Порядок камнеломковые (Saxifragales)
Семейство брунеллиевые (Brunelliaceae) (А. Е. Бородина)
Это тропическое американское семейство содержит единственный род брунеллия (Brunellia) с 50 видами, которые встречаются во влажных лесах гор Южной Мексики, Центральной Америки, Вест-Индии и в Андах Южной Америки от Венесуэлы до Боливии на высотах 1800-3000 м.
Брунеллиевые - невысокие или иногда довольно крупные, высотой до 20 м, вечнозеленые деревья с цилиндрическими, ветвящимися наверху стволами с серой часто трещиноватой корой. Листья довольно крупные, супротивные или мутовчатые, простые, трехлисточковые или непарноперистые, с маленькими опадающими прилистниками. Листья и молодые стебли обычно опушены короткими рыжеватыми, толстостенными одноклеточными волосками. Мелкие, невзрачные, актиноморфные, двудомные, безлепестные цветки собраны в пазушных или верхушечных метелках. Чашелистиков 4-5, густо опушенных. Тычинок 8-10, они прикреплены к основанию 8-10-лопастного нектарного диска, который, так же как и тычиночные нити, густо покрыт рыжими волосками. Гинецей апокарпный, состоит из 4-5 плодолистиков с вытянутыми шиловидными столбиками. Хотя цветки и однополые, однако в женских цветках имеются рудименты тычинок, а в мужских цветках - рудимент гинецея. Плод - 4-5-листовка. Интересной особенностью плода является расщепление его стенки при созревании: тонкий эндокарпий сжимается и отделяется от кожистого экзокарпия; шаровидные семена при этом выталкиваются из плода, но остаются прикрепленными на извилистой ножке, образованной фуникулусом и плацентой. Блестящие коричневые или красноватые семена, имеющие большой зародыш и обильный эндосперм, привлекают птиц, которые охотно поедают их.
Семейство Кунониевые (Cunoniaceae) (Н. Н. Цвелев)
Относительно небольшое (27 родов и около 350 видов) семейство кунониевых почти полностью распространено в южном полушарии. Лишь немногие из многочисленных (около 190) видов рода вейнманния (Weinmannia) распространены к северу от экватора и доходят до Филиппин, Южной Мексики и Антильских островов. На материке Евразия кунониевые полностью отсутствуют, а в Африке, не считая Мадагаскара, только в Капском флористическом царстве встречаются эндемичный монотипный род платилофус (Platylophus) и кунония капская (Cunonia capensis).
Кунониевые - деревья высотой до 30 м или кустарники с кожистыми простыми или сложными (тройчатыми или непарноперистыми) листьями, обычно расположенными супротивно, реже мутовчато. У рода бауэра (Ваиега) и некоторых других родов супротивные тройчатые листья не имеют черешков и их листочки производят впечатление мутовчато расположенных простых листьев. Очень характерно для всего семейства присутствие опадающих прилистников различной формы, часто срастающихся попарно с прилистниками противоположного листа. Лишь у бауэры прилистники отсутствуют, а у афанопеталума (Aphanopetalum) имеются лишь их рудименты.
Очень мелкие, обычно обоеполые цветки кунониевых собраны в кистевидные или метельчатые соцветия; реже образуют густые шаровидные головки (у родов трибы панхериевых - Pancherieae). Исключением здесь снова является бауэра, относительно крупные цветки которой расположены по одному в пазухах листьев. Несмотря на невзрачность и частое отсутствие лепестков, цветки кунониевых обычно имеют нектарники - придатки блюдцевидного околопестичного диска - и опыляются насекомыми с короткими хоботками, или жвалами. К двудомным растениям принадлежат все виды новокаледонского рода панхерия (Pancheria) и некоторые виды кодии (Codia). He полностью двудомными являются многие виды спиреантемума (Spiraeanthemum), у которых женские цветки нередко имеют фертильные тычинки.
Тычинок в цветках кунониевых обычно 4-5, как и чашелистиков, или вдвое больше, редко (у родов трибы беланжеровых - Belangereae) их более 20. Гинецей большинства кунониевых состоит из 2 в разной степени сросшихся плодолистиков со свободными столбиками. Из него образуется многосемянная коробочка, раскрывающаяся 2 створками. Однако у родов более примитивной трибы спиреантемовых (Spiraeanthemeae) плодолистиков (2)3-5, образующих синкарпный или апокарпный плод. Имеются и нераскрывающиеся односемянные плоды. У кодии, афанопеталума и цератопеталума (Ceratopetalum) они ореховидные и опадающие вместе с чашечкой, способствующей распространению плодов с помощью ветра и водных потоков. Виды австралийско-новогвинейского рода схизомерия (Schizomeria) имеют относительно крупные, мясистые, костянкообразные плоды, распространяющиеся преимущественно птицами. У большинства же кунониевых семена просто высыпаются из раскрывшихся коробочек и разносятся ветром на небольшие расстояния, чему способствует присутствие на них волосков или крыловидных выростов.
Кунониевые делятся на 2 подсемейства: монотипное - бауэровые (Baueroideae) и собственно кунониевые (Cunonioideae), которое, в свою очередь, делится на 5 триб. От наиболее богатой родами и видами трибы кунониевых (Cunonieae) другие трибы отличаются характерными для них особенностями: беланжеровые - многочисленными пыльниками, спиреантемовые - гинецеем из 3-5 плодолистиков, панхериевые - цветками в шаровидных головках, пуллеевые (Pulleae) - полунижней завязью.
Экологически большинство видов кунониевых приурочено к нагорным лесам тропиков и субтропиков. Все они влаголюбивы и часто обитают в поясе туманов. В Новой Зеландии и Чили некоторые кустарниковые виды поднимаются в горах за пределы субтропиков и переносят кратковременные заморозки.
Многие кунониевые дают ценную древесину. Так, древесина кунонии капской известна в Южной Африке под названием "красного железного дерева" благодаря красивому цвету и высокой прочности. Легкая, прочная и имеющая приятный запах древесина восточно-австралийского цератопеталума безлепестного (Ceratopetalum apetalum) используется в вагоностроительной промышленности и для производства мебели. Кора многих кунониевых содержит ценные дубильные вещества, а также используется в народной медицине в качестве вяжущего или тонизирующего средства. Кора вейнманнии красильной (Weinmannia tinctoria) дает хорошую краску для кожи. Наконец, некоторые кунониевые, например австралийская калликома пилъчатолистная (Gallicoma serratifolia), имеют декоративное значение и культивируются в ботанических садах и оранжереях.
Семейство эукрифиевые (Eucryphiaceae) (А. Е. Бородина)
Единственный род эукрифия (Eucryphia) встречается по обе стороны южной части Тихого океана: 2 вида - эукрифия сердцевиднолистная (E. cordifolia, рис. 76) и эукрифия клейкая (E. glutinosa) растут в горных лесах Южного Чили, а 3 вида - в горах Тасмании и юго-восточной части Австралии (карта 8). Обычно это высокие, стройные и красивые деревья. Наибольшей высоты (30-40 м) достигают чилийские эукрифии. Реже встречаются кустарники - некоторые австралийские эукрифии. Ветве- и листорасположение супротивное, листья плотнокожистые, вечнозеленые. В поверхностных тканях молодых побегов находятся многочисленные друзы, в большом количестве выделяющие красную воскообразную ароматную смолу; вследствие этого листья и молодые стебли блестят, а на поверхности некоторых австралийских растений образуются довольно большие скопления смолы. В марте эукрифии обильно зацветают. Крупные, диаметром до 5 см, красивые и ароматные цветки распускаются по одному в пазухах листьев. Цветок состоит из 4 кожистых сцепленных верхушками и опадающих в виде колпачка чашелистиков, обычно из 4 широких черепитчатых белых лепестков, множества тычинок, многорядно расположенных на удлиненной конусообразной оси цветка и вторичносинкарпного 4-14-плодолистикового гинецея. Благодаря 4 лепесткам и множеству тычинок, цветок внешне напоминает цветок зверобоя. Цветки посещают насекомые, производящие опыление. Плод эукрифии - кожистая или деревянистая коробочка, при созревании расщепляющаяся на лодочковидные створки, которые затем отделяются от оси плода и раскрываются вдоль шва, обнажая одно или несколько продолговатых семян с большим зародышем и обильным эндоспермом.
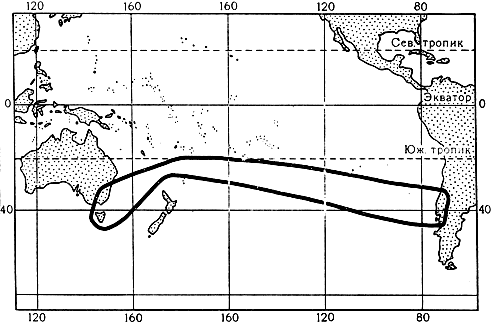
Карта 8. Ареал рода эукрифия
Эукрифии очень декоративны, их нередко культивируют в парках Великобритании и США (особенно чилийские виды). Крепкую, твердую и очень плотную древесину чилийских эукрифии используют для строительства домов, изготовления мебели, железнодорожных шпал, телеграфных столбов и повозок. Кора эукрифии служит для получения дубителей, цветы являются источником превосходного меда.

Рис. 76. Эукрифия сердцевиднолистная (Eucryphia cordifolia): 1 - ветвь с цветками; 2 - цветок с удаленными околоцветником и тычинками (а - зона прикрепления тычинок, б - зона гинецея и стилодия); 3 - поперечный разрез завязи; 4 - часть раскрывшегося плода; 5 - семя
Семейство эскаллониевые (Escalloniaceae) (Н. Н. Цвелев)
Семейство эскаллониевые отличается большим разнообразием входящих в него родов, которые распределяются по 6 подсемействам, иногда принимаемым за самостоятельные семейства. Из принадлежащих к нему 16 родов и около 200 видов значительное большинство распространено в тропических и субтропических областях южного полушария. В северных субтропиках встречается лишь обособленный род итеа (Itea), имеющий очень характерный для многих третичных реликтов разрыв ареала между Восточной Азией и Приатлантической Северной Америкой.
Эскаллониевые - преимущественно кустарники или деревья высотой до 20 м, очень редко полукустарники или лианы. В отличие от близкого семейства кунониевых их кожистые цельные листья обычно расположены очередно (редко супротивно или мутовчато) и, как правило, лишены прилистников. Очень мелкие быстро опадающие прилистники имеются только уродов брексия (Brexia) и филлонома (Phyllonoma, рис. 77). Последний род, виды которого распространены в горах Мексики и Южной Америки, замечателен расположением цветков (одиночных или собранных в кисти) не на ветвях, а на средних жилках верхней стороны листьев, обычно выше их середины, напоминая цветки иглицы (Ruscus), расположенные на листовидных стеблях - филлокладиях. Однако у филлономы листья настоящие, и такое расположение цветков объясняется срастанием цветоножек и ножек соцветий со средней жилкой листа. У большинства же других родов семейства мелкие или средней величины цветки собраны в верхушечные или пазушные кистевидные или метелковидные соцветия. Очень редко цветки располагаются по одному в пазухах листьев, а у оригинального стелющегося кустарника Огненной Земли и Южного Чили трибелеса южного (Tribeles australis) одиночные белые цветки заканчивают собою густо облиственные боковые веточки.
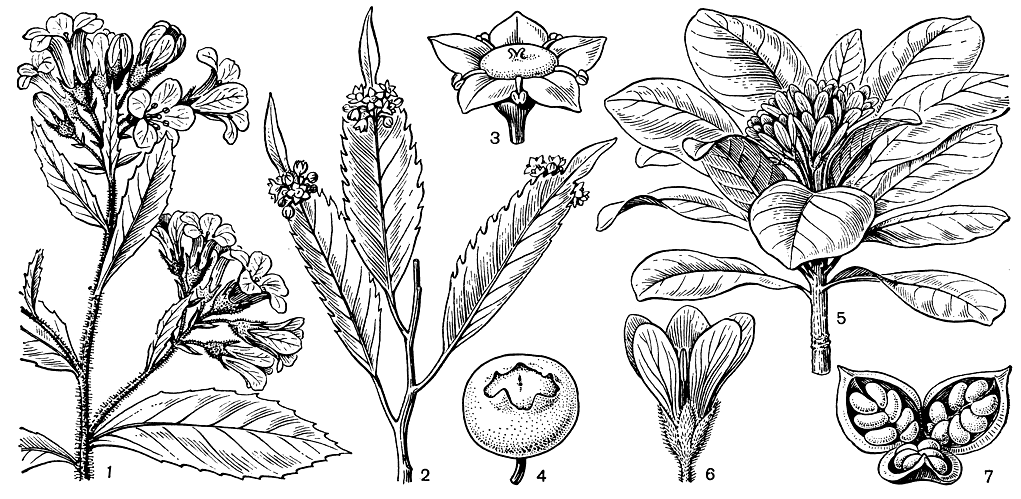
Рис. 77. Эскаллониевые и питтоспоровые. Эскаллония Клаусена (Escallonia claussenii): 1 - цветущая ветвь. Филлонома широкоостриевая (Phyllonoma laticuspis): 2 - цветущая ветвь; 3 - цветок: 4 - плод. Питтоспорум обыкновенный (Pittosporum tobira): 5 - цветущая ветвь; 6 - цветок; 7 - раскрывшийся плод
Цветки эскаллониевых обычно обоеполые, очень варьирующие по строению и опыляющиеся короткохоботковыми насекомымп. Чашелистиков и лепестков чаще всего 5, реже 6-9; причем чашелистики могут срастаться у основания в трубку, с которой нередко срастается и завязь, становясь полунижней или нижней. В некоторых родах наблюдается переход к двудомности. Так, у новозеландского карподетуса пильчатого (Carpodetus serratus) женские и обоеполые цветки встречаются на разных растениях, причем на обоеполых растениях только 12% цветков образует плоды. Наиболее обычное число тычинок в цветках эскаллониевых 5, реже 4-9, плодолистиков - от 2 до (5) 7. Апокарпный гинецей из 4 плодолистиков и плод-многолистовка имеются только у тасманского кустарника тетракарпеи (Tetracarpaea). Большинство видов имеют паракарпный или вторичносинкарпный гинецей и плод-коробочку, раскрывающуюся 2-3 створками. Выпадающие из коробочек семена обычно рассеиваются ветром (у аноптеруса - Anopterus - они снабжены широким перепончатым крылом). Встречаются и мясистые, ягодообразные плоды, которые могут быть как односемянными (у полиосмы - Polyosma), так и многосемянными (у филлономы с 3-6 семенами).
