемейство собственно колокольчиковые самое крупное по количеству родов, но по числу видов оно уступает лобелиевым. В зависимости от взглядов разных ботаников на объем рода, в нем насчитывают около 50 родов. Что касается числа видов, то, по современным данным, оно доходит примерно до 1000. Для подсемейства характерны актиноморфные (лишь иногда слегка зигоморфные) цветки и обычно свободные (только в бутоне обычно склеенные) пыльники, которые лишь изредка плотно соединены как бы в трубку или даже сросшиеся (рис. 237). Тычинки прикреплены у основания венчика или к нектарному диску. Лопастей чашечки, лепестков и тычинок обычно по 5, изредка 3-4 или 6 -10. Число плодолистиков (и соответственно число гнезд завязи и лопастей рыльца) 3, реже 5, 2 или до 8-10 (переднеазиатский род мишоксия - Michauxia).
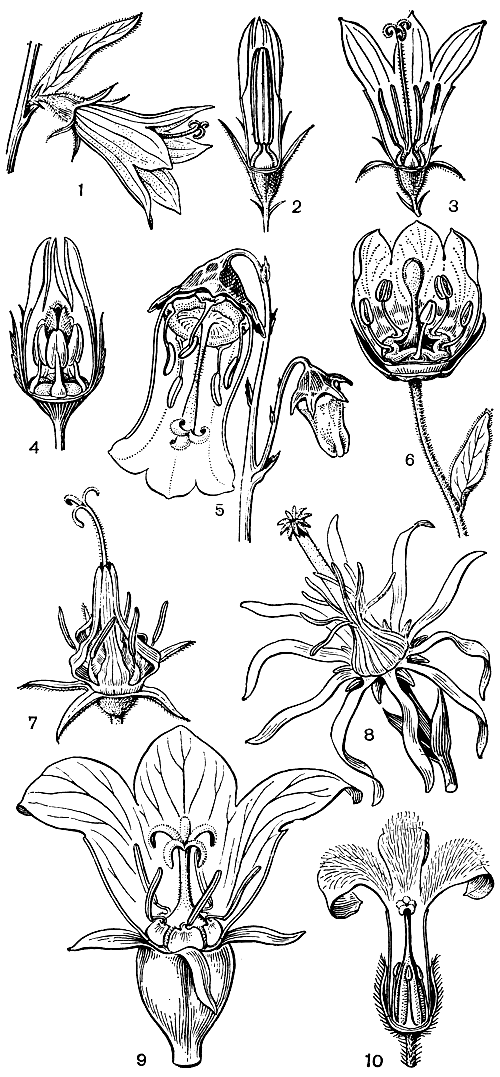
Рис. 237. Различные типы строения цветка у подсемейства колокольчиковых: 1, 2, 3 - колокольчик репчатовидный (Campanula rapunculoides); 4 - гаделлия молочноцветковая (Gadellia lactiflora); 5 - азорина Видаля (Azorina vidalii); 6 - кодонспсис клематисовидный (Godonopsis clematidea); 7 - кольник эдрайантусолистный (Phyteuma hetraianthifolium); 8 - мишоксия гладкая (Michauxia laevigata); 9 - платикодон крупноцветковый (Platycodon grandiflorus) 10 - цианантус мелколистный (Cyananthus microphyllus)
Плод - коробочка, раскрывающаяся створками или порами у основания, у вершины или же одним верхушечным отверстием, изредка раскрывающаяся на створки сверху донизу на неправильные фрагменты, образуя как бы воронку (эдрайантус - Edraianthus), редко почти нераскрывающаяся. Листья обычно цельные, редко перистые, как у эндемичного для острова Крит монотипного рода петромарула (Petromarula).
Биологические особенности многих представителей этого подсемейства изучены довольно детально. В частности, хорошо изучен механизм опыления, представляющий у колокольчиковых особый интерес. Как и у большинства остальных представителей семейства, нектар выделяется диском и, за немногим исключением, как азорина и гаделлия, защищен более или менее расширенными и часто довольно тесно сомкнутыми основаниями тычиночных нитей. Доступ к нектару, как правило, бывает открыт только на верхушке этого купола над нектарным диском, где он несколько отстает от столбика и, как правило, насекомые могут добывать нектар только сверху. Однако, как указывает С. Шетлер (1979), эта защита нектара не всегда достаточно надежна, так как у видов с колесовидным венчиком насекомые, проникая сбоку, могут просто раздвигать основания тычиночных нитей и таким образом извлекать нектар. Цветки характеризуются очень сильно выраженной протандрией, и пыльники вскрываются еще в бутоне. В связи с этим подача пыльцы принимает очень своеобразный характер. Это своеобразие заключается в том, что верхняя часть столбика, включая наружную поверхность плотно сомкнутых лопастей рыльца (а у петромарулы только рыльце), густо усажена правильно расположенными продольными рядами коротких одноклеточных волосков, которые и служат своего рода коллектором пыльцы, или "пыльцеснимателем". В бутоне, где пыльники плотно окружают покрытую волосками часть столбика, пыльца после ее освобождения из гнезд пыльника, прилипает к этим волоскам. После того как гнезда пыльника освободились от пыльцы, венчик раскрывается и все тычинки, кроме оснований тычиночных нитей, засыхают и скручиваются. Что же касается столбика, то он постепенно удлиняется и выносит пыльцу наружу. Некоторое время столбик служит лишь для подачи пыльцы опылителям, которые в поисках нектара неизбежно касаются покрытых пыльцой волосков столбика, находящихся на прямом их пути к нектарному диску.
С окончанием функции пыльцеотдачи волоски "пыльцеснимателя" обычно исчезают. Как показали специальные исследования, восходящие еще к А. Броньяру (1839) и Ф. Мейену (1839), волоски не опадают (как можно было думать), а как бы втягиваются внутрь. Они обладают совершенно уникальной способностью втягиваться и погружаться в наружную ткань столбика. Еще А. Броньяр указывал на то, что это явление совершенно исключительное в растительном мире. В процессе этого погружения (ретракции или инвагинации, как называют это явление в специальной литературе), которое происходит особенно быстро во время наибольшей активности опылителей, волоски втягиваются внутрь большого базального гнезда основания волоска до тех пор, пока лишь кончик волоска останется над поверхностью эпидермы.
А. X. Xасл (1841) считал, что втягивание волосков объясняется высыханием их содержимого. По мнению же С. Шетлера (1979), весь этот процесс представляет собой чисто осмотическое явление. О. Кирхнер (1897) считал, что насекомые не в состоянии собрать пыльцу, пока волоски не втянутся и тем самым не освободят ее. Правда, как показали последующие исследования, пыльцеотдача и "ретрактильный" процесс происходят одновременно и только во время активности опылителей. Но это только несколько видоизменяет и уточняет первоначальную гипотезу Кирхнера.
С исчезновением "пыльцеснимателя" кончается мужская фаза процесса опыления. Далее наступает вторая, женская стадия. Она начинается с того, что плотно сомкнутые до тех пор лопасти рыльца развертываются в виде звезды, тем самым обнажая его воспринимающую поверхность. Теперь уже при посещении цветка опылитель так или иначе заденет сосочки рыльца, и если на нем есть пыльца, то должно произойти опыление.
