Порядок санталовые (Santalales)
Семейство олаксовые (Olacaceae) (Э. С. Терехин)
В семейство входит около 27 родов и 260 видов. Оно распространено в тропических и отчасти субтропических областях Африки, муссонной Азии, Австралии и Южной Америки. Род неимения (Ximenia) пантропический.
Семейство представлено обычно вечнозелеными деревьями и кустарниками, иногда лиановыми. Многие, если не большинство, представителей семейства являются зелеными корневыми паразитами (полупаразитами). Род олакс (Olax) сочетает в себе черты и свойства разных жизненных форм. У палеотропического рода эритропалум (Erythropalum), иногда выделяемого в отдельное семейство, можно наблюдать побеги, преобразованные в настоящие усики. Листья цельные, очередные, с перистым жилкованием. Цветки в пазушных соцветиях или иногда одиночные, обычно мелкие, как правило, обоеполые, очень редко однополые и двудомные. Чашечка очень маленькая, стростнолистная. Венчик из 3-6 свободных или более пли менее сросшихся лепестков, обычно довольно невзрачный, но у американского рода хаунохитон (Chaunochiton) желто-белые лепестки достигают в длину 8 см. Внутренняя поверхность венчика у представителей некоторых родов покрыта густой массой желто-коричневых волосков. Тычинки в равном числе с лепестками или чаще в несколько раз превышают их число. Их расположение довольно любопытно. У западноафриканской кулы съедобной (Coula edulis) по 3-4 тычинки располагаются напротив каждого лепестка и еще по одной тычинке расположено между каждыми смежными лепестками. У олакса зеленого (О. viridis) по одной тычинке расположено между каждой парой лепестков и по одному стаминодию напротив каждого лепестка. Пыльники вскрываются общей продольной щелью.
Опыление изучено очень слабо, но, судя по строению и запаху цветков, можно предполагать энтомофилию. Цветки многих представителей семейства имеют хорошо развитый нектарный диск и развитую систему стаминодиев, а в цветках эритропалума (Erythropalum) присутствует особый стаминодийный круг. Интересно, что О. Поршом (1929) было высказано предположение о возможности орнитофилии у хаунохитона лорантовидного (Chaunochiton loranthoides), строение цветков которого очень напоминает соцветие орнитофильного рода пситтакантус (Psittacanthus) из семейства ремнецветниковых.
Гинецей из (2)3(4-5) плодолистиков с коротким или длинным простым столбиком и 2 -5-лопастным рыльцем; завязь обычно верхняя, редко полунижняя или нижняя. Разные варианты ее положения встречаются иногда в пределах одного рода, например у рода олакс. Завязь полностью или только в базальной части 2-5-гнездная. В завязи, как правило, формируется центральная колонка, срастающаяся с верхушкой полости завязи или не доходящая до нее. Семязачатки висячие, большей частью анатропные, с двойным или простым интегументом; иногда интегумент отсутствует (олакс черепитчатый - O. imbricata, рис. 167). Хотя в завязи образуется обычно несколько семязачатков, ко времени созревания плодов в каждом из них находится лишь одно семя. Плоды костянковидные или ореховидные, иногда заключены в сильно разросшуюся чашечку. Плоды некоторых видов съедобны.
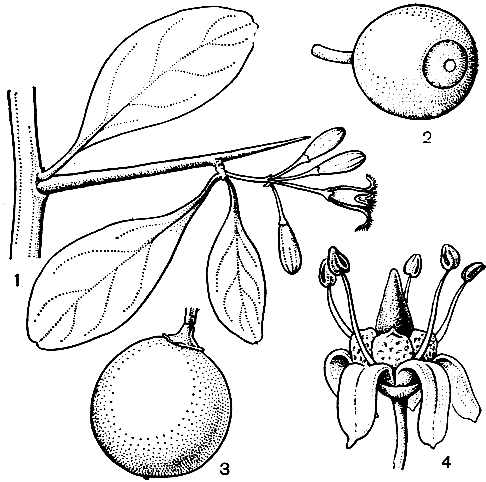
Рис. 167. Олаксовые и опилиевые. Неимения американская (Ximenia americana): 1 - часть ветви с цветком и бутонами. Олакс черепитчатый (Olax imbricata): 2 - плод. Агоиандра высокая (Agonandra excelsa): 3 - плод. Опилия каркасолистная (Opilia celtidifolia): 4 - цветок
Древесину ксимении, как и некоторых других представителей семейства, широко используют в качестве заменителя сандаловой древесины.
К олаксовым очень близко небольшое (9 родов и около 60 видов), но широко распространенное семейство опилиевые (Opiliaceae). Его представители встречаются в Юго-Восточной Азии и Австралии, на многих островах в зоне тропиков и субтропиков (от Тайваня до Новой Гвинеи и Новой Зеландии), в тропической Африке и тропической Америке (от Мексики до Аргентины). Обычно это небольшие деревья, иногда лазящие кустарники с очередными, цельными, перистонервными листьями. Виды отдельных родов - корневые зеленые паразиты иногда с очень сложно устроенными гаусториями, как у рода кансъера (Cansjera).
Семейство санталовые (Santalaceae) (А. Л. Тахтаджян)
В семействе около 35 родов и примерно 425 видов. Санталовые широко распространены в тропических, субтропических и умеренных областях обоих полушарий. Лишь немногие виды встречаются в странах с холодным климатом. Дальше всех на север продвинулись виды рода ленец (Thesium), дошедшие до Швеции и Эстонии, и монотипный североамериканский род геокаулон (Geocaulon), который достиг даже Аляски. В южном полушарии один из видов экзокарпоса - экзокарпос Бидвилла (Exocarpos bidwillii, рис. 168) - встречается на острове Южный в Новой Зеландии, а роды нанодея (Nanodea) и архона (Arjona) достигают Огненной Земли. В нашей стране встречаются только представители самого большого в семействе рода - ленец (Thesium), к которому относится более половины видов санталовых.
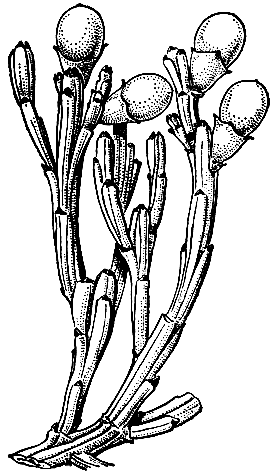
Рис. 168. Плодущие побеги экзокарпоса Бидвилла (Exocarpos bidwillii)
Санталовые большей частью кустарники или травы, многолетние или очень редко однолетние, как средиземноморский ленец низкий (T. humile), реже небольшие деревья. Большинство санталовых - зеленые корневые паразиты (полупаразиты), редко эндопаразиты. Паразиты имеются как среди трав, так и среди кустарников и деревьев. Самые крупные из них относятся к роду санталум (Santalum), отдельные экземпляры которого достигают в высоту 15 м и более и в диаметре 1-1,2 м, а самые маленькие принадлежат южноамериканским родам архона, квинчамалиум (Quinchamalium, около 25 видов в Андах) и монотипному роду нанодея (умеренные области Южной Америки).