Хозяйственное значение эскаллониевых невелико. Местные жители используют древесину многих представителей семейства, хотя она не отличается высокими техническими качествами. Многие виды эскаллонии (эскаллония розовая - Escallonia rosea, эскаллония красная - E. rubra и др.) и некоторые виды других родов очень декоративны и культивируются в ботанических садах, а в умеренно теплых областях - в оранжереях.
Семейство гортензиевые, или гидрангиевые (Hydrangeaceae) (Н. Н. Цвелев)
Широко известны такие декоративные кустарники, как чубушник душистый (Philadelphus coronarius), обычно называемый "диким жасмином", и гидрангия садовая, или "гортензия" (Hydrangea hortensis). Оба эти вида принадлежат к относительно небольшому (20 родов и около 260 видов) семейству гидрангиевых, распространенному преимущественно в субтропических, а отчасти также в умеренно теплых и тропических областях северного полушария. Особенно много гидрангиевых в Восточной и Юго-Восточной Азии и в восточной части Северной Америки - в районах, наиболее богатых прежде широко распространенными третичными реликтами.
Значительное большинство гидрангиевых - листопадные или вечнозеленые кустарники, реже небольшие деревья, лианы или полукустарники. К корневищным травам принадлежат лишь очень оригинальная и выделяемая в особое подсемейство японская киренгешома пальчатая (Kirengeshoma palmata, рис. 78) и виды небольшого японо-китайского рода кардиандра (Cardiandra). Листья гидрангиевых обычно цельные (очень редко перисто - или пальчатолопастные), без прилистников, расположенные супротивно, очень редко мутовчато или очередно (у кардиандры).
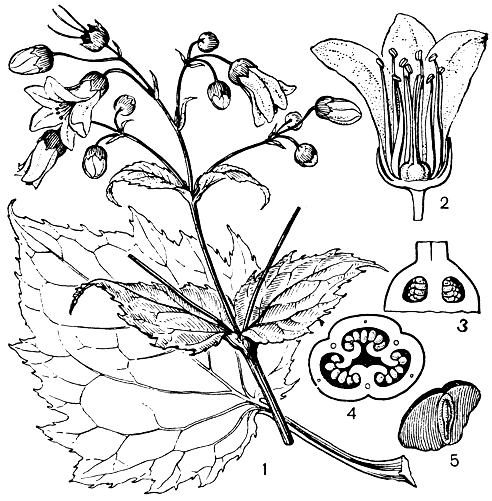
Рис. 78. Киренгешома пальчатая (Kirengeshoma palmata): 1 - часть растения с бутонами и цветками; 2 - цветок в разрезе; 3 - продольный разрез завязи; 4 - поперечный разрез завязи; 5 - семя
Почти всегда обоеполые (лишь у гавайской бруссесии - Broussaisia - однополые, нередко довольно крупные цветки гидрангиевых обычно собраны в кистевидные, щитковидные или метельчатые соцветия. Лишь у североамериканского кустарника фендлеры скальной (Fendlera rupicola) крупные белые цветки расположены по одному на верхушках густо облиственных боковых ветвей. Все гидрангиевые - энтомофильные растения. Их цветки содержат нектар и нередко имеют приятный запах, особенно сильный и напоминающий аромат жасмина у многих видов чубушника, опыляемых преимущественно ночными бабочками. У родов подсемейства гидрангиевых (Hydrangeoideae), в отличие от подсемейства чубушниковых (Philadelphoideae), краевые цветки щитковидных или кистевидных соцветий обычно стерильны и имеют сильно увеличенные лепестковидные чашелистики при отсутствии настоящих лепестков, что служит дополнительным средством привлечения насекомых (табл. 21). У видов гидрангии такие краевые цветки обычно немного зигоморфные и состоят из 3-5 лепестковидных чашелистиков, имеющих различную (белую, розовую, голубоватую) окраску, а у восточноазиатской схизофрагмы (Schizophragma) краевые цветки имеют всего один, но очень крупный чашелистик.
Фертильные цветки гидрангиевых обычно имеют чашелистиков и лепестков по 4-5, тычинок 4-5, 8-10, или они очень многочисленные, плодолистиков (2) 3-5 (10), образующих вторично синкарпный или паракарпный гинецей с нижней или полунижней завязью и свободными или сросшимися столбиками. Наиболее обычный тип плода у гидрангиевых - коробочка, раскрывающаяся створками или продольными щелями, с многочисленными семенами, распространяющимися с помощью ветра (иногда на них имеются крыловидные придатки, увеличивающие парусность). Встречаются также мясистые плоды, распространяемые птицами. Так, виды азиатского рода дихроа (Dichroa) имеют плод - голубую многосемянную ягоду.
Многие виды гидрангиевых очень декоративны и культивируются в садах, оранжереях и в качестве комнатных растений. Выше уже упоминались самые известные из них - "гортензия" и "дикий жасмин". Родоначальником многочисленных сортов садовой "гортензии" является японская гидрангия крупнолистная (H. macrophylla), культивировавшаяся в Японии и Китае с давних времен. В Европу "гортензия" была впервые привезена из Китая в 1789 г. английским ботаником Д. Банксом и быстро стала популярным декоративным растением. У большинства садовых сортов "гортензий", размножаемых черенками, все цветки стерильны, образуя крупные почти шаровидные соцветия. В более южных районах в открытом грунте культивируется еще несколько видов гидрангии: гидрангия метельчатая (H. paniculata), гидрангия древовидная (H. arborescens) и др. Из видов чубушника в наших садах и парках особенно широко культивируется чубушник душистый с душистыми белыми цветками в кистях, происходящий из Южной Европы. Встречаются в культуре и дикорастущие в СССР виды чубушник кавказский (P. caucasicus, табл. 21) и дальневосточные - чубушник тонколистный (P. tenuifolia) и чубушник Шренка (P. schrenkii). Несколько реже встречаются в культуре виды другого красиво цветущего рода гидрангиевых - дейции (Deutzia), 2 вида которого (дейция мелкоцветковая - D. parviflora и дейция головатая - D. glabrata) встречаются дико в лесах Дальнего Востока. Наиболее же декоративна японская дейция стройная (D. gracilis).
Семейство роридуловые (Roridulaceae) (В. Н. Гладкова)
Единственный род семейства роридула (Roridula) объединяет два эндемичных для Капского царства (Южная Африка) вида кустарников - роридулу зубчатую (R. dentata, рис. 79) и роридулу Горгоны (R. gorgonias). Оба вида растут в засушливых в летнее время районах на горных склонах с умеренным увлажнением среди зарослей склерофильных кустарников средиземноморского типа: роридула Горгоны - невысокий (30-50 см) кустарник - на болотистых или с просачивающейся влагой участках, нередко заросших сфагнумом, а роридула зубчатая, более высокое (до 150 см) и более ксероморфное растение,- на песчаных и пересыхающих летом местах, но обычно с доступными подземными водами от проходящих поблизости водотоков.
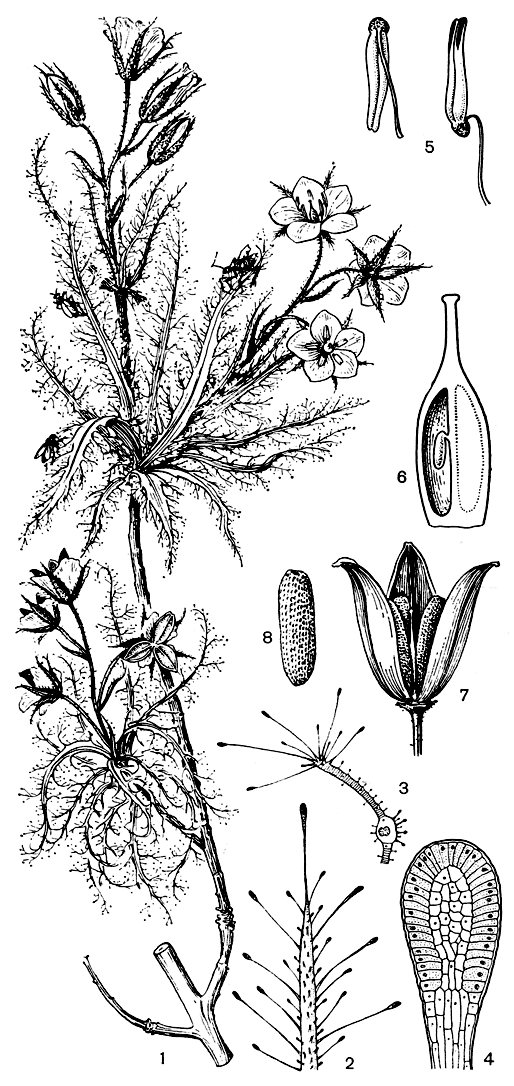
Рис. 79. Роридула зубчатая (Roridula dentata): 1 - цветущий побег; 2 - верхушка пластинки листа; 3 -поперечный разрез пластинки листа через среднюю жилку; 4 - продольное сечение крупной желёзки листа; 5 - тычинки; 6 - продольный разрез завязи; 7 - раскрывшаяся коробочка; 8 - семя
Характерный облик этим кустарникам придают удлиненные цельные или перистораздельные ланцетные или шиловидные листья желто-зеленого цвета, сближенные на концах ветвей и поблескивающие от капелек бальзама, выделяемого железистыми волосками, сплошь покрывающими эти растения. Отмершие бурые листья у роридулы Горгоны свисают вниз от основания олиственной части побегов, слегка закручиваясь на концах, словно змеи с головы Горгоны Медузы.
На листьях можно видеть множество прилипших к их клейкой поверхности погибших или барахтающихся в тщетных попытках освободиться насекомых, и пауков и клопов, высасывающих их соки.
Растения имеют красивые красные, белые или розовые цветки, собранные в немногоцветковые конечные кисти. Чашелистиков, лепестков и тычинок по 5, чашелистики срослись у основания, завязь голая, трехгнездная, с 1-4 висячими семязачатками в гнезде. Плод - 3-створчатая локулицидная коробочка, содержащая несколько довольно крупных семян с зернисто-шиповатой ослизняющейся при увлажнении поверхностью.
Долгое время роридулу причисляли к насекомоядным растениям и даже относили к семейству росянковых, пока работами Р. Марлота (1925) и Ф. Ллойда (1934) не было показано, что роридула не переваривает пойманных насекомых, так как секрет ее желёзок не содержит протеолитического фермента и состоит из смеси смолистых и каучукоподобных веществ. Желёзки роридулы имеют и принципиально иное, чем у росянок, строение: их ножка лишена проводящей ткани, а в эллипсоидальной головке проходят расширенные в нижней части канальцы, в которых собирается секрет, выделяемый эпидермальными и, возможно, паренхимными клетками головки. Канальцы сообщаются со схизолитическим отверстием на верхушке головки, и через это отверстие смолистый секрет капельками выступает наружу. По предположению Р. Марлота, секрет предназначен для отпугивания гусениц и улиток. Возможно, однако, что и гибнущие в липком секрете насекомые не совсем бесполезны для растения: попадая на землю вместе с отмершими листьями, они обогащают азотом бедную почву, на которой растет роридула.
Пять видов насекомых (2 - клопов из рода Pameridea и 3 - пауков из рода Synaema) постоянно живут на кустах роридулы, совершенно беспрепятственно передвигаясь по липкой поверхности листьев и цветков. Пауки устраивают среди оснований старых листьев гнезда и прячутся там, поджидая добычу; клопы сидят на листьях там, где мало желёзок Как только движение ветки выдает присутствие пойманного насекомого, пауки и клопы быстро устремляются к нему и высасывают из него соки. Каким образом им удается передвигаться по липкой поверхности, к которой приклеиваются все другие насекомые, даже такие крупные, как шмели, остается недостаточно ясным. Ф. Ллойд, производивший специальные наблюдения этого явления, объяснил его несколькими фактами.
Тело и ножки этих видов насекомых покрыты тонкими острыми щетинками, тем самым площадь соприкосновения насекомого с липкой поверхностью сведена до минимума. Ножки к тому же довольно длинные и во время движения достаточно высоко поднимают тело над поверхностью, так что оно ее почти не касается. Кроме того, эти насекомые бегают так быстро, что тонкие щетинки ножек не успевают погрузиться в секрет, который на поверхности менее липкий, чем внутри. Возможно также, что насекомые выделяют какое-то вещество, оберегающее их от прилипания.