Во многих случаях лопасти рыльца, загибаясь назад, касаются своими сосочками столбика. Многие исследователи предполагают, что в тех случаях, когда на столбике еще сохранились остатки пыльцы, может произойти самоопыление. Самоопыление принципиально возможно также у видов со свисающими вниз цветками, у которых пыльца, если она еще осталась на столбике, может падать на отогнутые лопасти рыльца. Но сколь часто происходит самоопыление в действительности? Начиная с классической работы Ч. Дарвина (1876), много раз высказывалось мнение, что в отсутствие опылителей лишь относительно немногие колокольчиковые способны к самоопылению.
У некоторых представителей подсемейства колокольчиковых, например у видов родов легузия (Legousia), триоданис (Triodanis), колокольчик и лобелия известна клейстогамия.
Все представители подсемейства специализировались в тех или иных направлениях, и среди них не сохранилось родов, которые мы могли бы поставить сколько-нибудь уверенно у основания родословного древа колокольчиковых. Тем не менее имеется ряд родов, у которых еще сохранились такие признаки, как верхняя завязь, бороздные (не борозднопоровые) пыльцевые зерна и некоторые другие примитивные черты. К числу таких родов относятся цианантус (Cyananthus), лептокодон, кодонопсис, островския.
В роде цианантус около 30 видов, распространенных в Гималаях, Тибете и Юго-Западном Китае. Особенностью этих низких многолетних трав является столь редкая в семействе колокольчиковых верхняя завязь. Другой также примитивной чертой цианантуса является, как показали Г. Эрдтман (1952) и Е. М. Аветисян (1967), строение оболочки его пыльцевых зерен, у которых борозды (в числе от 6 до 9) не только с неровными и неясно выраженными краями, но, что еще важнее, не имеют отверстия (оры). Кроме того, поверхность пленок этих борозд более или менее сходна с общей поверхностью зерна, что также является признаком примитивности. Подобное же строение оболочки пыльцевого зерна установлено для родов кодонопсис, лептокодон и островския. Монотипный род лептокодон встречается в Гималаях и в Юго-Западном Китае, где вообще сосредоточено много древних типов. Он характеризуется нектарным диском с 5 булавовидными желёзками по краю. Относительно большой род кодонопсис, насчитывающий более 30 видов, распространен в Средней Азии, в Гималаях, Ассаме, Восточной Азии и Малезии. У кодонопсисов уже гораздо больше признаков специализации: некоторые из них вьющиеся, корни обычно клубневидно утолщенные, тем не менее пыльцевые зерна у них бороздные, а завязь у некоторых видов верхняя. У очень близкого к кодонопсису и часто объединяемого с ним рода кампанумея (Campanumaea), рас пространенного в Южной, Восточной и Юго-Восточной Азии и отличающегося главным образом плодами-ягодами, пыльцевые зерна уже трехбороздно-поровые. Не менее специализирована островския, характеризующаяся редкой для колокольчиковых особенностью - мутовчатыми листьями. Островския встречается в Средней Азии (Гиссарский и Угамский хребты) и на севере Афганистана. Это типичный геофит с клубневидно утолщенным гипокотилем и главным корнем. Цветки у нее очень крупные, самые крупные в подсемействе. По наблюдениям Т. В. Шулькиной (1978), первое его цветение наблюдается на 10-й, а иногда даже только на 16-й год. Лишенные чешуи почки возобновления островскии ежегодно втягиваются глубоко в почву контрактильным корнем. В подсемействе колокольчиковых этот род стоит довольно изолированно и был с полным основанием выделен Ан. А. Федоровым (1957) в отдельную трибу островскиевых (Ostrowskieae).
Относительно примитивен род канарина, в состав которого входят 3 вида, 2 из которых встречаются в тропической Восточной Африке, а третий - на Канарских островах. Это наземные или эпифитные травы с утолщенными, мясистыми корнями. Листья у. них супротивные или сидят по 3. У африканской канарины Эмина стебли прямостоячие и лазящие или висячие, длиной до нескольких метров. У другого африканского вида - канарины абиссинской (C. abyssinica) - стебли также достигают нескольких метров длины, но они цепляются за опору посредством закрученных черешков и цветоножек. Канарский вид (C. canadensis) также более или менее цепляющийся. Цветки канарины довольно крупные, иногда достигающие в длину 7,5 см, одиночные, висячие, (5)6 (7)-членные, желтоватые, оранжевые или красные. В отличие от упомянутых выше родов пыльцевые зерна канарины трехбороздно-поровые. Плод - мясистая коробочка (иногда неправильно называемая ягодой).
Самый большой род в подсемействе собственно колокольчиковых - это род колокольчик (Campanula, табл. 63), насчитывающий до 350 видов. Это многолетние травы, реже однолетники или полукустарники. Виды колокольчика встречаются почти всюду (причем немного меньше половины видов произрастает на территории СССР) во внетропических областях северного полушария, за исключением лишь части Арктики. В Африке некоторые виды встречаются в тропической зоне, а колокольчик съедобный (C. edulis) достигает Танзании. На Аравийском полуострове этот же вид встречается в Йемене. Род колокольчик обладает очень широкой экологической амплитудой. Его виды встречаются в самых разных климатических зонах и в самых различных сообществах. В тропической Африке они встречаются в горных грасслендах; в умеренных же областях виды колокольчиков можно видеть в лесах, кустарниковых зарослях, на лугах и в степях, на высокогорных лугах и в альпийских коврах, а некоторые из них встречаются даже в полупустынях и пустынях. Многие своеобразные и в систематическом отношении изолированные виды произрастают на известняковых скалах. Одним из наиболее замечательных видов, приуроченных специально к известнякам, является эндемичный для Южных Альп колокольчик каменный (C. petraea), мелкие бледно-желтые цветки которого собраны в плотную головку. Не менее своеобразны произрастающие на известняковых скалах в ущельях Западной Грузии колокольчик удивительный (C. mirabilis) и колокольчик парадоксальный (C. paradoxa) или встречающийся на известняковых скалах Армении колокольчик скребница (C. radula, табл. 63).

Таблица 63. Колокольчиковые: 1 - колокольчик раскидистый(Campanula patula), Московская область

Таблица 63. Колокольчиковые: 2 - колокольчик скребница (C. radula), Армения
Процессу опыления колокольчиков посвящено множество работ, начиная с X. К. Шпренгеля (1793) и кончая С. Фогелем (1975) и С. Шетлером (1979). Колокольчики опыляются преимущественно пчелами и шмелями, хоботки которых легко достигают нектарного диска. Как уже было сказано выше, пыльники вскрываются уже в бутоне, где отдают свою пыльцу волосяному покрову на верхней части столбика, после чего пыльники увядают, тычиночные нити засыхают и более или менее скручиваются и только их защищающие нектар расширенные основания сохраняют тургидность. В только что раскрывшемся цветке тычинки бывают видны лишь в виде засохших остатков на дне трубки венчика. Тем временем столбик удлиняется и пыльца готова к выгрузке. В это время рыльце еще не раскрыто и поэтому самоопыление исключается. Но после того как добывающие нектар насекомые очистили пыльцу с волосяного покрова столбика и волоски втянулись внутрь, лопасти рыльца расходятся и рыльцевая поверхность готова к восприятию пыльцы. Опылители, успевшие побывать на цветках, находящихся на мужской стадии, по пути к нектару касаются рыльцевых лопастей и производят опыление. Очень сходный способ опыления наблюдается у многих близких к колокольчику родов.