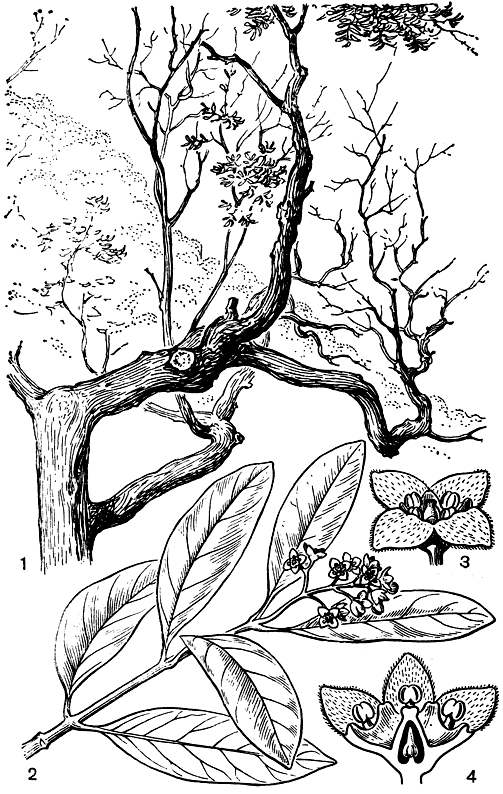
Рис. 169. Санталум фернандесский (Santalum fernandezianum): 1 - общий вид растения; 2 - ветвь с соцветием; 3 - цветок; 4 - продольный разрез цветка
Санталовые паразитируют главным образом на корнях, но среди них известен и целый ряд стеблевых паразитов, например виды родов дендротрофа (Dendrotrophe) и фацеллария (Phacellaria), паразитирующие на ветвях тропических деревьев. Дендротрофа, как и некоторые виды экзокарпоса, могут паразитировать как на ветвях, так и на корнях. Но дальше всех продвинулось приспособление к паразитическому образу жизни у тропического азиатского рода фацеллария, виды которого стали стеблевыми эндопаразитами. Все вегетативное развитие этих растений протекает в тканях стеблей растения-хозяина. Но самое интересное заключается в том, что фацеллария может паразитировать и на паразитах, а именно: на других представителях семейства санталовых, а также на ремнецветниковых (см. ниже).
Листья санталовых очередные или супротивные, цельные, от нормально развитых до чешуевидных, иногда, как у фацелларии, практически отсутствуют. Иногда листья бывают редуцированы только на побегах, образующих гаустории. Листья южноамериканской йодины ромболистной (Jodina rhombifolia, рис. 170), снабженные по краям тремя колючими заострениями, несколько напоминают некоторые типы листьев падуба остролистного.
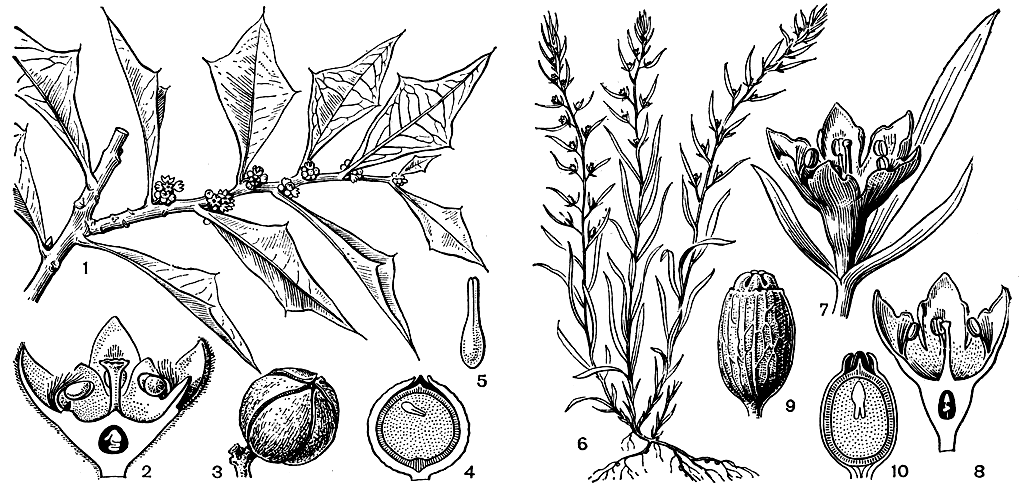
Рис. 170. Санталовые. Йодина ромболистная (Jodina rhombifolia): 1 - ветвь с цветками; 2 - продольный разрез цветка; 3 - плод; 4 - продольный разрез плода; 5 - зародыш. Ленец распростертый (Thesium divaricatum): 6 - общий вид растения; 7 - цветок; 8 - продольный разрез цветка; 9 - плод; 10 - продольный разрез плода
Цветки санталовых мелкие, невзрачные, обычно более или менее зеленоватые, большей частью собраны в разного рода соцветия, реже одиночные, как, например, у рода ленец. Они обычно обоеполые, реже однодомные, двудомные или полигамные. Интересно, что в пределах рода фацеллария имеются как однодомные, двудомные и полигамные виды, так и виды, у которых развиваются только обоеполые цветки (например, у китайской фацелларии Фарже - Phacellaria fargesii). Околоцветник у санталовых простой, представлен только чашечкой, которая состоит из 3-6, чаще всего 4-5 зеленых, более или менее лепестковидных, свободных или сросшихся, часто мясистых чашелистиков (некоторые ботаники считают их не чашелистиками, а лепестками, чашечку же считают редуцированной). За чашечкой следует обычно хорошо развитый лопастный нектарный диск. Как указывает канадский ботаник Дж. Кайт (1969), у санталума и других родов этот диск производит впечатление образования, которое могло возникнуть в результате редукции венчика. Когда лопасти этого диска бывают хорошо развиты, как, например, у рода баклия (Buckleya), они чередуются с чашелистиками, что подтверждает их происхождение из редуцированного венчика. Тычинки в одинаковом числе с чашелистиками и с лопастями диска, обычно приросшие к диску и к чашечке; нити короткие, а иногда (австралийский род хоретрум - Choretrum) пыльники почти сидячие. Пыльники вскрываются продольной щелью. Гинецей из (2) 3-5 плодолистиков, с простым столбиком с головчатым или лопастным рыльцем, завязь верхняя (австралийский род антоболус - Anthobolus - и часть видов рода экзокарпос), полунижняя или нижняя, сросшаяся с основанием нектарного диска. Как показали исследования Ф. и Е. Смитов (1943), нижняя завязь у санталовых и близких семейств возникла в результате образования чашеобразного углубления на верхушке цветоложа, обрастающего завязь (другими словами, она погружена в цветоложе). Завязь санталовых вполне 1-гнездная или с перегородками у основания, со свободной центральной плацентой, несущей 1-3, реже 4- 5 висячих семязачатков. Семязачатки иногда унитегмальные, как у ленца, командры (Comandra) или йодины, но у большинства санталовых они не дифференцированы на интегумент и нуцеллус, т. е. фактически беспокровные. Как показали исследования Ф. Фагерлинда (1939) и С. П. Бхатнагара и Р. Джоши (1965), наибольшая редукция семязачатка наблюдается у рода экзокарпос. У этого замечательного растения семязачаток не только беспокровный, но у него даже нет ясной дифференциации между семязачатком и плацентой. Другой интересной особенностью санталовых является наблюдаемое у некоторых родов, в частности у санталума, удлинение микропилярного конца женского гаметофита, который на 4-ядерной стадии или позднее выходит за пределы семязачатка и может даже достичь канала столбика. Плоды санталовых нераскрывающиеся, орехи или костянки. Интегумент, когда он имеется, потребляется растущим эндоспермом, и поэтому во всех случаях семя лишено кожуры. Зародыш у санталовых окружен обильным эндоспермом.