Извлекает ли растение какую-либо пользу от такого сосуществования? Пауки являются, по-видимому, просто иждивенцами, клопы же, по наблюдениям Р. Марлота, производят опыление. Оба вида роридулы имеют раздражимые тычинки, связник которых в нижней части сильно утолщен, а пыльники в бутоне и в неопыленном цветке инвертированы (перевернуты) и прижаты к тычиночной нити (рис. 79). Утолщения связников пяти тычинок образуют кольцо вокруг столбика, который вначале короче тычиночных нитей. Внутри утолщенной части связников содержится сахаристый сок. Добывая его, клоп прокалывает связник хоботком, и в ответ на это действие пыльники внезапно распрямляются, поворачиваясь на 180°, и раскрываются апикальной порой, извергая облако пыльцы на рыльца цветков и на насекомое, которое переносит пыльцу на другие более удаленные цветки.
Семена роридулы высыпаются при растрескивании коробочек и особых приспособлений для распространения не имеют. Неровная поверхность, возможно, помогает им удерживаться в шерсти животных.
Местное население приносит в дома ветки этих растений и использует их в качестве мухоловок-липучек .
Семейство питтоспоровые, или смолосемянниковые (Pittosporaceae) (Н. Н. Цвелев)
Ареал этого относительно небольшого (9 родов и около 240 видов) семейства ограничен тропическими и отчасти субтропическими областями восточного полушария. Значительное большинство его видов распространено в Австралии: 8 родов являются эндемичными для этого континента (включая Тасманию), а наиболее крупный (около 150 видов) род питтоспорум, или смолосемянник (Pittosporum, табл. 21), представлен в различных экологических условиях, от тропических дождевых лесов до скрэба (зарослей ксерофильных кустарников).
Среди питтоспоровых преобладают вечнозеленые деревья и кустарники с очередными кожистыми цельнокрайними, реже зубчатыми или лопастными листьями без прилистников. Некоторые из древесных видов достигают в высоту 20 м и более. Среди видов скрэба немало колючих кустарников и эрикоидных кустарничков с очень мелкими толстоватыми листьями. Напротив, у гавайского вида дождевых лесов питтоспорума Хозмера (Pittosporum hosmeri) листья достигают 30 см в длину. Некоторые смолосемянниковые начинают свое развитие в качестве эпифитов - на стволах других деревьев. Для всего семейства очень характерно присутствие в коре, а обычно и в других частях растения смолоносных канальцев.
Пятичленные, иногда немного зигоморфные цветки питтоспоровых обычно расположены зонтиковидными щитками или метелками. Однако у небольших кустарников и полукустарников из рода биллардъера (Billardiera) с обвивающимися вокруг опоры ветвями довольно крупные, повисающие на тонких цветоножках цветки расположены по одному на верхушках ветвей, а некоторые виды мариантуса (Marianthus) имеют также одиночные, но пазушные цветки. У питтоспорума Ледермана (P. ledermanii) цветки в щитке тесно скучены, образуя головчатое соцветие. Отмечается в семействе и каулифлория: у питтоспорума ветвецветного (P. ramiflorum) цветки образуются на старых ветвях, а у питтоспорума стеблецветного (P. cauliflorum) - на стволах. Однополые цветки известны только у некоторых видов питтоспорума. Лепестки питтоспоровых обычно срастаются основаниями, образуя трубку, достигающую у видов биллардьеры довольно большой длины. Обычными опылителями цветков являются насекомые: перепончатокрылые, мухи и жуки.
Гинецей питтоспоровых всегда паракарпный из 2, реже из 3-5 плодолистиков с простым столбиком. Главным образом по строению плода семейство делится на 2 трибы: смолосемянниковых (Pittosporeae) с плодом-коробочкой и биллардьеровых (Billardiereae) с плодом-ягодой. Раскрывшиеся коробочки большинства видов питтоспорума выглядят очень эффектно: их внутренняя сторона окрашена в красный или желтый цвет и покрыта клейкой смоловидной массой того же цвета, в которой расположены темно окрашенные семена. Птицы поедают клейкое содержимое коробочки вместе с семенами и, таким образом, способствуют их распространению. Виды, имеющие ягодообразные плоды, также распространяются с помощью птиц. Лишь у немногих видов семейства семена распространяются с помощью ветра. Так, у монотипного новогвинейско-австралийского рода гименоспорум (Hymenosporum) коробочки лишены клейкого содержимого, а семена снабжены крыловидной каймой, увеличивающей их парусность.
Хозяйственное значение питтоспоровых невелико. Их древесина используется местным населением, но не отличается высокими качествами. Многие кустарники этого семейства очень декоративны, но за пределами тропиков могут культивироваться только в оранжереях. Лишь японо-китайский вид питтоспорум обыкновенный (P. tobira, рис. 77) широко культивируется в открытом грунте в странах Средиземноморья, а также на Черноморском побережье. Он имеет блестящие кожистые листья и цветки, по форме, цвету и запаху напоминающие цветки апельсина.
Семейство библисовые (Byblidaceae) (В. Н. Гладкова)
Библисовые - небольшое семейство насекомоядных трав, содержащее всего один род библис (Byblis) с двумя видами, один из которых библис гигантский (В. gigantea, рис. 80) - произрастает на западе Австралии, а другой - библис льноцветковый (В. liniflora) - в северной тропической части Австралии и на крайнем юге Новой Гвинеи.

Рис. 80. Библис гигантский (Byblis gigantea): 1 - общий вид растения; 2 - верхушка листа; 3 - тычинки и гинецей (околоцветник удален); 4 - тычинка; 5 - гинецей; 6 - поперечный разрез завязи; 7 - зрелая коробочка с остающимися чашелистиками; 8 - семя
Библисы - невысокие, с тонким корневищем одно-двулетние растения (библис льноцветковый) или довольно крупные (высотой до 0,5 м) многолетники с древеснеющими корневищами (библис гигантский), обитающие на влажных кислых почвах по берегам рек, болотам или на песках, заболачивающихся в зимний период и пересыхающих летом. Библис гигантский нередко поселяется также на гарях и нарушенных местообитаниях, исчезая из этих мест по мере восстановления естественной растительности.
Листья библисов очередные, без прилистников, длинные, тонкие, линейные по форме у библиса льноцветкового в почке и в молодом состоянии улиткообразно закручены. Толстостенные эпидермальные клетки и механические ткани проводящих пучков придают листьям жесткость и упругость. Их верхушки шишковидно утолщены, и эти утолщения, содержащие внутри массу трахеидальных клеток, связанных с сосудами доходящего почти до верхушки проводящего пучка, и, вероятно, функционируют как гидатоды. Цветки пазушные, одиночные, на длинных цветоносах, с нежными голубыми, голубовато-фиолетовыми или розовато-красными венчиками. Чашелистиков и лепестков по 5, сросшихся в основании, лепестки обычно зубчатые по краю. Тычинок 5, свободных или внизу приросших к основаниям лепестков, пыльники прикреплены основанием и раскрываются короткой щелью или верхушечной порой. Гинецей синкарпный, из 2 плодолистиков; завязь двугнездная, с многочисленными семязачатками. Плод - округлая или обратнояйцевидная коробочка, семена с гребневидно морщинистой тестой. При плодах удлиняются и сохраняются чашелистики.
Стебли, листья, цветки библисов покрыты железистым опушением. Желёзки двух типов: стебельчатые и сидячие. Первые встречаются рассеянно или скоплениями на всех частях растений. Они состоят из длинных ножек и дисковидных головок из обычно 32 расходящихся радиально от центра клеток и внешне напоминают маленькие зонтики. Клетки ножек имеют толстые целлюлозные стенки с косой исчерченностью, благодаря чему ножки способны винтообразно складываться (при пересыхании) и снова распрямляться.
Сидячие желёзки более характерны для стеблей и листьев, на которых они располагаются ровными рядами в параллельных бороздках эпидермиса, прерываясь приподнятыми устьицами. Сидячая желёзка состоит из двух базальных клеток, короткой поддерживающей клетки и небольшой головки из 8-16 клеток.
Желёзки богаты плазмой и в живом состоянии обычно красного цвета. Через поры кутикулы они выделяют слизистый секрет. Более липкий секрет стебельчатых желёзок служит для улавливания насекомых, а секрет сидячих желёзок по некоторым наблюдениям обладает протеолитическими свойствами и служит для их переваривания.
Как и на некоторых других насекомоядных растениях и на представителе родственного семейства роридуловых роде роридула (Roridula), на библисах живут мелкие бескрылые клопы, питающиеся соками пойманных растением насекомых. Обычно они прячутся на верхней поверхности листьев, где меньше липких стебельчатых желёзок, но способны мчаться к добыче в любом направлении (см. роридуловые). Неизвестно, способствуют ли эти клопы опылению или их сожительство с библисами представляет собой род комменсализма, когда один вид кормится остатками пищи другого, не принося ему ни вреда, ни пользы.
Семейство камнеломковые (Saxifragaceae) (Г. Л. Кудряшова)
Камнеломковые - большое и разнообразное семейство, объединяет около 30 родов и до 600 видов, распространенных преимущественно в холодной и умеренной зонах северного полушария, за исключением немногих видов нескольких родов, известных в южном полушарии (в Андах Южной Америки и в Голантарктическом царстве).
Среди камнеломковых нет древесных растений. Все они травы, многолетние или редко однолетние. Листья простые, цельные или реже пальчатые или перистые, как правило, очередные, обычно без прилистников (рис. 81). Цветки одиночные или собранные в разного типа верхушечные соцветия, обоеполые или редко однополые и двудомные, как у японо-китайского рода танакея (Тапакеа, рис. 82), актиноморфные или реже зигоморфные, околоцветник обычно 5-членный, реже 3-членный, образующий более или менее хорошо развитую цветочную трубку, свободную или в различной степени приросшую к основанию завязи (рис. 81, 2). Лепестки иногда отсутствуют, например у селезеночника (Chrysosplenium). Тычинок 5-10, редко только 3, как у североамериканского рода толмия (Tolmiea). Гинецей апокарпный, как у примитивного азиатско-североамериканского рода астилъбе (Astilbe) или чаще синкарпный или паракарпный, из 2-5 плодолистиков; столбики свободные или реже сросшиеся; завязь верхняя, полунижняя или нижняя, со многими или несколькими семязачатками. Плод - коробочка, раскрывающаяся по перегородкам. Семена мелкие, с маленьким зародышем, окруженным обильным эндоспермом.
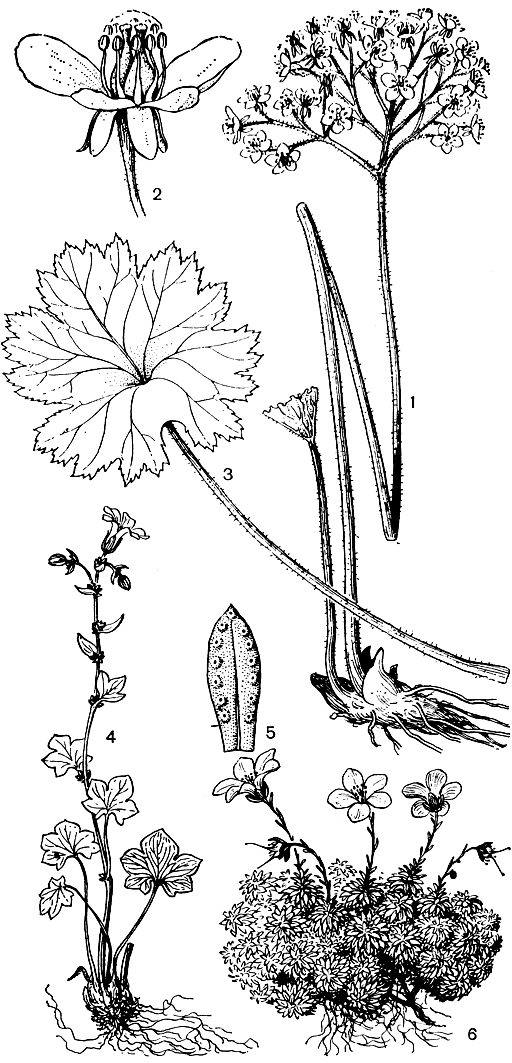
Рис. 81. Камнеломковые. Пелтифиллум щитковидный (Peltiphyllum peltatum): 1 - общий вид растения; 2 - цветок; 3 - лист. Камнеломка поникающия (Saxifraga cernua): 4 - общий вид растения (в пазухах стеблевидных листьев видны луковички). Камнеломка желто-зеленая (S. luteoviridis): 5 - прикорневой лист (по краю видны гидатоды, выделяющие известь). Камнеломка Динника (S. dinnikii): 6 - общий вид растения-подушки с цветками и плодами
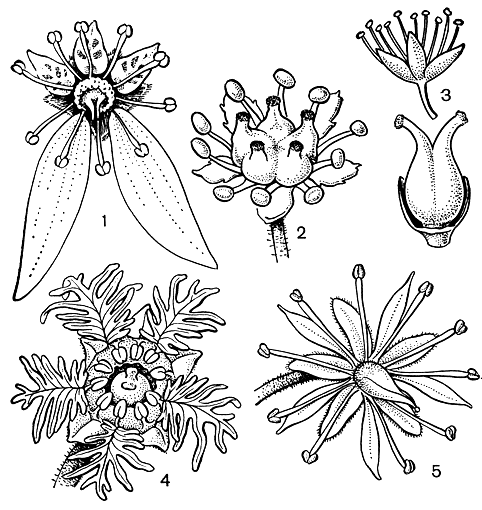
Рис. 82. Цветки представителей семейства камнеломковых: 1 - камнеломка столоносная (Saxifraga stolonifers), зигоморфный цветок; 2 - пенторум низкорослый (Penthorum sedoides), цветок с простым чашечковидным околоцветником; 3 - танакея укореняющаяся (Tanakea radicans), мужской и женский цветки двудомного растения; 4 - мителла двулистная (Mitella diphylla), цветок с рассеченными лепестками; 5 - тиарелла Верри (Tiarella wherryi), актиноморфный цветок
На громадном пространстве своего ареала камнеломковые распределены очень неравномерно. По всему ареалу семейства представлены виды только одного рода - камнеломка (Saxifraga, см. рис. 81) - самого большого по количеству видов (около 370). Основное разнообразие родов и видов камнеломковых сосредоточено в горных странах Азии, Европы, Америки. Участие камнеломковых в растительном покрове особенно бросается в глаза в Арктике и высокогорьях. В южных Альпах, на Кавказе, в Гималаях, на Скалистых горах Америки, особенно на известняках, камнеломковые, особенно род камнеломка, представлены наибольшим богатством узколокальных эндемичных форм. Второе место по широте распространения занимает род селезеночник (до 60 видов), представленный в Азии (большинство видов), Америке, Европе и Северной Африке. Несколько небольших родов (1 -12 видов) американо-азиатские. Около половины родов семейства ограничены в своем распространении горными областями Северной и Южной Америки и насчитывают в своем составе не более 1-5 видов. Исключение составляют род геухера (Heuchera, около 70 видов) и литофрагма (Litophragma, 20 видов). Восемь родов семейства распространены только в Азии. Некоторые секции самых больших родов камнеломка и селезеночник целиком или почти целиком азиатские.