Из колокольчиков в качестве отдельного рода обычно выделяется симфиандра (Symphyandra), объединяющая около 10 видов, распространенных от Восточного Средиземноморья до Кавказа и Северного Ирана. В отличие от типичных колокольчиков, пыльники у симфиандры соединены как бы в трубку или даже сросшиеся. Однако существование большого числа промежуточных по этому признаку видов вызывает серьезное сомнение в целесообразности выделения симфиандры в отдельный род. Некоторые систематики, в их числе Ан. А. Федоров (1957) и А. А. Колаковский (1980), поэтому считают, что было бы более правильно включить виды симфиандры в род колокольчик.
К колокольчику очень близок эндемичный для Азорских островов монотипный род азорина (Azorina). Как и у многих других островных колокольчиковых, стебель у азорины деревянистый, высотой до 2 м и увенчан на верхушке розеткой довольно суккулентных и кожистых листьев. Это очень красивое растение с висячими крупными цветками длиной до 3,5 см. Многие ботаники продолжают относить это растение к колокольчикам.
В систематическом отношении значительно дальше от колокольчиков стоит род кольник. В настоящее время в роде кольник насчитывается около 40 видов, распространенных преимущественно в Европе, особенно в Пиренеях, Альпах и Карпатах. Кольники очень интересны механизмом опыления. Цветки у них мелкие, многочисленные, почти сидячие, собранные в короткое колосовидное или почти шаровидное головчатое соцветие. Так же как у колокольчика, пыльца откладывается на волосках столбика в еще закрытом венчике. После того как пыльники высыпали пыльцу и тычинки засохли, венчик начинает раскрываться. Однако у кольников это происходит иначе, чем у колокольчиков. Длинные и тонкие лопасти венчика расходятся у них только в нижней части его, в верхней же половине они остаются сцепленными, образуя тонкую трубку, очень тесно прилегающую к волоскам столбика. Благодаря тому что венчик растет намного быстрее, чем столбик, нижние свободные части его лопастей дуговидно выгибаются наружу (рис. 237, 7). В течение определенного времени венчик продолжает держать столбик, который в результате втягивания волосков лежит в нем свободно. Когда опылители кольника, крупные пчелиные, садятся на нижнюю, вогнутую часть венчика, то тем самым оказывают давление на верхнюю, трубчатую его часть. В результате этого венчик полностью раскрывается и пыльца экспонируется. После этого раскрываются также лопасти рыльца и цветок переходит в женскую фазу. Биологическое преимущество этой модели механизма опыления заключается в том, что пыльца сохраняется внутри трубки венчика до тех пор, пока не приблизится время его подачи.
Определенный прогресс в эволюции опыления мы наблюдаем также у рода букашник. Это относительно небольшой род (около 20 видов), распространенный в Европе, Средиземноморье и Малой Азии. Цветки у букашников мелкие, собранные в густые верхушечные головки, окруженные оберткой из прицветников. Нектар доступен даже короткохоботковым насекомым, так как венчик расщепляется на линейно-ланцетные лопасти по направлению снизу вверх, а тонкие тычиночные нити свободны. Пыльники сросшиеся своими основаниями в трубку. Столбик от середины доверху волосистый, с волосками, расположенными в 10 продольных рядов. Удлиняясь, столбик выносит покрытую пыльцой часть над лопастями венчика, а затем, после отдачи пыльцы и исчезновения волосков, открывает 2-лопастное (очень редко 3-лопастное) рыльце. При каждом посещении насекомые опыляют сразу много цветков и таким образом густое соцветие вполне компенсирует малые размеры цветков, что у сложноцветных становится правилом.
Одним из самых крупных родов подсемейства колокольчиковые по числу видов (около 200), уступающему только колокольчику, является валенбергия (Wahlenbergia). Виды валенбергии распространены главным образом в умеренных областях южного полушария, особенно в Африке, с немногими видами в Европе, континентальной Азии, на острове Тайвань и в Малезии. На территории СССР виды этого рода отсутствуют. От колокольчиковых валенбергия отличается главным образом тем, что плоды раскрываются локулицидыо верхушечными створками, образующими на верху коробки одно отверстие. Виды валенбергии - многолетние или однолетние травы, полукустарники или иногда маленькие кустарники. Строение цветка и механизм опыления очень сходны с колокольчиками, но строение и способ раскрывания коробочки сходны с кодонопсисом, от которого, однако, валенбергия отличается гораздо более специализированными трехпоровыми пыльцевыми зернами.
У следующего подсемейства - цифиевые - цветки зигоморфные, 5-членные, обоеполые, мелкие, собранные в различного рода бокоцветные соцветия (рис. 239). Венчик сростнолепестный, более или менее двугубый. Тычинки обычно прикреплены к трубке венчика; нити свободные или более или менее сросшиеся, иногда у основания расширенные; пыльники свободные. Гинецей из 2 плодолистиков; завязь полунижняя или нижняя, 2-гнездная или реже почти 1-гнездная, с многочисленными семязачатками в каждом гнезде. Плод обычно коробочка, раскрывающаяся 2 створками или реже боковыми щелями (цифокарпус - Gyphocarpus); у рода паришелла (Parishella) плод - криночка. Однолетние или многолетние травы, иногда с клубневидно утолщенным корнем, как некоторые виды цифии (Cyphia).

Рис. 239. Цифия лещинолистная (Cyphia coryiifolia): 1 - ветвь с цветками; 2 - цветок; 3 - тычинка; 4 - плод; 5 - поперечный разрез завязи
В подсемействе цифиевые 5 родов и около 95 видов. Монотипный род цифокарпус эндемичен для Чили, а другой монотипный род - паришелла - для Калифорнии (пустыня Мохаве). Род немакладус (Nemacladus, около 12 видов) распространен на юго-западе США и в Мексике, монотипный род псевдонемакладус (Pseudonemacladus) - в Мексике, а самый большой в подсемействе род цифия (около 80 видов) - в тропической и субтропической Африке (особенно в Южной Африке) и на островах Зеленого Мыса.
В некоторых отношениях, особенно по строению оболочки пыльцевых зерен, цифиевые занимают как бы промежуточное положение между собственно колокольчиковыми и лобелиевыми (А. Дунбар, 1975). В то время как паришелла калифорнийская (P. californica) и немакладус красноватый (N. rubescens) по строению пыльцы сходны с некоторыми собственно колокольчиковыми, род цифокарпус очень похож на некоторые лобелиевые, например на род изотома (Isotoma). Но цифиевые в целом ближе к лобелиевым, чем к колокольчиковым.
Процесс опыления у цифиевых изучен недостаточно. Характерна протандрия. Наряду с перекрестным опылением наблюдается и самоопыление, особенно у рода цифия. У паришеллы, немакладуса и псевдонемакладуса пыльник раскрывается прежде, чем столбик начинает выходить из трубки тычиночных нитей.
