Растительные сообщества (Т. А. Работнов)
Выдающийся советский ученый В. И. Вернадский разработал представление о биосфере - наружной оболочке Земли, свойства которой определяются жизнедеятельностью организмов.
В. И. Вернадский понимал биосферу широко, включая в нее не только часть земного шара, заселенную организмами в настоящее время, но и земную кору, созданную организмами в прошлые геологические эпохи, например мощные толщи известняков и т. п.
Большинство ученых не разделяют такое широкое понимание биосферы.
Под биосферой обычно понимают сферу современной жизни, т. е. часть наружной оболочки Земли, населенную в настоящее время организмами.
Биосфера состоит из биогеоценозов, образованных организмами совместно со свойственной им косной (неживой) средой (рис. 63).
По теории В. Н. Сукачева - создателя биогеоценологии (науки о биогеоценозах), биогеоценозы состоят из двух основных компонентов - биоценоза (сообщества организмов) и экотопа (косной среды). В состав биоценоза входят растения, образующие растительное сообщество (фитоценоз), животные и микроорганизмы. Среда, в которой обитают организмы (экотоп), определяется условиями климата, гидрологией, почвообразующей породой, почвой. Между организмами и их средой в биогеоценозах существуют сложные взаимосвязи (рис. 64). Биогеоценозы иногда называют экосистемами.
Так как микроорганизмы представлены либо микроскопическими растениями (бактерии, актиномицеты, водоросли, грибы), либо микроскопическими животными (простейшие и др.), то можно говорить, что в состав биоценозов входят и растения и животные. Еще более существенно разделение организмов, входящих в состав биоценозов, на автотрофов и гетеротрофов.
Автотрофы в основном представлены фототрофами - зелеными растениями, способными создавать органическое вещество, используя энергию солнечных лучей. К автотрофам также относятся и хемотрофы - бактерии, получающие энергию в результате окисления некоторых химических соединений, например нитрифицирующие бактерии, окисляющие аммоний в нитриты, а затем в нитраты. Роль хемотрофов в энергетике биогеоценозов невелика. Энергетическую базу для жизнедеятельности биогеоценозов, включая процесс почвообразования, создают зеленые растения. Они продуцируют основную массу органического вещества (первичную продукцию), и потому их называют продуцентами.
Гетеротрофы не способны создавать органическое вещество, используя энергию солнечных лучей или химических соединений. Они могут существовать лишь при обеспечении их энергией органического вещества, созданного другими организмами.
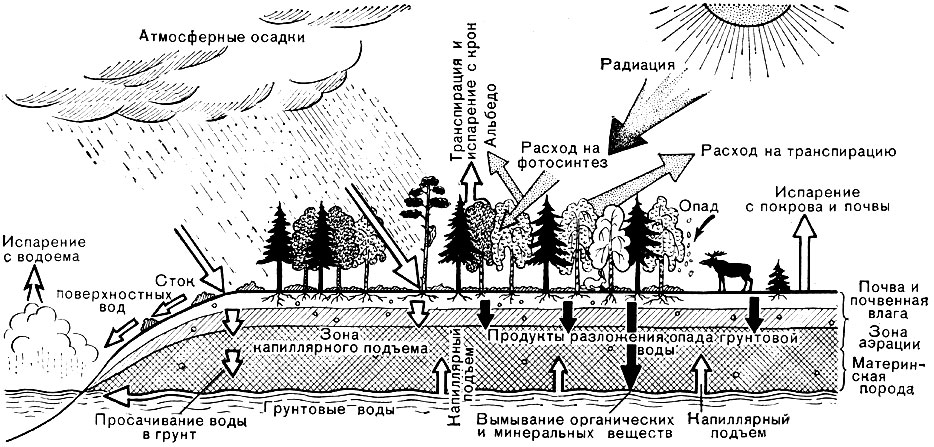
Рис. 63. Схема взаимодействия компонентов биоценоза
К гетеротрофам относятся все животные и все бесхлорофильные растения, включая цветковые паразиты и сапрофиты, грибы, актиномицеты, бактерии (за исключением хемотроф-ных и немногих фототрофных). Гетеротрофы также продуцируют органическое вещество (вторичную продукцию), но в значительно меньших количествах, чем автотрофы-фототрофы. Биоценозы можно рассматривать как закономерные системы взаимозависимых двух групп организмов - автотрофов и гетеротрофов. Гетеротрофы не могут существовать без автотрофов, поскольку получают от них энергию. Однако и автотрофы не могут существовать в отсутствие гетеротрофов, точнее, в отсутствие сапротрофов - организмов, использующих энергию отмерших органов растений, а также энергию, содержащуюся в экскрементах и трупах животных. В результате жизнедеятельности сапротрофов происходит минерализация так называемого мертвого органического вещества. Минерализация в основном происходит в результате деятельности бактерий, грибов и актиномицетов. Однако роль животных в этом процессе также очень велика. Размельчая растительные остатки, поедая их и выделяя как экскременты, а также создавая в почве более благоприятные условия для деятельности сапротрофных микроорганизмов, они ускоряют процесс минерализации отмерших органов растений. Без этого процесса, ведущего к поступлению в почву доступных форм минерального питания, растения-автотрофы быстро использовали бы наличные запасы доступных форм макро- и микроэлементов и не смогли бы жить; биогеоценозы превратились бы в кладбища, переполненные трупами растений и животных.