Хотя еще в прошлом веке Г. Мюллер наблюдал опыление цветков насекомыми у ленца, а Г. Иеронимус - у йодины ромболистной, биология опыления санталовых почти не изучена. Но душистость цветков ряда санталовых, венчиковидный околоцветник многих из них, строение соцветий, наличие диска, функционирующего как нектарник, и протандрия говорят о том, что насекомые являются если не единственными, то главными переносчиками пыльцы. В пользу насекомоопыления говорит также гетеростилия, которая наблюдается у ленца промежуточного (Thesium intermedium) и у рода архона. При этом особенно интересно, что у архоны гетеростилия сопровождается диморфизмом пыльцы. Но все же насекомые не являются, вероятно, единственными опылителями санталовых. Есть некоторые основания предполагать, что в двух по крайней мере случаях речь может идти об орнитофилии. По мнению О. Порша (1930), у гавайских видов санталума возможно опыление птицами. Орнитофилия возможна также у андийского рода квинчамалиум. По мнению Дж. Кайта (1969), виды этого рода, особенно чилийский квинчамалиум майский (Q. majus), характеризуются некоторыми особенностями, указывающими на возможность орнитофилии, в том числе довольно большая и длинная венчиковидная чашечка, окрашенная в красный, оранжевый или желтый цвет. По мнению Г. Иеронимуса (1889), у некоторых санталовых не вполне исключено и ветроопыление, особенно у двудомных видов с очень большим числом мужских цветков, как, например, озирис белый (Osyris alba).
В ботанической литературе биологии опыления видов рода ленца уделялось особое внимание. Это объясняется тем, что род ленец хорошо представлен во флоре Европы и поэтому для европейских ботаников был наиболее доступным объектом исследований. В своей "Жизни растений", переведенной еще в начале века на русский язык, австрийский ботаник А. Кернер фон Марилаун подробно описал реакцию пыльников ленца на изменение влажности. При наступлении неблагоприятной погоды пыльники ленца закрываются и тем самым защищают пыльцу. Обыкновенно им нужно для этого лишь несколько минут, а во многих случаях и меньше. Если смочить пыльники ленца альпийского (T. alpinum), то они закрываются уже через 30 с. Кернер считает, что у этого растения смачивание пыльника производится посредством своеобразного пучка волосков, отходящих от околоцветника. Дело в том, что цветки ленца обращены своим зевом кверху и остаются в этом положении постоянно и при любой погоде. Поэтому падающие сверху капли дождя и росы попадают прямо в раскрытый цветок. Однако форма зева и особенности его тканей не дают цветку смачиваться; дождь и роса отлагаются на отгибе и не сразу попадают на пыльники. Тем не менее пыльники тотчас же закрываются, что объясняется тем, что лепестки соединены со стоячими перед ними тычинками пучком скрученных волосков, которые не только сами легко смачиваются, но и проводят воду к пыльнику подобно фитилю у керосиновой лампы; вследствие этого пыльник и закрывается. Но, по мнению английского ботаника М. Ф. Юарта (1892), значение этих волосков заключается в том, что они удерживают пыльцу вблизи зева околоцветника, а также предотвращают продвижение насекомого в поисках нектара по неправильному пути. У видов ленца с волосистым околоцветником волоски задерживают пыльцу посредством выделений своих желёзок. Наконец, было высказано также предположение, что во время цветения волоски как бы регулируют положение пыльников относительно рыльца: волоски удерживают пыльники от соприкосновения с рыльцем и тем самым предотвращают самоопыление. Однако у тех видов ленца, у которых пыльники и рыльце находятся на одном уровне, встречается также самоопыление. Оно возможно также у короткостолбчатых форм гетеростильных видов. Имеются также сведения о клейстогамии у ленца горного (T. montanum), околоцветник которого остается полностью закрытым.
Несколько больше известно о биологии плодов санталовых. Распространение плодов происходит у них почти исключительно с помощью животных, главным образом птиц, а также грызунов, и иногда муравьев. Хотя распространение ветром не наблюдалось ни у одного вида, но наличие у зрелых плодов баклии четырех разросшихся чашелистиков наводит на мысль о возможной анемохории. У санталовых наиболее обычна орнитохория. Экзокарпий у костянок часто красный, оранжевый или желтый, что, несомненно, привлекает птиц. Но интересно, что орнитохорные приспособления имеются и у некоторых орехов, превратившихся в своего рода "ложные костянки". У антоболуса (Anthobolus) и близких родов та часть плодоножки, которая находится непосредственно под плодом, разрастается и становится очень мясистой. Эта разросшаяся часть плодоножки, исполняющая функцию мясистого околоплодника костянки, часто бывает ярко окрашенной в контрастирующие цвета, как у некоторых видов экзокарпоса. Плодами экзокарпоса питаются не только летающие птицы, но и не летающий австралийский эму. Плоды вполне съедобны и для человека. Забавно, что многие писавшие о природе Австралии называли плоды экзокарпоса кипарисовидного (Е. cupressiformis) "вишней с наружной косточкой". Что касается мирмекохории, то еще Е. Ульбрих (1907) обратил внимание на то, что у европейского ленца альпийского плодоножка при созревании плода становится мясистой и может исполнять функцию элайосомы, привлекая муравьев.
У тех видов санталовых, которые паразитируют на ветвях, околоцветник снабжен различными придатками в виде волосков, щетинок и прочее; предполагается, что эти придатки служат для прикрепления плодов к растению-хозяину.
Семейство санталовых подразделяется на 3 трибы - санталовые (Santaleae), ленцовые (Thesieae) и антоболовые (Anthoboleae).
