Порядок аралиевые (Araliales)
Семейство аралиевые (Araliaceae) (И. В. Грушвичкий)
Семейство включает более 850 видов, относящихся примерно к 70 родам. Самый крупный - род шефлера (Schefflera, табл. 41), 250-300 видов которого представляют аралиевые во всех влажнотропических областях Земли, особенно богато в тропической Азии. Более 120 видов рода ореопанакс (Огеорапах) распространены в тропической и субтропической Америке. Наконец, в тропиках Старого Света обитает примерно 100 видов рода полисциас (Polyscias). Три тропических в основном рода - и более половины всех видов семейства! Зато большинство остальных родов включает всего по 2-5 (олиготипные) или по одному (монотипные) виду, примерно по 40% тех и других. И все эти виды - тоже большей частью жители тропиков.

Таблица 41. Аралиевые: 1 - шефлера Скортечини (Schefflera scortechinii), Малайзия
Таким образом аралиевые - семейство в основном тропическое и субтропическое. Наиболее богато они представлены в Восточной и Юго-Восточной Азии, на островах Тихого океана и в Австралии, а также в тропической Америке, т. е. в областях, относящихся преимущественно к влажным тропикам и субтропикам.
Лишь немногие виды аралиевых распространены в умеренной зоне, при этом наблюдается их приуроченность к приокеаническим территориям материков. На западе Европы единственным представителем является плющ обыкновенный (Hedera helix), но только на юге советского Дальнего Востока, помимо женьшеня (Panax ginseng), в лесах распространено еще 9 видов. Это и высокое (до 28 м) дерево первого яруса южноуссурийских лесов калопанакс семилопастный (Kalopanax septemlobus), и колючие кустарники элеутерококк колючий (Eleutherococcus senticosus, табл. 41), акантопанакс сидячецветковый (Acanthopanax sessiliflorus) и высокогорная заманиха, или оплопанакс высокий (Oplopanax elatus, табл. 41). Наконец, это пять видов рода аралия (Aralia). Поразительно сходно с этим распространение аралиевых в умеренной зоне Северной Америки. На ее северо-западе до Южной Аляски встречается всего один вид - оплопанакс ощетиненный (O. horridus), настолько близкий к восточноазиатской заманихе, что в свое время их считали одним видом. А в лесах восточной части материка обитают два вида из того же рода панакс, что и женьшень и четыре вида аралии.

Таблица 41. Аралиевые: 6 - элеутерококк колючий (Eleutherococcus senticosus), Приморский край

Таблица 41. Аралиевые: 7 - оплопанакс высокий (Oplopanax elatus), Приморский край

Таблица 41. Аралиевые: 2 - плющ крымский (Hedera taurica) на стволе платана восточного, Никитский ботанический сад
Аралиевые большей частью деревья и кустарники (в том числе лианы, эпифиты и полуэпифиты), редко кустарнички, полукустарники и многолетние корневищные травы. Самое высокое аралиевое - дерево тропических дождевых лесов Новой Гвинеи и ближайших (к востоку от нее) островов гастония великолепная (Gastonia spectabilis) со стволом высотой до 40 м, диаметром до 175 см. Насколько известно, это единственное аралиевое с досковидными корнями. Большинство же деревьев из аралиевых низкорослы, высотой до 10-15 м. Некоторые из них (представители разных родов) относятся к особой форме так называемых розеточных деревьев. Ствол их, по крайней мере в молодом возрасте, неветвящийся, крупные листья собраны расходящимся пучком наверху ствола, образуя ровную шаровидную крону. Аралиевые такого габитуса уже издали легко узнаются среди огромного разнообразия других тропических деревьев. С возрастом ствол может разделиться ложновильчато на две ветви, затем каждая из них на одном уровне может дать два или три новых ответвления. но и такая разветвленная верхушка нередко остается скрытой в пределах по-прежнему шаровидной кроны.
Многие роды аралиевых целиком или почти целиком представлены кустарниками. Типичным кустарником является элеутерококк колючий (табл. 41), ветвящиеся от основания побеги которого покрыты тонкими игольчатыми шипами, с чем связаны бытующие на Дальнем Востоке его народные названия "чертов куст" и "нетронник". Попутно отметим, что колючесть стволов и ветвей в ряде случаев также черешков и листовых пластинок, характерная для многих аралиевых и, несомненно, защищающая их от животных, является, как правило, хорошим родовым признаком, в частности диагностическим признаком таких родов, как тревезия (Trevesia), брассайопсис (Brassaiopsis), оплопанакс (Oplopanax) и ряда других.
Лианы в семействе только древовидные, но относятся к разным типам. Наиболее примитивный тип представлен монотипным родом тупидантус (Tupidanthus). Одно из древнейших аралиевых тупидантус колпачковый (T. calyptratus), обитающий в горных лесах Индии (Восточные Гималаи), Китая (Юньнань), Таиланда, Бирмы и Вьетнама, первые годы растет как обычное неветвящееся дерево, но позднее обвивает близстоящее дерево-хозяин, поднимаясь по нему на высоту 20 м и более. Кустарниковые лианы, напротив, почти с самого начала развития приобретают черты лиан, примером чего может служить широко распространенный в тропической Азии акантопанакс трехлистный (Acanthopanax trifoliatus) с его гибкими шиповатыми побегами, цепляющимися за окружающие кустарники.
Наиболее специализированный тип лиан представляют виды плюща. У этих "лиан-корнелазов" молодые вегетативные побеги прикрепляются к опоре придаточными корнями-прицепками. Совершенно другой характер имеют их репродуктивные побеги (табл. 39). Они короткие, свободно возвышаются над мозаичным покровом плюща, лишены корней-прицепок и несут листья столь отличные от листьев стерильных побегов, что те и другие можно принять за листья разных растений. С возрастом главные оси стеблей, утолщаясь, превращаются в искривленные, покрытые коркой стволы, иногда достигающие в обхвате 2 м. От Канарских островов до Тихого океана распространено около 15 видов плюща. Из них особо следует упомянуть крупнолистный плющ колхидский (Hedera colchica, табл. 41) целым водопадом листьев покрывающий стволы деревьев в богатых колхидских лесах. Близкие виды растут в Китае. Для корней плюща обыкновенного (как и у ряда других аралиевых) характерна эндомикориза. В то же время на них можно встретить паразитирующую заразиху плющовую (Orobanche hederae). По другую сторону экватора на корнях ряда новозеландских аралиевых паразитирует эндемичный для Новой Зеландии своеобразный цветковый паразит из семейства баланофоровых (Balanophoraceae) дактилантус Тейлора (Dactylanthus taylorii).

Таблица 41. Аралиевые: 5 - плющ колхидский (H. colchica), Уссурийский заповедник, Приморский край
Некоторые тропические аралиевые проявляют способность к эпифитному и полуэпифитному образу жизни. К полуэпифитам принадлежит ряд видов шефлеры, как азиатских, так и южноамериканских. Они поселяются на стволах деревьев тропического дождевого леса, часто на большой высоте. Там и здесь под пологом крон можно увидеть свисающий вдоль ствола дерева-хозяина и укоренившийся в почве канатовидный воздушный корень, и только подняв голову, заметить в просветах между ветвями сам полуэпифит с его пальчатосложными листьями.