Почти все камнеломковые - многолетние травы от нескольких сантиметров до полутора метров высотой. Камнеломковые в основном гигрофиты и мезофиты и лишь небольшая часть субксерофиты. Исключительно или преимущественно влаголюбивы лесные виды родов астильбе, роджерсия (Rodgersia), лептарена (Leptarrhena), бойкиния (Boykinia), пелтифиллум (Peltiphyllum), боландра (Bolandra), суксдорфия (Suksdorfia), элмера (Elmera) и др. Все они имеют, как правило, мочковатые корни, крупные, сочные листья с рыхлым мезофиллом и слабо кутинизированной эпидермой. Субксерофильные виды, обитающие в низменных районах, имеют хорошо развитые корни, листья опушенные или кожистые с сильно кутинизированной эпидермой. Высокогорные субксерофильные виды способны переносить недостаток влаги благодаря мясистым сильно кутинизированным листьям. Кроме того, в розетках листьев может собираться атмосферная влага и роса, которые служат для развития побегов. У обитателей скал и осыпей часто раздвоенный, уходящий глубоко в почву главный корень, наиболее распространена подушечная форма роста. На укороченных и прижатых к земле побегах при этом наблюдается значительная редукция листовых пластинок, характеризующихся, кроме того, толстой кожистой эпидермой. Замечательной особенностью субксерофильных высокогорных видов камнеломок является способность выделять известь на поверхности листьев. Выделение извести происходит через гидатоды, которые имеются и у видов других секций, но не выделяют никакой извести. Гидатоды представляют собой группы мелких паренхимных клеток, бедных хлорофиллом или даже без него, которые лежат под эпидермой над проводящими пучками (рис. 81, 5). У видов, выделяющих известь, гидатоды расположены в углублениях, имеющих вид ямок, которые находятся на верхней стороне листа по краю или только на кончике. У видов с зубчатым или пильчатым краем эти ямки находятся у основания зубцов. Углекислый кальций в растворенном виде поступает из проводящего пучка в ямку, где после испарения воды остается осадок, к которому снизу добавляются новые частицы углекислого кальция. Постепенно ямка не только полностью заполняется, но даже за пределами ее оказывается известковый налет, так что, если ямки расположены близко друг от друга, по краю листа образуется сплошная известковая полоса, как у камнеломки метельчатой (S. paniculata). Количество выделяемой извести довольно значительно. В зависимости от секреции извести находится и глубина ямок. Последние на листьях проростков почти плоские и становятся глубже позднее, когда отделение извести усиливается, Известковый налет, образующийся на листьях, по-видимому, затрудняет отдачу воды и служит защитой от слишком интенсивного освещения.
Биология цветения камнеломковых, многие из которых приспособлены к крайним условиям существования на севере и в высокогорьях, имеет ряд особенностей. Эти растения характеризует ускоренный ритм прохождения всех сезонных фаз, от появления первых листьев и молодых побегов до образования зрелых плодов и семян. Сроки цветения зависят от сформированное репродуктивного побега в почке возобновления к концу предыдущего вегетационного периода. Раньше всех образуются зачатки цветков будущего года у камнеломки супротивнолистной (S. oppositifolia, табл. 22). В поздние дни лета у нее появляются цветочные побеги, которые осенью развиваются и в таком состоянии зимуют. Внутри цветочной почки в это время уже есть завязь, тычинки, чашелистики и окрашенные лепестки. Поэтому цветение у нее начинается ранней весной, прямо из-под снега. У большинства видов цветочные почки закладываются в июле, у некоторых еще позднее, в августе (камнеломка болотная - S. hirculus), и у них соответственно позднее наступает цветение на будущий год. Как правило, семена у позднецветущих видов не вызревают к наступлению зимы, нередко они даже уходят под снег в цветущем состоянии. Зато их незрелые семена прекрасно прорастают и цикл развития от семени до семени оказывается завершенным.

Таблица 22. Камнеломковые: 1 - бадан тихоокеанский (Bergenia pacifica), Дальний Восток; 2 - камнеломка супротивнолистная (Saxifraga oppositifolia), Франция; 3 - итея японская (Itea japonica), Батумский ботанический сад
Все камнеломковые - насекомоопыляемые растения. На севере и в высокогорьях это является ограничительным фактором перекрестного опыления, так как насекомых-опылителей там недостаточно. Для привлечения насекомых служат разнообразно окрашенные лепестки с более яркими точками или жилками при основании, которые, в свою очередь, указывают местонахождение нектара. Он выделяется у основания завязи, или на диске, окружающем гинецей, или на внутренней стороне лепестков в больших количествах. Ткань нектарников состоит из более мелких, чем окружающие, клеток и содержит глюкозу. Секреторную функцию выполняют устьица или поры, рассеянные по всей эпидерме нектарников. Открытые цветки камнеломковых с легко доступным нектаром чаще всего посещают короткохоботковые насекомые (мухи). Последних наблюдали даже на высоте 3000 м на цветках камнеломки проломниковой (S. androsacea). Нектар у камнеломки супротивнолистной находится глубже и доступен только мотылькам и шмелям. И как раз у этого раннецветущего высокогорного вида чаще происходит самоопыление из-за отсутствия насекомых-опылителей. Случайное посещение цветков муравьями наблюдалось у камнеломки жестколистной (S. aizoides). Перекрестному опылению у камнеломковых способствует также дихогамия, неодновременное созревание тычинок и рыльца (протандричные или протогиничные цветки соответственно), хотя наряду с этим существует и гомогамия (одновременная готовность тычинок и рыльца к опылению). Нередки виды, которые бывают то дихогамными, то гомогамными. Самоопыление известно и в том и в другом случае, когда не произошло перекрестное опыление. У некоторых видов литофрагмы самоопыление исключается благодаря несовместимости.
Камнеломковые обычно образуют очень много мелких семян, за исключением тех видов, у которых большую роль играет вегетативное размножение. Ко времени созревания семян коробочки раскрываются и семена могут высыпаться, когда растения раскачиваются от ветра, толчков животных и т. д. (распространение по типу баллистов). У лесных видов селезеночника, мителлы (Mitella), тиареллы (Tiarella) семена вылетают из коробочек при падении на них водяных капель. У многих видов высохшие плоды целиком разносятся потоками воды, растаскиваются мышами, повисают на шерсти крупных животных (в последнем случае благодаря присутствию железистых липких волосков). Семена разных родов камнеломковых кажутся очень похожими между собой из-за малых размеров (длиной 0,5 мм). Однако на самом деле они очень разнообразны по форме, скульптуре оболочки, снабжены различными придатками в зависимости от способа распространения. Семена видов - обитателей сильно увлажняемых мест - обычно с гладкой поверхностью, обладают хорошей плавучестью и переносятся водой на большие расстояния. Исследования показали, что семена камнеломки жестколистной остаются плавучими более 3 недель, у камнеломки снежной (S. nivalis), камнеломки звездчатой (S. stellaris), также влаголюбивых видов, семена могут оставаться в воде 18 дней, не теряя жизнеспособности, в то время как семена скальных видов - камнеломки моховидной (S. bryoides), камнеломки метельчатой, камнеломки мускусной (S. moschata) тонули через два дня.
Кроме семенного размножения большую роль у камнеломковых играет вегетативное размножение. Наиболее распространенным и продуктивным способом вегетативного размножения среди камнеломковых является способность к образованию выводковых почек в пазухах листьев. Выводковые почки состоят из укороченной оси и мясистых, бедных хлорофиллом листочков, прижатых друг к другу и напоминающих луковичку. В литературе их нередко так и называют - луковичками или бульбочками. По месту их образования различают воздушные бульбочки - в пазухах стеблевых и прицветных листьев (у видов камнеломок) и подземные или прикорневые (виды родов камнеломка, литофрагма, суксдорфия). Некоторые камнеломки, назовем для примера наши северные виды - камнеломка листочковая (S. foliolosa), камнеломка кимвальная (S. cymbalaris),- размножаются и распространяются только благодаря своей способности к вивипарии и практически не образуют плодов и семян. У большинства же видов вегетативный способ размножения существует параллельно с семенным. Кроме образования выводковых почек, у камнеломковых существуют и другие способы вегетативного размножения: образование зачатков новых растений на концах ползучих побегов (камнеломка усатая - S. flagellaris, виды рода мителла), столонов (камнеломка ручейная - S. rivularis), на листьях (виды рода толмия), повторное деление мясистого главного корня и образование таким путем целых колоний новых растений у видов рода геухера, тиарелла, литофрагма, джепсония (Jepsonia).
Многие камнеломковые издавна известны человеку как красиво цветущие растения, а некоторые и как лекарственные. Название "камнеломка" чаще всего связывают с высокогорными видами, растущими на скалах (считалось, что они образуют в них трещины). Другое объяснение названия "камнеломка" связывают с применением каких-то видов в народной медицине против камней в почках. Есть данные о бактерицидных свойствах некоторых камнеломок. В Сибири и на Алтае используют листья бадана толстолистного (Bergenia crassifolia) как народное лекарство и суррогат чая. Более важное значение имеет бадан как дубильное растение. В листьях его содержится до 23 % , а в корневищах до 27% танидов. В связи с этим в 1927 г. бадан введен в культуру. В течение 8-10 лет используют листовую массу, а затем корневища в кожевенной промышленности. Из листьев получают гликозид арбутин, применяемый в медицине и фотографии. Клетки с дубильными веществами обнаружены также у ряда других видов камнеломковых (астильбе, роджерсия, бойкиния, камнеломка, селезеночник), но в незначительных количествах. В основном же камнеломковые вводятся в культуру как декоративные растения. Виды астильбе, бадана, пельтифиллума, геухеры имеют крупные пирамидальные, метельчатые, зонтиковидные, кистевидные соцветия из белых, розовых или пурпурных цветков, появляющихся ранней весной. Они не требовательны к условиям произрастания и широко распространены в садах и парках, особенно на каменистых горках и в затененных местах. В культуре выведено много гибридов, превосходящих по красоте исходные виды, например между астильбе японской (Astilbe japonica) и астильбе китайской (A. chinensis). Своеобразна красота многих камнеломок, особенно высокогорных и арктических видов, образующих подушки из розеток довольно широких, нередко толстых и мясистых, напоминающих толстянковые, листьев или чаще из многочисленных побегов, покрытых узкими кожистыми, иногда колючими листьями. Весной подушки камнеломок, достигающих в диаметре нередко 1 м, почти сплошь покрыты желтыми, белыми, пурпурными, розовыми цветками, одиночными или собранными в небольшие соцветия, создающими замечательный контраст с густыми жесткими подушками. Многие камнеломки, главным образом европейские виды, уже давно выращиваются в садах на каменистых горках, но несравненно красивей камнеломки в природе, на скалах, в неожиданных сочетаниях с другими растениями. Любителям комнатного цветоводства хорошо знакома камнеломка столононосная (S. stolonifera). Область ее естественного распространения лежит в Китае и Японии. Она красива не только кистевидно-метельчатыми соцветиями изящных цветков с неравными лепестками, но и округлыми ярко-зелеными с красными жилками, сильно опушенными листьями. От основания листьев отходит множество нитевидных побегов разной длины, несущих на концах зачатки новых растений, так что это и красивое ампельное растение.
Семейство толстянковые (Crassulaceae) (В. М. Виноградова)
Характерный облик представителей семейства толстянковых отражен в названии семейства. Слово "crassus" на латинском языке означает "толстый". Мясистые, сочные стебли и листья составляют примечательную черту этого семейства. Толстянковые, подобно кактусовым, относятся к экологической группе растений засушливых областей - суккулентам. В семейство входят более 30 родов и 1500 видов, распространенных очень широко, но главным образом в теплых и засушливых областях. Наиболее богата родами толстянковых Африка, особенно Южная Африка, к которой приурочено большинство видов таких крупных родов, как толстянка (Crassula, более 300 видов), каланхое (Kalanchoe, около 200 видов). Богато представлены толстянковые также в сухих областях Америки, от юго-запада США до Мексики и Центральной Америки. Но самый крупный род в семействе - очиток (Sedum), насчитывающий, вероятно, до 600 видов, обитающих в основном в умеренной полосе в пределах северного полушария, преимущественно в Евразии. Толстянковые совершенно отсутствуют в Австралии и Полинезии.