Подсемейство сфеноклеевые представлено лишь 1 родом сфеноклея (Sphenoclea, рис. 240), состоящим из 2 видов, один из которых широко распространен в тропиках Старого Света, заходит в Иран и в Среднюю Азию (Южный Узбекистан и Южный Таджикистан) и занесен в Южную Америку, а второй эндемичен для Западной Африки. Виды сфеноклеи - однолетние травы с мочковатой корневой системой, несколько суккулентными стеблями и очередными цельными листьями. Внешне они напоминают лаконос (Phytolacca), хотя в систематическом отношении чрезвычайно далеки от лаконосовых. Цветки мелкие, белые, невзрачные, собраны в очень густые верхушечные колосья, актиноморфные, обоеполые, снабженные 2 прицветничками. Чашечка сростнолистная, с вогнутыми лопастями. Венчик сростнолепестный, опадающий, в отличие от подсемейства колокол ьчиковых черепитчатый. Тычинки прикреплены к трубке венчика, с очень короткими, у основания расширенными нитями; пыльники свободные. Морфологически 3-бороздно-поровые пыльцевые зерна сфеноклеи сходны с таковыми представителей подсемейства цифиевые и лобелиевые, особенно с родом цифия (А. Дунбар и Г. Валлентанус, 1976). Гинецей из 2-3 плодолистиков, с очень коротким и лишенным волосков столбиком и головчатым рыльцем; завязь верхняя или полунижняя, с многочисленными семязачатками, сидячими на больших грибообразно расширенных губчатых плацентах. Плод - 2-гнездная перепончатая криночка с многочисленными семенами. Семена очень мелкие, с очень скудным эндоспермом или даже без эндосперма.
Виды сфеноклеи произрастают во всякого рода влажных местообитаниях, но особенно часто встречаются на рисовых полях и в канавах. К сожалению, механизм опыления этого очень своеобразного и интересного рода не изучен.
С предыдущими подсемействами тесно связано подсемейство лобелиевые, насчитывающее около 30 родов и более 1200 видов. Оно распространено главным образом в тропиках и в умеренной зоне южного полушария, некоторые виды встречаются в северной умеренной зоне. Наибольшая концентрация лобелиевых наблюдается в Америке (около 800 видов), за которой следуют Африка, Австралия, Азия и Новая Зеландия. В Европе 2 вида лобелии и 1 вид лаурентии (Laurentia). В нашей стране встречаются 2 вида лобелии (в европейской части, в Восточной Сибири и на Дальнем Востоке). Дальше всех на север идет водная лобелия Дортманна, достигающая Северной Европы (у нас до Архангельской области) и Канады, и лобелия сидячелистная (L. sessilifolia), доходящая до Восточной Сибири и Камчатки.
Цветки лобелиевых собраны в бокоцветные или редко верхоцветные соцветия, реже одиночные, зигоморфные или очень редко почти актиноморфные, 5-членные, обоеполые или очень редко однополые и тогда двудомные (некоторые виды лобелии с плодами-ягодами). Чашечка 5-лопастная. Венчик сростнолепестный, но часто более или менее расщеплен (редко лепестки почти до основания свободные), большей частью двугубый. Тычинок 5, приросших к трубке венчика или свободных; пыльники довольно плотно спаянные в трубку. Гинецей из 2 плодолистиков; столбик нитевидный, с 2-лопастным или очень, редко цельным рыльцем; под рыльцем имеется воротничок из волосков; завязь обычно нижняя, редко полунижняя или даже полностью верхняя, 2-гнездная или редко 1-гнездная, обычно с многочисленными семязачатками. Плод - коробочка или ягода. Семена мелкие, обычно многочисленные, с маленьким зародышем в обильном мясистом эндосперме. Многолетние или однолетние травы или реже древовидные растения. Листья обычно очередные, редко супротивные или мутовчатые, цельные или различным образом расчлененные.
Разнообразие жизненных форм лобелиевых исключительно большое, значительно большее, чем у собственно колокольчиковых. По размерам они варьируют от крошечных растений длиной всего лишь в несколько сантиметров, как, например, малезийская лобелия Арчболда, новозеландская лобелия наименьшая (L. perpusilla) или родственная ей австралийская лобелия дарлинговская (L. darlingensis), до произрастающих на Гавайских островах, часто похожих на пальмы древовидных форм, достигающих в высоту иногда 15 м, как, например, обитающая в дождевых лесах цианея мелкочешуйчатая (Cyanea leptostegia). Больших размеров достигают не только древовидные формы, но и травянистые виды, как некоторые африканские лобелии (например, лобелия ланурийская - L. lanuriensis), достигающие в высоту 8-10 м. Это подлинные гиганты среди трав, не имеющие аналогов среди других двудольных. Большого разнообразия достигли у лобелиевых также расположение, размеры, форма и строение листьев. Обычно листья распределены по всему стеблю, но нередко они сосредоточены у основания стеблей, как особенно у розеточных африканских лобелий, или же, напротив, сосредоточены на верхушке стебля, как у суккулентной гавайской брайамии замечательной (рис. 238). Очень варьируют размеры листьев, но особенно замечательно разнообразие форм листовой пластинки, наиболее ярко выраженное у гавайского рода цианея. У большинства видов этого рода листья более или менее лопастные. Но наряду с ними встречаются виды с цельными листьями, которые у цианеи линейполистной (C. linearifolia), произрастающей на более солнечных, открытых местообитаниях, очень узкие, линейные. Те же виды, которые произрастают в тенистых лесах, имеют более или менее расчлененные листья. У цианеи асплениелистной (C. aspleniifolia) и цианеи глимеса (C. glimesiana) листья перистые, а у цианеи Шипмана (C. shipmanii) сегменты перистого листа, в свою очередь, перисто лопастные. Но наиболее уклоняются от основного типа листа лобелиевых пальчатораздельные листья колумбийского сифокампилуса удивительного (S. mirabilis) и суданской лобелии рассеченной (L. dissecta).
Интересными жизненными формами лобелиевых являются пальмовидные розеточные древовидные формы эндемичных гавайских родов. Как указывает известный американский ботаник Ш. Карлквист (1965) в своей книге "Островная жизнь", гавайские лобелиевые развивались параллельно и, вероятно, под влиянием эндемичного для Гавайских островов семейства гавайских цветочниц (Drepanididae). Это касается не только древовидных форм, но и трав, как крупноцветные розеточные травянистые виды рода цианея. Обе эти группы, биологически тесно связанные процессом опыления, представляют один из наиболее ярких примеров радиации от исходной предковой формы, некогда заселившей Гавайские острова. На Гавайских островах имеются свои очень интересные виды лобелии, например лобелия "слава горы" (L. gloriamontis). Однако гораздо более замечательны такие эндемичные гавайские роды, как очень близкий к лобелии род трематокарпус (Trematocarpus, или Trematolobelia), клермонтия (Clermontia), цианея, делиссея (Delissea), ролландия (Rollandia) и особенно брайамия. По словам Ш. Карлквиста, брайамия - одно из наиболее курьезных растений в мире, которое эволюционировало в очень своеобразном направлении. Она растет в расщелинах почти недоступных крутых скал, образующих северные побережья островов Кауаи и Молокаи. Сравнительно невысокий (до 1 м) толстый и сочный стебель этого растения у основания более или менее округлый, что дает ему возможность при порывах ветра качаться, не падая, из стороны в сторону, подобно "ваньке-встаньке". Листья растения также толстые и суккулентные, что характерно для многих скальных растений.
