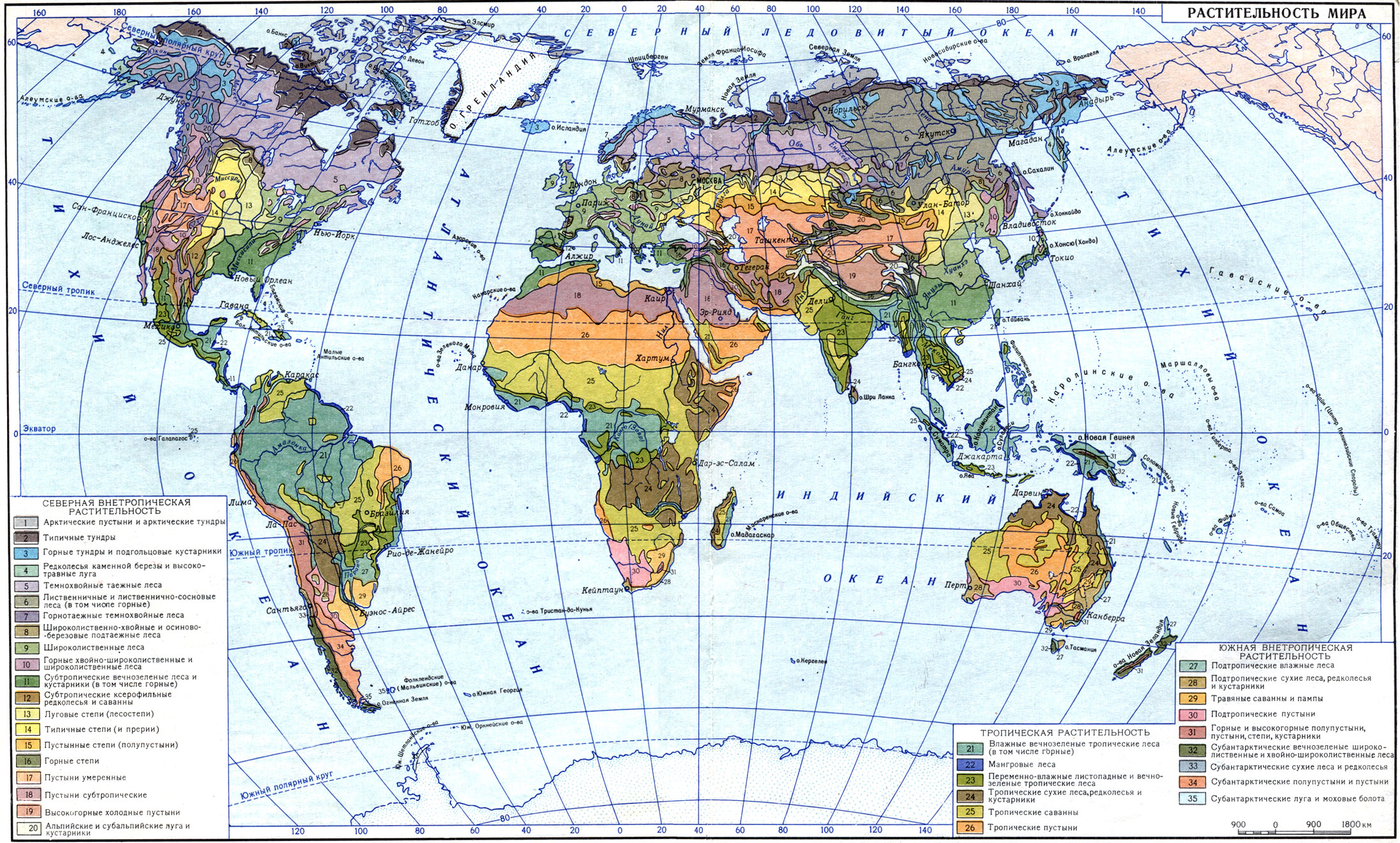
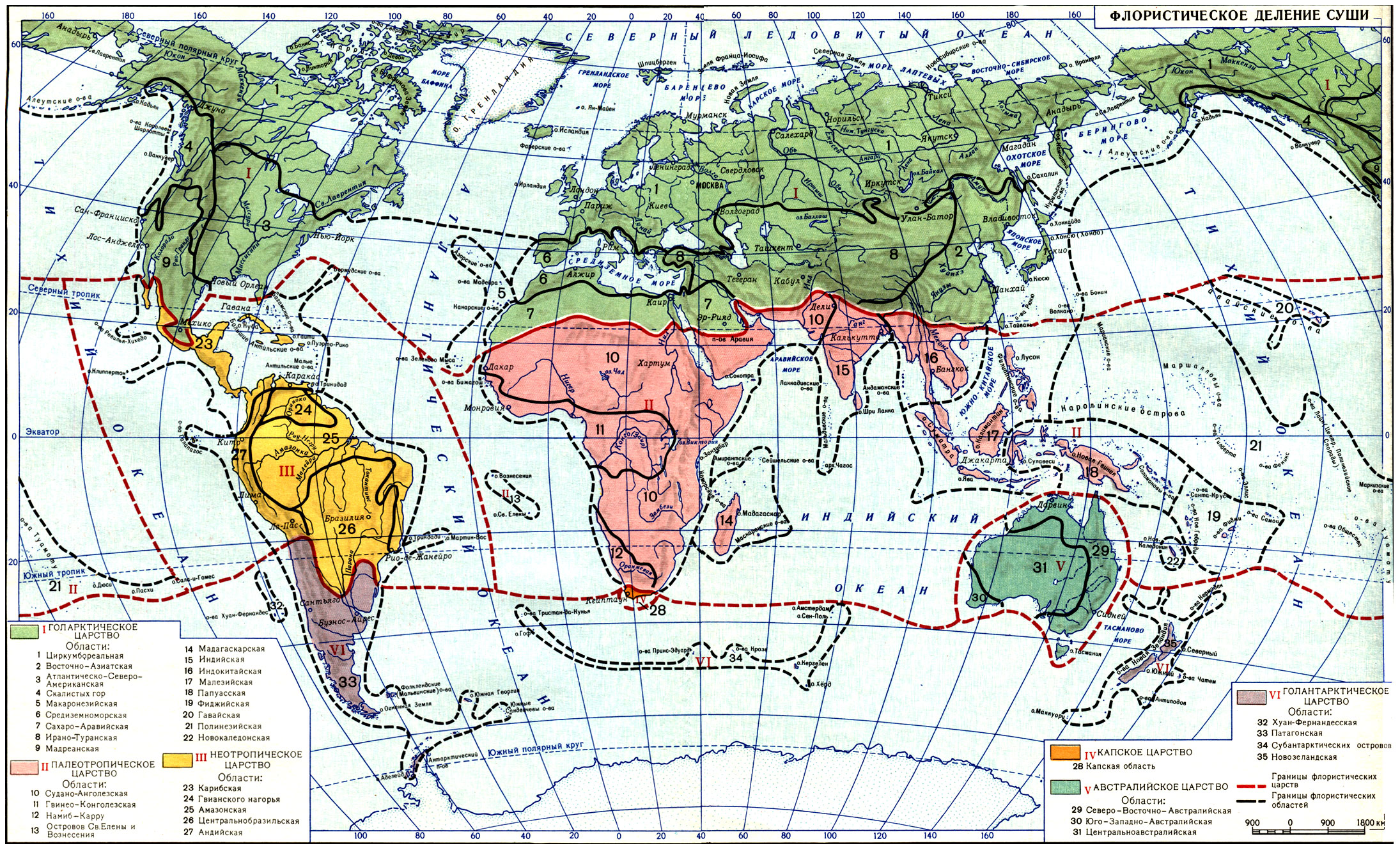
Жизненные формы растении (Т. И. Серебрякова)
Когда мы путешествуем и попадаем в края с непривычными, экзотическими для нас ландшафтами, поневоле в первую очередь обращаем внимание на общий облик, или габитус, растений, создающих этот ландшафт. Житель умеренной лесной полосы в тундре непременно обратит внимание на низкорослые, иногда ползучие (шпалерные) кустарники и кустарнички - полярные виды березы и ивы, толокнянку (табл. 14), водянику; в высокогорьях Памира будет с любопытством рассматривать "растения-подушки" - терескен, акантолимон (табл. 19); в целинных казахстанских степях - крупные плотные дерновины ковылей.