Формирование биогеоценозов происходило в течение длительного времени, в период которого подбирались виды организмов, способные существовать совместно друг с другом в данных условиях среды. Это обстоятельство необходимо учитывать для понимания сложных взаимоотношений, существующих в биогеоценозах между отдельными видами организмов, автотрофами и гетеротрофами, организмами и средой.
Знакомясь с любым растительным сообществом (например, в лесу, на лугу, в степи, в поле, занятом посевом), нельзя забывать, что луг, лес, болото, степь (рис. 66 - 69), поле - это биогеоценозы, составной частью которых являются существующие растительные сообщества (лесные, луговые, степные, посевы). Поскольку растительное сообщество всегда входит как компонент в биоценоз и биогеоценоз, то, естественно, его свойства существенно зависят от зоокомпонентов и косной среды биогеоценоза.
В состав растительных сообществ входят не только сосудистые растения, мхи и лишайники, но и грибы, бактерии, актиномицеты, микроскопические водоросли. Поэтому определение полного флористического состава растительных сообществ представляет большие трудности и возможно лишь при использовании специальных методов исследования. Однако многие очень существенные признаки растительных сообществ (например, внешний вид, структура, продуктивность) определяются составом сосудистых растений. Поэтому состав этой группы растений можно считать важнейшим признаком растительных сообществ.
Большинство растительных сообществ характеризуется достаточно богатым флористическим составом группы сосудистых растений. Например, в хвойных лесах (еловых, сосновых, лиственничных) нашей страны обычно совместно произрастает до 30 и более видов; в дубовых лесах - до 40 - 50; на лугах - 30 - 50; в луговых степях - до 100 и более видов; в типичных дерновинно-злаковых степях - 50 - 70 видов и т. д.
Наибольшего богатства флористический состав растительных сообществ достигает в тропических лесах, где совместно произрастает до 100 видов деревьев, не считая видов травянистых растений. Лишь в особых условиях, исключающих возможность произрастания большей части видов растений (например, при сильном засолении), формируются растительные сообщества из немногих видов, в редких случаях из одного вида сосудистых растений. Иногда одновидовые растительные сообщества представляют сравнительно непродолжительную возрастную стадию развития какого-либо биогеоценоза. Примером могут быть так называемые мертвопокровные леса, где из-за недостатка света под пологом деревьев (от сомкнутости их крон) другие растения произрастать не могут.
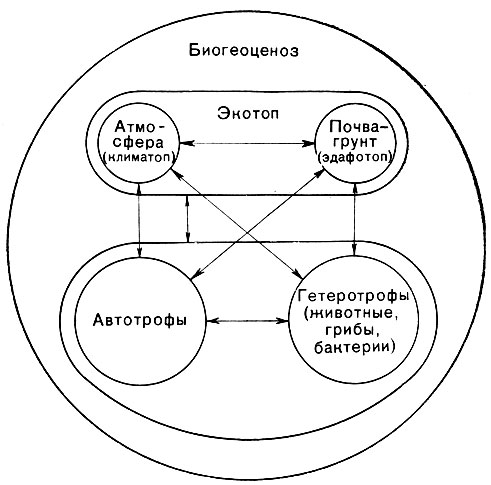
Рис. 64. Схема взаимного влияния лесного фитоценоза и условий среды
Не меньшее значение для характеристики растительных сообществ имеет то, что они образованы не только многими видами-, но и видами, различными по своим экологическим и биологическим свойствам и по значимости в определении особенностей растительного сообщества.