Многолетние травянистые растения в семействе немногочисленны. В северном полушарии это 7-8 видов рода панакс, а в южном - род стилъбокарпа (Stilbocarpa), включающий три вечнозеленых вида, образующих заросли в суровых условиях крайнего юга Новой Зеландии и прилегающих субантарктических островов. Немногочисленные травянистые и полукустарниковые виды имеются, кроме того, среди аралий и в немногих других родах.
Весьма своеобразна жизненная форма видов рода панакс, в частности женьшеня. Для этого реликтового растения тенистых хвойно-широколиственных лесов юга советского Дальнего Востока, севера Кореи и северо-востока Китая характерно крайне медленное развитие. Семена в природе прорастают не ранее чем на второй год после рассеивания. Строение взрослого надземного побега достигается обычно только у 8-10-летних растений. У взрослого женьшеня на верхушке прямого тонкого стебля изящной правильной розеткой располагаются на длинных черешках пятипальчатосложные листья, а из центра этой розетки как продолжение стебля поднимается цветонос с простым зонтиком мелких невзрачных цветков, позднее ярко-красных плодов-костянок (табл. 41).
Постоянными многолетними органами женьшеня являются корневище и мясистый главный корень, причем это редкий среди корневищных трав случай длительного, в течение многих десятилетий, сосуществования того и другого. При ежегодном осеннем опадении листьев со стеблем на корневище остаются рубцы, по числу которых можно установить возраст растения. Самые крупные из найденных "корней", массой до 300-400 г, могут быть старше многих окружающих женьшень древесных пород, имея возраст до 200 и, возможно, более лет. И сколько же интересного и необычного происходит за столь долгую жизнь! Это и ежегодное сокращение корня и втягивание им в почву подрастающего корневища, вследствие чего сам корень прячется в землю и постепенно приобретает наклонное, а затем и горизонтальное положение. Это и способность корня с корневищем после сильных повреждений впадать в "сон", продолжающийся годы и даже десятки лет, и многое другое, дававшее в древности пищу для всяческих суеверий.

Таблица 41. Аралиевые: 4 - дикорастущий женьшень (Panax ginseng), Уссурийский заповедник, Приморский край
Листорасположение у большинства аралиевых очередное. Листья в подавляющем большинстве сложные, иногда очень крупные в семействе листья, достигающие вместе с черешком трехметровой длины. Наиболее часты пальчатосложные листья, характерные для видов огромного рода шефлера, а также для многих других родов. У разных представителей они значительно отличаются по величине, по числу листочков в листе, по их форме и расчлененности. И. В. Грушвицким и Н. Т. Скворцовой (1970) было показано, что у некоторых видов шефлеры пальчатосложные листья их предков в ходе эволюции преобразовались в своеобразные пучковатосложные листья, многочисленные листочки которых расположены на конце черешка не веером, как обычно, а пучком как цветки в зонтике.
Аралиевых с перистосложными (до трижды перистых) листьями меньше. Помимо крупного рода полисциас, сюда из упомянутых выше относятся виды родов аралия и гастония.
Разнообразны в семействе и различные типы простых листьев от цельных, иногда очень крупных (длиной почти 1,5 м) до пальчатых и перистых с различной степенью расчлененности листовой пластинки. Наконец, на простые цельные листья внешне похожи сложные однолисточковые листья некоторых аралиевых, например из числа мадагаскарских видов полисциаса.
Общим для подавляющего большинства аралиевых является роднящее их с зонтичными наличие в основании черешка широкого и более или менее охватывающего стебель влагалища. При опадении листьев на стебле остаются заметные листовые рубцы серповидной формы с 5, 7, 9 и значительно большим числом следов проводящих пучков.
В жизни тропических видов сильно развитые влагалища имеют большое значение: они поддерживают крупные и тяжелые листья, а вместе с прилистниками, то образующими каемки по краям влагалища, то срастающимися в пазушный язычок, защищают спящие и особенно верхушечные почки с зачатками побегов, заменяя отсутствующие в этом случае почечные чешуи.
Соцветия у аралиевых очень разнообразны как по размерам, так и по строению. В большинстве случаев это сильно разветвленные соцветия, которые при всех отличиях до сих пор принято обозначать одним сборным термином - метелка. Элементарной частью в них у большинства родов является зонтик, реже головка, кисть или колос. На разных представителях аралиевых можно проследить фактически все этапы упрощения от крупных сложно разветвленных соцветий до максимально редуцированных, состоящих всего из одного простого зонтика, как у женьшеня (табл. 41). Метельчатые соцветия аралиевых могут быть очень крупными.
У аралиевых поликарпиков в зависимости от темпов развития соцветия оно зацветает в одних случаях как терминальное (при быстром развитии), в других как боковое по положению. В последнем, нередком у аралиевых случае пробуждающуюся на верхушке побега цветочную почку перевершинивают одна (при симподиальном ветвлении ствола) или две (при ветвлении ложновильчатом) вегетативные ветви, и к моменту цветения соцветие соответственно представляется боковым или оказывается зажатым в развилке ствола. И уж совсем необычный вид имеют репродуктивные побеги таких примитивных аралиевых, как тупидантус, гоастония великолепная и некоторые шефлеры, развитие соцветия у которых растягивается на несколько лет. По длине побега у них можно видеть одновременно соцветия на всех этапах формирования от терминальной цветочной почки, ниже - развертывающегося соцветия в бутонах, еще ниже - вполне развернувшегося соцветия и до соцветия со зрелыми плодами (рис. 156).