Большинство толстянковых обычно растут на более или менее сухих открытых местах, очень часто среди камней и в трещинах скал. Иногда они встречаются в кустарниках, на лугах и в тенистых ущельях, а некоторые виды даже живут в воде (толстянка водная - С. aquatica). Толстянковые встречаются от уровня моря до альпийского пояса. Иногда они бывают эдификаторами растительного покрова. Например, в Капской области как на побережье, так и в горах широко распространены ассоциации древовидных толстянковых, в которых доминируют виды рода котиледон (Cotyledon). Очиток едкий (S. acre) в период цветения дает роскошный желтый ковер. Широко распространенный по всей Европе этот вид предпочитает сухие возвышенные места с песчаными почвами.
Большинство толстянковых - травы, но среди них встречаются также полукустарники и небольшие кустарники. Установлено, однако, что стебли кустарниковидных форм имеют травянистую анатомическую структуру. Древовидную форму имеют, например, южноафриканские виды толстянка древовидная (C. arborescens) и толстянка серебристая (C. argentea), у которых имеется один слегка одревесневающий стебель, достигающий в высоту 2-3 м. Каланхое перистый (K. pinnata) имеет более типичную кустарниковидную форму роста - он развивает несколько стеблей. Многие виды толстянок несут густо облиственные толстые колонновидные стволики, на верхушке которых формируются соцветия (толстянка пирамидальная - C. pyramidalis). Некоторые из них (толстянка колончатая - C. columnaris) в первые годы жизни имеют вид круглых камней размером от грецкого ореха и больше, и только через 5-10 лет, когда они переходят к цветению, "камень" вырастает в короткую колонку с плотно прилегающими листьями и соцветием на верхушке. Эти виды толстянок-монокарпики; они цветут один раз, а затем отмирают, успев образовать в пазухах листьев при основании стебля отпрыски с плотными розетками листьев.
Наиболее часто в семействе встречается травянистая форма роста; прямостоячие, полегающие и ползучие травы имеют чаще всего многочисленные ветвящиеся стебли. Среди них есть как однолетние, так и двулетние и многолетние растения. Прекрасным примером многообразия форм роста толстянковых являются некоторые виды почти исключительно макаронезийского рода эониум (Aeonium, рис. 83), некоторые из них, по словам Ш. Карлквиста (1974), представляют собой "миниатюрные, монокарпические розеточные деревья". Древовидная, кустарниковидная и травянистая формы роста переходят здесь одна в другую через изменения в длине, толщине, густоте ветвей и соцветий.

Рис. 83. Формы роста у видов рода эоннум (Aeonium) с розеточными листьями. Древовидная форма роста: 1 - побеги толстостебельные ветвящиеся у эониума древовидного (A. arboreum); 2 - побеги канделябровидные у эониума благородного (A. nobile). Кустарниковидная форма роста: 3 - побеги толстостебельные у эониума реснитчатого (A. ciliatum); 4 - побеги ползучие у эониума Саундерса (A. saundersii); 5 - побеги полегающие у эониума очитколистного (A. sedifolium). Травянистая форма роста: 6 - со стеблем у эониума клейкого (A. glutinosum); 7 - у бесстебельного эониума Смита (A. smithii); 8 - у многолетника эониума железистого (A. glandulosum); 9 - у монокарпика эониума Канарского (A. canariense)
Листья толстянковых обычно цельные, очередные или супротивные, большей частью сидячие, часто собраны в розетки. Благодаря образованию розеток, имеющих чашевидную, цилиндрическую и луковицеобразную форму, достигается защита от интенсивной инсоляции и испарения. Устьица листьев толстянковых в сухую погоду днем бывают закрыты. Ночью при открытых устьицах CO2 поступает в растение. В ходе усвоения CO2 образуются органические кислоты, которые днем разрушаются, освобождая CO2. Последний днем используется для образования новых запасов углеводов. Такой необычный тип фотосинтеза получил название "крассулатип". Мясистые листья могут быть плоскими (очиток, каланхое), цилиндрическими (каланхое трубколистный - K. tubiflorum), щитковидными (умбиликус скальный - Umbilicus rupestris) или почти округлыми (ряд видов очитка). У каланхое перистого (K. pinnata) наряду с простыми листьями развиваются и сложные непарноперистые. Обычно листья зеленые, но часто также желтоватые или серо-голубоватые от воскового налета или с красноватым от антоциана оттенком, иногда сильно опушенные.
Цветки толстянковых, как правило, обоеполые, редко однополые, актиноморфные, собранные в верхушечные верхоцветные соцветия. У видов рода монантес (Monanthes) цветки одиночные либо в монохазиях или дихазиях. Околоцветник обычно 5-членный, реже 4-членный, 3-9-членный до 32-членного, причем число частей варьирует и непостоянно даже у одного и того же растения. Чашелистики, как правило, свободные либо сросшиеся в основании (эчеверия - Echeveria). Лепестки свободные или более или менее сросшиеся, редко полностью сросшиеся. Тычиночные нити могут быть прикреплены к основанию цветочной трубки, около ее середины или в верхней части. Тычинки в одинаковом числе с лепестками, либо их вдвое больше. Плодолистики обычно свободные или слабо сросшиеся в основании. В их нижней части находятся небольшие придатки, функционирующие в качестве нектарников. Сильно развитыми лепестковидными нектарниками выделяется в семействе род монантес, большинство видов которого растет на Канарских островах. Плод - многолистовка, редко коробочковидный (североамериканский род диаморфа - Diamorpha). Семена без эндосперма или со скудным эндоспермом.
Толстянковые - перекрестноопыляемые растения. Для многих видов характерна протандрия (умбиликус повислый - U. pendulis) и протогиния (виды рода очиток). Нектар, обильно отделяемый чешуевидными нектарниками у основания плодолистиков, защищен в начале цветения наклоненными друг к другу лепестками и основаниями тычинок, прижатых к нектарникам. У некоторых видов нектар глубоко спрятан в трубчатом цветке и доступен только длиннохоботковым насекомым. Цветки толстянковых обычно посещаются шмелями, пчелами, мухами и бабочками.
Висячие трубчатые цветки некоторых видов каланхое (например каланхое перистого), а также цветки эчеверии и южноафриканских видов котиледона опыляются птицами. Птица просовывает клюв и переднюю часть головы в длинную трубку венчика, сосет нектар и производит опыление.
У толстянковых известно также самоопыление. Так, у толстянки водной и родственных ей видов клеистогамные и автогамные цветки почти никогда не открываются и цветение, как и созревание плодов, иногда происходит под водой.
Толстянковые обычно производят большое количество семян, которые распространяются ветром, водой, муравьями. Семена обычно мелкие и легкие. Например, у мадагаскарского каланхое Блоссфельда (K. blossfeldiana) подсчитано 80 000 семян на грамм. Семена очитка едкого распространяются дождем: они вымываются из созревших открытых сверху листовок очитка по мере наполнения листовок водой. Нередко размножение видов очитка происходит вегетативным путем: на стеблях и листьях появляются придаточные корни.
Всем представителям семейства свойственна исключительная способность к вегетативному размножению. Легкость образования придаточных корней на стеблях, почек на листьях, новых розеток листьев на ползучих побегах определяет необычную жизнеспособность видов толстянковых. Особенно активен процесс почкообразования на листьях с последующим развитием молодых растений - деток. Эти маленькие растеньица с крохотными корешками можно видеть на листе каланхое Дегремона (К. daigremontiana, рис. 84). В меньшей степени образование деток свойственно каланхое перистому, а у каланхое трубколистного детки образуются на конце длинного трубчатого листа.

Рис. 84. Толстянковые. Толстянка колончатая (Crassula columnaris): 1 - общий вид растениям цвету; 2 - цветок в разрезе. Молодило Функa (Sempervivum funkii): 3 - общий вид растения в цвету; 4 - цветок в разрезе; 5 - плод-листовка. Каланхое Блоссфельда (Kalanchoe blossfeldiana): 6 - общий вид растения в цвету; 7 - цветок в разрезе. Каланхое Дегремона (К. daigremontiana): 8 - часть листа с живыми растеньицами на нем. Очиток тополелистный (Sedum populifolium): 9 - общий вид растения в цвету; 10 - цветок
Виды рода молодило (Sempervivum, табл. 23) формируют шаровидные розетки листьев на ползучих побегах. Они отделяются от материнского растения и завоевывают новые местообитания. Размножение молодила отпрыскового (S. soboliferum) тесно связано с сосновым лесом, где сухие сосновые шишки, падая в заросли молодила, разрывают стебельки, находящиеся между розетками листьев, и отбрасывают розетки на несколько метров в сторону.
Семейство толстянковых делится на 6 подсемейств. К подсемейству очитковых (Sedoideae) относятся многолетние травянистые растения с супротивными, очередными, мутовчатыми или в прикорневой розетке листьями. Цветки 3(5-12)-членные. Чашелистики и лепестки свободные. Тычинки в числе лепестков или их в два раза больше (см. рис. 84). Сюда входят роды очиток и диаморфа.
В подсемействе молодиловых (Sempervivoideae) у основных его представителей (молодило, эониум, монантес) листья очередные, большей частью в розетке, прикорневые всегда в розетке. Цветки обычно 5-32-членные, лепестки свободные, у основания сросшиеся, тычинки расположенные в два круга (см. рис. 84).
Основным представителем подсемейства эчевериевых (Echeverioideae) является крупный американский род эчеверия, насчитывающий около 150 видов. Листья очередные, обычно в розетке. Цветки 5-членные, лепестки у основания сросшиеся, тычинок в два раза больше, чем лепестков (рис. 85).

Рис. 85. Толстянковые. Эчеверия Пекокка (Echeveria peacockii): 1 - общий вид растения в цвету; 2 - цветок в развернутом виде. Адромискус Пельница (Adromishcus poellnitzianus): 3 - общий вид растения в цвету; 4 - цветок в развернутом виде
В подсемействе котиледоновые (Cotyledonoideae) у его основных родов котиледон, адромисхус (Adromischus), умбиликус, розеточница (Rosularia) листья очередные или супротивные, на черешках. Цветки 5-членные; лепестки, сросшиеся в трубку; тычинки расположены в два круга (см. рис. 85).
В подсемейство каланхоевых (Kalanchoideae) входит большой род каланхое, включая бриофиллум (Bryophyllum), который в настоящее время не выделяют из каланхое. Листья супротивные, на черешках. Цветки 4-членные; лепестки, сросшиеся в трубку; тычинок 8, расположенных в два круга и прикрепленных к трубке венчика (см. рис. 84).
К подсемейству толстянковых (Crassuloideae, табл. 23) относятся большой род толстянка и ряд небольших родов. Листья у них супротивные. Цветки мелкие, 5(4-9)-членные, одиночные или в соцветиях. Лепестки свободные, тычинки в равном числе с лепестками и прикреплены к их основаниям (см. рис. 84).

Таблица 23. Толстянковые: 1 - толстянка плауновидная (Crassula lycopodioides), оранжереи Ботанического института АН СССР в Ленинграде; 2 - молодило закавказское (Sempervivum transcaucasicum); 3 - молодило кавказское (S. Caucasicum), Кавказ, Чечено-Ингушетия; 4 - очиток розовый (Sedum rosea)
Многие толстянковые - декоративные растения. Из-за оригинальности внешнего облика их часто выращивают как комнатные растения, а также в парках, садах и оранжереях. Хорошо знакомы любителям комнатных растений каланхое (бриофиллум) Дегремона и каланхое (бриофиллум) перистый. Родина каланхое Дегремона - остров Мадагаскар, а каланхое перистый встречается в тропиках и субтропиках обоих полушарий. Родину этого вида установить теперь трудно из-за активной деятельности человека по его распространению. Листья этих видов каланхое используют в быту при самых разных заболеваниях, и не случайно эти растения называют "деревом жизни" или "комнатным женьшенем". Молодило кровельное (S. tectorum) издавна введено в культуру в Европе как красиво цветущее растение. Раньше его применяли для покрытия глиняных крыш, отчего и происходит его видовое название. Свежие листья толстянки серебристой используют в Южной Африке в качестве корма для животных. Ряд видов используют в быту как лекарственные: более 400 лет популярным средством народной медицины является очиток розовый, или золотой корень (Sedum rosea, табл. 23). Эониум бальзамический (A. balsamifera) и эониум палъмский (A. palmense) выделяются среди видов рода эониум содержанием бальзама в листьях.