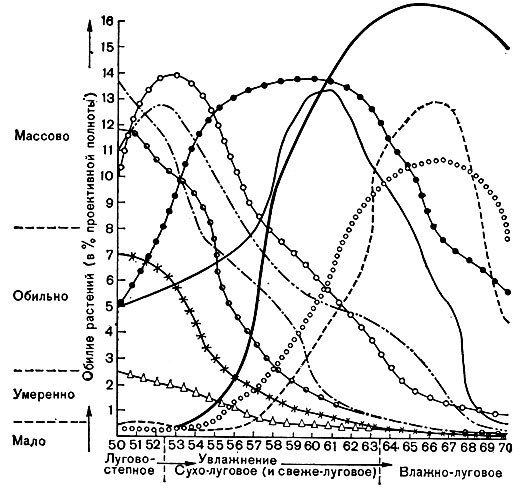
Рис. 65. Распределение отдельных видов растений (каждому виду соответствует определенная кривая) в зависимости от увлажнения
Выдающийся советский геоботаник Л. Г. Раменский обосновал положение об экологической индивидуальности видов растений. Нет двух видов, одинаково относящихся к условиям произрастания (к обеспечению светом, водой, элементами минерального питания и т. п.), имеющих одинаковые кривые распределения при изменении условий произрастания в пространстве или во времени. Это видно из графика, составленного Л. Г. Раменским, демонстрирующего изменения проективного покрытия отдельных видов луговых растений в зависимости от условий произрастания (цвет. табл. 20), а также на рисунке 65, где показано распространение видов растений в зависимости от увлажнения и богатства почвы.

Рис. 66. Вейниковый луг

Рис. 67. Сосновый лес
Отдельные виды растений своеобразны не только экологически (по их отношению к условиям произрастания), но и биологически (по приспособлениям, обеспечивающим им возможность существования и размножения в определенных условиях среды).

Рис. 68. Кочкарное болото

Рис. 69. Ковыльная степь
Растительные сообщества образованы многими видами, а это означает, что в состав сообщества входят виды, различные по своим экологическим и биологическим свойствам. Такие различия нередко настолько значительны, что виды, входящие в состав одного и того же сообщества, представлены различными жизненными формами (деревьями, кустарниками, травами и пр.), а помимо того, относятся к различным типам по длительности жизни, по способам размножения и т. д.
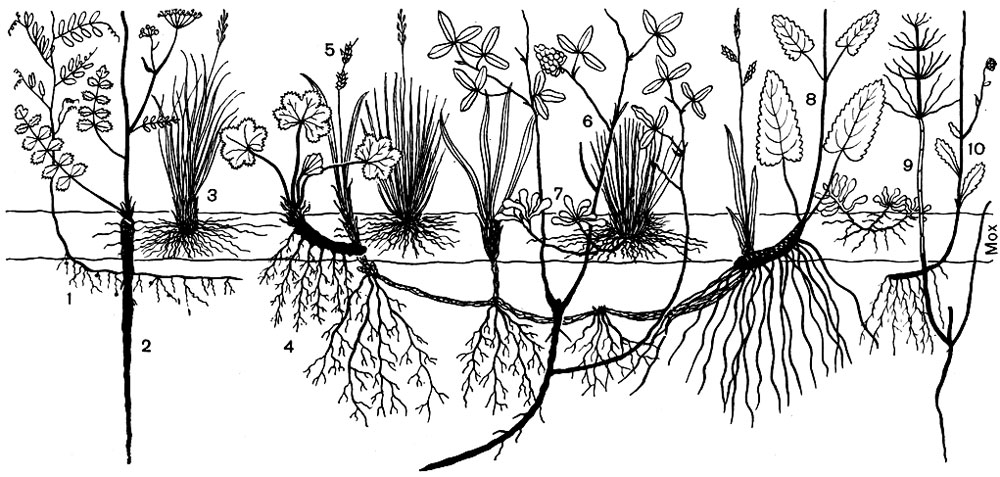
Рис. 70. Вертикальный разрез через надземную и подземную части биогеоценоза суходольного луга. 1 - мышиный горошек; 2 - бедренец-камнеломка; 3 - овечья овсяница; 4 - манжетка; 5 - просяная осока; 6 - средний клевер; 7 - кошачья лапка; 8 - лекарственная буквица; 9 - хвощ; 10 - нивяник
Примером могут быть растительные сообщества еловых лесов, в состав которых входят деревья (ель), кустарники (крушина ломкая, жимолость лесная), кустарнички (брусника, черника), травы (кислица обыкновенная, майник, седмичник и др.), гипновые мхи. Среди трав преобладают многолетники, размножающиеся вегетативным путем, но встречаются и однолетники, размножающиеся исключительно семенами (например, полупаразитное растение марьяник луговой).

Соотношение (в %) видов трав с различной глубиной укоренения на материковых лугах Карельской АССР
Кустарнички представлены как вечнозеленой брусникой, так и ежегодно теряющей осенью листья черникой.
В состав растительных сообществ широколиственных лесов, в частности в дубравах, входят деревья (дуб, клен, липа), кустарники (лещина), многие виды трав. Среди трав выделяются две группы видов, отличающиеся по приуроченности периода вегетации либо к весне, либо к лету. К первой группе относятся весенние эфемероиды (хохлатки, ветреница лютиковая, пролеска) - растения, начинающие вегетацию рано весной. Они используют благоприятные условия освещения до начала развертывания листьев на деревьях и успевают за короткий срок образовать соцветия, дать плоды и накопить достаточное количество энергетического материала в органах запаса.