Триба санталовые - самая большая в семействе. Она включает около 27 родов. Несмотря на то что завязь у них нижняя и лишь иногда полунижняя, эта триба считается относительно самой примитивной в семействе. Род санталум - самый известный представитель этой трибы. Он заключает более 20 видов, распространенных в восточной части острова Ява и на Малых Зондских островах, на островах Огасавара (Бонин), на Гавайских островах, на юге Новой Гвинеи, в Австралии, на Новых Гебридах, в Новой Каледонии, на островах Фиджи, островах Тонга, островах Общества, островах Туамоту, на острове Рапа, Маркизских островах и на островах Хуан-Фернандес. На этих последних островах, оторванно от общего ареала рода, некогда произрастал эндемичный санталум фернандесский (S. fernandezianum, рис. 170), который уже в первых десятилетиях нашего века был полностью уничтожен. Последний экземпляр этого вида, старое дерево высотой около 9 м, был сфотографирован в 1908 г. известным шведским ботаником К. Скоттсбергом, но уже в 1916 г. при повторном посещении этого места оно оказалось мертвым. Санталум фернандесский произрастал в лесах, покрывавших часть острова Робинзон-Крузо. Еще в 1624 г. его было много, но в результате хищнической добычи его ценной, долго сохраняющей свой приятный аромат, красной древесины, которую отправляли в Перу, уже к 1740 г. он стал очень редким. В 1978 г. этот вид был уже "посмертно" включен в "Красную книгу" мировой флоры, изданную
Международным союзом охраны природы и природных ресурсов. Слова, написанные в этой книге о санталуме фернандесском, звучат для нас как эпитафия. Виды санталума - деревья или кустарники с вечнозелеными супротивными или редко очередными листьями, с зеленоватыми до желтых или красными обоеполыми цветками и плодами-костянками. Санталум паразитирует на корнях очень многих двудольных и даже на некоторых однодольных. Он может даже паразитировать на корнях других паразитов, например рода кансьера из семейства олаксовых и даже на других особях своего же вида. В Индии насчитывают почти 500 видов растений-хозяев для культивируемого и широко натурализовавшегося там санталума белого (Santalum album). Однако, как указывает английской ботаник Н. Л. Бор (1953) в своей книге "Руководство по индийской лесной ботанике", далеко не все растения одинаково пригодны для санталума, а некоторые из них даже вредны и могут привести к гибели полупаразита. По наблюдениям Бора гибельно для санталума большинство представителей семейства анакардиевых, а также такие растения, как эритрина индийская (Erythrina indica), сапиндус трехлисточковый (Sapindus trifoliatus) и папайя. Но, как ни странно, санталум белый вполне успешно развивается на таком крайне ядовитом растении-хозяине, как стрихнос (Strychnos nuxvomica). Некоторое количество алкалоида стрихнина переходит при этом в ткань сантал ума, доходя до листьев, но он не приносит ему никакого вреда.
В трибу ленцовые входит 5 родов, в том числе ленец, архона и квинчамалиум.
В самую специализированную в семействе трибу антоболовых входят австралийский род антоболус и широко распространенный род экзокарпос (в Малезии, Австралии, Новой Каледонии, на Гавайских островах и в Новой Зеландии).
Для человека главное значение санталовых заключается в ценной древесине некоторых их представителей. Широко известна и имеет наибольшее значение древесина санталума белого (Santalum album), известная под названием "сандаловой древесины" или "сандала" (это название происходит от санскритского candana). Сандаловая древесина - одна из самых драгоценных древесин мира. Она очень душистая, твердая, тяжелая и прочная, устойчивая против термитов. В ядре этой древесины содержится 3-6%, а в корнях - до 10% ценного эфирного масла. Древесину санталума белого использовали в Китае и Индии начиная с V в. до н. э. В древности ее экспортировали в Египет, Грецию и Рим. В настоящее время ее широко используют в Китае и особенно в Индии для резьбы по дереву, изготовления идолов, статуэток, вееров, шкатулок, шкафчиков, курительных палочек и различных сувениров. Измельченная в виде пудры, древесина служит для курения фимиама при совершении различных религиозных церемоний и похоронных ритуалов, а также для косметических целей. Кроме того, получаемое из древесины ствола и корня исключительно ароматное сандаловое масло применяют в парфюмерии и медицине. В диком состоянии санталум белый встречается на Малайском архипелаге от восточной части острова Ява до острова Тимор. Считалось, а некоторыми ботаниками считается и теперь, что этот вид дико произрастает также в Индии. Но ряд ботаников, особенно Ч. Фишер (1927, 1928, 1938), привели убедительные доводы в пользу того, что санталум белый, который уже с незапамятных времен культивируется в Индии, здесь натурализовался и получил широкое распространение. Это мнение поддерживает также Н. Л. Бор (1953). Индия, особенно штат Майсур, является главным в мире поставщиком сандаловой древесины и сандалового масла. Сандаловую древесину в той или иной степени заменяет древесина некоторых других видов санталума, произрастающих на Гавайских островах, островах Фиджи и в Новой Каледонии, а также австралийской эвкарии колосковой (Eucarya spicata), ксимении американской (Ximenia americana), озириса тонколистного (Osyris tenuifolia) и даже некоторых представителей других семейств. Наиболее знаменита древесина эндемичного для Фиджи и Тонга санталума яси (S. yasi). В течение долгого времени ее высоко ценили в Китае, где использовали для совершения различных религиозных обрядов. Очень высокая цена этой древесины в начале XIX в. вызвала целое нашествие жадных до наживы авантюристов на острова Фиджи. "Освоение" островов Фиджи европейцами фактически началось именно в связи с торговлей сандаловой древесиной между 1800 и 1805 гг. "Бум" продолжался только десять лет (между 1804 и 1816 гг.), и в результате "яси" представляет собой очень редкое растение, находящееся под охраной. Вся история торговли фиджийской сандаловой древесиной полна жестокостей, убийств и кровавой резни.
Некоторые санталовые, как ксимения американская, доставляют довольно хорошую древесину, употребляемую для производства мебели. Следует упомянуть также капский колпоон сжатый (Colpoon compressum), кору которого используют как дубильное сырье.
Есть среди санталовых и пищевые растения. Плоды некоторых из них, как, например, южноамериканский акантосирис серповидный (Acanthosyris falcata), гималайская пирулария съедобная (Pyrularia edulis), так же как утолщенные мясистые плодоножки некоторых видов (особенно экзокарпоса кипарисовидного - Exocarpos cupressiformis), вполне съедобны. Съедобны также сладкие клубни "макачи", или архоны клубненосной (Arjona tuberosa),- многолетней травы, обитающей в Патагонии.
Среди санталовых имеются также лекарственные растения.