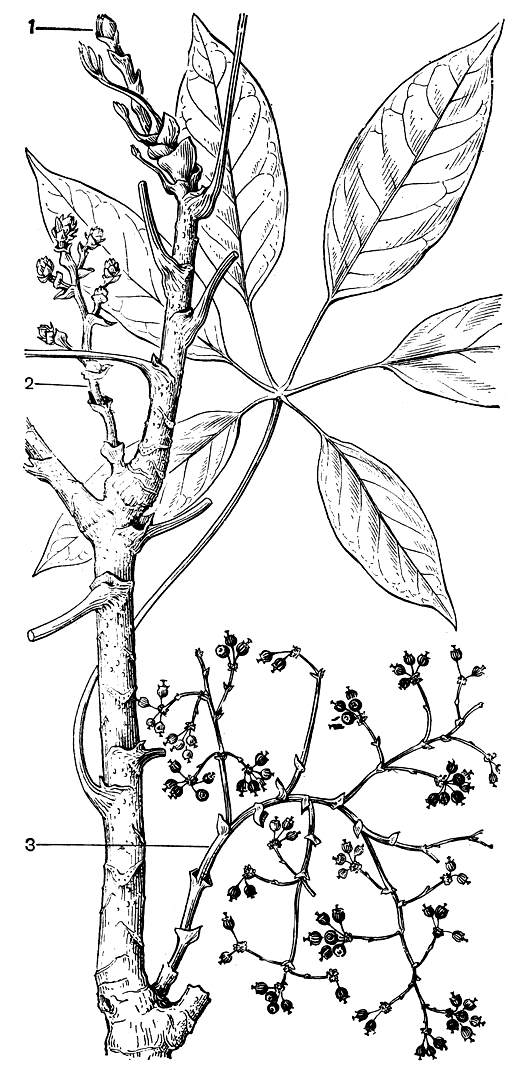
Рис. 156. Ветвь шефлеры многокосточковой (Schefflera polypyrena) с соцветиями на разных этапах развития: 1 - развертывающаяся терминальная цветочная почка; 2 - соцветие в бутонах; 3 - соцветие со зрелыми и опадающими плодами
Насколько крупными и заметными уже издали являются соцветия аралиевых, настолько мелки и невзрачны, как правило, их цветки. Если для примера рассмотреть их у наиболее известных представителей - плюща и женьшеня, то цветок аралиевых можно характеризовать как свободнолепестный, правильный, обоеполый, пятичленный, с мелкими зубчиками чашечки, с пятью неяркими широкими в основании лепестками, с пятью чередующимися с ними тычинками, наконец, с нижней пяти-(у плюща) или двугнездной (у женьшеня) завязью, увенчанной соответственно пятью или двумя столбиками, у основания которых, прикрывая завязь, расположен нектароносный диск. Такой цветок действительно наиболее характерен для аралиевых и по своему плану очень близок к цветку зонтичных. Однако в пределах рассматриваемого семейства наблюдаются значительные отличия от этого плана. Так, для наиболее примитивных аралиевых, объединяемых в трибу плерандровых (Plerandreae), а также некоторых видов шефлеры и других родов характерны полимерные цветки. Так, в цветках уже упомянутой выше гастонии великолепной 6-12 лепестков, 25-66 тычинок и 6 - 12-гнездная завязь.
У тупидантуса колпачкового число лепестков установить трудно, так как они, как и у ряда других плерандровых, срастаются в деревянистую калиптру, или колпачок, который целиком отпадает в момент зацветания. Но тычинок здесь от 90 до 160, а гнезд завязи даже до 200.
Интереснейшим исключением также являются цветки с верхней завязью, характерные для тетраплазандры голоплодной (Tetraplasandra gymnocarpa, рис. 157). У этого невысокого дерева гавайских тропических лесов, как показали обстоятельные исследования американских специалистов Р. Айда и Ч. Тсенга (1969), верхняя завязь произошла из нижней завязи предков, как бы в порядке "обратной эволюции".
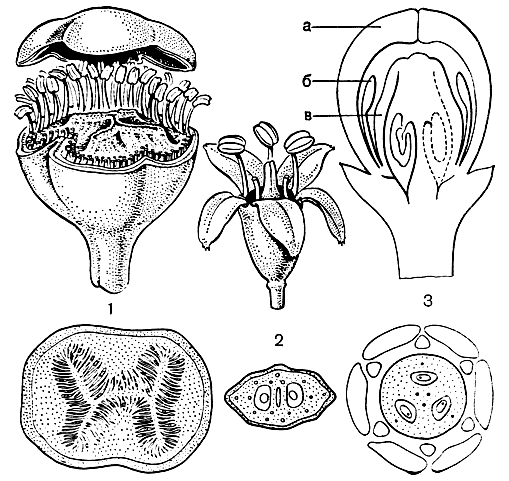
Рис. 157. Цветки аралиевых: 1 - тупидантус колпачковый (Tupidanthus calyptratus), полимерный цветок с нижней завязью; 2 - женьшень (Panax ginseng), пятичленный цветок с нижней завязью; 3 - тетраплазандра голоплодная (Tetraplasandra gymnocarpa), цветок с верхней завязью: а - венчик; б - тычинки; в - верхняя завязь. Сверху: 1-2 - общий вид цветка (спереди часть тычинок удалена); 3 - продольный срез цветка. Снизу: 1-2 - поперечный срез завязи; 3 - диаграмма цветка
У ряда представителей аралиевых - однодомных, полигамных и двудомных - имеют место однополые цветки, в других случаях лишь функционально однополые. Поразительные примеры разделения полов описали Филипсон (1970) у псевдопанакса дикого (Pseudopanax ferox) и Айди Тсенг (1971) у мериты Синклера (Meryta sinclairii). У этих двудомных растений женские соцветия одного типа строения, а мужские соцветия другого типа.
Аралиевые - растения энтомофильные. Цветки их доступны для посещения самыми различными насекомыми, которых привлекают и заметные издали соцветия, и распространяемый цветками аромат, и наличие выделяемого железистым диском нектара. Заросли и посадки плюща обыкновенного, прекрасного медоноса, дающего позднеосенний белый и очень плотный, так называемый "каменный" мед, в период цветения буквально гудят от множества пчел, но посетителями цветков являются одновременно и многие другие насекомые. В ходе многолетнего изучения цветения у элеутерококка колючего Э. А. Елумеев констатировал посещение цветков этого растения 27 видами насекомых, в том числе 16 видами из перепончатокрылых, 7 - из двукрылых, двумя - из чешуекрылых и по одному виду из сетчато - и жесткокрылых.
Приспособлением к перекрестному опылению у большинства аралиевых является протандрия. Имеются указания на опыление у некоторых видов шефлеры птицами.
Плод аралиевых - костянка, много -, пяти или двукосточковая, редко даже однокосточковая. Как правило, костянки сочные и распространителями этих плодов с их ярким (красным, оранжевым, желтым, синим, блестяще-черным, иногда пестрым) экзокарпием, с мясистым мезокарпием и твердым эндокарпием (косточкой) являются плодоядные птицы. Нельзя не упомянуть, однако, о плодах немногих аралиевых, проявляющих черты удивительного сходства с широкоизвестными сухими плодами (вислоплодниками) зонтичных. Черты этой общности (сухой перикарпий, распадение зрелых плодов на два мерикарпия, наличие в центре плода колонки - карпофора и т. д.) в той или иной степени присущи видам небольшого рода миодокарпус (Myodocarpus) из Новой Каледонии и названным выше видам стильбокарпы.
В каждой косточке плода аралиевых развивается, как правило, лишь одно семя. Его мощный эндосперм, гладкий у одних родов, руминированный - у других, является богатой кладовой запасных питательных веществ - белков и жиров. Напротив, зародыш у аралиевых очень маленький. Среди аралиевых нет видов широкого экономического значения. Однако некоторые представители зарекомендовали себя одни как эффектные декоративные растения, другие - как источники ценных лекарственных средств. Помимо плюща обыкновенного, культивируемого в десятках садовых форм не только в Европе, но и далеко за ее пределами, не только в открытом грунте, но и как комнатное растение, можно назвать ряд других популярных декоративных видов. Широко известна, в частности, фатсия японская (Fatsia japonica), иногда неправильно называемая аралией, а также гибрид между нею и плющом обыкновенным - фатсхедера Лизе (Fatshedera lizei). В тропических странах наряду с другими часты в культуре декоративные виды из рода полисциас, в частности полисциас кустарниковый (Polyscias iruticosa), листья которого издавна применяют и в народной медицине, и как ароматическую приправу к пище.