Группа трав летней вегетации представлена в дубравах как растениями, размножающимися исключительно или преимущественно вегетативным путем (сныть обыкновенная, осока волосистая), так и травами, размножающимися исключительно семенами (ежа сборная, бор развесистый).
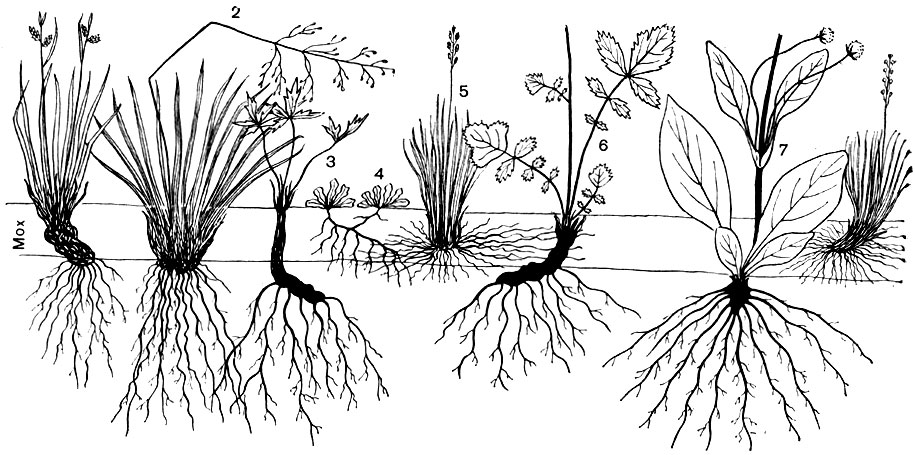
Рис. 71. Вертикальный разрез через надземную и подземную части биогеоценоза суходольного луга. 1 - осока бледная; 2 - щучка дернистая; 3 - лютик едкий; 4 - кошачья лапка; 5 - овсяница овечья; 6 - гравилат речной; 7 - сивец луговой
Приведенные выше примеры характеризуют также различия видов, входящих в состав растительных сообществ, по высоте их надземных органов (деревья, кустарники, травы, мхи). Такие же различия наблюдаются и по глубине проникновения корней отдельных видов, о чем можно судить по данным таблицы, полученным при изучении трех типов лугов, а также по рисункам 70 и 71.
В составе многовидового растительного сообщества всегда можно обнаружить виды, относящиеся к различным экологическим и биологическим типам растений. Следовательно, к числу существенных признаков растительных сообществ можно отнести разнообразие экобиоморфного состава их компонентов.
Разнообразие состава растительных сообществ объясняется еще тем, что, как правило, каждый вид представлен в них большим числом особей, отличающихся по возрасту и жизненному состоянию. Совокупность особей любого вида в пределах конкретного растительного сообщества образует ценотическую популяцию, или ценопопуляцию. В состав ценопопуляций видов цветковых растений могут входить жизнеспособные семена, находящиеся в почве или на ее поверхности, всходы, ювенилы (молодые растения, отличающиеся от взрослых размерами и формой листьев), переходные от ювенильных к взрослым (имматурные) и взрослые растения. В состав популяций некоторых трав входят также особи, находящиеся в состоянии вторичного покоя (корневища, луковицы, клубни, неразвившиеся надземные побеги).
Ценопопуляций отдельных видов в одном и том же растительном сообществе могут сильно отличаться друг от друга как по числу особей на единицу площади, так и по соотношению особей различных возрастных групп (возрастному спектру). Сложный состав популяции сообщества является приспособлением, обеспечивающим большую устойчивость видов в растительных сообществах.
Важное значение имеет свойство семян многих видов растений долго сохранять всхожесть при погребении их в почве. С этим связано большое накопление жизнеспособных семян в почвах лугов, лесов, степей. Число их колеблется в почвах лугов от нескольких тысяч до нескольких десятков тысяч на 1 м2 ; в почвах лесов - от нескольких сотен до нескольких тысяч: на 1 м2 ; в почвах луговой степи под Курском обнаружено 18 000 семян на 1 м2, а в почвах пустынь Средней Азии - от нескольких десятков до нескольких сотен. Благодаря запасу жизнеспособных семян в почве даже при отсутствии обсеменения растений этих видов может происходить их семенное возобновление. Участие в составе ценопопуляций молодых особей (всходов, ювенилов и имматуров) означает, что в благоприятных условиях они могут превратиться во взрослые растения, что обеспечивает устойчивое положение вида в сообществе.