Семейство цефалотовые (Cephalotaceae) (А. Л. Тахтаджян)
К этому монотипному семейству относится одно из наиболее замечательных насекомоядных растений - цефалотус мешочковидный (Cephalotus follicularis, рис. 86). Цефалотус - эндемик Юго-Западной Австралийской флористической провинции. Он встречается на ограниченной территории, которая в форме полумесяца протягивается вдоль морского побережья на крайнем юго-востоке провинции. Цефалотус растет на относительно сухих местах по окраинам торфяных болот. Это небольшое травянистое растение с подземным корневищем. Ежегодно образуется розетка тесно расположенных прикорневых листьев. Листья двух типов - верхние (внутренние) плоские, цельные, толстые, с желёзками на черешке и на нижней стороне пластинки, а нижние (внешние) превращены в сложно устроенные ловчие кувшинчики, расположенные более или менее косо на поверхности почвы. Плоские листья развиваются в течение австралийской осени (март - апрель) и достигают полного развития весной (август - сентябрь), в то время как кувшинчатые листья развиваются зимой и весной и полностью формируются и активно функционируют летом (ноябрь - январь), когда насекомые наиболее обильны. В ноябре - декабре из середины розетки поднимается очень длинный безлистный цветонос, несущий наверху соцветие, состоящее из мелких боковых дихазиев, каждый из которых состоит из 3-8 цветков. Цветение происходит в январе и в начале февраля. Цветки мелкие, беловатые, обоеполые, безлепестные. Чашечка сростнолистная, 6-лопастная. Тычинок 12, в двух чередующихся кругах, прикрепленных к верхушке трубки чашечки у внешнего края толстого диска. С внешней стороны связника образуется полушаровидная клеточная масса (см. рис. 86), что и послужило французскому ботанику Ж. де Лабиллардьер (1806), впервые описавшему это растение, поводом назвать его цефалотусом (греч. kephalotos - головчатый). Гинецей апокарпный, из 6 плодолистиков, расположенных в одном кругу; плодолистики вытянуты в слегка согнутый столбик и на вентральной стороне верхней части покрыты очень мелкими рыльцевыми сосочками. В каждом плодолистике обычно 1 (редко 2) базальных семязачатков. Плод - многолистовка, созревающая в феврале или марте. Плодики покрыты отогнутыми вниз волосками; столбики остаются при плодах, удлиняются и крючковидно загибаются наружу. Волоски и крючковидный столбик способствуют зоохорному распространению. Семена с очень маленьким зародышем, окруженным обильным мясистым эндоспермом.
В строении и жизнедеятельности цефалотуса наибольший интерес вызывают кувшинчатые листья, морфологии и биологии которых посвящена довольно большая литература. Кувшинчатые листья состоят из яйцевидного кувшинчика длиной от 0,5 до 3 см и ориентированного почти перпендикулярно к ее оси волосистого черешка. В молодом состоянии кувшинчик закрыт крышечкой, впоследствии открывающейся. Кувшинчик с крышечкой являются результатом инвагинации пластинки листа. Как известно, листья типа асцидий (от греч. askidion - мешочек) встречаются иногда в качестве аномалий у растений с нормальными плоскими листьями, что вызвано неравномерным ростом тканей. У цефалотуса довольно часто встречаются аномальные листья, представляющие разные стадии превращения обыкновенного плоского листа в кувшинчатый лист, описанные английским ботаником А. Диксоном (1882). Они до некоторой степени соответствуют стадиями онтогенетического развития кувшинчика, впервые тщательно изученного немецким ботаником А. В. Эйхлером (1881).
Строение кувшинчатого листа цефалотуса настолько замечательно, что об этом нужно рассказать несколько подробнее. Эпидерма внешней поверхности кувшинчика, состоящая из толстостенных клеток, снабжена устьицами и погруженными желёзками. Кроме того, вдоль всей длины кувшинчика тянутся три несколько уплощенных гребня. На поперечном разрезе эти гребни более или менее Т-образные, особенно средний из них. Все три гребня покрыты длинными волосками. Но наиболее интересна внутренняя поверхность кувшинчика, знакомство с которой мы начнем с его косо-вверх направленного отверстия, или зева. Край отверстия обрамлен довольно толстым ободком, или перистомом (от греч. peri - вокруг, около и stoma - рот), который прерывается лишь в месте отхождения крышечки. Ободок как бы рифленый, с чередующимися гребнями и желобками, причем каждый гребень образует когтевидный зубец, направленный книзу, внутрь урны (см. рис. 86). Зубцы темно-красного цвета и хорошо контрастируют со светло-зеленым цветом глубоких желобков.
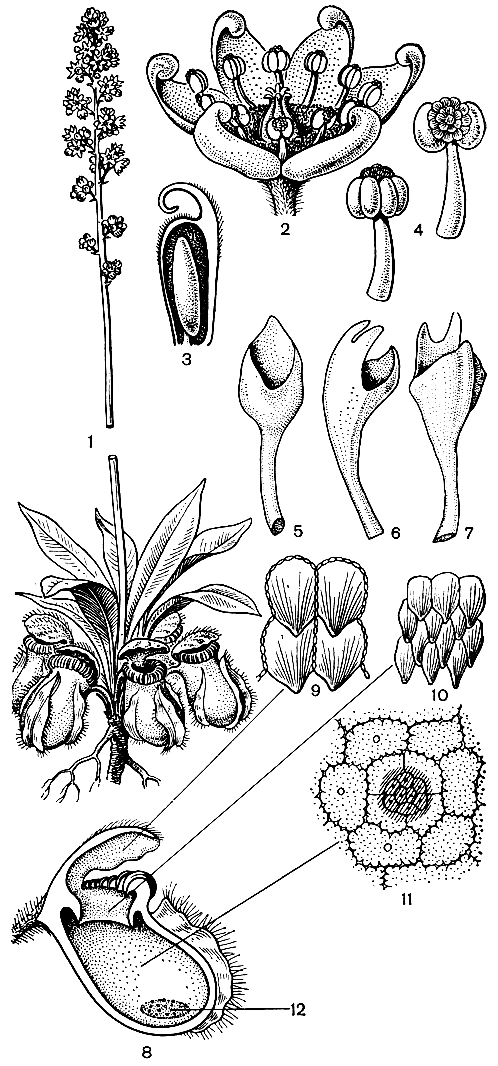
Рис. 86. Цефалотус мешочковидный (Cephalotus follicularis): 1 - общий вид растения; 2 - цветок; 3 - продольный разрез плодолистика; 4 - тычинки; 5, 6, 7 - аномальные листья, представляющие разные стадии развития кувшинчика; 8 - продольный разрез кувшинчика; 9 - клетки внутренней поверхности крышечки; 10 - клетки поверхности воротничка; 11 - клетки и желёзка верхней части внутренней поверхности кувшинчика; 12 - железистый валик на внутренней поверхности кувшинчика
Если теперь сделать продольный разрез кувшинчика, то в верхней его части мы увидим бледно-зеленый воротничок от 2 до 8 мм шириной, который является продолжением перистома и нависает своим нижним острым краем в виде карниза. Воротничок состоит главным образом из губчатой паренхимы, которая образует самую толстую часть стенки кувшинчика. Он покрыт очень своеобразными черепитчато налегающими друг на друга эпидермальными клетками, поверхность которых характеризуется тонкой радиальной штриховатостью. Каждая из этих клеток вытянута в направленный книзу остроконечный отросток. Вместе с когтевидными зубцами перистома эти отростки образуют "задерживающее кольцо", мешающее насекомому выбраться наружу, и "зону скольжения", способствующую его падению внутрь кувшинчика. Расположенная под воротничком внутренняя часть кувшинчика состоит из паренхимных клеток с волнистыми стенками. В этих клетках часто содержится темно-красный пигмент. За исключением узкой полоски, лежащей непосредственно под карнизом воротничка, в верхней половине этой части полости кувшинчика имеются многочисленные небольшие погруженные желёзки, которые по направлению вниз (т. е. к той части кувшинчика, которая расположена ближе к земле) постепенно становятся более крупными. Эти желёзки выделяют протеолитический экзофермент протеазу, т. е. несут чисто пищеварительную функцию. По обе стороны нижней половины этой зоны полости кувшинчика имеется по одной косо расположенной темно-красной выпуклости или валика, содержащей многочисленные крупные погруженные пищеварительные желёзки. Особенно богата желёзками верхняя часть валика. Эти желёзки играют главную роль в переваривании попавших в западню насекомых. Нижняя часть валика, покрытая эпидермой с волнистыми клеточными стенками, снабжена исключительно большим числом устьиц. Эти устьица, однако, необычного типа. Их замыкающие клетки потеряли способность к тургорным движениям, и отверстие устьица все время широко открыто. В сущности это уже не настоящие устьица. Известный немецкий ботаник К. Гёбель (1891), впервые описавший эти своеобразные структуры, назвал их "водными порами", т. е. гидатодами. Весьма вероятно, что нижняя часть кувшинчика наполняется жидкостью именно через эти гидатоды, хотя не все исследователи с этим согласны. Самая нижняя часть внутренней поверхности кувшинчика совершенно лишена желёзок.
Не менее интересно строение крышечки кувшинчика, являющейся немаловажной частью ловчего аппарата. По верхней стороне крышечки проходят однажды или дважды вильчатые радиальные участки зеленой ткани. Эпидерма этих участков состоит из клеток с более или менее волнистыми краями и снабжена волосками. Эта ткань снабжена как привлекательными для насекомых (аттрактивными) погруженными желёзками, так и устьицами. С внутренней стороны крышечки она темно-красная. Промежутки между зелеными участками лишены хлорофилла и устьиц, но с желёзками. В отличие от зеленых участков клетки эпидермы здесь прямые. Насекомым эти почти полупрозрачные участки кажутся открытыми. В своих попытках выбраться из ловушки они, налетая на эти участки, отталкиваются от них и погружаются в полость кувшинчика. Края крышечки волнистые. Черепитчато налегающие друг на друга, эпидермальные клетки внутренней стороны крышечки вытянуты каждая в отросток, который направлен вниз, к основанию крышечки. Эти клетки, подобно эпидермальным клеткам воротничка, с тонкой штриховкой, сходящейся к концу отростка (см. рис. 85). Между эпидермальными клетками находятся аттрактивные желёзки, сходные с желёзками внешней стороны крышечки.
Кувшинчатые листья цефалотуса представляют собой чрезвычайно остроумную западню для насекомых. Три плоских гребня, проходящих вдоль кувшинчика, вероятно, облегчают ползающим насекомым доступ к зеву кувшинчика. Пестрая окраска кувшинчика и обилие желёзок имитируют цветок и служат таким образом приманкой для летающих насекомых. Соблазнившись выделениями этих желёзок, насекомое двигается по направлению к зеву кувшинчика и приближается к его полости, где, как указывает А. Дж. Хамилтон (1904), изучавший биологию цефалотуса в природе, насекомое долгое время лижет поверхность воротничка, прежде чем идти дальше вниз. Попав на внутренюю сторону очень гладкого и скользкого зева урны, оно легко соскальзывает вниз и почти неизбежно становится жертвой цефалотуса. Основными жертвами цефалотуса являются муравьи. Насекомые перевариваются как ферментами, выделяемыми поверхностью кувшинчика, так, вероятно, и бактериями. В урне находят хитиновые остатки насекомых, что говорит о том, что желёзки цефалотуса не выделяют хитиназы.
Семейство крыжовниковые (Gkossulakiaceae) (Н. Н. Цвелев)
К относительно небольшому семейству крыжовниковые принадлежит только один род смородина (Ribes), включающий более 150 видов распространенных в умеренно теплых и субтропических областях северного полушария, а также в горных районах Центральной и Южной Америки до Огненной Земли. Правда, один из подродов этого очень полиморфного рода нередко выделяется в самостоятельный род крыжовник (Grossularia) и действительно имеет ряд особенностей, отличающих принадлежащие к нему виды от других смородин.
Все крыжовниковые - кустарники с очередными пальчатолопастными листьями без прилистников. Как и многие кустарниковые виды других семейств (например, виды шиповника - Rosa), смородины имеют длительно живущие подземные стволики, которые ежегодно дают неразветвленные вегетативные побеги. На следующий год эти побеги ветвятся и становятся репродуктивными побегами. После 4-8 лет жизни они отмирают, так что кусты смородин и крыжовников постоянно обновляются. Ветви некоторых смородин, в особенности принадлежащих к крыжовникам, покрыты шипами - выростами коры, из которых наиболее крупные в числе 1 или 3 (реже 5) располагаются в узлах стебля под основанием листьев. Эти шипы хорошо защищают растение от поедания травоядными животными.
Цветки крыжовниковых обычно небольшие, 4-5-членные и расположенные кистями, у крыжовников редуцированными до 1-3 цветков. Чашелистики у всех видов срастаются основаниями в трубку и часто лепестковидно окрашены; лепестки очень мелкие (нередко в виде чешуек), свободные; тычинки чередуются с лепестками; гинецей паракарпный, из 2 плодолистиков. Некоторые виды, в том числе смородина черная (Ribes nigrum), могут самоопыляться, но преобладают перекрестно опыляющиеся виды. Во многих случаях перекрестному опылению способствует протогиния, особенно хорошо выраженная у смородины красной (Ribes rubrum) и крыжовника обыкновенного (R. uvacrispa, или Grossularia reclinata). Имеются и двудомные виды, к которым принадлежит, например, смородина альпийская (R. alpinum). Опылители большинства видов смородин не специфичны, и ими могут быть различные перепончатокрылые, мухи и жуки. Цветки видов с длинной и узкой трубкой чашечки, например у смородины золотистой (R. aureum), опыляются длиннохоботковыми пчелами. В горных районах Америки имеются виды, опыляемые колибри, например особенно крупноцветковая смородина красивая (R. speciosum) с далеко выступающими из околоцветника нитями тычинок.
Завязь у крыжовниковых нижняя, и образующиеся позднее плоды - различным образом окрашенные ягоды - несут на своей верхушке засохшую чашечку. У большинства собственно смородин между цветками и цветоножкой имеется сочленение, по которому зрелые ягоды легко опадают, а у крыжовников такого сочленения нет и ягоды долго сохраняются на цветоножках. Имеется и другое различие: ягоды типичных смородин почти всегда шаровидные, голые, а ягоды крыжовников обычно имеют широкоэллипсоидальную форму и бывают покрыты щетинками или железистыми волосками. Все виды семейства распространяются с помощью птиц, а отчасти также других животных, поедающих ягоды.