В тропических странах ему бросятся в глаза пальмы, и он отметит, что крона их состоит не из многочисленных толстых и тонких ветвей, покрытых на концах довольно мелкими листьями, как у наших привычных деревьев, а из огромных перистых или пальчатых листьев, собранных на верхушке ствола в пучок или розетку. У мадагаскарского "дерева путешественников" (Ravenala), напоминающего пальму (табл. 19), нашего туриста поразит особое расположение листьев - веерное, в одной плоскости, а у родственного ему банана - форма "ствола", от самого подножия окутанного длинными трубчатыми основаниями листьев. Оказывается, что "ствол" этот в действительности ложный, а само растение - не дерево, а гигантская трава с подземным клубневидным многолетним стеблем. В мексиканских пустынях мы будем удивляться огромным безлистным кактусам с колоннообразными сочными стволами, а в высокогорьях Анд, где-нибудь в парамосах Колумбии, - высоким (5 - 6 м) фигурам эспелеций (Espeletia) - своеобразнейших древовидных сложноцветных, которые местные жители называют "монашками". Столь же причудливы древовидные крестовники (Senecio), тоже из сложноцветных (в горах тропической Африки).
Все необычные для нас формы растений мы поневоле будем сравнивать с обычными для нас, господствующими в умеренном ландшафте, - лесными деревьями, кустарниками, травами.
Первым обратил внимание на ландшафтную роль "основных форм" растений крупнейший немецкий ученый-путешественник, "отец географии растений" А. Гумбольдт (1806). Он разделил все растения на 16 "форм", названия которых часто совпадают с наименованиями крупных систематических групп ("форма пальм", "форма мимозовых", "форма алоэ"), однако он имел в виду не родственные отношения, а конвергентное сходство в облике. Он отметил, например, что к "форме алоэ" надо отнести не только виды рода Aloё и некоторые другие лилейные, но и многие бромелиевые (например, ананас), имеющие такие же сочные заостренные листья, скученные на верхушке стеблей, а к "форме казуарин" - не только своеобразные австралийские деревья казуарины с безлистными зелеными веточками, но и такие же безлистные крупные африканские хвощи, эфедры, среднеазиатский джузгун (Calligonum) и т. д. К "форме кактусов" он отнес также поразительно с ними сходные по внешнему виду африканские молочаи.
Естественно, в создании ландшафта, "физиономии" растительного покрова, той или иной страны играют решающую роль формы надземных органов растений: величина и ветвление стволов, формы крон, направление роста стеблей, размеры и формы листьев и т. п. Но это лишь часть габитуальных признаков.
Скрыты от непосредственного наблюдения, но не в меньшей степени важны для характеристики общего облика растения его подземные органы. Разгребая лесную подстилку, копая яму или пропалывая грядки, мы непременно заметим, что у ландыша и пырея отдельные побеги, которые нам без раскопки казались самостоятельными индивидами, под землей соединены длинными ветвящимися горизонтальными корневищами, покрытыми придаточными корнями; у люцерны и горного клевера-бело-головки (Trifolium montanum) вглубь уходит длинный и толстый стержневой корень; у цветущей ранней весной хохлатки (Corydalis halleri) под землей круглый желтый клубень, а у одного из сорных видов лука (Allium rotundum) интересная луковица, похожая на луковицу культурного чеснока. Все эти подземные органы дополняют картину общего облика, особенно у травянистых растений, где надземные части, ежегодно отмирающие к осени, на первый взгляд кажутся однообразными. По подземным частям можно судить о способах перезимовки, вегетативного возобновления и размножения растений.
Таким образом, габитус растений определяется формой и величиной их вегетативных надземных и подземных органов, составляющих в совокупности систему побегов и корневую систему. Часть побегов и корней или даже все они могут быть значительно видоизменены.
Именно вегетативные органы обеспечивают питание, рост, всю индивидуальную жизнь растения. Они постоянны и необходимы, тогда как органы репродуктивные - соцветия, цветки, плоды, семена, шишки, спорангии - могут в некоторых случаях вообще не появиться на растении, а если появляются, то в большинстве случаев заметно не влияют на габитус, тем более что существование их временно.
Форма побеговой и корневой систем - результат роста. Поэтому часто в ботанической литературе употребляется термин "форма роста" как синоним общего габитуса растения. Но не менее часто встречаются и другие понятия - "жизненная форма", или "биоморфа", которые не вполне равноценны "форме роста" и габитусу.
Термин "жизненная форма" был введен в 80-х годах прошлого века известным датским ботаником Е. Вармингом - одним из основоположников экологии растений. Варминг понимал под жизненной формой "форму, в которой вегетативное тело растения (индивида) находится в гармонии с внешней средой в течение всей его жизни, от колыбели до гроба, от семени до отмирания". Это очень глубокое и емкое определение.
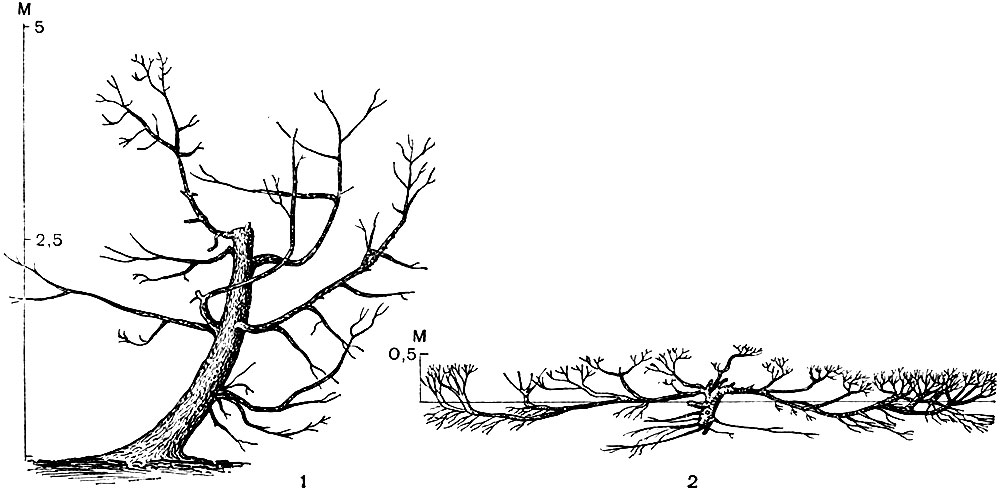
Рис. 58. Древовидная (слева) и стланцевая (сярава) форма туркестанского можжевельника на склонах хребта Терскей-Алатау: 1 - на высоте 2900 м над уровнем моря (лугово-лесной пояс); 2 - на высоте 3200 м над уровнем моря (субальпийский пояс)
Во-первых, оно подчеркивает, что жизненная форма, т. е. форма вегетативного тела, индивида не остается постоянной, а меняется во времени по мере взросления и старения растения.