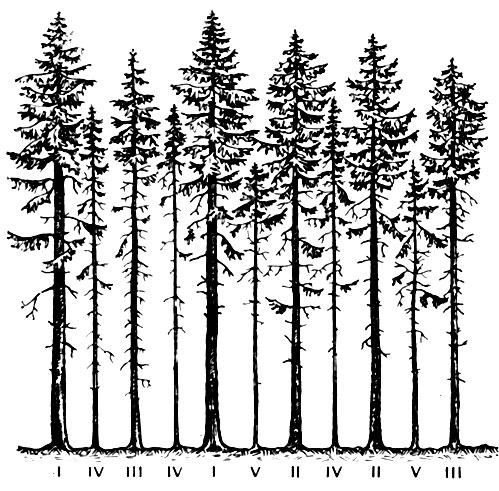
Рис. 72. Распределение деревьев в сообществе по классам Крафта
Большое значение также имеют различия в жизненном состоянии растений в пределах группы взрослых особей. Это особенно заметно в лесах, при изучении которых лесоводы давно используют классификацию Крафта. По этой классификации деревья в лесу разделяются в соответствии с их жизненным состоянием на пять классов. Наиболее мощноразвитые деревья относятся к первому классу, а сильноугнетенные деревья, кроны которых целиком размещаются под пологом более высоких деревьев, - к пятому классу (рис. 72). Деревья, занимающие по жизненному состоянию промежуточное положение, соответственно относятся ко второму, третьему и четвертому классам. Такую же дифференциацию взрослых особей по их жизненному состоянию можно наблюдать и у многолетних травянистых растений в травяных растительных сообществах: одни особи цветут и плодоносят; другие не образуют генеративных побегов, но их вегетативные органы развиты нормально; третьи явно угнетены.

Рис. 73. Луговое растительное сообщество - луг с преобладанием кровохлебки
К очень существенным признакам многовидовых растительных сообществ относится дифференциация видов, входящих в состав растительных сообществ, по их значимости в определении свойств сообществ. В любом растительном сообществе можно различать преобладающие виды, образующие основную массу надземных органов, - доминанты - и ВИДЫ, принимающие относительно небольшое участие в сложении сообщества.
В растительных сообществах, состоящих из видов, относящихся к различным жизненным формам, образующим достаточно сомкнутые покровы (ярусы), можно различать несколько преобладающих растений - доминантов. Например, в еловом лесу с выраженным ярусом из черники и гипновых мхов можно различить три доминанта: ель - в древесном ярусе, чернику - в кустарничковом ярусе, гипновый мох - в моховом ярусе. В некоторых растительных сообществах (например, в тропических лесах) очень трудно выделить один-два доминанта, поскольку в их составе имеется несколько или даже много видов, принимающих Примерно одинаковое участие в сложении сообщества. Это же наблюдается в некоторых травяных, в частности луговых, растительных сообществах (рис. 73).
В растительных сообществах создается особая среда - особый климат (фитоклимат), особые почвенные условия. Доминанты, обладающие большой средообразующей способностью, получили название эдификаторов - строителей сообществ. Примером эдификаторов может быть ель в сообществах незаболоченных еловых лесов. Образуя сомкнутый ярус, ель создает особый фитоклимат: резкое снижение освещенности, особый температурный режим, повышенную влажность воздуха и т. д. В связи с большим количеством опада (хвои, веток, коры, шишек), из которого формируется лесная подстилка, ель оказывает большое влияние на почву. Однако в сообществах заболоченных лесов, где сомкнутый ярус из ели отсутствует, а сфагновые мхи образуют сплошной мощный моховой покров, основное средообразующее воздействие оказывает не ель, а сфагновые мхи, и потому они, а не ель являются эдификатором таких сообществ.
В результате особых условий произрастания, создающихся в растительных сообществах, растения, входящие в их состав, отличаются от одновозрастных особей тех же видов, выросших вне сообществ. Это было давно установлено при изучении деревьев - ели, сосны, дуба и др. Деревья, выросшие вне леса, т. е. без воздействия на них других деревьев, обычно имеют более толстый ствол с хорошо развитой почти от основания кроной. Деревья, выросшие в лесу, характеризуются большей высотой; их ствол более постепенно, чем вне леса, сужается кверху; крона менее развита и приурочена к самой верхней части ствола (рис. 74). Изменения в форме роста наблюдаются и у травянистых растений (рис. 75).