Не менее разнообразно применение южнокитайского кустарника тетрапанакса бумагоносного (Tetrapanax papyriferus), который давно и широко культивируется в Китае. Сильно развитая белая и губчатая сердцевина его стеблей дает материал для изготовления искусственных бумажных цветов, но также используется и в медицине в основном как средство лактогенное, усиливающее функцию молочных желез.
Необходимо, наконец, еще раз вернуться к женьшеню. Препараты из его корней, содержащих тритерпеновые гликозиды редкого в природе типа, как показали исследования советских фармакологов и врачей, обладают тонизирующим и стимулирующим действием. Женьшень оказался и первым растительным адаптогеном - источником средств, повышающих общую сопротивляемость организма к различным неблагоприятным воздействиям. Крайняя редкость женьшеня в природе, обусловленная многовековыми поисками этого "растения-клада", уже давно, около 600 лет назад, побудила к введению его в культуру в Корее, много позднее также в Японии и Китае. В нашей стране дикорастущий женьшень взят под охрану. Культивируется женьшень в специальном совхозе "Женьшень" на юге Приморского края, ведутся опыты по его выращиванию и в других районах страны. В последние годы в СССР обоснована возможность промышленного выращивания лекарственного сырья женьшеня в полностью искусственных условиях, методом культуры изолированных тканей. Наконец, весьма успешными оказались поиски заменителей женьшеня среди остальных дальневосточных аралиевых; применение в научной медицине получили препараты из подземных органов заманихи, аралии и особенно ценные из корней элеутерококка колючего, как считают, полноценно заменяющие женьшень.
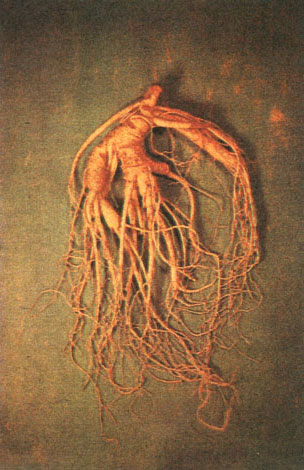
Таблица 41. Аралиевые: 3 - корень женьшеня
Семейство зонтичные (Apiaceae, или Umbelliferae) (Н. Н. Цвелев)
Семейство зонтичные принадлежит к числу наиболее крупных и наиболее важных в хозяйственном отношении семейств цветковых растений. Оно включает около 300 родов и 3000 видов, распространенных почти по всей суше земного шара. Однако наиболее многочисленны зонтичные в умереннотеплых и субтропических областях северного полушария, а в тропических странах приурочены главным образом к горным районам.
Среди зонтичных преобладают однолетние и многолетние травы разнообразного облика. Значительно реже встречаются полукустарники, а кустарники и древовидные жизненные формы встречаются лишь в немногих родах (мирридендрон - Myrrhidendron, гетероморфа - Heteromorpha, синеголовник - Eryngium, володушка - Bupleurum).
Стебли обычно прямостоячие, реже лежачие, часто с полыми междоузлиями, даже у травянистых форм нередко достигающие в высоту 3 м.
Однако среди зонтичных имеются и виды, образующие густые подушкообразные дерновины (азорелла - Azorella, рис. 158), а также карликовые бесстебельные или почти бесстебельные однолетники, например хохенакерия бесстебельная (Holienackeria exscapa), встречающаяся и в СССР в полупустынных районах Закавказья (рис. 159). В стеблях, как и во всех других органах зонтичных, имеются секреторные каналы, содержащие эфирные масла и смолистые вещества, определяющие их специфический запах.

Рис. 158. Зонтичные. Азорелла голая (Azorella glabra): 1 - общий вид растения; 2 - цветок с оберткой; 3 - плод с оберткой. Болот-нозонтичник ползучий (Helosciadium repens): 4 - общий вид растения; 5 - плод. Синеголовник ложноситниковый (Eryngium pseudojunceum): 6 - общий вид растения; 7 - головка; 8 - цветок. Арктопус колючий (Arctopus echinatus): 9 - общий вид особи с мужскими цветками; 10 - мужской цветок; 11 - зонтичек с женскими цветками; 12 - женский цветок
Листья зонтичных обычно очередные, без прилистников и разделены на сильно расчлененную (нередко с нитевидными долями) пластинку, черешок и охватывающее стебель влагалище. Однако исключений в этом отношении довольно много. Так, у многочисленных видов рода володушка имеются цельные и цельнокрайные листья без влагалищ и обычно с параллельным или дуговидным жилкованием. Их форма варьирует от узколинейной до сердцевидной и почти круглой, а у некоторых видов верхние листья могут быть даже пронзенными стеблем (см. рис. 159). Прежде предполагали, что листья володушки представляют собой пластинкообразно расширенные черешки, однако анатомоморфологические исследования не подтвердили: этого предположения. Полная редукция листовых пластинок до шиловидной средней жилки встречается среди зонтичных только у видов лилеопсис (Lilaeopsis) - стелющихся болотных растений, распространенных в Америке, Австралии и Новой Зеландии (рис. 160).
Цельные, но зубчатые по краю листовые пластинки встречаются во многих трибах зонтичных. У видов щитолистника (Hydrocotyle) они имеют почти округлую форму и сидят на длинных черешках, причем у единственного европейского вида этого рода - щитолистника обыкновенного (H. vulgaris)) - листья даже щитовидные (см. рис. 159). У этого рода, как и у других родов более примитивного подсемейства щитолистниковых (Hydrocotyloideae), при основании черешков имеются небольшие прилистники, отсутствующие у других зонтичных. Другое редкое исключение - супротивное расположение листьев - встречается у видов 2 небольших родов из Америки: бовлезии (Bowlesia) и спананте (Spananthe), а также у эндемика Канарских островов - друзы супротивнолистной (Drusa oppositifolia).

Рис. 159. Зонтичные. Володушка кустарниковая (Bupleurum fruticosum): 1 - цветущая ветвь; 2 - цветок; 3 - плод. Володушка Хельдрейха (В. heldreichii): 4 - верхушка стебля с соцветием; 5 - частная обертка; 6 - плод. Нираратамнус копытнелистный (Nirarathamnus asarifolius): 7 - общий вид растения; 8 - цветок. Хохенакерия бесстебельная (Hohenackeria exscapa): 9 - общий вид растения. Щитолистник обыкновенный (Hydrocotyle vulgaris): 10 - общий вид растения
Очень мелкие, густо расположенные, часто цельные и цельнокрайные листья имеют виды азореллы и некоторых других высокогорных зонтичных Южной Америки. Часто двурядно расположенные, по краю колючезубчатые, ланцетные или линейные листья некоторых видов синеголовника и австралийско-новозеландского рода ацифилла (Aciphylla) напоминают листья бромелиевых (Bromeliaceae) или некоторых агав (Agave). He менее оригинальны виды южноамериканского кампоса (саванно-подобной растительности) - синеголовник ситниковый (Eryngium junceum) и синеголовник пухоносный (E. eriophorum) с жесткими узколинейными листьями длиной до 80 см и шириной до 5 мм, а также синеголовник ложноситниковый (E. pseudojunceum) из Чили, имеющий полые узколинейные листья с поперечными перегородками, как у многих видов ситника (Juncus). Сильно мясистые листья имеются у нескольких видов синеголовника и у некоторых других, обычно галофильных родов, например у средиземноморского литорального рода критмум (Crithmum), известного в СССР в Крыму и на Кавказе.