Семейство мизодендровые (Misodendraceae) (З. И. Никитичева)
К санталовым, особенно к родам архона и квинчамалиум, стоит близко семейство мизодендровых. Оно состоит из одного-единственного рода мизодендрум (Misodendrum, рис. 171), заключающего около 11 видов. Все они обитают в субантарктических нотофагусовых лесах Чили и Аргентины от 33° ю. ш. до Магелланова пролива.

Рис. 171. Мизодендрум точечный (Misodendrum punctulatum) на веточке нотофагуса антарктического: 1 - общий вид растения; 2 - мужское соцветие; 3 - мужской цветок; 4 - женское соцветие; 5 - соплодие; 6 - завязь
Виды мизодендрума - кустарнички, ведущие полупаразитический образ жизни исключительно на видах рода нотофагус (Nothofagus) из семейства буковых. Большинство видов мизодендрума с мелкими очередными листьями. Но есть виды с листьями, редуцированными до чешуи, что создает впечатление членистости ветвей. Мизодендрумы - двудомные растения с очень мелкими цветками, собранными в сложные кистевидные соцветия. Мужские цветки без околоцветника, с 3 или 2 тычинками, сидячими вокруг небольшого подкововидного нектарного диска. Женские цветки несут гинецей из 3 плодолистиков, с толстым столбиком и с 3 рыльцами. Завязь заключена в 3 приросших к ней, но свободных по краям чашелистика. В щель между краями этого своеобразного околоцветника высовываются стаминодии, превращенные в щетинки, покрытые одноклеточными волосками. После опыления эти щетинки сильно удлиняются, достигая от 10 до 85 мм, и сохраняются при плодах. В полости завязи образуется толстая центральная колонка с 3 лопастями, представляющими редуцированные беспокровные семязачатки, из которых только один созревает в семя. Зародышевый мешок развивает длинный трубчатый гаусторий в плацентарную колонку, достигающий проводящих пучков в основании цветка. По этой особенности мизодендрум сходен с олаксовыми.
Процесс опыления не изучен. По-видимому, имеет место перекрестное опыление с помощью небольших насекомых. Плодики содержат одно семя с редуцированным покровом, зеленым эндоспермом и прямым зародышем, с небольшими сросшимися семядолями и корешком. В сезон плодоношения женские растения украшены массой щетинок. Из-за них мизодендрум иногда называют "оперенной омелой". Небольшие легкие плоды, снабженные тремя щетинками, покрытыми остроконечными или крючковатыми волосками, разносятся ветром. Они цепляются за ветви окружающих нотофагусов и прикрепляются к ним. Здесь они и прорастают. Из корневого конца зародыша развивается толстая дисковидная присоска с тонким липким краем. Ее внутренняя часть внедряется в ветвь растения-хозяина. Вокруг радиально разросшейся гаустории образуются чашеподобные наплывы ткани хозяина. Симподиально ветвящиеся стебли паразита могут быть толще ветви растения-хозяина, и последняя обычно отмирает за местом его прикрепления.
Семейство ремнецветниковые, или лорантовые (Lobanthaceae) (З. И. Никитичева)
Семейство ремнецветниковых - наиболее крупное в порядке санталовых. Оно включает от 30 до 65 родов (в зависимости от понимания объема рода) и не менее 900 видов, широко распространенных по земному шару, главным образом в тропических странах. Лишь отдельные виды заходят в субтропическую и умеренную зону. Наиболее крупный род семейства - ремнецветник (Loranthus), содержащий даже при узком его понимании более 200 видов. Почти все они обитают в тропиках Старого Света, только несколько видов внетропические. В нашей стране, в юго-западных областях, встречается ремнецветник европейский (L. europaeus, рис. 172) - небольшой листопадный кустарник, растущий на ветвях дуба черешчатого и дуба скального.
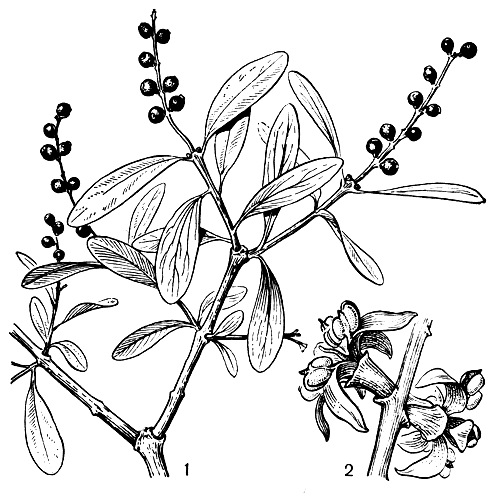
Рис. 172. Ремнецветник европейский (Loranthus europaeus): 1 - ветвь с плодами; 2 - цветки
Подавляющее большинство ремнецветниковых - зеленые стеблевые паразиты (полупаразиты), использующие в качестве хозяев широкий круг растений из различных семейств цветковых растений. Некоторые виды являются "наземными" деревьями и кустарниками, развивающими корни или корнеподобные органы до нескольких метров длиной. Эндемик Западной Австралии - нуйтсия обилъноцветущая (Nuytsia floribunda), называемая "рождественским деревом" за яркие желтовато-оранжевые цветки, появляющиеся к новому году, достигает в высоту 10 м. Только недавно был установлен паразитизм этого растения и обнаружены гаусториальные связи корней нуйтсии с корнями различных соседних растений, в том числе культивируемой моркови.
В семействе ремнецветниковых поражает разнообразие жизненных форм и способов прикрепления к питающим растениям. Виды рода гайадендрон (Gaiadendron), растущие в Америке (в Андах от Перу до Колумбии), встречаются как корневые и стеблевые паразиты. В Бразилии встречается лиана фригилантус остролистный (Phrygilanthus acutifolius) с подземными корнями и деревянистым стеблем, высоко поднимающим облиственные ветви и цветки. Как на корнях, так и на стебле этого паразита образуются присоски (гаустории), внедряющиеся в соответствующие органы растения-хозяина.