Произрастание в условиях сомкнутых растительных сообществ оказывает большое влияние не только на форму роста растений, но и на способность размножаться семенами и вегетативным путем. Растения, возникшие из семян, в условиях сообществ развиваются медленнее, чем вне сообществ, достигая способности цвести и плодоносить через более продолжительный срок. Например, многие луговые растения в питомниках (на грядках) зацветают на второй год жизни или даже в первый год жизни, если семена высеваются с осени. А в луговых сообществах большинство особей этих видов зацветает не ранее, как в возрасте 3 - 5 лет, а многие из них - лишь в возрасте около 10 и более лет. То же наблюдается и у древесных растений, образующих лесные сообщества. Деревья (ель, дуб, сосна) начинают плодоносить в лесах позже и затем плодоносят реже и менее обильно, чем деревья тех же видов вне леса.
Как правило, растения в растительных сообществах в какой-либо степени угнетены по сравнению с одновозрастными особями тех же видов, выросшими в тех же условиях почв и климата вне сообществ.
Однако существуют некоторые виды, которые не могут произрастать вне определенных растительных сообществ. К ним относится, например, бесхлорофильное растение-сапрофит подъельник, распространенный в тенистых лесах, где он использует с помощью грибного симбионта в качестве источника энергии лесную подстилку и корни деревьев.

Рис. 74. Влияние условий произрастания на форму дерева: слева - вне леса; справа - в лесу
Многие виды растений приурочены лишь к определенным растительным сообществам, будучи хорошо приспособлены к свойственной этим сообществам фитосреде. Примером может быть кислица обыкновенная, успешно произрастающая в темнохвойных лесах в условиях значительного затенения и обычно быстро исчезающая при уничтожении леса. Однако в опытах выращивания кислицы из семян вне природных сообществ она росла при полном солнечном освещении лучше, чем при затенении в условиях леса.

Рис. 75. Влияние условий произрастания на клевер розовый; слева - на открытом месте; справа - в луговом травостое
Для уяснения причин угнетения растений при произрастании их в растительных сообществах необходимо рассмотреть сложные взаимоотношения, возникающие между растениями при их совместном произрастании.
Из всех форм взаимодействия между растениями в растительных сообществах наибольшее значение имеет изменение среды под влиянием жизнедеятельности самих растений. Эти изменения происходят в результате того, что каждое растение в процессе своей жизнедеятельности либо использует солнечную энергию, воду, элементы минерального питания и пр., в связи с чем обеспеченность соседних растений соответствующими условиями снижается; либо выделяет в окружающую среду продукты обмена веществ. Помимо того, большое значение в изменении среды имеет, во-первых, поступление отмерших органов растений в почву и на ее поверхность; во-вторых, создание особого фитоклимата в результате образования надземными органами растений более или менее сомкнутого покрова.
Потребление растениями необходимых для их жизни энергии и веществ создает в растительных сообществах условия, при которых не хватает наличных ресурсов для удовлетворения потребностей в них всех растений, входящих в состав сообщества. Возникает состояние, получившее название конкуренции за свет, воду, минеральное питание и т. д. Растения, обладающие какими-либо приспособлениями, позволяющими им более полно использовать необходимые для их жизни ресурсы или какой-либо из них (например, свет), имеют преимущества перед растениями, не обладающими этими приспособлениями. Первые растения более конкурентоспособны, чем другие. Конкурентная способность растений может изменяться от одного растительного сообщества к другому, в зависимости от условий произрастания. Она также зависит от воздействия на растения других компонентов сообществ (паразитных растений, животных-фитофагов и т. д.). Поражение паразитными грибами и бактериями, повреждение животными, ослабляя растения, снижают их конкурентную способность и косвенно повышают конкурентную способность их неослабленных соседей.
Каждое растение в процессе своей жизни выделяет в окружающую среду (в атмосферу, в почву) продукты своей жизнедеятельности, в том числе различные органические соединения. Продукты обмена веществ выделяют все органы растений, в особенности корни. Содержащиеся в выделениях вещества, в том числе и высокомолекулярные, могут поглощаться соседними растениями.
В многочисленных опытах, проведенных в лабораторных условиях, в чашках Петри, в вазонах, на искусственных средах (фильтровальная бумага, дистиллированная вода, песок), было установлено, что в этих условиях прижизненные выделения одних растений могут оказывать значительное влияние на другие растения. Однако результаты таких опытов нельзя переносить на условия, имеющиеся в природных растительных сообществах.
В настоящее время нет достоверных фактов о непосредственном токсическом влиянии корневых выделений одних растений на другие растения в природных сообществах. Для объяснения этого явления следует иметь в виду, что состав корневых выделений и их количество, по-видимому, специфичны для каждого вида, а в пределах вида - для особей различного возраста. Этот состав зависит и от условий произрастания. В частности, при отклонении условий жизни от оптимальных выделение активных веществ может возрастать. Кроме того, неоднократно высказывалось положение о токсичности продуктов метаболизма растений для особей того же вида, и в связи с этим весьма вероятно, что корневые выделения растений содержат вещества токсичные для особей того же вида и других. А так как почва, в отличие от водной и воздушной среды, характеризуется неподвижностью, то сухопутные растения могли существовать лишь при условии детоксикации их корневых выделений, а она в основном осуществляется микроорганизмами.