Хотя у большинства зонтичных мелкие цветки собраны в очень характерные для этого семейства соцветия - сложные зонтики (табл. 42), многие роды различного родства имеют цветки в простых зонтиках, переходящих у видов рода подлесник (Sanicula) в головкообразные соцветия. Многочисленным видам синеголовника свойственны соцветия - плотные головки, в которых каждый цветок имеет у своего основания прицветник (брактею), а прицветники краевых цветков обычно более или менее увеличены, образуя обертку. У южноафриканской центеллы красивозубчатой (Centella calliodus) и западноавстралийской ксантозии (Xanthosia singuliflora) цветки одиночные вследствие редукции всех цветков простого зонтика до одного. Одиночные цветки встречаются и у многих видов азореллы, верхушечные простые зонтики которых обычно состоят из 2-5 цветков. Оригинальные верхушечные соцветия в форме дихазиев имеет эндемик Сицилии - петагния подлесниколистная (Petagnia saniculifolia). В развилках вильчато разветвленного соцветия и на концах его ветвей располагаются простые зонтики, состоящие из одного срединного женского цветка и 1-3 мужских цветков на длинных ножках, срастающихся в своей нижней части с жилками чашечки женского цветка (см. рис. 160).

Таблица 42. Зонтичные: 3 - реброплодник уральский (Pleurospermum uralense), Камчатка
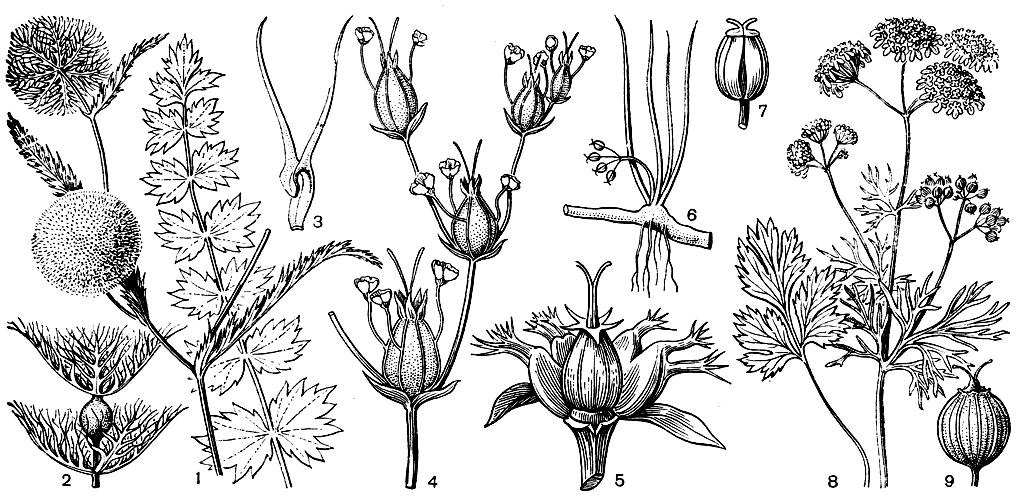
Рис. 160. Зонтичные. Лагеция кминовидная (Lagoecia cuminoides): 1 - общий вид растения; 2 - одноцветковый зонтичек; 3 - лепесток. Петагния подлесниколистная (Petagnia saniculifolia): 4 - часть соцветия. Колюченосник колючий (Echinophora spinosa): 5 - плодоносящий зонтичек с удаленными спереди тычиночными цветками. Лилеопсис линейный (Lilaeopsis lineata): 6 - общий вид растения; 7 - плод. Кориандр посевной (Coriandrum sativum): 8 - общий вид растения; 9 - плод
У основания простых зонтиков и вторичных зонтиков (или зонтичков) сложного зонтика часто имеются небольшие листочки, образующие частную обертку (или оберточку) и являющиеся прицветниками наружных цветков. Внутренние цветки, как правило, лишены прицветников. Если у основания первичных лучей сложного зонтика имеются верхушечные видоизмененные листья, то они образуют общую обертку сложного зонтика или просто обертку. Листочки оберток обычно бывают цельными и цельнокрайними, однако у некоторых родов, например у моркови (Daucus), они могут быть перисторассеченными. Листочки частных оберток и оберток простых зонтиков могут различным образом срастаться и быть ярко окрашенными, играя в этом случае некоторую роль в привлечении насекомых-опылителей. Например, у видов астранции (Astrantia, табл. 42) сильно увеличенные листочки обертки окрашены в розовый цвет, а у володушки золотистой (Bupleurum aureum) - в ярко-желтый. У володушки Хельдрейха (B. heldreichii) сильно увеличены и срастаются друг с другом только 3 наружных листочка обертки из 5 (см. рис. 159). Обычно же листочки обертки и оберточки служат средством защиты бутонов от внешних воздействий и у некоторых видов после распускания цветков опадают.