Очень интересное биологическое приспособление наблюдается у трематокарпуса крупноколосого (T. macrostachys) - единственного представителя этого рода. Незрелые плоды трематокарпуса представляют собой обычные ягоды. Но в процессе созревания мясистая часть плода превращается в своего рода кашу, которая вымывается дождем. В результате сохраняется только проводящий скелет, внутри которого находится пара бумажистых мешочков, которые при смачивании закрываются, а при сухой погоде вновь открываются. Заключенные внутри этих мешочков семена в сухую погоду высвобождаются, и, когда порывы ветра раскачивают эти небольшие древовидные растения, семена вытряхиваются через отверстия в остове плода. Более того, в отличие от всех остальных лобелиевых, семена этого замечательного растения плоские и крылатые и могут поэтому уноситься ветром на далекие расстояния. По Ш. Карлквисту, этим, вероятно, можно объяснить, что трематокарпус является единственным представителем лобелиевых, который распространился на всех крупных Гавайских островах.
Другим интересным приспособлением, наблюдаемым только у гавайских лобелиевых, является наличие многочисленных шипов у некоторых видов цианей, причем не только на стеблях и листьях, но иногда, как у цианей колючецветковой (C. aculeatiflora), также на цветах и плодах. Так как на Гавайских островах не было млекопитающих, то можно лишь предположить, что эти шипы защищают их от личинок насекомых и наземных моллюсков. Как указывает Ш. Карлквист, особенно густо покрыты шипами молодые и, следовательно, более уязвимые листья колючих цианей. Наконец, у видов клермонтий наблюдается эпифитизм, а клермонтия Пеле (C. peleana) ведет исключительно эпифитный образ жизни.
Опыление у лобелиевых имеет некоторые особенности, указывающие на большую их эволюционную подвинутость. Так же как у растений подсемейства колокольчиковых, пыльники вскрываются еще в бутоне. Но, в отличие от колокольчиковых, пыльники не засыхают после этого, а остаются в спаянном состоянии, образуя чехол вокруг пыльцы. Иначе ведет себя также столбик. Так как верхушка столбика с воротничком волосков расположена у них у основания или в самой нижней части пыльнкковой трубки, то волоски уже не играют роли "пыльцеснимателя", как это мы видели у колокольчиковых. В процессе эволюции произошла смена функции и роль волосков у лобелиевых стала другой. Сначала верхушка столбика и волоски лишь задерживают выпадение пыльцы из пыльниковой трубки. Но затем, по мере удлинения столбика, она принимает на себя более активную роль своеобразного поршня, выталкивающего пыльцу из пыльниковой трубки наружу, где она удерживается пучком крепких волосков на верхушке пыльников. На этой стадии и вплоть до окончания выгрузки пыльцы рыльце остается закрытым и самоопыление тем самым исключается. С раскрытием венчика начинается выгрузка пыльцы опылителями. Вскоре после этого рыльце раскрывается и начинается женская стадия. Но так как собственная пыльца уже высыпалась или удалена опылителями, то, как правило, опыление возможно только посредством чужой пыльцы. Однако самоопыление все же не полностью исключено. Оно может произойти вследствие того, что оставшаяся в волосках пыльников пыльца может упасть на лопасти слегка наклоненного вниз рыльца. Более того, некоторые виды лобелии и других родов в той или иной степени перешли к самоопылению. Наиболее ярким примером самоопыления у лобелиевых является лобелия Дортманна. Это растение настолько приспособилось к самоопылению, что рыльце уже никогда не выходит из трубки пыльника. Опыление происходит вследствие того, что пыльца проникает к лопастям рыльца через внутреннюю часть трубки пыльника. Иногда же дело доходит до клейстогамии, т. е. опыление происходит внутри еще не раскрывшегося цветка, особенно у подводных цветков. Но чаще всего самоопыление у лобелиевых не является обязательным и представляет собой запасной способ опыления, к которому растение прибегает при недостатке опылителей.
Опылителями лобелиевых чаще всего являются различные насекомые, прежде всего пчелиные (Apidae) и чешуекрылые, а также жуки и др. Очень большую роль в опылении многих лобелиевых играют птицы. Почти все крупноцветковые лобелиевые опыляются птицами. К числу орнитофильных лобелиевых принадлежат прежде всего американские роды сифокампилус (Siphocampylus, рис. 241), центропогон (Centropogon), бурмайстера (Burmeistera), все гавайские роды, а также отдельные виды большого рода лобелия. Цветки этих орнитофильных лобелиевых крупные (длиной от 2 до 8 см), массивные, часто узкотрубчатые, по крайней мере в нижней части венчика, где трубка имеет в поперечнике едва 2 мм, обычно более или менее сильно изогнуты, а у некоторых видов сифокампилуса изогнуты под прямым углом или даже серповидно, редко прямые и широкотрубчатые, с обильным нектаром, большей частью без запаха, с торчащими наружу жесткими столбиками и тычинками, без посадочной площадки. Окраскадгветков очень разная, особенно у видов рода цианея, но чаще всего красная, или оранжевая, или белая с пурпуровыми или красными прожилками.
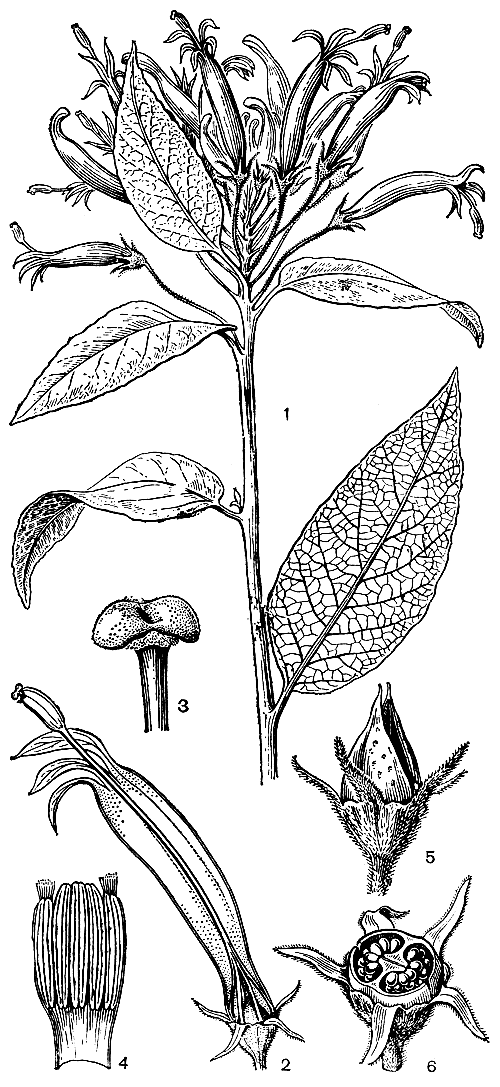
Рис. 241. Сифокампилус сетчатый (Siphocampylus reticulatus): 1 - ветвь с цветками; 2 - цветок; 3 - верхушка столбика с раскрывшимся рыльцем; 4 - развернутая верхняя часть трубки пыльников; 5 - плод; 6 - поперечный разрез плода
Орнитофильные цветки настолько приспособлены к опылению птицами, что уже не могут опыляться насекомыми. Еще Ч. Дарвин (1859) отметил, что произраставшую в его саду орнитофильную мексиканскую лобелию блестящую (L. splendens) не посещали насекомые. Там, где рядом растут орнитофильная лобелия пурпуровая (L. cardinalis, табл. 63, рис. 242) с красными цветками и энтомофильная лобелия сифилитическая (L. syphilitica) с синими цветками, колибри опыляли только первый вид, в то время как второй опыляли исключительно насекомые. В связи с этим отметим описанный Б. М. Козо-Полянским (1946) интересный случай, когда в ботаническом саду Воронежского университета лобелия пурпуровая опылялась крупным дневным бражником из рода макроглосса (Macroglossa), который по размерам, форме тела и манере сосать нектар близок к колибри. Однако это замещение птиц насекомыми - случай, вероятно, очень редкий. Что переход от орнитофилии к энтомофилии очень труден, доказывается тем, что произошло на Гавайских островах, где лобелиевые опыляют цветочницы. В тех местах, где цветочницы вымерли, эти растения, например цианея коротковолосистая (C. hirtella), перешли по необходимости к самоопылению.