Многие из видов крыжовниковых - широко культивируемые ягодные кустарники, имеющие большое хозяйственное значение. Из них крыжовник обыкновенный представлен множеством сортов, которые делятся на 3 основные группы, различающиеся по цвету ягод (зеленые, желтовато-белые, черные или красные), их форме и величине, а также по присутствию или отсутствию на них щетинок и железистых волосков. Многочисленными сортами представлены и культивируемые смородины: черная, красная, золотистая. Для черной смородины, кроме черных ягод в немногоцветковых кистях, очень характерно присутствие на нижней стороне листьев и других частях растения желтоватых желёзок, выделяющих ароматические вещества. Поэтому листья черной смородины часто используют в качестве ароматизирующего средства при консервации овощей и грибов. Не менее богатая витаминами красная смородина имеет красные, реже желтовато-белые ягоды в многоцветковых кистях. Реже используются красные или черные ягоды североамериканской смородины золотистой, которая чаще разводится в качестве декоративного растения, имеющего золотисто-желтые цветки с приятным запахом. Многие другие дикорастущие виды смородины (некоторые из них встречаются и в культуре) также имеют вполне съедобные, богатые витаминами ягоды. Из декоративных смородин можно отметить еще смородину кроваво-красную (R. sanguineum) с красивыми пурпурными цветками.
Семейство белозоровые (Parnassiaceae) (Г. Л. Кудряшова)
Небольшое семейство белозоровых включает два рода (рис. 87) - белозор (Parnassia), насчитывающий более 50 видов, и монотипный род лепуропеталон (Lepuropetalon). Ареал рода белозор - внетропическая зона северного полушария. Единственный вид рода лепуропеталон - лепуропеталон лопатчатый (L. spathulata) - распространен на юго-востоке Северной Америки, в Мексике и в Чили, где он достигает 30° ю. ш. Северная граница ареала семейства проходит в Арктике (70-80° с. ш.), куда заходит белозор Коцебу (P. kotzebuei).

Рис. 87. Белозоровые. Белозор болотный (Parnassia palustris): 1 - общий вид растения; 2 - цветок (видны тычинки и стаминодии). Белозор альпийский (P. alpicola): 3 - цветок с бахромчатыми лепестками. Лепуропеталон лопатчатый (Lepuropetalon spathulata): 4 - общий вид растения; 5 - отдельный побег; 6 - цветок к концу цветения
Представители семейства - многолетние (белозор) или однолетние (лепуропеталон) травы высотой 2-30 см, с простыми, иногда ветвистыми, многочисленными или одиночными стеблями. Растения, как правило, имеют черешчатые прикорневые листья, образующие розетки, и сидячие стеблевые листья (у некоторых видов листья стеблеобъемлющие) без прилистников. Цветки одиночные на верхушке стебля (белозор) или в пазухах прикорневых листьев (лепуропеталон), актиноморфные или слабозигоморфные, обоеполые. Чашелистиков и лепестков по 5. Цельнокрайние или бахромчатые по краю лепестки обычно длиннее чашелистиков (белозор) или редко короче, недоразвиты или даже отсутствуют (лепуропеталон). Тычинок 5, чередующихся с лепестками и с 5 стаминодиями. Пыльники яйцевидные (белозор) или шаровидные (лепуропеталон). Гинецей паракарпный, из 3 - 4 плодолистиков; столбики очень короткие или отсутствуют, с 3 - 4 свободными комиссуральными рыльцами; завязь верхняя, или полунижняя (белозор), или же нижняя (лепуропеталон), одногнездная, в которой развиваются многочисленные семязачатки на 3 или 4 постенных плацентах. Плод - коробочка, раскрывающаяся 4 створками, со многими мелкими семенами, с цилиндрическим зародышем и очень скудным эндоспермом или без него. Характерным признаком семейства белозоровых являются также вместилища танина в специальных клетках эпидермы листьев.
Главные центры распространения рода белозор - Восточная Азия и Северо-Западная Америка. В Европе, в Средиземноморье и в Западной Азии растет только белозор болотный (Parnassia palustris, рис. 87). Все виды белозора - обитатели влажных лугов или даже болотистых мест низменностей и особенно высокогорий. Встречаясь иногда и на более сухих лужайках в горах, они указывают в этих случаях на присутствие грунтовых вод.
Латинское название белозора - Parnassia - происходит от упоминаемой уже у Диоскорида "парнасской травы" с плющевидными листьями и белыми цветками.
Биология цветения и устройство цветка очень детально изучены у белозора болотного. Уже давно известно, что белозор - насекомоопыляемое растение. Его белые, довольно крупные цветки (диаметром до 1 см) с темными жилками на лепестках хорошо заметны для насекомых. В цветке же особенно привлекательны для насекомых стаминодии. Каждый из них состоит из короткого стебелька, расширяющегося в мясистую пластинку, рассеченную затем на тонкие дланевидные дольки, заканчивающиеся золотисто-желтыми блестящими шариками, очень похожими на капельки меда. Эти чисто морфологические, структуры, называемые в литературе "желёзками", ничего, однако, не выделяют и не имеют запаха. Нектар же в небольшом количестве выделяется в двух маленьких углублениях на верхней стороне пластинки стаминодия. Но именно круглые блестящие желтые "желёзки" привлекают насекомых. Было замечено, что насекомые из семейства сирфидиевых (Syrphidae) сферофория скрипта (Sphaerophoria scripta), питающаяся пыльцой, или сиритта пипиенс (Syritta pipiens), прилетающая на белозор за нектаром, сначала "пробуют" эти более заметные части цветка. Получается, что цветок больше обещает, чем может дать, за что его называют "цветком-обманщиком". Иногда притягательность стаминодиев для насекомых объясняется не только внешним видом, имитирующим капельки меда, но и следами пыльцы или нектара, оставляемыми на них предыдущими насекомыми. Во всяком случае, стаминодии способствуют тому, что насекомые дольше остаются на цветке и больше двигаются, что повышает шансы опыления.
Важным приспособлением для перекрестного опыления у белозора является также дихогамия его цветка, хорошо выраженная протандричность. Рассмотрим детально все фазы цветения. Когда цветок раскрывается, рыльце еще не развито, тычинки очень короткие и прижаты к яйцевидной завязи. Затем в течение пяти дней тычинки начинают вытягиваться, но не одновременно, а по одной в день. В первый день достигает максимальной длины тычинка, противостоящая наружному чашелистику; при этом она загибается внутрь так, что пыльник оказывается как раз над вершиной завязи, прикрывая место будущего рыльца. Когда пыльник раскроется вверх и наружу, пыльца неизбежно попадет на насекомое, которое посетит цветок, но на рыльце, даже если бы оно уже и существовало, пыльца не может попасть, так как оно защищено тыльной частью пыльника. Продержавшись один день в таком положении, тычинка отгибается к периферии, уступая место следующей по очереди, и увядает. Через пять дней все тычинки, повторив по очереди эти движения, оказываются отогнутыми кнаружи цветка, и вскоре пыльники опадают. На шестой день тычинки увядают, рыльце еще не созрело. С седьмого дня начинается развитие крупного четырехлопастного слегка неправильного сидячего рыльца. Такой строгий ритм в развитии цветка белозора послужил поводом для еще одного его названия - "цветок, который сообщает свой возраст". Все эти особенности развития цветка направлены на обеспечение перекрестного опыления насекомыми.
Плодоношение у белозора наступает в конце лета - осенью. Созревшая коробочка растрескивается на верхушке, и при раскачивании стебля из нее вылетают многочисленные мелкие семена, распространяемые в основном ветром.
Второй род семейства белозоровых - лепуропеталон. Его название происходит от греческих слов lepuron - шелуха, скорлупа и petalon - лепесток; оно говорит об очень маленьких лепестках, напоминающих белые чешуйки между толстыми долями чашечки. Лепуропеталон лопатчатый - крошечное мясистое растеньице, высотой не более 2 см, может служить примером одного из самых маленьких цветковых растений. Растет лепуропеталон на влажных песчаных почвах, на кладбищах, вдоль дорог, по краям обрывов. Растение мало изучено биологически. Известно только, что цветет оно ранней весной и что, по-видимому, является самоопылителем, так как замечено, что пыльники растрескиваются, когда они обращены к рыльцу, а в более позднюю стадию завязь увеличивается и отталкивает пыльники в стороны. Семена созревают весной, но прорастание их задерживается до середины зимы, когда состояние покоя нарушается, вероятно, в связи с увеличением длины дня.
Семейство росянковые (Droseraceae) (Г. А. Денисова)
К семейству относят 4 рода, включающие более 100 видов. Росянковые - многолетние корневищные болотные или водные травянистые растения (очень редко - полукустарники), обладающие специальными приспособлениями для ловли насекомых. Листья у них очередные, простые, цельные, обычно собраны в розетки и, как правило, усажены разнообразными железистыми волосками, щетинистыми выростами и чувствительными щетинками.
Цветки у представителей семейства актиноморфные, обоеполые, собраны в простые верхоцветные соцветия. Чашелистики более или менее сросшиеся у основания, лепестки свободные. Тычинок чаще всего 5, свободные или редко сросшиеся у основания. Гинецей обычно паракарпный из 3 плодолистиков или лизикарпный из 5 плодолистиков, обычно со свободными столбиками. Завязь верхняя, с многочисленными или реже несколькими семязачатками. Плод - коробочка. Семена с маслянистым эндоспермом.
Три монотипных рода росянковых - росолист (Drosophyllum), венерина мухоловка (Dionaea.) и альдрованда (Aldrovanda) - имеют более или менее узкое распространение. Самый же крупный род семейства - росянка (Droseга), насчитывающий до 100 видов, является космополитом. Представители этого рода встречаются почти во всех климатических поясах обоих полушарий. Особенно много видов росянок произрастает в Австралии и в Новой Зеландии.
Росянки - многолетние, как правило, болотные травы с коротким ползучим или клубневидным корневищем и розеткой прикорневых листьев, покрытых красноватыми железистыми ловчими волосками-щупальцами с красной железистой головкой, выделяющей липкий секрет.
Мелкие цветки собраны в конечные негустые кистевидные или метельчатые соцветия; не сросшиеся белые лепестки цветка росянки при основании вытянуты в желтоватый ноготок. Шаровидная завязь несет 3 столбика, иногда 5, каждый из которых разделен наверху на два лопатчатых рыльца, в зрелом состоянии покрытых чрезвычайно липкими выделениями. Когда цветок раскрыт, рыльца в нем размещены горизонтально, а тычинки, наоборот, подняты вертикально, перекрещиваясь с рыльцами под прямым углом, и пыльники таким образом оказываются над рыльцами. При закрывании цветка пыльца из пыльников высыпается на рыльца, отчего происходит самоопыление. Чашечка цветка трубчатоколокольчатая, немного короче лепестков. Плод - коробочка, открывающаяся 3-5 створками, одетая остающимися чашечкой и венчиком. Семена многочисленные, светло-бурые, почти гладкие.
В СССР произрастают 4 вида росянок, из которых наиболее широко распространена росянка круглолистная (D. rotundifolia, табл. 24). Это чрезвычайно мелкое растение особенно часто встречается в северных и центральных районах нашей страны, в основном по торфяным болотам. Округлые листья собраны в прикорневую розетку. Вся верхняя сторона и края каждого листа усажены волосками-щупальцами с красной железистой головкой. В центре листа железистые волоски короткие, а по краям более длинные. Головку волоска окружает прозрачная капелька густой липкой тягучей слизи. Мелкие мухи или муравьи, привлеченные блеском этих капелек, садятся или вползают на лист и прилипают к нему. Насекомое мечется и бьется, пытаясь освободиться из ловушки, и при этом неизбежно задевает за соседние липкие капли. Все волоски потревоженного листа изгибаются навстречу добыче и скоро обволакивают ее слизью. Раньше считалось, что насекомые просто прилипают к этой слизи, но недавно ученым удалось выделить из сока росянки два вещества (одно из них - алкалоид кониин), относящиеся к классу аминов, которые оказывают парализующее действие на насекомых. Край листа медленно загибается и покрывает свою добычу, которая здесь же вскоре начинает перевариваться. Этот процесс возможен благодаря тому, что слизь железистых волосков содержит пищеварительные ферменты, напоминающие по составу пищеварительный сок животных. Как указывает английская исследовательница И. Xеслоп-Харрисон (1976), биохимическими исследованиями определены следующие ферменты, вырабатываемые и выделяемые желёзками насекомоядных растений: пероксидаза, альбонуклеаза, липаза, эстераза, кислая фосфатаза, амилаза. В слизи также обнаружены кислые полисахариды, состоящие из ксилозы, маннозы, галактозы и клюкуроновой кислоты. Под действием такого сложного секрета за сравнительно короткое время насекомые разлагаются и постепенно всасываются теми же желёзками внутрь растения.

Таблица 24. Росянковые: 1 - росянка нитевидная, разновидность Трейса (Drosera filiformis var. tracyi), Флорида; 2 - верхняя часть молодого листа росянки нитевидной, там же; 3 - росянка круглолистная (D. rotundifolia), Ленинградская область; 4 - венерина мухоловка (Dionaea muscipula), оранжереи Ботанического института АН СССР в Ленинграде
Какой же фактор приводит железистые волоски листа в движение Профессор Н. Г. Xолодный (1948) считает, что такая реакция наблюдается в тех случаях, когда вслед за механическим раздражением листа следует его химическое раздражение, вызываемое веществами, диффундирующими из тела пойманного насекомого. Оказывается, лист росянки не только чувствует груз упавшего на него тела, но даже воспринимает его запахи.