Таблица 17. Жизненные формы: вверху слева - бутылочные деревья (Австралия) рода Brachychiton; вверху справа - древовидный крестовик; внизу - вельвичия (пустыня Намиб)
В самом деле, растение в течение всей своей жизни растет, увеличивается в размерах, по мере возможности захватывает новое пространство над землей и под землей, образует новые побеги, корни, теряет более старые части, иногда вегетативно размножается и перестает быть единым индивидом. Однолетний сеянец дуба еще ничем не напоминает могучее дерево с толстым стволом и ветвистой кроной, а пневая поросль, которая вырастает после того, как дуб срубили, имеет облик, не похожий ни на сеянец, ни на дерево, хотя все это разные этапы жизни одной и той же особи.

Таблица 18. Жизненные формы: вверху слева - мангровые заросли (видны ходульные корни); вверху справа - многоствольное дерево
Во-вторых, из определения явствует, что важнейшую роль в становлении жизненной формы, во всех ее изменениях играет внешняя среда. И в самом деле, сеянец дуба, развивающийся в питомнике при хорошем освещении и питании, очень скоро становится настоящим деревцем с главным стволом, а сеянец, выросший в лесу, в густой тени, долго остается низкорослым корявым "торчком" (лесоводственный термин) без явного главного стволика. Взрослая особь обыкновенной ели в лесной зоне имеет привычный для нас облик стройного дерева с острой верхушкой, а на Крайнем Севере, на пределе своего распространения, она растет горизонтально, образуя плотно прижатый к субстрату стланец.

Таблица 19. Жизненные формы: вверху слева - подушка акантолимона на Памире; вверху справа - одна из этих подушек; внизу слева - 'дерево путешественников'; внизу справа - древовидные кактусы
Но гармония с внешней средой не означает, конечно, что жизненная форма любого растения бесконечно пластична и зависит только от непосредственно действующих на него в данный момент условий. Каждый вид растений реагирует на внешние воздействия в рамках своих наследственно закрепленных возможностей, запрограммированных генетическим кодом. У особей каждого вида проявляется своя "норма реакции", не позволяющая, допустим, землянике стать развесистым деревом даже в самой благоприятной для роста и ветвления обстановке. И далеко не всякий вид дерева может в крайних условиях существования приобрести стелющуюся форму, как это бывает у ели и туркестанского можжевельника (рис. 58). А главное, говоря о гармонии с внешней средой, мы подразумеваем, что во всем ходе формообразования, особенно в уже сложившейся типичной жизненной форме взрослой особи данного вида, проявляются черты наследственной, исторически выработавшейся в процессе естественного отбора приспособленности растения к тому комплексу внешних факторов, который господствует в области его распространения.
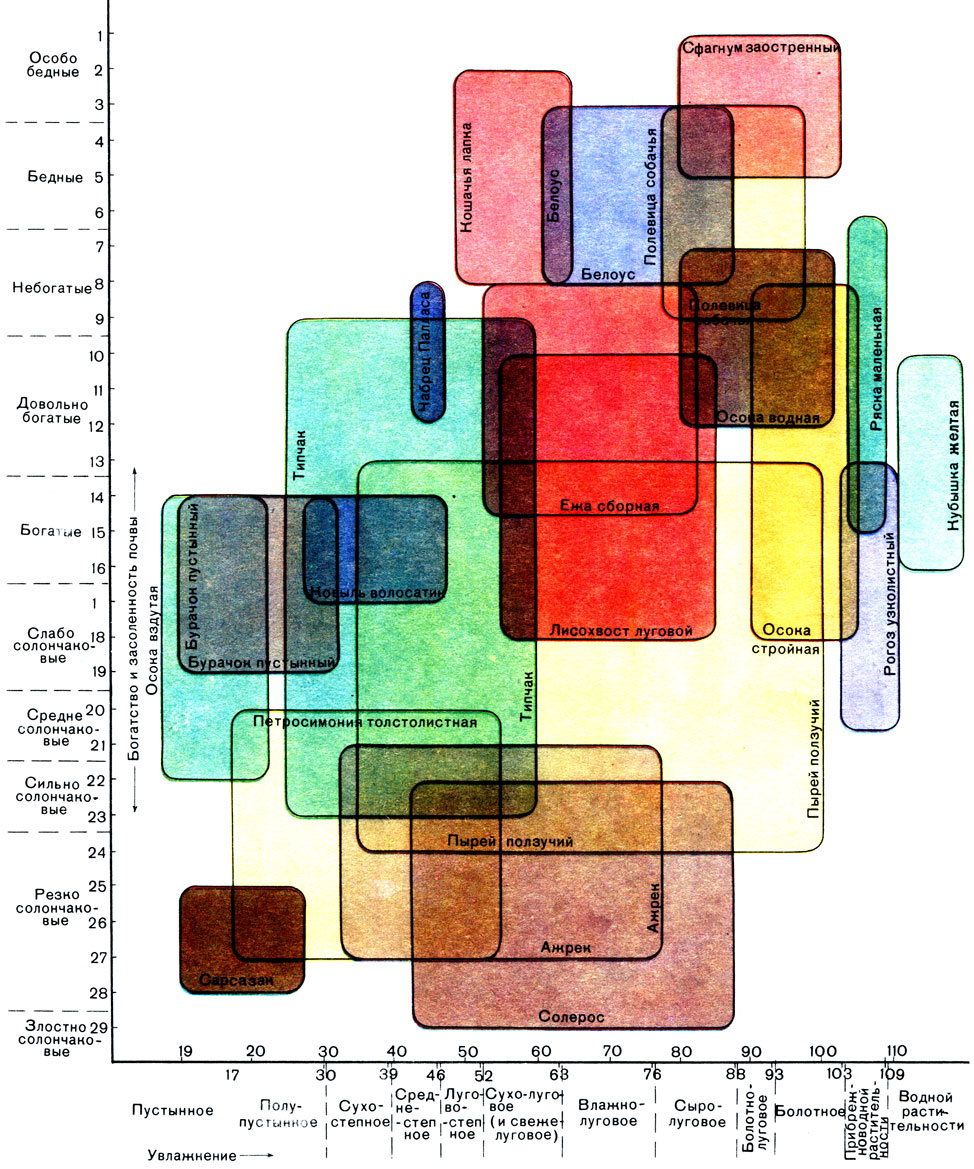
Таблица 20. Распределение некоторых видов растений в зависимости от увлажнения и богатства почв (схема)
В понятие "жизненная форма" со времен Варминга непременно вкладывается представление о соответствии ее структурных особенностей условиям жизни, об адаптивном, приспособительном значении тех или иных габитуальных признаков, используемых для характеристики жизненной формы.
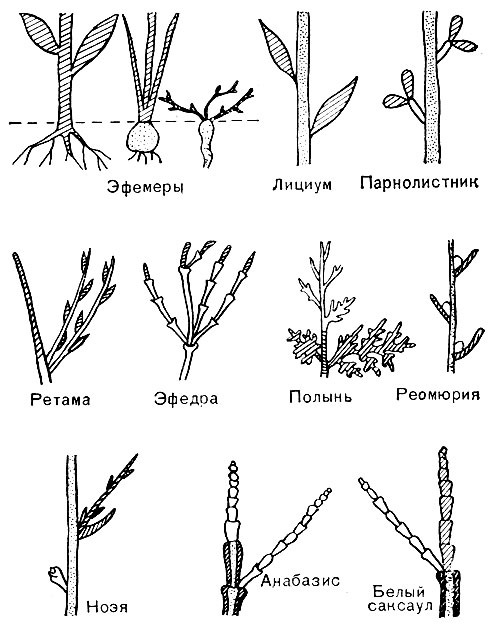
Рис. 59. Побеги разных пустынных растений Ближнего Востока (схема). Заштрихованы ассимилирующие части, которые при недостатке влаги опадают (белые - остающиеся части). Точками заполнены многолетние неассимилирующие части
И. Г. Серебряков называет жизненной формой своеобразный габитус определенных групп растений, возникающий в онтогенезе в результате роста и развития в определенных условиях среды и исторически сложившийся в данных почвенно-климатических и ценоти-ческих условиях как выражение приспособленности к этим условиям.