В процессе естественного отбора формировались связи наземных растений с микроорганизмами, массово обитающими в непосредственной близости к корням - в ризосфере, способными к детоксикации корневых выделений. В растительных сообществах корни особей отдельных видов обычно входят в контакт с корнями особей других видов. Поэтому естественно, что формирование видового состава сообществ происходило на основе подбора видов, способных произрастать совместно, включая подбор видов, в ризосферах которых имеются микроорганизмы, способные к детоксикации метаболитов не только данного вида, но и других видов, произрастающих вместе с ним. Этим можно объяснить отсутствие или крайнюю скудость данных о токсическом влиянии корневых выделений одних растений на другие в природных условиях.
Корневые выделения отдельных видов растений, оказывая специфическое воздействие на микроорганизмы, в том числе на азотобактер, денитрификаторы и прочие, косвенно влияют на взаимоотношения между видами в сообществах, определяя их конкурентную способность.
Среди прижизненных выделений растений особое место занимают органические вещества, вымываемые из надземных органов растений. В ряде случаев эти вещества оказывают токсическое влияние на соседние растения. Например, вещества, вымываемые из листьев горькой полыни, отрицательно влияют на произрастающие совместно с ней растения (фенхель, любисток, перечную мяту и др.). Но это бывает не всегда. Кроме того, во многих случаях токсическое влияние веществ, вымываемых из листьев, наблюдается лишь для видов, которые совместно в природных условиях не произрастают.
Большое значение в определении взаимоотношений между видами растений может иметь опад (отмершие части растений, поступающие в почву и на ее поверхность). В сообществах, не использующихся или лишь эпизодически использующихся человеком (например, в лесах), опад может скапливаться в больших количествах, образуя подстилку. В результате жизнедеятельности сапротрофных организмов (животных, грибов, бактерий, актиномицетов) происходит минерализация отмерших остатков растений; при этом продукты жизнедеятельности некоторых сапротрофных грибов могут оказывать токсическое влияние на сосудистые растения.
Особую форму взаимоотношений между растениями в сообществах представляют паразитизм, симбиоз. Органы паразитов проникают внутрь тканей хозяина, в его межклетники и даже в клетки. Иногда тело паразита целиком располагается в теле хозяина. Аналогичное взаиморасположение бывает и при симбиозе. Тесный контакт паразита с хозяином или двух симбионтов создает возможность прижизненного обмена веществ между ними. Паразитирование одного растения на другом широко распространено в растительных сообществах. Среди паразитных растений различают настоящие (полные) паразиты и полупаразиты. Первые получают от автотрофов и энергию и вещества (включая воду и элементы минерального питания). Вторые имеют зеленые листья, способные к фотосинтезу, и от растений-хозяев получают лишь воду и минеральные вещества, а иногда и энергетический материал. Между настоящими паразитами и полупаразитами имеются переходные формы.
Наиболее широко распространены паразитные грибы и бактерии. Вероятно, нет ни одного растительного сообщества, в котором бы они отсутствовали. Они поселяются на надземных и подземных органах древесных и травянистых растений.
В состав некоторых растительных сообществ входят и цветковые паразиты. Известно около 2000 видов цветковых полупаразитов, относящихся к 8 семействам и 83 родам, и свыше 500 видов настоящих паразитов из 9 семейств и 52 родов. Особенно много видов полных паразитов в семействах раффлезиевых, баланофоровых, заразиховых и повиликовых. Полные цветковые паразиты наиболее широко распространены в тропических странах. В СССР паразитных цветковых растений относительно немного, наиболее часто из них встречаются повилики и заразихи, паразитирующие главным образом на травах, реже на кустарниках (некоторые повилики).

Рис. 76. Подберезовики в березовом лесу
Основную часть полупаразитных растений составляют представители семейств ремнецветных (около 1000 видов), норичниковых (около 500 видов) и санталовых (около 400 видов). В СССР наиболее широко распространены полупаразиты из семейства норичниковых, как однолетние (погремки, очанки, марьяники и др.), так и многолетние (большинство видов мытников, кастиллеи). На деревьях в юго-западных и южных районах нередко поселяется вечнозеленый полупаразит - омела.
В одном и том же сообществе могут быть виды и особи, и пораженные паразитами, и вовсе ими не пораженные. Соответственно и конкурентная способность одних видов или особей возрастает, а других снижается.
В растительных сообществах широко распространены симбиотические (взаимно полезные) взаимоотношения между растениями. В зависимости от того, какой организм вступает в симбиотические отношения с сосудистыми растениями, различают микотрофию (симбиоз с грибом), бактериотрофию (симбиоз с бактериями или актиномицетами), альготрофию (симбиоз с водорослью).