Таблица 42. Зонтичные: 2 - астранция трёхнадрезная (Astrantia trifida), Кавказ
Простые или сложные зонтики могут располагаться по одному, заканчивая собой стебель или выходя из пазух листьев стелющегося побега, верхушка которого остается вегетативной (например, у щитолистника, см. рис. 159). Однако чаще зонтики образуют более или менее разветвленное общее соцветие, в котором центральный зонтик является наиболее развитым. У некоторых зонтичных центральный зонтик несет обоеполые или женские цветки, а зонтики боковых ветвей - мужские цветки с более или менее редуцированным гинецеем. Однополые цветки вообще не редки среди зонтичных, причем они могут быть как однодомные, так и двудомные. Примером двудомных зонтичных могут служить южноафриканский род арктопус (Arctopus, см. рис. 158) и виды рода триния (Trinia), обитающие в степях Евразии. Однополые, но однодомные цветки имеют виды древнесредиземноморского рода колюченосник (Echinophora). Зонтики сложных зонтиков состоят у них из сидячего центрального женского цветка, окруженного более мелкими мужскими цветками на толстых, сильно отвердевающих при плодах ножках (см. рис. 160). Цветки большинства зонтичных построены относительно однообразно. Обычно они актиноморфные и 5-членные, однако у видов многих родов, особенно орлайи (Orlaya) и лизеи (Lisaea), наружные лепестки краевых цветков сложных зонтиков могут быть более или менее увеличенными. Зубцы чашечки обычно слаборазвиты и мало заметны, но имеются и исключения в этом отношении. Так, у западноавстралийского рода схенолена (Schoenolaena) зубцы чашечки очень крупные, сердцевидные, но быстро опадающие. У средиземноморского однолетника лагеции кминовидной (Lagoecia cuminoides) они еще более крупные, перисто-рассеченные, играют при плодах роль хохолка или летучки. Зонтички лагеции одноцветковые, но с частной оберткой из перисторассеченных листочков, а составленные из таких зонтичков сложные зонтики имеют почти шаровидную форму, что придает всему растению очень оригинальный облик (см. рис. 160). Лепестки обычно белые, желтые или желтовато-зеленые, редко синеватые или розоватые. У более примитивного подсемейства щитолистниковые форма их варьирует от яйцевидной до ланцетной, однако у большинства зонтичных лепестки построены более сложно: из узкого основания они расширяются в пластинку, затем вновь суживаются и суженной частью загибаются внутрь цветка, производя впечатление двух- или трехлопастных. У лагеции боковые части лепестков, кроме того, оттянуты в длинные рожки. Лепестки австралийского рода актинотус (Actinotus) редуцированы до нитевидных образований, напоминающих стаминодии. Тычинок в цветке зонтичных всегда 5, чередующихся с лепестками, прикрепленных к железистому диску, и в молодости загнутых внутрь цветка. Гинецей синкарпный и состоит из 2 плодолистиков. Нижняя двугнездная завязь на верхушке переходит в подстолбие (или стилоподий), часто называемое еще железистым или нектарным диском, так как на нем выделяется нектар. От железистого диска, который может иметь различную форму (у актинотуса он редуцирован до 2 небольших желёзок), отходят 2 столбика, заканчивающиеся притуплённым, реже головчатым или заостренным рыльцем. Обычно в каждом гнезде находятся 1-2 семязачатка, из которых только один развивается в семя. У лагеции, петагнии, колюченосника и некоторых других родов плод становится односемянным вследствие редукции одного из гнезд.
Значительное большинство зонтичных имеет сухой плод - вислоплодник. При созревании он обычно распадается на 2 половинки - мерикарпии, которые некоторое время висят на вильчато разветвленной колонке, образованной брюшной (вентральной) частью плодолистиков и называемой столбочком или карпофором. Карпофор отсутствует в пределах более архаичных подсемейств щитолистниковых и подлесниковых (Saniculoideae), однако и среди подсемейства сельдереевых (Apioideae) он не всегда заметен, не отчленяясь от мерикарпиев (например, у кориандра - Coriandrum) или оставаясь заключенным внутри нераспадающегося плода (например, у омежника - Оеnаnthe).
Строению плодов придается особенно большое значение в систематике зонтичных, в связи с чем разработана специальная терминология. Брюшную сторону мерикарпиев называют спайкой или комиссурой. На выпуклой спинной стороне имеются 5 главных или первичных продольных ребер, образованных проводящими пучками с окружающей их тканью. В промежутках между ними, называемых ложбинками, могут находиться вторичные ребра. В среднем слое околоплодника - мезокарпии - имеются продольные масляные каналы, обычно располагающиеся под ложбинками и со стороны комиссуры. Детали строения плодов могут очень варьировать в связи с приспособлением к распространению различными агентами. В частности, ребра могут быть крыловидными или нести различные придатки в виде шипиков, чешуек, бугорков и так далее (рис. 161).
Зонтичные обычно не занимают господствующего положения в растительных группировках, но нередко играют в них видную роль. Следует особенно отметить большое значение видов ферулы (Ferula, табл. 42), доремы (Dorema), прангоса (Prangos) в низкогорных ландшафтах Средней и Западной Азии. Среди растительности приречных лугов Камчатки выделяются такие гигантские зонтичные, как дудник медвежий (Angelica ursina) и борщевик сладкий (Heracleum dulce, табл. 42). В травянистом ярусе лиственных и смешанных лесов в качестве эдификаторов нередко выступают виды сныти (Aegopodium), подлесника, купыря (Anthriscus) и других родов зонтичных. Виды из родов поручейник (Sium), омежник, вех (Cicuta) нередко в изобилии встречаются по берегам водоемов и на болотах. Для степей и открытых песчаных мест не менее характерны многие виды родов горичник (Peucedanum), жабрица (Seseli), бедренец (Pimpinella) и др. Древовидные жизненные формы высотой до 5 м известны среди зонтичных только в роде мирридендрон, 5 видов которого встречаются на горных склонах Центральной и Южной Америки. Они имеют очень крупные, трижды перистораздельные листья и сложные зонтики белых цветков. Среди немногих кустарниковых зонтичных в высоту 5-6 м достигает восточно- и южноафриканская гетероморфа древовидная (Heteromorplm arborescens) с перистыми, тройчатыми или цельными кожистыми листьями. Не превышают высоту 2-2,5 м кустарники из родов синеголовник (Eryngium): синеголовник володушковидный (E. bupleuroides) и синеголовник толстолистный (E. sarcophyllum) - эндемики острова Хуан-Фернандес - и володушка (Bupleurum): володушка седоватая (B. canescens) и володушка кустарниковая (B. fruticosum) из Средиземноморья. Последний из этих видов, имеющий цельные и цельнокрайние кожистые листья, встречается в качестве одичавшего растения на Южном берегу Крыма (см. рис. 159).

Таблица 42. Зонтичные: 1 - борщевик сладкий (Heracleum dulce), о. Карагинский

Таблица 42. Зонтичные: 5 - ферула Иешке (Ferula jaeschkeana), Дарваз
Из более многочисленных в семействе полукустарников наиболее оригинален эндемик острова Сокотра нираратамнос копытнелистный (Nirarathamnos asarifolius) с почти округлыми кожистыми цельными цельнокрайними пластинками листьев (см. рис. 159).

Таблица 42. Зонтичные: 4 - нираратамнос копытнелистный (Nirarathamnos asarifolius), остров Сокотра
Среди травянистых зонтичных особенно много однолетников, двулетников и многолетников с прямостоячим стеблем и стержневым корнем. Многолетники представлены как поликарпиками, цветущими и плодоносящими в течение целого ряда лет, так и монокарпиками, после плодоношения полностью отмирающими. Обычно в течение 3-15 лет многолетники-монокарпики образуют лишь розетки прикорневых листьев, накапливая питательные вещества в корне, а затем дают мощный цветущий стебель. К ним. в частности, принадлежат многие виды рода ферула, распространенные в полупустынях и низкогорьях Средней и Западной Азии. Их высокие толстые стебли с сильно разветвленным соцветием часто несут лишь значительно редуцированные влагалищеобразные листья, у некоторых видов (например, у ферулы яйценогой - F. oopoda) имеющие своеобразную чашевидную форму. Еще более замечательный облик имеют цветущие экземпляры видов небольшого средне - и западноазиатского рода дорема. Их стебли, нередко достигающие в высоту 2-3 м, также несут лишь редуцированные листья и очень крупные соцветия, на ветвях которого располагаются многочисленные простые зонтики густо расположенных желтых, реже зеленовато-желтых цветков.
Очень больших размеров (высотой до 3 м) могут достигать и стебли луговых и лесных зонтичных из родов дудник и борщевик, многие из которых также являются многолетниками-монокарпиками.