Е. М. Лавренко, предпочитающий термину "жизненная форма" термин "экобиоморфа", подчеркивает, что экобиоморфы являются "как бы типовыми адаптационными организменными системами, существующими в определенных условиях среды".
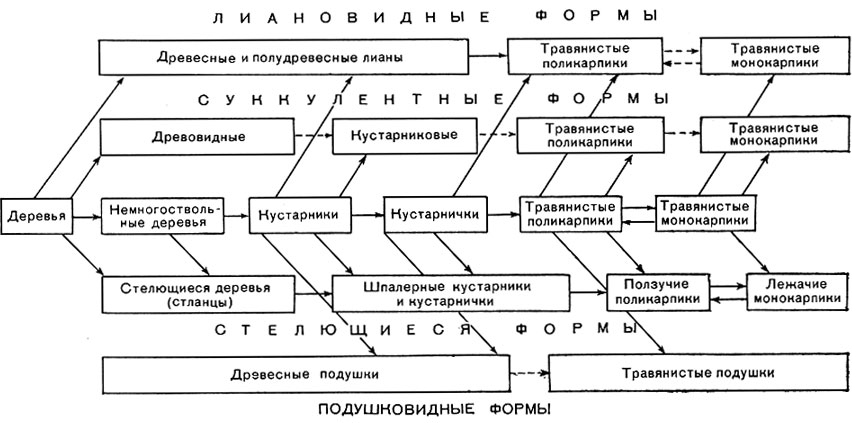
Взаимосвязь жизненной формы со средой и ее приспособительный смысл можно иллюстрировать, скажем, на примере лиан или лазающих растений. Лианы характерны главным образом для влажнотропического леса и там весьма разнообразны; жизненную форму лианы приобретают многочисленные совсем не родственные между собой виды. В условиях высокой влажности почвы и особенно воздуха, при обильном почвенном питании и обилии тепла, но при сильном затенении под пологом древесных крон побеги лиан сначала очень быстро растут в длину. При этом они несут черты этиоляции: стебли сильно вытягиваются, но остаются слабыми, листья на них недоразвитые. Слабые стебли вынуждены опираться на соседние растения, часто они обвивают их благодаря круговым движениям верхушечных почек. Только впоследствии ткани стеблей деревянистых лиан укрепляются механическими тканями, сохраняя, однако, большую гибкость. В результате такого способа роста, тесно связанного с внешними условиями, растения быстро достигают верхних ярусов леса, где их побеги оказываются уже в условиях благоприятного освещения, не несут признаков этиолирования, развивают нормальные зеленые листья, цветки и плоды.
Так лиановидная форма роста - один из возможных путей приспособления растений к жизни во влажнотропическом лесу - оказывается биологически выгодной жизненной формой, помогающей "преодолеть" недостаток солнечного освещения.
Полегание и горизонтальный рост побегов арктических кустарников и кустарничков обусловлены комплексом климатических и почвенных условий тундры: повышенной влажностью субстрата в сочетании с низкими температурами воздуха и почвы, недостатком минерального питания и т. д. Но шпалерная жизненная форма в этих условиях оказывается приспособительной, биологически выгодной, увеличивая стойкость растений при постоянных холодных и иссушающих ветрах, во время суровой малоснежной зимы.
Плотнодерновинный облик степных злаков способствует сохранению влаги в дернине во время летней засухи. Но в тех же засушливых условиях не менее биологически выгодной оказывается и луковичная жизненная форма (например, у степных тюльпанов), когда влага сохраняется в подземных водозапасающих органах луковицы, надземные же побеги во время засухи отмирают: растение "убегает" от засухи.
Последние два примера показывают, что структурные приспособления к одним и тем же условиям могут быть совсем неодинаковыми у разных видов растений. Подтверждение этому можно видеть и на рисунке 59, где схематически показаны разные приспособления к перенесению сухого времени года у растений пустынь Ближнего Востока. Видно, что одни растения при наступлении засухи погибают целиком, оставляя лишь жаростойкие семена (однолетние эфемеры); другие (эфемероиды) "убегают" от засухи, сохраняя только подземные многолетние органы (корни, луковицы); третьи летом частично или полностью сбрасывают листья, части листьев, целые зеленые безлистные веточки и даже зеленую ассимилирующую кору, сохраняя над землей надежно защищенные покровными тканями системы многолетних стеблей с почками возобновления. Все эти признаки габитуальные, определяющие жизненную форму растений, и притом динамические, отражающие сезонную изменчивость жизненной формы каждой особи. На этих же примерах можно показать разницу между жизненной формой и экологической группой.
Все обсуждаемые засухоустойчивые растения по их отношению к влаге можно отнести к экологической группе ксерофитов, но по структурным особенностям и всему внешнему облику, т. е. по жизненной форме, они неодинаковы.
Таким образом, жизненные формы как типы приспособительных структур демонстрируют, с одной стороны, разнообразие путей приспособления разных видов растений даже к одним и тем же условиям, а с другой - возможность сходства этих путей у растений совершенно неродственных, принадлежащих к разным видам, родам, семействам. Поэтому классификация жизненных форм - а при большом разнообразии без нее не обойтись - не может совпадать с обычной классификацией систематиков, основанной на строении репродуктивных органов и отражающей общность происхождения, "кровное" родство растений. Классификация жизненных форм основана на структуре вегетативных органов и отражает параллельные и конвергентные пути экологической эволюции.
Итак, каждая особь в течение своей жизни постоянно меняет жизненную форму. Но жизненная форма как классификационная единица, объединяющая группы сходных по облику растений, должна быть более определенной и ограниченной. Обычно, говоря о типичных жизненных формах того или иного вида какого-либо фитоценоза, имеют в виду жизненные формы взрослых, нормально развитых особей. Признаки, на которых строится классификация, разнообразны и разномасштабны. Гумбольдт, например, подметил наиболее броские физиономические черты, особо не выделяя приспособительное их значение. Варминг отмечал, что для характеристики жизненных форм нужны очень многие биолого-морфологические признаки, в том числе отражающие поведение растения и его органов во времени: общая длительность жизни особи, длительность жизни отдельных побегов и листьев, способность к вегетативному возобновлению и размножению и характер органов, обеспечивающих эти процессы. Но вследствие большого числа признаков четкой системы жизненных форм он не построил.
Наибольшую популярность не только среди ботаников, но и среди неспециалистов завоевала классификация жизненных форм, предложенная крупным датским ботаником К. Раункиером. Раункиер очень удачно выделил из всей совокупности признаков жизненных форм один чрезвычайно важный признак, характеризующий приспособление растений к перенесению неблагоприятного времени года - холодного или сухого. Этот признак - положение почек возобновления на растении по отношению к уровню субстрата и снегового покрова. Раункиер связал это с защитой почек в неблагоприятное время года.
По Раункиеру, жизненные формы растений можно подразделить на пять главнейших типов: фанерофиты (Ph), xамефиты (Ch), гемикриптофиты (НК), криптофиты (К) и терофиты (Th) (от греч. слов "фанерос" - открытый, явный; "хаме" - низкий, приземистый; "геми" - полу-; "кри-птос" - скрытый; "терос" - лето; "фитон" - растение). Схематически эти типы показаны на рисунке 60 (вверху).