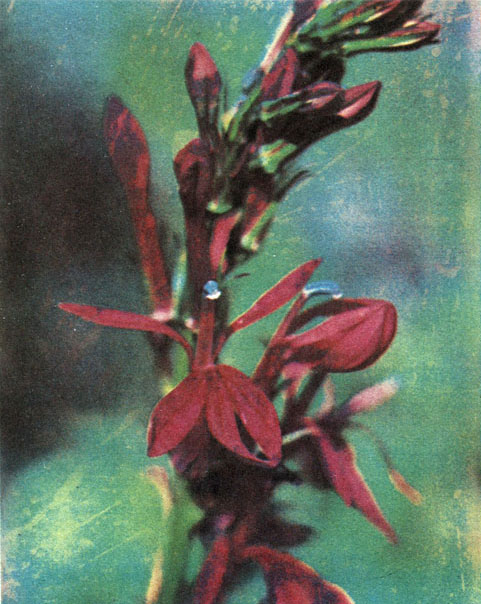
Таблица 63. Колокольчиковые: 3 - лобелия пурпуровая (Lobelia cardinalis), парк Ботанического института АН СССР в Ленинграде

Таблица 63. Колокольчиковые: 4 - лобелия Лешенольта (L. Leschenaultia), Индия
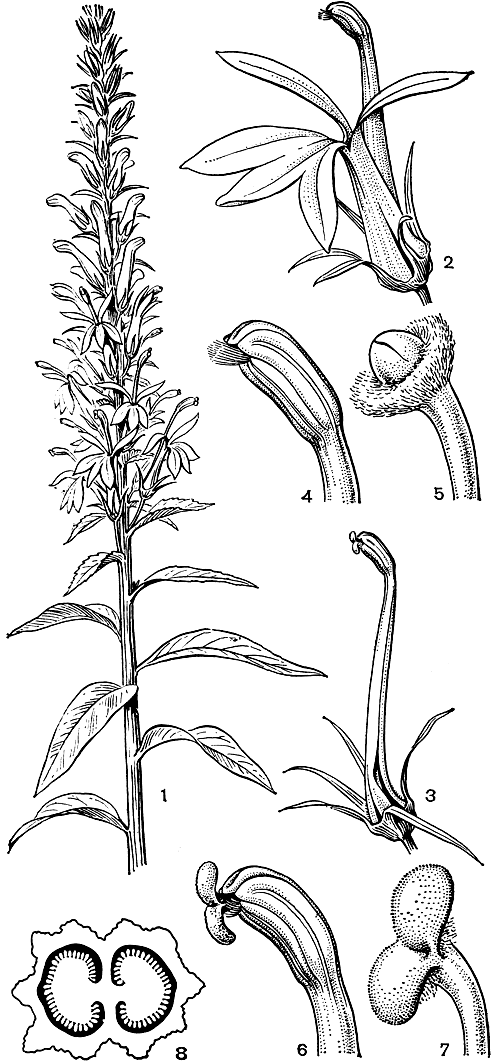
Рис. 242. Лобелия пурпуровая (Lobelia cardinalis): 1 - ветвь с цветками; 2 - цветок (лепестки у основания свободные); 3 - цветок с удаленным венчиком, рыльце начинает раскрываться; 4 - андроцей (видны два нижних пыльника с пучком волосков); 5 - верхушка столбика с нераскрывшимся рыльцем; 6 - пыльники с выступающим раскрывшимся рыльцем; 7 - рыльце; 8 - поперечный разрез завязи
В подсемействе лобелиевые, как и в семействе колокольчиковые в целом, самым большим родом является лобелия. По Ф. Вильмеру (1968), в него входит 388 видов, но если согласиться с теми ботаниками, которые род пратия (Pratia), насчитывающий 36 видов, объединяют с лобелией, то тогда число видов последнего превысит 430. Это почти космополитный род, очень широко распространенный на всех континентах, но главным образом в тропических и субтропических странах, особенно в Америке. В пределах этого рода наблюдается почти весь спектр жизненных форм подсемейства.
Второй по величине род - центропогон - насчитывает около 220 видов. Это род чисто неотропический, с наибольшим числом видов в Колумбии. Почти равен ему неотропический род сифокампилус (около 215 видов). На третьем же месте - гавайский род цианея (около 60 видов).
Совершенно особняком стоит в семействе колокольчиковых очень своеобразный род пентафрагма (Pentaphragma), в котором около 30 видов, распространенных от Нижней Бирмы, Южного Китая и Индокитая через Малезию до Новой Гвинеи. В Малезии виды пентафрагмы отсутствуют только на Яве и на Малых Зондских островах. Это более или менее суккулентные многолетние травы высотой от 5- 10 см до 2,5 м, часто у основания несколько деревянистые, большей частью покрытые ветвистыми многоклеточными волосками. Листья очередные, большей частью более или менее асимметричные. Цветки небольшие, собраны в пазушные, верхоцветные, часто скорпиоидные (завиткообразные), напоминающие соцветия некоторых бурачниковых, актиноморфные, обычно обоеполые. Чашелистиков 5, неравных, остающихся. Лепестков 5 или редко 4, более или менее сросшихся или реже свободных дочти до основания, остающихся. Тычинок 5, прикрепленных к венчику, свободных; нити остающиеся; пыльники свободные. Гинецей из 2 плодолистиков с коротким толстым столбиком, заканчивающимся массивным, продолговато-цилиндрическим рыльцем; завязь нижняя, приросшая к цветочной трубке посредством продольных перегородок, образованных продолжением тычиночных нитей; семязачатки очень мелкие и очень многочисленные, висячие на плацентах. Плод - ягода. Семена с очень маленьким зародышем и обильным эндоспермом. Виды пентафрагмы обитают обычно в очень влажных первичных дождевых лесах, часто на скалах вдоль ручьев, поднимаясь иногда до высоты 4000 м над уровнем моря.