Большинство исследователей считает, что у ремнецветниковых корневой паразитизм первичен, а стеблевой - вторичен. Одна группа стеблевых паразитов образует корнеподобные органы (эпикортикальные корни), растущие по поверхности ветвей растения-хозяина. Эти сильно преобразованные органы морфологически, вероятно, гомологичны придаточным корням. Через определенные интервалы на них образуются погружающиеся в ткань растения-хозяина гаустории, с помощью которых паразит закрепляется на ветви и получает из нее воду и питательные вещества. Такой способ паразитирования характерен, например, для вида струтантуса (Struthanthus) и представителей некоторых других родов. У более специализированных паразитов мощная гаусториальная система в виде длинных ветвящихся тяжей или аморфных образований развивается внутри стеблей растения-хозяина. На их поверхности появляются только побеги и соцветия паразита, возникающие в разных точках из внутренних тканей. Такой глубокий паразитизм характерен для некоторых видов ремнецветника и пситтакантуса (Psittacanthus). Почти все ремнецветниковые размножаются вегетативно. Гаустории у этих паразитов образуются из всех вегетативных органов - корней, стеблей и даже листьев. У тропического американского вида струтантуса округлого (S. orbicularis) свисающие тонкие молодые ветви имеют крепкие загнутые листья с крючковатой вершиной. Когда такой лист зацепляется за ветвь дерева, черешок закручивается вокруг нее, и вскоре в этом узле образуются эпикортикальные органы, охватывающие ветвь хозяина и внедряющие в нее свои гаустории. Известны случаи самопаразитизма наземных и эпикортикальных корней. В условиях тропического леса с его обильной растительностью часто наблюдается прикрепление одного паразита сразу к нескольким растениям-хозяевам и даже к другим паразитным растениям. Ремнецветниковые - кустарники или небольшие деревья. Самым крупным представителем семейства является монотипная западно-австралийская нуйтсия (Nuytsia) - дерево высотой до 10 м. Листья цельные, тонкие или мясистые, обычно расположенные супротивно, очень редко очередные или мутовчатые. Характерны отчетливые 3-5 жилок, перисто-расположенные или проходящие вдоль листа.
Лишь у некоторых растений листья редуцированы до чешуи, таков фригилантус безлистный (Phrygilanthus aphyllus). Цветки у ремнецветниковых обычно собраны в верхоцветные соцветия, актиноморфные или слабо зигоморфные, обоеполые или однополые (большей частью двудомные). Под околоцветником находится слегка лопастное или более или менее усеченное кольцевидное образование, называемое каликулусом.
Многие ботаники считают каликулус сильно редуцированной чашечкой, хотя и не все согласны с этим. Наиболее убедительным доводом в пользу этого мнения считается хорошо развитый 5-7-лопастный каликулус нуйтсии, который по своему анатомическому строению очень похож на чашечку. Вопрос, однако, остается открытым. Околоцветник (венчик, или чашечка, в зависимости от того, отождествляем ли мы каликулус с чашечкой или нет) состоит из 4-6 (до 9) долей, свободных или сросшихся в трубку. Тычинки в одинаковом числе с долями околоцветника и расположены супротивно им. Они чрезвычайно разнообразного строения. Пыльники при созревании раскрываются продольной щелью или верхушечными порами. Гинецей образован 3-4 сросшимися плодолистиками, с длинным или сильно укороченным столбиком; завязь нижняя, 1-гнездная, с толстой центральной колонкой, несущей полностью слившиеся с ней семязачатки. У ряда представителей семейства можно наблюдать постепенную редукцию колонки вплоть до полного ее исчезновения. В последнем случае женский гаметофит развивается в ткани основания завязи. В развитии зародышевых мешков их много удивительных особенностей. Прежде всего они необычайно сильно удлиняются и врастают в столбик, а у некоторых видов достигают даже рыльца. После оплодотворения зигота опускается в полость завязи благодаря росту подвеска. Кроме того, в завязи часто развиваются 6-7 зародышевых мешков, сливающихся на стадии развития эндосперма в общую структуру с несколькими молодыми зародышами, из которых только один достигает зрелости. Зародыш обычно крупный, с развитыми прямыми семядолями, которые выше почечки часто сливаются, образуя трубку.
У фригилантуса безлистного зародыш небольшой, без семядолей. Корневой конец зародыша без зачаточного корешка, обычно расширенный и плоский, и, по существу, представляет собой гипокотиль. Клетки его эпидермы с сосочковидными выростами, играющими определенную роль в закреплении зародыша на ветвях растения-хозяина. У большинства ремнецветниковых эндосперм обильный, но у некоторых родов он может отсутствовать. Плоды обычно ягодообразные, односемянные, с ярко окрашенным и мясистым околоплодником, редко сухие или костянковидные. Под мясистым наружным слоем расположен клейкий слой - висцин, заключающий внутренние ткани перикарпия и эндосперм с зародышем. У отдельных видов висцин представлен кольцом ткани в базальной или терминальной части плода. Этот слой из ослизняющихся клеток является замечательной особенностью ремнецветниковых, связанной с их приспособлением к стеблевому паразитизму и распространением плодов с помощью птиц.
Ремнецветниковые опыляются насекомыми или птицами. Многие тропические виды имеют крупные, ярко окрашенные цветки, приспособленные к опылению птицами. В Америке главными опылителями являются колибри, в Старом Свете - цветососовые (Dicaeidae) и нектарнициевые (Nectariidae), а в Австралазии также медососовые (Meliphagidae). Привлекаемые обильным нектаром и сладкими выделениями на рыльце, птицы принимаются за работу еще до восхода солнца, когда многие цветки еще не открылись. Птицы надавливают на верхушку цветка своим клювом или вставляют его между долями околоцветника, и цветок раскрывается. Австралийский орнитолог С. А. Паркер (1977) так описывает опыление лизианы внеплодниковой (Lysiana exocarpi), паразитирующей в большом количестве на видах акации. Цветки оказались наполненными около одной трети нектаром. Во время выделения нектара лопасти трубчатого околоцветника плотно сомкнуты, что, вероятно, защищает нектар от нежелательных посетителей. Но, как только клюв медососа проникает в околоцветник, лопасти отходят в сторону. Вентральная лопасть трубки околоцветника, которая более свободна, чем остальные пять, отклоняется сильнее, почти на половину длины всей трубки, что позволяет птице достичь нектара снизу.
С. А. Паркер исследовал также цветки двух австралийских видов рода амиема (Аmуеmа). Их строение сходно с лизианой, за исключением того, что доли околоцветника свободны до основания. Но и в этом случае доли околоцветника плотно сомкнуты во время выделения нектара.
Интересно, что в Индии малабарский дрозд (Sturnus malabaricus) иногда добывает нектар из цветков ремнецветника.
У ремнецветника европейского собранные в кисти ягодообразные желтые плоды издали заметны и охотно поедаются дроздами. У тропических видов семейства плоды обычно распространяются теми же видами птиц, которые являются их опылителями. Из желудка птиц вместе с экскрементами семена, окруженные клейким слоем, попадают на ветви деревьев и приклеиваются к ним. Ось зародыша удлиняется и соприкасается с поверхностью ветви, плотно прилипает к ней и расширяется. Из образовавшейся присоски возникает группа агрессивных клеток, внедряющихся в ткань ветви растения-хозяина до проводящего пучка. Эта погруженная первичная гаустория осуществляет снабжение проростка паразита питательными веществами. После заякоривания концы семядолей освобождаются от эндосперма, а из почечки развивается побег паразита. От первичной гаустории развиваются впоследствии придаточные корнеподобные органы, или интраматрикальные тяжи, а из них - вторичные гаусториальные органы и побеги паразита.