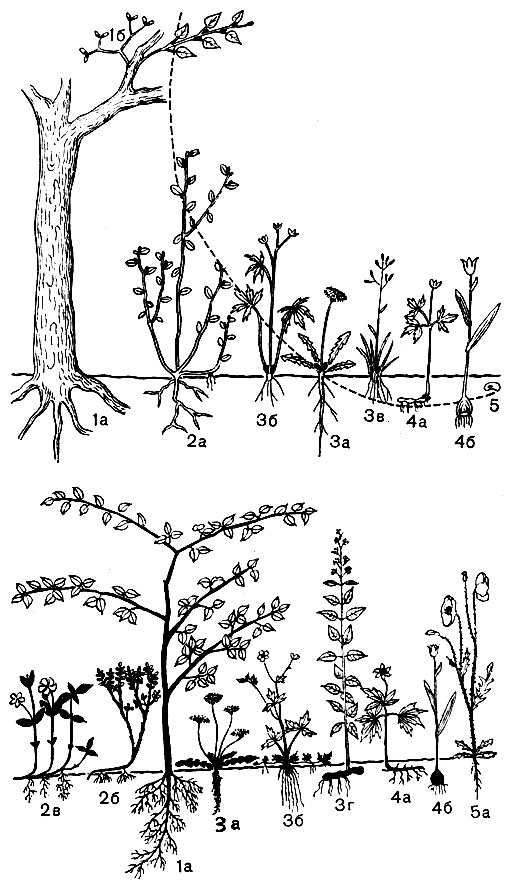
Рис. 60. Жизненные формы растений но Раункиеру (схема): 1 - фанер офиты (1а - тополь, 16 - омела); 2 - хамефиты (2а - брусника, 26 - черника, 2в - барвинок); 3 - гемикриптофиты (3а - одуванчик, розеточный гемикриптофит, 3б - лютик, 3в - кустовой злак, 3г - вербейник обыкновенный, 'протогемикриптофит'); 4 - геофиты (4а - ветреница, корневищный геофит, 4б - тюльпан, луковичный геофит); 5 - терофиты (5а - мак-самосейка). Вверху - черным показаны зимующие почки возобновления (пунктирная линия - уровень их расположения); внизу - соотношение отмирающих и перезимовывающих частей (черным - остающиеся, белым - отмирающие на зиму)
У фанерофитов почки зимуют или переносят засушливый период "открыто", достаточно высоко над землей (деревья, кустарники, деревянистые лианы, эпифиты). В связи с этим они обычно защищены специальными почечными чешуями, имеющими ряд приспособлений главным образом для сохранения конуса нарастания и молодых зачатков листьев, заключенных в них от потери влаги. Почки хамефитов располагаются почти на уровне почвы или не выше 20 - 30 см над ней (кустарнички, полукустарнички, стелющиеся растения). В холодном и умеренном климате эти почки очень часто получают зимой дополнительную защиту, помимо собственных почечных чешуи: они зимуют под снегом. Гемикриптофиты - обычно травянистые растения; их почки возобновления находятся на уровне почвы или погружены очень неглубоко, главным образом в подстилку, образуемую листовым и прочим мертвым растительным отпадом, - это еще один дополнительный "покров" для почек. Среди гемикриптофитов Раункиер различает "протогемикриптофиты" с удлиненными .побегами, ежегодно отмирающими до основания, где располагаются почки возобновления, и розеточные гемикриптофиты, у которых укороченные побеги могут зимовать на уровне почвы целиком (рис. 60). Криптофиты представлены либо геофитами (G), у которых почки находятся в земле на некоторой глубине (они подразделяются на корневищные, клубневые, луковичные), либо гидрофитам и, у которых почки зимуют под водой. Терофиты - особая группа; это однолетники, у которых все вегетативные части отмирают к концу сезона и зимующих почек не остается, - эти растения возобновляются на следующий год из семян, перезимовывающих или переживающих сухой период на почве или в почве.
Ясно, что раункиеровские типы - категории очень крупные, сборные. Раункиер подразделял их по разным признакам, в частности фанерофиты - по размерам (мега-, мезо-, нано-, микрофанерофиты), по характеру почечных покровов, по признаку вечнозелености или листопадности, особо выделял суккуленты и лианы; для подразделения гемикриптофитов и геофитов он пользовался структурой их "летних побегов", характером подземных органов.
Раункиер применил свою систему для выяснения взаимосвязи жизненных форм растений и климата, и получилась замечательно четкая картина. В так называемых "биологических спектрах" он показал участие (в %) своих типов жизненных форм в составе флоры разных зон и стран.
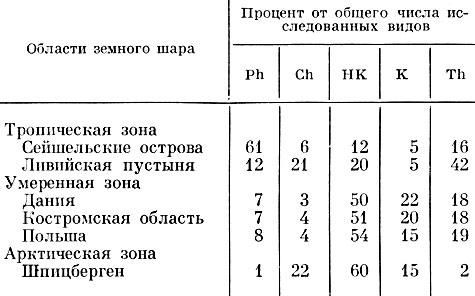
Биологические спектры растительности в разных зонах земного шара
В дальнейшем такими спектрами пользовались многие авторы. На основании анализа биологических спектров климат влажных тропиков был назван климатом фанерофитов, климат умеренно холодных областей - климатом гемикриптофитов, терофиты оказались господствующей группой в пустынях средиземноморского типа, а хамефиты активно участвуют и в тундровой и в пустынной растительности (что, конечно, указывает на неоднородность этой группы).
Подобные спектры могут быть очень показательными при анализе жизненных форм и в разных сообществах одной и той же климатической зоны.
Например, в пределах климата гемикриптофитов тем не менее можно различить сообщества, более близкие к тропическим по составу жизненных форм (широколиственные леса), несущие черты арктические (хвойные леса, высокогорья) и черты средиземноморские в смысле господства терофитов (полевые сорняки). Это легко объясняется различиями в условиях обитания этих сообществ, в частности их резко различным микроклиматом, степенью увлажнения, характером субстрата и т. д.
Скрупулезный анализ известных науке ископаемых растений на предмет их отнесения к той или иной группе жизненных форм показал, что в историческом аспекте группы эти неравноценны. Последовательность их возникновения и наиболее массового развития отражает смену климатических и других мощных физико-географических комплексов условий в различные геологические периоды. Наиболее древними оказались мега- и мезофанерофиты, получившие максимальное развитие в меловом периоде. В палеогене господствовали микрофанерофиты и лианы, в неогене развивались главным образом нанофанерофиты и гемикриптофиты. Самые молодые жизненные формы - хамефиты, геофиты и терофиты - максимально распространились в четвертичном периоде.
Интересно, что раункиеровские типы жизненных форм, отражающие приспособление к неблагоприятному времени года, оказались "универсальной реальностью", и признак положения почек всегда более или менее четко коррелирует с комплексом других, в том числе чисто физиономических, признаков. Поэтому классификацией Раункиера охотно пользуются не только ботаники, изучающие растительность холодных, умеренных, сезоннозасушливых областей, но и "тропические ботаники", имеющие дело с равномерно благоприятным климатом дождевых тропических лесов.