Примером многолетнего зонтичного с ползучим корневищем может служить широко распространенная в лесах СССР сныть обыкновенная (A. podagraria). Многие болотные и прибрежные зонтичные имеют стелющиеся и укореняющиеся в узлах стебли. Таковы, например, европейские виды щитолистник обыкновенный и болотнозонтичник ползучий (Helosciadium repens, см. рис. 158).
Очень густые и плотные подушкообразные дерновины в поперечнике до 4 м образуют многочисленные виды азореллы, распространенные в высокогорьях южноамериканских Анд и Антарктике. Особенно богато ими Чили, где встречается около 30 видов этого замечательного рода, внешне очень похожего на подушкообразующие камнеломки северного полушария (см. рис. 155). Азорелла селаго (A. selago) является одним из немногих видов цветковых растений антарктического острова Кергелен, где она покрывает значительную часть его суши. Подушкообразные дерновины азореллы очень богаты эфирными маслами и смолами и поэтому широко используются жителями высокогорных пустынь Анд в качестве отличного горючего материала.
Многие зонтичные накапливают питательные вещества в клубнях. Чаще встречаются одиночные клубни, располагающиеся или близ поверхности земли (например, у двулетника бутеня Прескотта - Chaerophyllum prescottii), или глубоко под землей (например, у видов рода буниум - Bunium). Пучок клубневидно утолщенных корней имеют некоторые виды омежника.
К сильно колючим растениям принадлежат многие виды синеголовника, имеющие колючки по краям листьев и на листочках обертки. Южноевропейский вид синеголовник полевой (E. campestre) отлично защищен колючками от поедания скотом и нередко в изобилии растет даже на самых выбитых выгонах. Колючие листочки обертки и колючки на поверхности листьев имеют виды оригинального южноафриканского рода арктопус (Arctopus, см. рис. 158), а многочисленные колючки стеблевого происхождения - западносредиземноморская володушка колючая (B. spinosum).
Выделяемый железистым диском широко раскрытых цветков зонтичных нектар доступен самым разнообразным насекомым-опылителям, в особенности короткохоботковым мухам, средством привлечения которых нередко является и специфический запах многих зонтичных. Однако самоопыление среди зонтичных случается лишь в очень редких случаях, так как ему препятствует очень строгая протандрия. Столбики в обоеполых цветках зонтичных, как правило, удлиняются и расходятся в стороны только после отмирания тычинок. А. Н. Пономаревым отмечено, что даже у видов с сильно разветвленным соцветием, состоящим из многих зонтиков, женская фаза цветения четко отграничена по времени от мужской. Так, у лугово-степного горичника эльзасского (Peucedanum alsaticum) все зонтики очень крупного общего соцветия зацветают почти одновременно и в течение 8-12 дней цветут в мужской фазе, а затем одновременно во всех зонтиках наступает переход к женской фазе, которая длится еще 6-7 дней. Одновременный переход к женской фазе цветения имеет место даже у разновременно расцветающих зонтиков одного и того же растения, причем мужская фаза ранее расцветших зонтиков длится гораздо дольше, чем у поздно зацветших. Как уже отмечалось, дополнительными средствами привлечения опылителей, кроме объединения мелких цветков в соцветия и запаха, могут быть увеличенные и ярко окрашенные листочки оберток и увеличенные краевые лепестки цветков.
Распространение диаспор, которыми у зонтичных обычно являются мерикарпии, обычно осуществляется с помощью ветра, водных потоков, животных и человека. Пожалуй, чаще всего встречается анемохория, причем парусность мерикарпиев может увеличиваться как за счет образования крыловидных продольных ребер, так и за счет развития очень легкой, пробкообразной ткани (например, у прангоса), что характерно также для мерикарпиев многих зонтичных, распространяющихся водными потоками (например, видов омежника). Обитающий в песчаных пустынях Средней Азии криптодискус песколюбивый (Cryptodiscus ammophilus) имеет густоволосистые мерикарпии, что еще больше увеличивает их парусность. Легко распространяются ветром также мелкие, покрытые чешуйками плоды многих видов синеголовника. Целый ряд степных и полупустынных видов зонтичных, в том числе синеголовник полевой и жабрица извилистая (Seseli tortuosum), имеют сильно и растопыренно разветвленные от основания стебли и распространяют плоды по типу перекати-поля.
Многие виды зонтичных распространяются экзозоохорно - на шерсти животных и одежде человека. Их мерикарпии обычно снабжены крючковидными шипами или щетинками (например, у торилиса - Torilis и прицепника - Caucalis). На ногах человека и животных легко разносятся с комочками почвы мерикарпии многих видов, не имеющих специальных приспособлений для распространения (например, тмина - Carum carvi). Редкий у зонтичных баллистический способ распространения известен у средиземноморского рода скандикс (Scandix), имеющего очень крупные (длиной до 7-8 см) плоды с длинным и тонким носиком (см. рис. 161). Когда зрелые плоды распадаются, образующая носик часть мерикарпия скручивается и отбрасывает весь мерикарпии в сторону. Относительно немногие зонтичные легко размножаются вегетативно: с помощью ползучих корневищ, корневых отпрысков, укореняющихся надземных побегов. Североамериканский вех луковичконосный (Cicuta bulbifera) несет пучки опадающих луковичек в пазухах верхних стеблевых листьев.
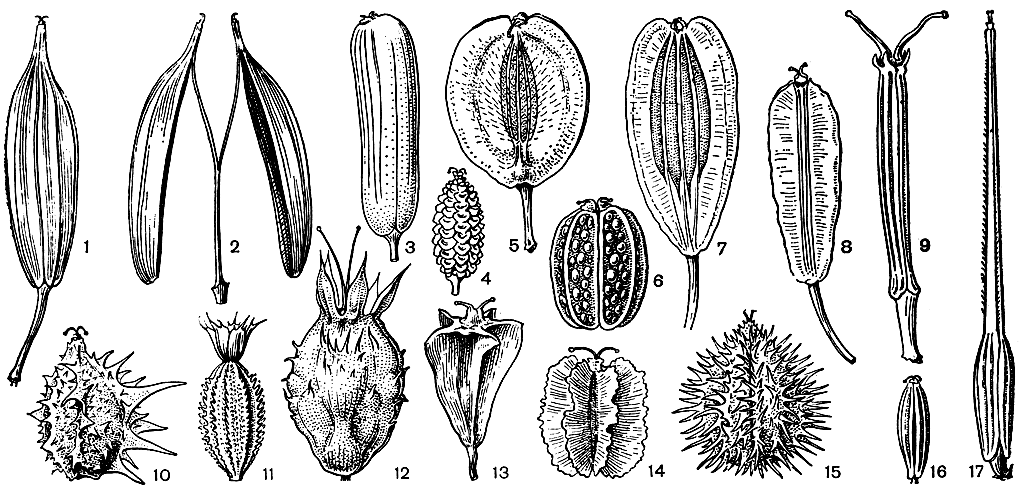
Рис. 161. Плоды зонтичных. 1, 2 - миррис душистая (Myrrhis odorata); 3 - прангос противозубный (Prangos odontalgica); 4 - совичия красивоплодная (Szovitsia callicarpa); 5 - зосима восточная (Zosima orientalis); 6 - смирниопсис Оше (Smyrniopsis aucheri); 7 - ферула гигантская (Ferula gigantea); 8 - граммосциадиум шероховатый (Grammosciadium scabridum); 9 - граммосциадиум крылоплодный (G. pterocarpum); 10 - лизея разноплодная (Lisaea lieterocarpa); 11 - актинолема крупнообертковая (Actinolema macrolema); 12 - синеголовник приморский (Eryngium maritimum); 13 - пирамидоптера кагульская (Pyramidoptera cagulica); 14 - гладыш щетинисто-волосистый (Laserpitium hispidum); 15 - тургеневия широколистная (Turgenia latifolia); 16 - тмин обыкновенный (Carura carvi); 17 - скандикс гребенчатый (Scandix pecten-veneris)
Семейство зонтичных обычно делят на 3 подсемейства и целый ряд триб, основываясь главным образом на строении плодов. Наиболее приближающееся к аралиевым и иногда выделяемое в особое семейство подсемейство щитолистниковых характеризуется костянкообразным плодом с деревянистым внутренним слоем околоплодника - эндокарпием, отсутствием свободного карпофора и масляных каналов (или последние помещаются в главных ребрах). Из 2 триб подсемейства собственно щитолистниковые (Hydrocotyleae) имеют сплюснутые с боков, а мулиновые (Mulineae) уплощенные или широко закругленные на спинке мерикарпии. 30 родов и около 400 видов подсемейства распространено преимущественно в южном полушарии, многие в горах тропиков, нередко играя существенную роль в их растительном покрове (например, виды азореллы).