Ремнецветники вызывают серьезные повреждения цитрусовых, эвкалиптов и других миртовых деревьев, какао, гевеи, камфорного лавра. Поражают они и насаждения криптомерии японской. Кроме ухудшения роста растений, на которых они поселяются, они вызывают образование на них гипертрофированных галлов, образующихся в результате сильного и причудливого разрастания тканей растения-хозяина в местах образования гаусториев паразита. Размер этих галлов может быть с человеческую голову, и они сильно отягощают несущие их ветви.
Семейство омеловые (Viscaceae) (З. И. Никитичева)
В облике омеловых имеется много общего с ремнецветниковыми и чаще всего они объединяются в одно семейство с двумя подсемействами. Но по современным представлениям омеловые имеют независимое от ремнецветниковых происхождение от санталовых, причем, вероятно, они происходят от другой группы санталовых, чем ремнецветниковые. В семействе 11 родов и около 510 видов. Оно распространено главным образом в тропических и субтропических областях и только отдельные виды заходят в умеренную зону. Относительно далеко на север продвинулись виды омелы (Viscum) и арцеутобиума (Arceuthobium) в материковой Евразии и форадендрона (Phoradendron) и арцеутобиума в Северной Америке. Один вид большого и широко распространенного рода корталселла (Korthalsella japonica) встречается в Японии.
Омеловые - кустарники или реже травы, ведущие обычно полупаразитический образ жизни на ветвях деревьев. Но некоторые виды арцеутобиума и форадендрона, с редукцией листьев, перешли к почти полному паразитизму. Омеловые паразитируют как на голосеменных, так и на цветковых растениях. Иногда (омела членистая - V. articulatum) они паразитируют на ремнецветниковых и даже на других омеловых (в данном случае на видах рода нототиксос - Notothixos). Листья, когда они развиты, супротивные, обычно с характерным дуговидным жилкованием. Имеются все переходы от хорошо развитых зеленых листьев к лишенным хлорофилла мелким чешуям, что можно особенно хорошо проследить в родах форадендрон, омела и дендрофтора (Dendrophtora). Роды же арцеутобиум, корталселла и эвбрахион (Eubrachion) имеют лишь чешуевидные листья. Эта редукция листьев не обязательно связана с понижением фотосинтетической активности, с более полным переходом к паразитизму. Арцеутобиум и корталселла имеют достаточно много хлорофилла, а форадендрон с чешуевидными листьями бросается в глаза своей яркой зеленью. У целого ряда родов омеловых, как дендрофтора, омела, форадендрон, корталселла и гиналлоа (Ginalloa), подобно южноамериканскому роду иксокактус (Ixocactus) из ремнецветниковых, происходит постепенное уплощение и расширение междоузлий, которые, таким образом, берут на себя функцию фотосинтеза.
Большинство омеловых развивает гаусториальную систему внутри тканей растения-хозяина. Но два неотропических рода - антидафна (Antidaphne) и эремолепис (Eremolepis)- характеризуются тем, что развивают на поверхности ветвей так называемые эпикортикальные корни, снабженные гаусториями.
За исключением специализированного рода иксокактус, цветки омеловых всегда бывают собраны в соцветия и всегда сидячие или почти сидячие. В отличие от ремыецветниковых цветки омеловых мелкие, длиной обычно менее 2 мм, а у многих видов дендрофторы даже менее 1 мм. Это невзрачные цветки обычно зеленоватые или желтоватые и лишь у некоторых видов арцеутобиума ярко-красные. Они всегда однополые, однодомные или двудомные, и лишь для арцеутобиума наименьшего (A. minutissimum) однажды указывалось наличие обоеполых цветков.
Околоцветник простой, 2-4-членный, сильно редуцированный, особенно в женских цветках, а у рода арцеутобиум он почти не выражен. Тычинки в одинаковом числе с долями околоцветника, свободные или приросшие к ним. В семействе омеловых наблюдается постепенная редукция тычинок, выражающаяся как в редукции тычиночной нити, так и в уменьшении числа гнезд пыльника, который у дендрофторы стал одногнездным. Гинецей омеловых состоит из 3-4 плодолистиков, с коротким столбиком, заканчивающимся массивным рыльцем; завязь нижняя, 1-гнездная, с короткой редуцированной центральной колонкой, иногда лопастной, несущей полностью слившиеся с ней семязачатки, которые у омеловых достигли крайней степени упрощения и морфологически уже не выражены. У некоторых видов омелы отсутствует даже центральная колонка и женский гаметофит развивается прямо в ткани основания завязи. Плоды ягодообразные, сочные и ярко окрашены, с 1 или реже 2- 3 семенами. В стенке плода развивается характерный клейкий слой (висцин), расположенный обычно внутри от проводящих пучков, в то время как у ремнецветниковых он расположен снаружи. Семена без кожуры, с мясистым эндоспермом, окружающим зародыш, кроме его корневой части.
Омеловые обычно опыляются насекомыми, но в некоторых случаях, как у омелы белой (V. album, рис. 173) и арцеутобиума, установлено также ветроопыление. У арцеутобиума в области рыльца выделяется капелька жидкости, которая служит для улавливания пыльцы. С усыханием этой капельки пыльца приходит в соприкосновение с рыльцем. Но главными опылителями омеловых являются насекомые, в основном перепончатокрылые, которых привлекает выделение нектарного диска и особенно пыльца. У некоторых родов, как у антидафна, пыльца является единственным аттрактантом для насекомых. Есть указания, что омелу белую опыляют мухи.
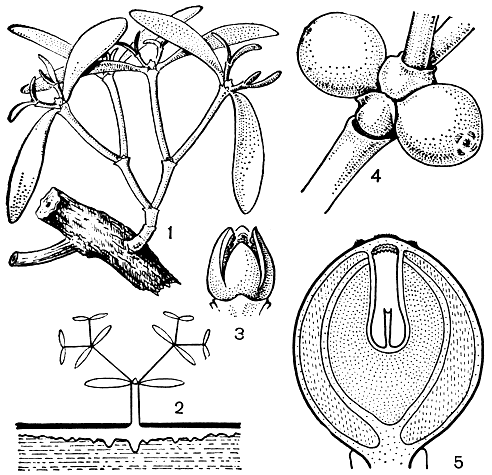
Рис. 173. Омеловые. Омела белая (Viscum album): 1 - молодое растение на стебле груши; 2 - схема строения омелы с олиственным тюбегом и гаусториальным тяжем, развивающимся под корой ветви растения-хозяина; 3 - мужской цветок. Омела окрашенная (V. coloratum): 4 - плоды; 5 - продольный разрез зрелого плода
В строении плода, семени и зародыша ярко выражены приспособительные признаки к особенностям распространения и прорастания омеловых. Их ягодообразные плоды и распространяются главным образом птицами. Наряду с эндозоохорией наблюдается также эпизоохория, выраженная в том, что птицы, очищая свой клюв на ветвях деревьев, оставляют там клейкие семена (как у видов омелы). Но орнитохория характерна не для всех омеловых. Плоды арцеутобиума, взрываясь, выбрасывают клейкие семена на значительное расстояние (пример активного баллиста).