Поскольку способность питаться животной пищей выработалась у росянковых как своеобразное приспособление к исключительно обедненному субстрату, растение извлекает из пойманных насекомых прежде всего соли натрия, калия, магния, фосфор и азот.
Через несколько дней после поимки насекомого лист постепенно раскрывается. На железистых волосках снова выступают прозрачные капельки слизи. Ловушка вновь приведена в боевую готовность.
Если же на лист росянки упадет кусок сухой травинки или еще что-нибудь несъедобное, волоски-щупальца только чуть-чуть дрогнут и снова останутся неподвижными. Ч. Дарвин отмечал, что падение на лист даже крупных тяжелых капель дождя не вызывает движения волосков. Однако капля слабого раствоpa аминокислот или аммиака является для листа сильным раздражителем.
Цветение росянки круглолистной может продолжаться в течение всего лета. Белые мелкие цветки этого энтомофильного растения поднимаются над розеткой листьев на длинном цветоносе. Осенью вызревают плоды-коробочки, заполненные семенами, у которых между эндоспермом и семенной оболочкой имеется воздушная полость; легкие, надутые воздухом семена росянки, не превышающие в поперечнике 0,2 мм, распространяются на большие расстояния в основном с водой, так как способны долго плавать на ее поверхности, не теряя всхожести.
Многие виды росянок, произрастающих в тропических странах, крупнее своих северных сородичей и могут ловить более крупных насекомых. Длинные волоски этих видов, окружающие центр листа, торчат во все стороны, как щупальца гидры. Они особенно быстро реагируют на любое, самое слабое прикосновение. В Австралии, например, произрастает росянка гигантская (D. gigantea), вполне оправдывающая свое название. Ее гибкий крепкий восходящий или цепляющийся стебель достигает в длину 60-100 см. Нижние стеблевые листья редуцированы до жестких шиловидных чешуек. По описаниям В. И. Талиева (1903) и Ф. Ллойда (1942), это своеобразное растение со своими растопыренными ветвями, усаженными длинночерешковыми ловчими листьями, подобно паутине, обладает обширной улавливающей поверхностью.
Близкий к росянкам полукустарник росолист лузитанский или "португальская мухоловка" (Drosophyllum lusitanicum), встречается в Испании, Португалии и Марокко, преимущественно на сухих каменистых почвах, обладая хорошо развитой корневой системой. От короткого, прямостоячего, в нижней части одревесневшего стебля отходят в сторону и вверх до двух десятков длинных узких линейных листьев, желобчатых сверху и выпуклых снизу. Их верхняя поверхность и края густо усыпаны ловчими желёзками двух форм: сидячими и на ножках. Последние постоянно выделяют чрезвычайно липкую густую слизь, содержащую кислые полисахариды, к которой насекомые, даже крупные, накрепко приклеиваются; при этом и железистые волоски, и сами листья остаются неподвижными.
Выделение пищеварительных ферментов производят только сидячие желёзки, и лишь после того, как они получат раздражение от движения пойманного насекомого; кислое содержимое железистых волосков, попадая на сидячие желёзки, усиливает их секреторную функцию. Как указывает Н. Г. Холодный (1948), переваривающая способность росолиста чрезвычайно велика: в течение дня одно растение средней величины успешно справляется с добычей, состоящей из нескольких десятков крупных мух и других насекомых. Поглощение продуктов распада осуществляется, вероятно, желёзками обеих форм. По данным И. Хеслоп-Харрисон (1978), секреторные желёзки всегда находятся в более или менее прямой связи с сосудами, питающими лист. В головку желёзки проходит пучок трахеид, связанный с проводящими сосудами ножки желёзки, которые, в свою очередь, входят в сосудистую систему листа.
Относящаяся к семейству росянковых венерина мухоловка (Dionaea muscipula, табл. 24) является эндемиком прибрежной части штата Северная Каролина в США. Она приурочена к скудным песчаным почвам с высоким уровнем грунтовых вод и нуждается в интенсивном освещении. Крупные белые цветки собраны в конечные малоцветковые соцветия на верхушке цветоноса. Плод - неравномерно растрескивающаяся коробочка, заполненная двумя десятками блестящих черных семян и окруженная увядшими лепестками. Хорошо развитые экземпляры венериной мухоловки могут переносить без вреда для себя как засуху, так и временное затопление. Листья этого растения, чуть приподнятые над землей, собраны розеткой вокруг длинного цветоноса (рис. 88). Черешок листа плоский и широкий, а пластинка листа превратилась в две округлые створки, лежащие под углом друг к другу; они снабжены по краям длинными крепкими зубцами. На верхней поверхности обеих половинок листа торчат вверх по три чувствительных волоска. Лишь только насекомое коснется одного из волосков, створки мухоловки стремительно захлопываются. При этом зубцы их заходят друг за друга, образуя подобие решетки. Если добыча мала, она может выползти из ловушки, но если внутри створок оказалось насекомое толще 3-4 мм, оно неизбежно погибнет. Чем отчаянней бьется оно в западне, тем крепче сжимаются створки листа, все сильнее прилегая одна к другой и сдавливая добычу. Верхняя (т. е. внутренняя) поверхность листа усеяна мелкими красными желёзками, выделяющими жидкость, которая содержит пищеварительные ферменты и муравьиную кислоту.
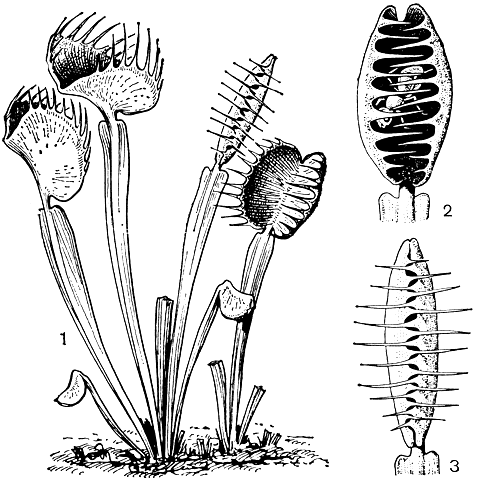
Рис. 88. Венерина мухоловка (Dionaea muscipula): 1 - общий вид растения; 2 - полузакрытый лист; 3 - закрытый лист
Е. В. Граве (1958) дает обзор гипотез, пытающихся объяснить механизм столь редко встречающегося в природе явления - мгновенного закрывания ловчего аппарата. Наиболее вероятной из них кажется гипотеза о распространении двигательного стимула путем передачи электрической энергии; в листе венериной мухоловки при раздражении происходят электрофизиологические явления, напоминающие процессы, которые наблюдаются при передаче возбуждения в нервно-мышечном аппарате животных. По представлениям других ученых, вещества, извлекаемые желёзками из тела насекомого, являются источником химического раздражения, вызывающего движение листа.
Как указывают Ф. Т. Лихтнер и С. Е. Вильяме (1977), а также И. Хеслоп-Харрисон (1978), тесно захлопнувшиеся ловушки растения удерживаются в таком состоянии довольно долго, обычно не менее 40 ч. При этом муравьи, составляющие в природе примерно одну треть всей добычи, остаются живыми в течение 8 ч и лишь после выделения желёзками кислой жидкости погибают. Причиной сохранения створок листа закрытыми авторы считают механическое раздражение живыми муравьями чувствительных волосков. Когда начинается переваривание насекомых, в растворе появляются ионы натрия, свободные аминокислоты и аммиак, являющиеся химическим стимулом, который поддерживает лист в закрытом состоянии.
По истечении определенного срока лист венериной мухоловки снова раскрывается, желёзки на его поверхности восстанавливаются и готовы к повторному функционированию.
Единственным водным растением семейства росянковых является альдрованда пузырчатая (A. vesiculosa), спорадически распространенная во внутриконтинентальных прибрежных водах всех климатических поясов Земли, исключая самые северные районы. У нас в стране это миниатюрное растеньице можно встретить и в центральных районах европейской части, и на Кавказе, и в Средней Азии, и на Дальнем Востоке.
Совсем не имея корней, альдрованда свободно плавает в водоеме. Ее тонкий, слабоветвистый травянистый стебель с мутовками из 7-9 листьев, похожих на листья венериной мухоловки, всегда находится под водой. Широкие плоские листообразные черешки листьев сужены возле листовой пластинки, где оканчиваются длинными шиловидными щетинками, которые, как пики, торчат во все стороны.
Листовая пластинка состоит из двух полукруглых, наклоненных одна к другой половинок. Это ловчий аппарат растения. Край листовой пластинки, усаженный заостренными одноклеточными волосками, загнут внутрь. Ближе к центру располагаются четырехлопастные крестовидные желёзки, которые выделяют слизь. В центральной части листовой пластинки, возле главной жилки, имеется множество сидячих округлых пищеварительных желёзок, а также большое количество чувствительных волосков. Добычей альдрованы служат рачки, туфельки и другие микроскопические обитатели водоемов. Задевая чувствительные волоски листа, они вызывают его закрывание; чем лист моложе, тем он быстрее закрывается. Поэтому ловят животных главным образом молодые листья двух - четырех верхних мутовок. Животные, попавшие в западню, некоторое время продолжают там плавать, но лист смыкается все плотнее и плотнее, пока его плоские края не соприкоснутся. При этом в центральной части листа образуется пузыревидная полость, из которой вода постепенно исчезает, заменяясь воздухом. Как предполагает Н. Г. Холодный (1948), исчезновение воды в пузырьках объясняется всасывающей способностью всех желёзок, а появление воздуха там, вероятно, связано с процессом фотосинтеза в окружающих зеленых клетках листа, которые на свету выделяют кислород. Дольше всего вода сохраняется в углублениях возле жилки, в месте скопления пищеварительных желёзок, где и собираются все пойманные животные. Фабиан, Галан и Салагену (1968) обнаружили, что у альдрованды животная пища усваивается только в зрелых листьях-ловушках, откуда питательные вещества переносятся прямо к точкам роста растения, минуя фотосинтезирующие участки листьев. Незрелые ловушки пищи не усваивают. Многие листья, переварив свой первый: улов, вскоре после этого отмирают. Но эту убыль вполне компенсирует быстрое образование новых листьев.
В мелких озерках стариц на Волге или Амударье можно наблюдать, как плавающие у поверхности воды особи альдрованды выставляют на воздух свои мелкие одиночные цветки. Пригретые и обсушенные солнцем, цветки раскрываются, освобождая крупные пестики с пятью торчащими столбиками и сердцевидные пыльники, заполненные зрелой пыльцой. Альдрованда - перекрестноопыляемое энтомофильное растение. Поскольку насекомых над водой бывает не очень много, цветки ее часто остаются неопыленными и процент завязавшихся семян очень низок. Поэтому альдрованда размножается главным образом вегетативно. С наступлением осенних холодов на концах ее веточек образуются зимующие почки, состоящие из плотно прижатых друг к другу зачатков листьев. Отделившись от растения, эти почки падают на дно и там перезимовывают. Весной из них развиваются новые молоденькие растения, которые легче воды, а поэтому всплывают наверх. Легкие цепкие стебли альдрованды без труда пристают к телу водоплавающих птиц, которые могут перенести их в другой водоем и там стряхнуть в воду. Таким образом растение получает новое жизненное пространство для своего распространения. По мнению академика А. Л. Тахтаджяна (1966), семейство росянковых ближе всего стоит к семейству белозоровых (Parnassiaceae) и к семейству непентовых (Nepenthaceae).
Семейство гуннеровые (Gunnebaceae) (А. Е. Бородина)
В семействе гуннеровых - один род гуннера (Gunnera), названный в честь норвежского ботаника Эрнста Гуннера (1718-1773). Этот род включает около 50 видов, распространенных в Южной и Центральной Америке, в Южной и Юго-Восточной Африке, на Мадагаскаре, в Тасмании, Новой Зеландии, на острове Новая Гвинея и на Гавайских островах. Обычно гуннеры - гигантские многолетние травы, растущие в сырых горных лесах и внешне напоминающие ревени. Только некоторые виды (например, в Новой Зеландии и на Огненной Земле) маленькие дернинообразующие растения. Листья у гуннер очередные, прикорневые, округлые или почковидные, иногда с лопастями или зубцами по краю, часто огромные (в поперечнике до 3 м), на черешках и часто снабжены чешуями.
На нижней поверхности листьев и особенно на погруженной в воду части черешков часто можно встретить колонии сине-зеленых водорослей ностока. Цветки очень мелкие, в крупных (до 2 м) безлистных ярко окрашенных метелках, обоеполые или однополые (нижние - женские, верхние - мужские). Чашелистиков 2, лепестков 2 или нет совсем, тычинки 2 с короткими нитями, гинецей состоит из 2 плодолистиков, завязь одногнездная с 1 семязачатком. Сочные, костянковидные, оранжевые или красные плоды, содержащие семена с обильным эндоспермом, охотно поедают рыбы и птицы.
В садах и парках Западной и Южной Европы около прудов высаживают одиночные крупные растения гуннеры шероховатой (G. scabra). На родине - это растение-великан. Рассказывают, что под одним ее листом от дождя могут укрыться сразу 3 всадника. В Европе гуннера не достигает таких исполинских размеров, однако экземпляры, имеющие в поперечнике 6 м, не являются большой редкостью. Корневище гуннеры используют для получения танинов (до 9% сухой массы), отвары корневищ африканской гуннеры точной (G. perpensa) служат для успокоения детей. Черешки листьев некоторых гуннер используют для приготовления овощных блюд.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'