По некоторым своим особенностям пентафрагма настолько своеобразна, что некоторыми ботаниками выделяется в отдельное семейство пентафрагмовые (Pentaphragmataceae), которое даже предлагают исключить из порядка колокол ьчиковые. Но далеко не все ботаники с этим согласны, и Дж. Хатчинсон, например, который вообще был склонен скорее к дроблению, чем к объединению семейств, в своей книге "Эволюция и филогения цветковых растений" (1969) решительно возражал против выделения этого рода в самостоятельное семейство. Родственная связь пентафрагмы с колокольчиковыми хорошо подтверждается данными эмбриологических исследований индийских ботаников Р. Н. Капила и М. Р. Виджаярагавана (1965). Об этом же говорят данные палинологии. Е. М. Аветисян (1967) приходит к выводу, что по морфологии пыльцевых зерен пентафрагма не отличается сколько-нибудь существенно от остальных колокольчиковых с бороздно-поровыми зернами, таких, как кодонопсис, платикодон и канарина.
В семействе колокольчиковых много полезных растений. Для человека колокольчиковые представляют прежде всего эстетическую ценность. Среди них очень много красивых растений, и часть из них уже давно вошла в золотой фонд декоративного садоводства. Особенно много декоративных растений среди видов рода колокольчик, некоторые из них широко разводят (например, колокольчик средний - С. medium или колокольчик карпатский - С. саграtica, представленные, как и некоторые другие виды, многими сортами). Кроме колокольчика, культивируют представителей родов канарина, конодопсис, цианантус, островския, платикодон, лобелия, центрапогон, Лаврентия и др.
Некоторые колокольчиковые, например платикодон, виды кодонопсиса, некоторые виды лобелии (особенно лобелия эринус - L. erinus) используют как лекарственные растения. Особенно широкое применение в медицине (в том числе народной) находят виды лобелии. В СССР как лекарственные растения используют североамериканскую лобелию вздутую (L. inflata) и реже западноевропейскую лобелию жгучую (L. urens). Надземные части этих растений содержат алкалоид лобелии, используемый главным образом как стимулятор дыхания. Среди колокольчиковых есть и некоторые съедобные растения. Так, колокольчик рапунцель (C. rapunculus) используют как овощное растение - корни первого года и листья едят как салат. Съедобны также ягоды некоторых видов кодонопсиса. На острове Ява молодые растения сфеноклеи, а также верхушки старых тушат и едят с рисом. Едят и молодые листья лобелий, например лобелии цейлонской (L. zeylanica), которые также тушат и едят с рисом. Съедобны даже виды гавайского рода цианея. Так, листья цианеи тритоманта (C. tritomantha) употреблялись в пищу аборигенами Гавайских островов как овощ.
Семейство стилидиевые (Stylidiaceae)
В семействе стилидиевые 5 родов и около 160 видов, распространенных главным образом во внетропических областях Австралии, а также в Тасмании, Новой Зеландии, Новой Гвинее, Южной и Юго-Восточной Азии и Голантарктической Южной Америке (Огненная Земля). Из пяти родов в Австралии (вместе с Тасманией) отсутствует только монотипный род ореостилидиум (Oreostylidium), эндемичный для Новой Зеландии. Шире всех распространен самый большой в семействе род стилидиум (Stylidium), заключающий около 140 видов, из которых около 8 видов встречается вне Австралии (Шри-Ланка, Сикким, горы Кхаси. Бенгал, Бирма, Южный Китай, Индокитай, Малезия). В роде левенгукия (Levenhookia) 10 видов, эндемичных для Южной Австралии. Но особенно интересно географическое распространение рода филлахна (Phyllachne, 4 вида), имеющего разорванный ареал. По одному виду филлахны встречаются в Тасмании и на Огненной Земле, а остальные виды приурочены к Новой Зеландии. Разорванный ареал имеет также род форстера (Forstera, 5 видов).
Стилидиевые - многолетние или однолетние (часто эфемеровые) травы, иногда похожие на мхи, а некоторые - на толстянковые и камнеломковые или реже небольшие кустарнички, обычно более или менее ксерофильные. Виды рода филлахна образуют низкие плотные подушки, приспособленные к высокогорному или субантарктическому климату. Новозеландский ореостилидиум выпускает тонкие столоны или корневища. По данным Ш. Карлквиста (1963), у стилидиума лиственницелистпого (S. laricii'olium) наблюдается слабо выраженный аномальный вторичный рост. Листья очередные, маленькие, цельные, большей частью узкие, похожие на листья злаков, стеблевые или чаще в прикорневых розетках. Растения часто с железистыми волосками. У некоторых видов стилидиума содержится инулин. Млечники отсутствуют. Цветки в верхушечных верхоцветных или бокоцветных соцветиях, редко одиночные (филлахна), зигоморфные или иногда почти актиноморфные (форстера), обоеполые или однополые (филлахна). Чашечка 5-7(9) - лопастная, часто покрыта клейкими железистыми волосками; лопасти свободные или чашечка сростнолистная и более или менее двугубая. Венчик 5-лопастный, лопасти большей частью неравные, и самая нижняя из них образует губу; у рода левенгукия губа имеет форму домашней туфли, которая вначале охватывает колонку, но при прикосновении отскакивает вниз. Тычинок 2, редко 3, полностью сросшихся со столбиком, образуя колонку или гиностемий (как у ластовневых). У видов стилидиума колонка в нижней своей части отличается раздражимостью и подвижностью; в нормальном положении колонка наклонена вперед, при соприкосновении отскакивает назад, после чего возвращается в исходное положение. Пыльники, в отличие от колокольчиковых, экстрорзные. Гинецей из 2 плодолистиков, иногда псевдомономерный; рыльце 2-лопастное, расположенное выше пыльников; завязь нижняя 2-гнездная или частично или полностью 1-гнездная, обычно с многочисленными семязачатками. Плод обычно септицпдная коробочка; у форстеры 1-гнездная коробочка открывается на верхушке, а у филлахны - нераскрывающаяся или неправильно раскрывающаяся. Семена обычно очень мелкие, с тонкой кожурой, очень маленьким зародышем и обильным мясистым эндоспермом.
Стилидиевые произрастают в самых различных местообитаниях - от морских побережий до высокогорий. Некоторые из них предпочитают влажные места, включая болота и рисовые поля; другие, напротив, произрастают в относительно сухих местообитаниях, включая прибрежные песчаные пространства с растительностью типа саванны.
Вероятно, все стилидиевые - насекомоопыляемые растения. Для них, как и для всех остальных семейств порядка колокольчиковые, характерна протандрия. Но подача пыльцы происходит у стилидиевых иначе, чем у всех остальных представителей порядка. Пыльца подается насекомым прямо из пыльников. В этом отношении стилидиевые, несмотря на множество признаков их специализации, более примитивны, чем все остальные члены порядка, за исключением близкого к стилидиевым семейства донатиевые (Donatiaceae), у которого, однако, тычинки свободные. Вполне понятно поэтому, что на столбике нет здесь волосков, снимающих пыльцу из раскрывшихся пыльников. Важнейшими опылителями стилидиевых являются пчелы. Возбудимость колонки, ее триггерная способность содействуют перекрестному опылению. Но если насекомые не приводят в действие этот триггерный механизм, то наступает самоопыление. У некоторых видов стилидиума наличие коленчато изогнутой колонки является специальным приспособлением, обеспечивающим самоопыление.