Процесс прорастания семян омеловых замечательно преобразован в связи с особенностями их паразитизма. Наиболее хорошо он изучен у омелы. Семена прорастают на ветвях растений-хозяев. Корневой конец зародыша изгибается к поверхности ветви и при соприкосновении с ней расширяется в дисковидную присоску. Из ее центра образуется группа клеток, которые внедряются в стебель растения-хозяина до его проводящего пучка. На следующий год развивается облиственный побег, а из присоски, вклиненной в ткань растения-хозяина, возникают так называемые "коровые корни" - зеленоватые, почти цилиндрические тяжи. Они растут вдоль питающей их ветви, между корой и древесиной, вверх и вниз от места внедрения первичного гаустория. В дальнейшем на этих тяжах, перпендикулярно к ним, образуются вторичные гаустории, из них развиваются новые побеги паразита, поражающего постепенно все тело растения-хозяина. Поэтому срезание кустов омелы не уничтожает паразита. У арцеутобиума основная часть тела представлена множеством переплетающихся тяжей в стебле питающего растения, достигающих точки роста побега. Устанавливается изофазность (одновременность) цветения паразита и хозяина. У некоторых омеловых все вегетативное тело паразита находится внутри стеблей растения-хозяина и на поверхности появляются только цветки.
Самые большие роды в семействе - это форадендрон (около 190 видов) и омела (около 100 видов). Все виды рода форадендрон обитают в тропической Америке. Это кустарники с ложнодихотомическим ветвлением, образующие густые пучки побегов на ветвях растений-хозяев. Большинство видов - вечнозеленые растения, с крупными, цельнокрайними, супротивно расположенными листьями. Но у некоторых видов листья редуцированы до чешуи. Форадендрон либоцедрусовый (P. libocedri) образует свисающие безлистные побеги. В качестве хозяев форадендрон предпочитает деревья с твердой древесиной, особенно хвойные (можжевельник, пихта, кипарис). Некоторые его виды в Калифорнии поражают деревья из семейства бобовых, другие растут на дубе, ильме, грецком орехе. Отмечалась деформация и гибель деревьев, пораженных паразитом. В месте его внедрения на стебле растения-хозяина образуются вздутия из-за разрастания тканей. Эти причудливо деформированные ветви используют для различных поделок.
Виды омелы распространены главным образом в тропических и субтропических областях Азии и особенно Африки, а также в Новой Гвинее, в Австралии и в умеренных областях Евразии. Виды омелы кустарники или травы. Ветви в поперечном сечении округлые, гранистые или уплощенные, прямостоячие или висячие. Листья супротивные или реже мутовчатые, у ряда видов редуцированы до чешуи, как у омелы непальской (V. nepalense), или даже полностью отсутствуют, как у омелы угловатой (V. angulatum). У южноафриканской омелы наименьшей (V. minimum), паразитирующей на видах молочая, редуцированы не только листья, но и стебли. Цветки однодомные или двудомные, зеленоватые, желтые или беловатые, в пазушных или верхушечных соцветиях. Плоды белые, желтые, оранжевые или красные, содержат клейкое вещество висцин.
В нашей стране 2 вида омелы: омела белая в средней и южной полосе европейской части СССР и на Кавказе и омела окрашенная (V. coloration) - на Дальнем Востоке. Оба эти вида настолько близки, что многие ботаники рассматривали омелу окрашенную как подвид или даже как разновидность омелы белой. У омелы белой различают 3 подвида. Один из них паразитирует на лиственных деревьях - груше, яблоне, тополе, клене, дубе; два других - на хвойных деревьях - пихте, сосне и лиственнице. Из плодов омелы получают так называемый птичий клей, используемый для ловли мелких птиц. Экстракт из молодых листьев омелы белой применяют для лечения ранних стадий гипертонической болезни.
Небольшой род арцеутобиум заключает 15 видов, распространенных главным образом во внетропических областях северного полушария, но некоторые его виды достигают Малайского полуострова, Явы и Вест-Индии. Это маленькие кустарники с супротивными чешуевидными листьями и однополыми цветками, паразитирующие исключительно на голосеменных растениях. Некоторые виды имеют крошечные размеры. Так, североамериканский арцеутобиум крошечный (A. pusillum) не превышает в высоту 1-2 см, а о западногималайском арцеутобиуме наикрошечном (A. minutissimum) знаменитый английский ботаник Дж. Xукер писал: "Это самое крошечное двудольное растение, какое я могу вспомнить". Наиболее широко распространен арцеутобиум можжевельниковый (A. oxycedri, рис. 174, табл.43), растущий на можжевельнике в Северной Америке, Африке, Азии и Европе. Этот род является карликовым паразитом, с небольшими пучками слабоодревесневающих тонких желтоватых ветвей. Мелкие чешуевидные листья срастаются в основании и придают ветвям членистый вид. Наибольшее число видов рода (5) встречается в Северной Америке, здесь они паразитируют на пихтах, елях и соснах. Паразит вызывает уменьшение прироста пораженных деревьев, их деформацию, а нередко и большой процент гибели, особенно в культурных посадках.
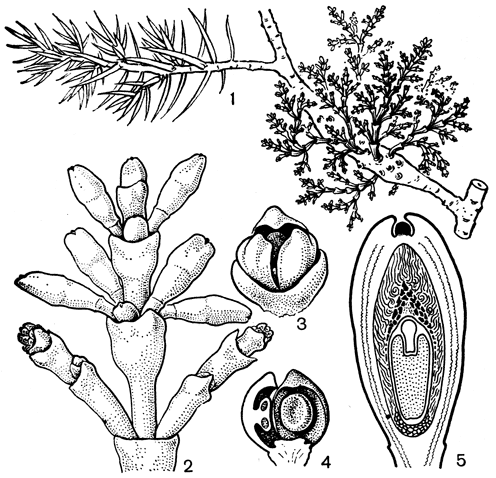
Рис. 174. Арцеутобиумможжевельниковый (Arceuthobium oxycedri): 1 - общий вид растения на ветке можжевельника; 2 - часть ветви с женскими (вверху) и мужскими (внизу) цветками; 3 - мужской цветок; 4 - мужской цветок в разрезе (видны тычинки, приросшие к листочкам околоцветника); 5 - продольный разрез зрелого плода (эндосперм и зародыш окружены клейким слоем внутренней части стенки плода)

Таблица 43. Омеловые и крушиновые: 1 - арцеутобиум можжевельниковый (Arceuthobium oxycedri), паразитирующий на можжевельнике, Кавказ
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'