Представители подсемейства подлесниковых, включающего целый ряд своеобразнейших по своему облику родов зонтичных (синеголовник, астранцию, лагецию, петагнию, арктопус и другие), имеют мягкий, паренхимный эндокарпий плодов, но, в отличие от сельдереевых, железистый диск у них в виде кольца, окружающего столбики с головчатыми рыльцами, и отсутствует свободный карпофор. Кроме того, цветки видов этого подсемейства собраны простыми зонтиками или головками, а не сложными зонтиками. Подлесниковые также делятся на 2 трибы: собственно подлесниковые (Saniculeae) с двугнезднои завязью и развитыми масляными каналами и лагециевые (Lagoecieae) с одногнездной завязью и неразвитыми масляными каналами. Из 9 родов и около 300 видов подлесниковых многие заходят в горные районы тропиков и Южную Африку.
Наконец, последнее подсемейство - собственно зонтичные или сельдереевые, представленное многочисленными родами и видами почти исключительно во внетропических странах северного полушария, при мягком эндокарпии плодов имеет не кольцевидный и обычно выпуклый железистый диск со столбиками, обычно заканчивающимися притуплёнными рыльцами, почти всегда хорошо развитый карпофор и хорошо развитые масляные каналы. Также на основании различий в строении плодов подсемейство делится на 8 триб: колюченосниковые (Echinophoreae), скандиксовые (Scandiceae), кориандровые (Coriandreae), смирниевые (Smyrnieae), аммиевые (Ammieae), горичниковые (Peucedaneae), гладышевые (Laserpitieae) и морковные (Dauceae).
Многие зонтичные имеют большое хозяйственное значение в качестве пищевых (преимущественно овощных и пряных), лекарственных, кормовых и технических растений. Среди них морковь посевная (Daucus sativus) является одной из основных овощных культур. Корнеплоды моркови очень богаты витаминами (особенно провитамином А - каротином) и содержат эфирное масло, придающее им специфический морковный запах. Морковь имеет также большое значение в диетическом питании, при лечении авитаминозов, малокровия. В культуру она введена около 4000 лет назад в странах Средиземноморья сначала как лекарственное, а затем как пищевое и кормовое растение.
Широко культивируют в качестве овощных и пряных растений также петрушку (Petroselium crispum) и сельдерей (Apium graveolens), у которых в пищу используют не только корнеплоды, но и листья, особенно богатые витамином С. Оба вида содержат специфические эфирные масла, которых особенно много в семенах. Широко известным в СССР пищевым и лекарственным растением является также укроп (Anethum graveolens). В пищу употребляют его листья, богатые витамином С и другими витаминами. Используют его также как ароматическую приправу и пряность при консервировании овощей. Отвары и настои плодов употребляют в медицине для улучшения аппетита и как успокаивающее средство. К числу основных эфирномасличных растений СССР принадлежит кориандр (Coriandrum sativum), плоды которого содержат 0,2-1,4% эфирного масла и до 28% жирного масла (см. рис. 160). Листья прикорневых розеток и плоды кориандра, имеющие сильный аромат и острый вкус под названием киндза или кинза, широко используют в пищу в качестве пряности в Средней Азии и Закавказье, где этот вид культивируют с древнейших времен. В Древнем Египте кориандр культивировали более чем за 1000 лет до н. э. Ценными эфирномасличными культурами являются также тмин и анис (Anisum vulgare), плоды, корни и листья которых используют в качестве пряности, а эфирные масла применяют в медицине и парфюмерии. Из менее известных в СССР овощных зонтичных следует упомянуть еще любисток (Levisticum officinale), пастернак (Pastinaca savita), фенхель (Foeniculum vulgare), купырь кервель (Anthriscus cerefolium). В Центральной и Южной Америке с древнейших времен культивируют в качестве овоща арракачу (Arracacia xanthorhiza), дающую корнеплоды толщиной до 20 см.
Многие дикорастущие зонтичные (например, сныть) можно использовать для приготовления салатов. В качестве пряных и эфирномасличных растений культивируют еще такие виды зонтичных, как айован (Trachyspermum ammi), амми (Ammi visnaga), миррис (Myrrhis odorata), кмин (Cuminum cyminum).
Из лекарственных зонтичных можно отметить еще центеллу азиатскую (Centella asiatica). издавна используемую в Южной Азии в качестве средства против проказы, слоновой болезни и кожных заболеваний, а также некоторые виды ферулы (особенно "персидский корень" - Ferula assafoetida), из отвердевшего сока которых готовят ценные лекарственные препараты. Некоторые гигантские виды борщевика введены в культуру в качестве кормовых и декоративных растений. Декоративное значение имеют также некоторые виды астранции и володушки, пригодные для культуры в парках и альпинариях, и некоторые виды синеголовника, сохраняющие в сухом состоянии свою окраску.- для сухих букетов.
Нельзя в заключение не упомянуть о таких широко известных ядовитых зонтичных, как вех, или цикута (Cicuta virosa), болиголов (Conium maculatum) и собачья петрушка (Aethusa cynapium), содержащих много алкалоидов (алкалоид болиголова - кониин - был первым искусственно синтезированным алкалоидом). В древних Афинах яд болиголова использовали как орудие казни. В частности, именно им, вероятно, был отравлен один из известных философов древности - Сократ.
Канапе и фуршетные закуски доставка www.furshetnedorogo.ru.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'