Очень мелкие семена большинства стилидиевых распространяются ветром, а у болотных видов также птицами. Благодаря этому некоторые стилидиумы могли распространиться очень далеко и достичь даже Шри-Ланки и Сиккима.
К стилидиевым стоит очень близко и иногда объединяется с ними маленькое семейство донатиевые, состоящее из 1 рода донатия (Donatia) и 2 видов, один из которых обитает в Тасмании и Новой Зеландии, а другой - в Голантарктической Южной Америке (Огненная Земля). От стилидиевых они отличаются главным образом тем, что свободные между собой тычинки в нижней части коротко приросли к основанию столбика и не образуют таким образом колонки. От стилидиевых они отличаются также морфологией пыльцевых зерен. Виды донатии -низкорослые подушкообразные травы с густо расположенными линейными листьями и одиночными и верхушечными, актиноморфными, обоеполыми цветами. Гинецей из 2-3 плодолистиков со свободными столбиками с головчатыми рыльцами. Некоторые ботаники переносят донатиевые в порядок камнеломковые, но по эмбриологическим и другим признакам они стоят гораздо ближе к стилидиевым.
Семейство гудениевые (Goodeniaceae)
Семейство гудениевые включает 15 родов и около 350 видов, распространенных главным образом в Австралии (особенно на юго-западе) и Тасмании, но также в Новой Зеландии, Японии, тропической Азии, в Африке и на Мадагаскаре, в тропической Америке и на островах Тихого океана.
Гудениевые - многолетние травы, полукустарники, кустарнички или кустарники с очередными или редко супротивными цельными листьями. Инулин имеется, но млечники отсутствуют. Цветки одиночные и пазушные в верхоцветных или бокоцветных, иногда головчатых соцветиях, обычно зигоморфные, редко актиноморфные или почти актиноморфные, обоеполые. Чашечка трубчатая, большей частью 5-лопастная. Венчик сростнолепестный, обычно двугубый или редко одногубый, у брунонии (Brunonia) 5-лопастный. Трубка венчика обычно открыта на одной стороне и лепестки часто с пленчатыми крыльями. Тычинок 5, обычно свободных от венчика, реже коротко приросших к его основанию; пыльники интрорзные, свободные или иногда соединены в трубку вокруг столбика. Диск отсутствует. Гинецей из 2 плодолистиков, с нитевидным столбиком с простым или редко 2-3-лопастным рыльцем (азиатско-австралийский род калогина - Calogyne); завязь нижняя, или иногда полунижняя, или даже вполне верхняя, как у австралийского рода веллейя (Velleia) и монотипного австралийско-тасманийского рода брунония, 1- 2-гнездная, редко 4-гнездная; в каждом гнезде 1, 2 или много семязачатков. Рыльце окружено расположенной непосредственно под ним пыльцевой чашей, которая иногда (род лешенолтия - Leschenaultia) бывает 2-3-лопастной; края чаши часто бывают усажены крепкими волосками; пыльцевая чаша представляет собой вырост наружных тканей столбика и поэтому не гомологична волоскам на столбиках колокольчиков. Плод большинства гудениевых - коробочка, раскрывающаяся створками или неправильно, реже плод костянковидный (сцевола - Scaevola) или орех. Семена с прямым зародышем и мясистым эндоспермом или редко без эндосперма.
Почти все гудениевые, за немногими исключениями, произрастают в относительно сухих или периодически сухих местообитаниях, особенно на песчаных побережьях, открытых склонах, в саваннах, вдоль лесных опушек, а также как сорняки на сухих рисовых полях. В сложении растительного покрова сколько-нибудь существенную роль играют лишь некоторые виды сцеволы, особенно сцевола таккада (S. taccada, рис. 243) - кустарник с крупными обратнояйцевидными несколько мясистыми листьями, бледно-голубыми цветками и мелкими белыми ягодообразными плодами. Она растет по песчаным и скалистым морским побережьям, образуя в некоторых местах чистые насаждения. Наиболее характерна сцевола для маленьких островов, особенно для тихоокеанских атоллов.
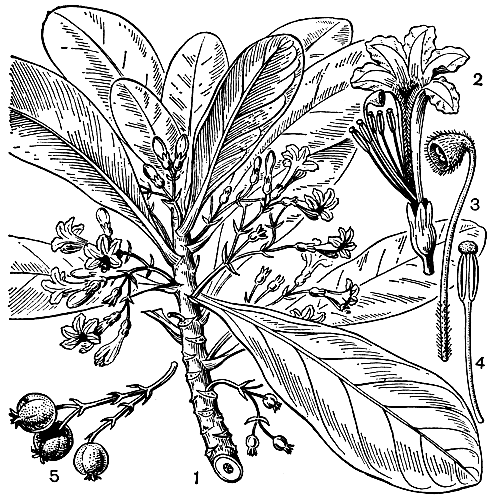
Рис. 243. Сцевола таккада (Scaevola taccada): 1 - ветвь с цветками; 2 - цветок; 3 - столбик и рыльце; 4 - тычинка; 5 - плоды
Цветки гудениевых протандричны и опыляются насекомыми, главным образом пчелами и бабочками. Как и у колокольчиков, тесно сомкнутые или спаянные в трубку пыльники раскрываются еще в бутоне. Но, в отличие от колокольчиковых, пыльца собирается не на волосках столбика, а откладывается в пыльцевой чаше. По мере удлинения столбика пыльцевая чаша продвигается вверх между пыльниками, постепенно собирая высыпающуюся пыльцу, которая в результате буквально переполняет ее. Мужская фаза заканчивается тем, что столбик наклоняется вниз, располагаясь над входом в горизонтальный цветок. Рыльце постепенно увеличивается в размерах и заполняет всю пыльцевую камеру, вытесняя оттуда всю пыльцу, которая остается между волосками чаши в ожидании опылителей. Насекомые, посещающие цветки гудениевых в поисках глубоко запрятанного нектара, неизбежно касаются своим телом пыльцы. Таким образом, как и у колокольчиковых, функция подачи пыльцы перешла у гудениевых от пыльников к столбику (у колокольчиковых - к волоскам, а у гудениевых - к чаше). После этого наступает женская фаза - тычинки засыхают, а рыльце готово к принятию пыльцы.
Обычно мелкие, легкие и плоские семена гудениевых с плодами-коробочками распространяются ветром. Но наряду с анемохорией имеются и другие способы распространения диаспор. Особенно интересно распространение костянок сцеволы. Косточка этих костянок очень твердая, деревянистая и иногда пробковидная. Распространяются они обычно птицами. Но у двух литоральных видов - сцеволы таккада и очень близкой к ней сцеволы Плюмъера (S. plumierii) косточки костянок благодаря их внешнему пробковому слою очень плавучие и в морской воде сохраняют свою всхожесть очень длительное время. Благодаря этому эти два вида получили очень широкое географическое распространение, особенно сцевола таккада (от Мадагаскара до островов Тихого океана).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'