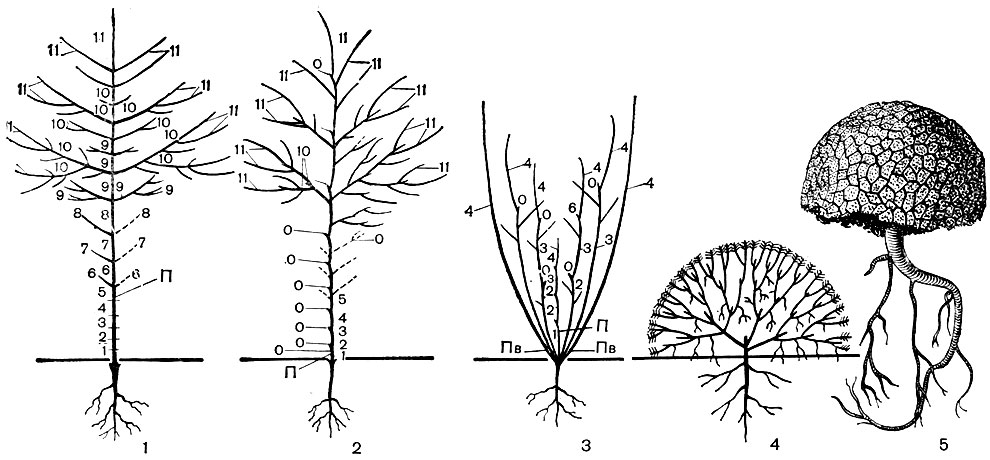
Рис. 61. Схема образования некоторых жизненных форм: 1,2 - дерево; 3 - кустарник; 4, 5 - подушка (1 - 4 - схемы, 5 - Azorella selago из семейства зонтичных с острова Кергелен). На рисунках 1 - 3 мелкими цифрами указаны последовательные годичные приросты (пунктиром - уже отмершие). П - первичный (главный) побег, О - отмершие кончики побегов, Пв - почки возобновления у кустарника. 1 - ствол дерева, представляющий собой длительно растущий главный побег (моноподиальный); 2 - ствол 'составной', т. е. сформирован из побегов последовательных порядков (симподиальный)
Другим примером использования одного важного признака жизненных форм для их классификации может служить подразделение, предложенное Г. М. 3озулиным. Принцип подразделения здесь совсем иной: жизненно важен прежде всего способ удержания растениями площади их обитания и способы распространения по ней, что в общем определяет сосуществование растений со своими соседями по фитоценозу. Выделено 5 основных типов жизненных форм: 1) реддитивные - многолетники, не возобновляющиеся при уничтожении их надземных частей ("уступающие"); 2) рестативные - многолетники, возобновляющиеся и "сопротивляющиеся" захвату площади другими особями; 3) иррумптивные - многолетники, не только возобновляющиеся, но и имеющие органы вегетативного разрастания и размножения, "вторгающиеся", "захватывающие" территорию у других растений; 4) вагативные - однолетние или двусезонные виды, не удерживающие площадь и прорастающие каждый раз на новом месте, "кочующие" или "блуждающие"; 5) инсидентные - не занимающие отдельной площади обитания, "сидящие" на других растениях (эпифиты, паразиты). Эти типы далее подразделяются по характеру отмирания органов, по типам побегов и т. д. и в конечном счете тоже закономерно распределяются по определенным климатическим областям. Однако здесь в центре внимания именно ценотические приспособления. Эта классификация развивает систему жизденных форм, предложенную еще в 1915 г. крупным почвоведом и геоботаником Г. Н. Высоцким, где в основу положена "степень вегетативной подвижности" растений. Так, стержнекорневые или луковичные жизненные формы оказываются вегетативно неподвижными, а длиннокорневищные - подвижными.
По ходу изложения мы все время пользуемся, как само собой разумеющимися, терминами "дерево", "кустарник", "стланец", "подушка", "лиана", "корневищный травянистый многолетник" и т. д. Однако все они - тоже названия крупных категорий жизненных форм, различавшихся еще с древних времен. Недаром большинство этих названий давным-давно вошло в обиходный язык, а некоторые, наоборот, взяты из обиходного языка. Не всегда очевидна в этой эколого-морфологической классификации приспособительность той или иной жизненной формы. Например, для лиан, или лазающих растений, она, как мы видели, ясна, а вот к чему приспособлено "дерево"? Но, оказывается, и здесь можно обнаружить соответствие определенному комплексу внешних условий. Статистические подсчеты показывают, что самый высокий процент деревьев - во флоре влажнотропических лесов (до 88% в Амазонской области Бразилии), а в тундре и высокогорьях нет ни одного настоящего прямостоячего дерева. В области таежных лесов умеренно холодной зоны деревья хотя и господствуют в ландшафте, однако это всего 1 - 2 или несколько видов, составляющих ничтожный процент от общего числа видов, и они, как ^правило, имеют специальные приспособления для перенесения зимы либо в виде особой анатомической структуры и физиологических особенностей листьев (хвои), либо в виде закономерного листопада и т. п. Во флоре умеренной лесной зоны Европы деревья составляют не более 10 - 12% от общего числа видов.
Таким образом, жизненная форма дерева оказывается выражением приспособления к наиболее благоприятным для роста условиям - климатическим и ценотическим. Жизнь в лесу в окружении соседних деревьев вызывает необходимость вынесения органов ассимиляции вверх. У деревьев наиболее полно выражена способность к интенсивному и длительному росту побегов; вследствие этого деревья достигают наибольших для высших растений размеров. Помещая свои кроны высоко над землей, они занимают максимальное пространство.
Отличительная черта всякого прямостоячего дерева - образование единственного ствола, биологически главной, "лидерной" оси, всегда стремящейся сохранить более или менее вертикальное направление роста и растущей интенсивнее остальных побегов (и в длину, и в толщину). Ветвление, если оно выражено, у дерева обычно акротонное, т. е. наиболее сильные ветви развиваются ближе к верхушке ствола и его крупных ответвлений, а в нижних частях ствола боковые ветви или совсем не развиваются, или развиваются слабо и быстро отмирают. Так формируется крона в верхней части ствола (рис. 61).
В некотором роде антагонистом дерева оказывается растение-подушка, воплощающая наибольшую заторможенность роста всех побегов, вследствие чего возникает многократное равномерное ветвление без выделения "главного ствола"; каждая веточка подуниш продолжает испытывать крайнее угнетение роста в длину (рис. 61). Подушковидные растения встречаются во всех зонах, но приурочены к наиболее неблагоприятным местообитаниям: с низкими температурами воздуха и почвы, с холодными штормовыми ветрами, с крайней сухостью почвы и низкой влажностью воздуха и т. п. Экологически разнообразные местообитания подушек (тундры, высокогорья, субантарктические острова и побережья, пустыни, скалы и осыпи) связывает один общий фактор: свободный доступ света, который играет, вероятно, немалую роль в подавлении роста их побегов.
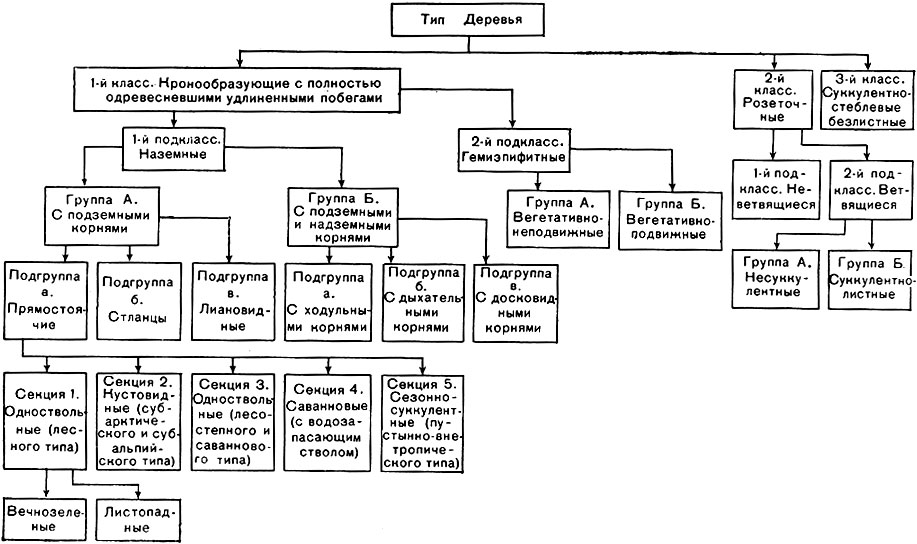
Использовав и обобщив ранее предложенные классификации жизненных форм по морфологическим признакам, И. Г. Серебряков положил в основу своей системы признак длительности жизни всего растения и его скелетных осей, как наиболее четко отражающий влияние внешних условий на морфогенез и рост. Эта система выглядит следующим образом:
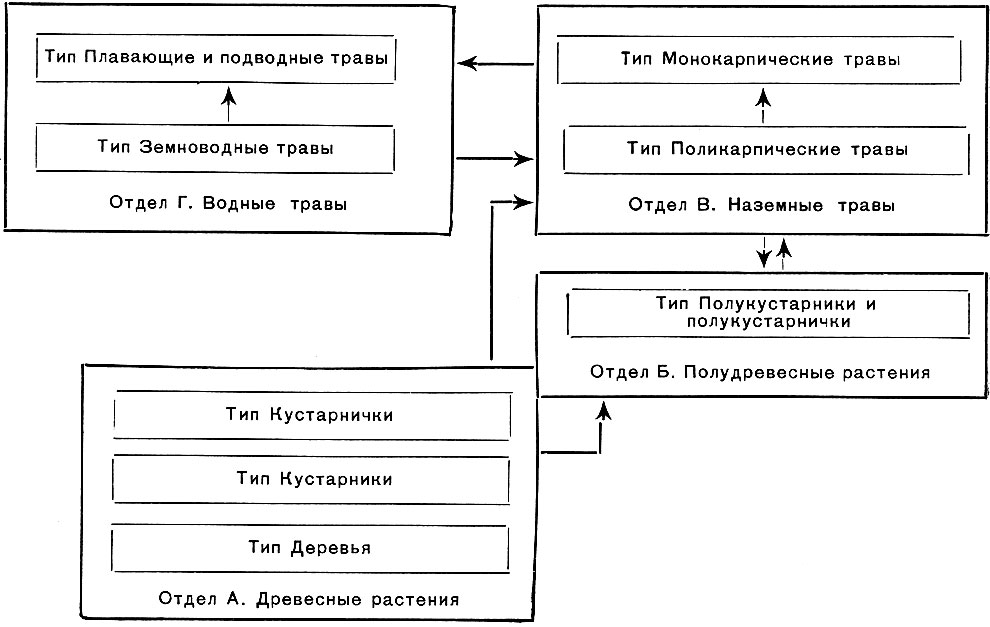
Соотношение отделов и типов жизненных форм покрытосеменных растении
Различие между деревьями, кустарниками, кустарничками, полукустарниками и полукустарничками и травянистыми растениями состоит, помимо разной степени одревеснения их стеблей, именно в длительности жизни и характере смены скелетных побегов в общей побеговой системе. Ствол у деревьев живет столько* же, сколько и все дерево целиком, - от нескольких десятков до нескольких сотен лет, а иногда и до тысяч лет (мамонтово дерево). Спящие почки у основания ствола дают сестринские стволы только в случае, если главный ствол срублен или поврежден иным путем (пневая поросль). У кустарников (рис. 61) главный побег ведет себя как небольшое деревцо, однако довольно рано, на 3, 5, 10-й год жизни, из спящих почек у основания стволика начинают расти новые стволики, часто перегоняющие материнскрий и постепенно сменяющие друг друга.
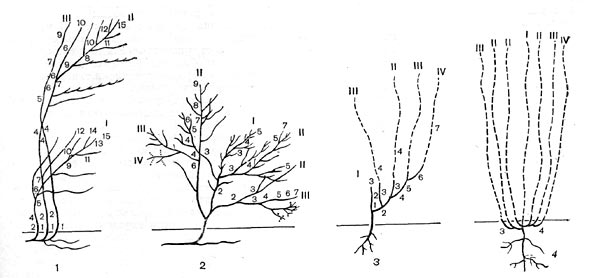
Рис. 62. Схема формирования куста: 1 - у кустарника; 2 - у кустарничка; 3 - у полукустарника; 4 - у длиннопобегового травянистого многолетника (протогемикриптофита). Точками и прерывистыми линиями обозначены ежегодно отмирающие части побегов; римскими цифрами - порядки основных структурных осей; мелкими арабскими цифрами - годичные приросты
В целом длительность жизни кустарника может быть тоже очень большой, в несколько сотен лет, но каждый из стволиков, или скелетных осей, живет в среднем 10 - 40 лет (крайние пределы - от 2 лет у малины до 60 с лишним лет у желтой акации, сирени и др.). Они сосуществуют во времени, сменяясь по мере отмирания главного и ближайших к нему дочерних стволиков в центре куста и появления новых на периферии куста.
Кустарнички представляют собой миниатюрные кустарники с тем же основным способом ветвления, однако они более низкорослы и длительность жизни отдельных скелетных осей у них меньше, 5 - 10 лет. Очень распространены кустарнички в тундрах, высоко в горах, на сфагновых болотах, под пологом хвойных таежных лесов (черника, брусника, голубика, клюква, вереск, водяника и т. д.). Многие из них принадлежат к семейству вересковых.
Соотношение между кустарниками, кустарничками, полукустарниками и многолетними травами с удлиненными побегами хорошо показано на рисунке 62. Если у кустарничков, как и у кустарников, в связи с цветением и плодоношением отмирают ежегодно очень небольшие части их побеговой системы, то у полудревесных, а особенно у травянистых жизненных форм это отмирание играет решающую роль в сложении общего облика растения. Полукустарники и полукустарнички, особенно характерные для пустынных и полупустынных областей (разные виды полыней, солянок), формируются по принципу кустарника, но имеют меньшую продолжительность жизни скелетных осей (5 - 8 лет) и к тому же ежегодно (во взрослом состоянии) теряют после цветения всю верхнюю часть своих годичных цветоносных побегов, иногда составляющую до 3/4 и более от общей высоты побегов. Остающаяся деревянистая многолетняя система "пеньков" несет на себе почки возобновления, располагающиеся над землей (хамефиты, по Раункиеру). Это отражает специфику пустынных полукустарничков: почки не могут находиться в почве, которая летом чрезмерно перегревается.
У многолетних травянистых растений прямостоячие надземные побеги живут один вегетационный сезон и после цветения и плодоношения отмирают до основания. Но на остающемся основании под землей или на уровне почвы формируются зимующие почки (по Раункиеру, это геофиты или гемикриптофиты). У некоторых трав, розеточных и ползучих, надземные стебли могут жить и несколько лет, но при условии, что они остаются плотно прижатыми к почве.
Подразделение наземных травянистых растений основано в системе И. Г. Серебрякова на признаке моно- или поликарпичности, т. е. способности к повторному плодоношению. Большинство многолетних трав - поликарпики, но бывают и монокарпики: несколько лет растут, оставаясь в виде вегетативной розетки, а потом зацветают и после плодоношения отмирают целиком. Так ведут себя многие наши зонтичные: тмин, порезник, дягиль - в средней полосе, ферулы - в Средней Азии.
К монокарпикам относятся и однолетники (терофиты), цветущие в первый год жизни; особенно краток жизненный цикл эфемеров, укладывающийся в считанные недели. В предгорных пустынях Средней Азии рано весной образуются эфемеровые луга, в составе которых преобладают однолетники. К началу мая они уже полностью исчезают, выгорают, оставляя в почве только семена.
Многолетние поликарпики подразделяются большей частью по форме подземных многолетних органов. Обычно различают стержне-корневые, кистекорневые, дерновые, коротко и длиннокорневищные, клубневые, луковичные многолетние травы. Особый отдел составляют водные травы (табл. 15); они подразделены по самому броскому физиономическому признаку на погруженные (элодея), плавающие (кувшинки, водокрас) и земноводные (частуха, стрелолист, белокрыльник).
В пределах типа деревья тоже очень разнообразны. В основу их классификации положено строение надземных органов, однако корневые системы играют не последнюю роль в создании облика некоторых деревьев, даже без раскопок. Стоит лишь вспомнить о своеобразных досковидных корнях, корнях-подпорках, корнях-ходулях у многих тропических деревьев. Особенно ясна связь корней-подпорок с условиями обитания у мангровых растений, образующих заросли в полосе прилива и отлива у побережий океанов в тропиках. У них же нередко можно наблюдать торчащие из ила дыхательные корни, доставляющие кислород через воздухоносную ткань к глубоко расположенным частям корневой системы, где избыток влаги создает плохую аэрацию, недостаточное воздухоснабжение. А какие своеобразные формы стволов у саван-новых "бутылочных деревьев" (табл. 17), запасающих в стволе воду! Известный африканский баобаб хотя и не принадлежит к "бутылочным", однако тоже отличается мощностью и толщиной ствола, богатого мягкой водозапасающей паренхимой. Еще более яркий пример деревьев с водозапасающими стволами - древовидные кактусы.
В качестве примера приводим одну из схем классификации деревьев (стр. 97). В ней использованы разнообразные габитуальные признаки, четко отражающие связь жизненных форм с условиями обитания (определения "лесные", "саванновые", "субарктические" и т. п. пришлось ввести в название групп).
Для саванновых деревьев, например, очень характерна плоская зонтиковидная крона, как у австралийских и африканских акаций.
Иногда такая форма встречается и не в саваннах, но обязательно в климате с засушливым летом (средиземноморские пинии). Наиболее велико разнообразие деревьев именно в тропических странах. Только там встречаются розеточные, суккулентные, лиановидные, полу-эпифитные деревья и деревья с разнообразными метаморфизированными корнями (табл. 18).
В ходе истории растительного мира жизненные формы эволюционировали, разумеется, не сами по себе, а как неотъемлемый комплекс признаков той или иной эволюционирующей систематической группы. На заре возникновения наземной растительности первенцы ее, вышедшие из моря на сушу, во многих отношениях сохраняли сходство со своими предками - водорослями. Это были некрупные растения, близкие к травянистым не только по размерам, но и по совокупности своих морфологических и анатомических черт. В дальнейшем развились и крупные древовидные формы, в том числе своеобразные древовидные папоротники с розеткой больших перистых листьев на верхушке "ствола", и древовидные лепидодендроны и сигиллярии, достигавшие 30 - 45 м высоты, и столь же крупные древовидные каламиты - предки хвощей. Наряду с этими формами, вероятно, с давних пор существовали и травянистые папоротниковидные. Например, травянистые папоротники, плауны и хвощи дожили до наших дней, тогда как значительная часть древовидных форм вымерла. Что касается мхов, то они в течение своей длительной истории остались "карликовыми травами". Голосеменные, наоборот, представляют преимущественно древесную группу, во всяком случае, среди ныне живущих голосеменных настоящих "классических" трав нет. Саговники-толстоствольные розеточные деревья разного размера, но среди них есть и совсем маленькие растения. Например, замия (Zamia pygmaea), живущая на Кубе, высотой всего 2 - 3 см - ее так же трудно отнести к деревьям, как и к травам. Хвойные, широко распространенные на Земле, имеют облик крупных деревьев, реже кустарников (обыкновенный можжевельник) и стланцев (в горах Восточной Сибири кедровый стланик, табл. 14).
Совершенно своеобразна среди голосеменных и не похожа по жизненной форме ни на какие другие растения вельвичия удивительная (табл. 17), растущая в пустыне Намиб и на юго-западном побережье Африки. Ствол этого "дерева-карлика" похож на обрубок или пень, очень низкий и толстый (до 50 см в высоту и до 1,2 м в диаметре). Он суживается книзу, а наверху несет два длинных кожистых листа, сохраняющихся в течение всей жизни растения и растущих у основания вставочно. Это собственно самые первые листья растения - семядольные, так что все растение как бы "взрослый проросток".
Цветковые наиболее разнообразны по жизненным формам. Широко признано, что в ходе эволюции они прошли путь от сравнительно невысоких толстоствольных маловетвящихся розеточных деревцев (такие сейчас встречаются главным образом в тропических лесах, например пальмы, дынное дерево Carica papaya) к крупным, "настоящим" деревьям с хорошо развитым стволом и мелковетвистой кроной, а от деревьев - к кустарникам, кустарничкам и разнообразным травам. Направление "от деревьев к травам" называют "редукционной эволюцией" или "соматической редукцией" и связывают с расселением цветковых из области их возникновения и первоначального развития (предположительно, в горах тропиков и субтропиков) в области и зоны с менее благоприятными, иногда очень суровыми условиями обитания. Травянистые растения лучше приспособлены для освоения новых экологических ниш и проникают буквально "в каждую щель".
Однако это не значит, что каждое конкретное семейство или род обязательно прошли в ходе своей эволюции весь путь "соматической редукции". Некоторые семейства, видимо, с самого начала были травянистыми, и в некоторых случаях от травянистых предков возникли более специализированные древесные формы (бамбуки в семействе злаков). В крайних условиях эволюция приводила то к стланикам, то к подушкам, то к луковичным геофитам, то к однолетним эфемерам. Эфемеры считаются эволюционно наиболее молодой группой жизненных форм, свойственной области древнего Средиземья, которая стала сушей при высыхании древнего Средиземного моря - Тетис.
Разные линии эволюции жизненных форм, параллельные и независимые в разных группах цветковых и "перекрещивающиеся", показаны на схеме (стр. 97). Это обобщенная схема, демонстрирующая многообразие возможных путей преобразования жизненных форм, но отнюдь не исчерпывающая их. Здесь не показаны, например, своеобразные линии эволюции, приведшие к возникновению среди цветковых растений жизненных форм с особыми способами питания (табл. 16): паразитных (петров крест), сапрофитных (некоторые орхидеи), насекомоядных (росянка, непентес и др.). Отдельные фрагменты этой схемы можно иллюстрировать примерами представителей из разных семейств. Так, почти весь редукционный ряд суккулентов - от древовидных (табл. 19) до травянистых - можно найти в семействе кактусовых. Ряд многолетних стелющихся форм, берущих начало, видимо, от прямостоячих деревьев, можно проследить среди берез и ив (вплоть до почти травянистой тундровой ивы Salix herbacea). В семействе тыквенных крупные тропические древесные лианы трансформировались в травянистые лианы с ежегодно отмирающим надземным стеблем (бриония), а от таких форм, вероятно, возникли стелющиеся травы, многолетние и однолетние, например огурец, арбуз, тыква. В семействе вересковых представлены ряды от кустарников к кустарничкам, прямостоячим, стелющимся и даже подушковидным, однако до травянистых форм в этом семействе дело не доходит. "Эстафету" принимает близкое к вересковым семейство грушанковых, где некоторые представители близки к кустарничкам (зимолюбка); другие можно считать настоящими корневищными многолетними травами (грушанка); наконец, одноцветка (Moneses uniflora) - это "вегетативный однолетник", ежегодно сменяющий не только надземные, но и подземные органы.
В заключение следует сказать, что изучение жизненных форм, их признаков, приспособлений к переживанию неблагоприятного периода, возрастных изменений, вегетативного возобновления и размножения и т. д. имеет не только чисто теоретический интерес, но и важное прикладное значение. Именно от этих особенностей зависит сохранение и возобновление дикорастущих растений, в том числе используемых человеком, например лекарственных, а также успех интродукции, т. е. переселения растений в новые для них районы.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'