При микротрофии гриб поселяется на корнях растений (снаружи корня или внутри его клеток); в отличие от грибных паразитов, микоризные грибы оказывают на сосудистые растения положительное влияние, выраженное более или менее заметно. Особенно благоприятно микоризные грибы, представленные шляпочными (подберезовик, подосиновик, белый гриб, масленок и др.), влияют на деревья (сосну, ель, дуб, бук и т. д.). Деревья, находящиеся в симбиотических отношениях с грибом, растут значительно лучше (рис. 76).
Менее широко в растительных сообществах распространена бактериотрофия. Она наиболее хорошо изучена у бобовых, на корнях которых бактерии, фиксирующие атмосферный азот, образуют клубеньки. Благодаря симбиозу с клубеньковыми бактериями бобовые обеспечены необходимым для них азотом. При отмирании клубеньков и корней бобовых содержащийся в них азот после минерализации отмерших органов становится доступным для растений, произрастающих совместно с ними. Аналогичные клубеньки в результате симбиоза с актиномицетами, способными фиксировать атмосферный азот, образуются на корнях ольхи, лоха, облепихи и др. Фиксация азота, особенно в ольховых лесах, происходит в значительных количествах, что создает условия для произрастания видов, особенно требовательных к обеспеченности азотом, например двудомной крапивы.
Симбиоз азотфиксирующих организмов с сосудистыми растениями может также проявляться в форме образования "желвачков" на листьях некоторых тропических растений, относящихся к семействам мареновых, дискорейных и др.
Примером альготрофии может служить симбиоз водного папоротника азолли с сине-зеленой водорослью анабеной, способной фиксировать атмосферный азот. Этот папоротник используется во Вьетнаме в посевах риса для повышения его урожая благодаря повышению обеспеченности культуры азотом.
На стволах и ветвях древесных растений нередко поселяются другие растения (водоросли, мхи, лишайники, папоротники, цветковые). Древесные растения служат для них субстратом для прикрепления, но не являются источником энергии и минеральных веществ. Такие растения получили название эпифитов. Взаимоотношения эпифитов с теми растениями, на которых они поселяются, - пример комменсализма, т. е. явления, когда один из партнеров получает пользу от связи с другим, а для второго партнера в этой связи нет ни пользы, ни вреда.
Эпифиты наиболее широко распространены в тропических лесах, где, помимо многочисленных травянистых эпифитов (папоротников, орхидных, бромелиевых), встречаются и кустарниковые эпифиты. В СССР из эпифитов широко распространены лишайники и частично мхи, поселяющиеся на нижних частях стволов деревьев. Эпифиты-лишайники наиболее обильно разрастаются на ветвях старых или вообще угнетенных деревьев. Это дает основание предполагать, что достаточно молодые и неугнетенные деревья обладают какой-то приспособленностью, препятствующей поселению на них лишайников-эпифитов, и что это свойство утрачивается при старении или угнетении деревьев. Нет сомнения, что обильное поселение эпифитов на ветвях деревьев может оказывать на них отрицательное влияние: оно увеличивает нагрузку на ветви; эпифиты конкурируют с древесными растениями за углекислоту; повышение влажности воздуха способствует поражению древесных растений грибными и бактериальными паразитами.
В тропических лесах, где широко распространены лианы - растения со стеблями, нуждающимися в опоре, чтобы вынести свои листья вверх, имеет ценотическое значение взаимоотношение между ними и деревьями.
При формировании биогеоценозов в результате жизнедеятельности многочисленных организмов, входящих в состав биоценоза, абиотическая среда изменяется и экотоп превращается в биотоп. Характерной особенностью биотопа является его неоднородность по вертикали и по горизонтали, а также динамичность во времени. Последняя связана с изменениями метеорологических и гидрологаческих условий в течение каждого года и ряда лет; косвенно этим обусловлены колебания численности биокомпонентов, в особенности животных, деятельность которых имеет большое средообразующее значение.
Неоднородность и динамичность среды в большинстве растительных сообществ создает условия, при которых ни один вид растений не может полностью использовать наличные ресурсы энергии и вещества и тем самым исключить возможность участия в сообществах других видов растений. Как следствие этого в течение очень длительного периода времени шел отбор видов растений и животных, включая микроскопические организмы, способные существовать совместно в определенных условиях среды. Создавались биогеоценозы, в которых каждый вид занимал определенное место - экологическую нишу. При этом количественные соотношения видов изменялись в определенных пределах в соответствии с изменением условий их обитания в течение года и год от года. Формировались растительные сообщества из растений, использующих различные горизонты среды (надземной и подземной), с разной интенсивностью в отдельные годы и периоды года.
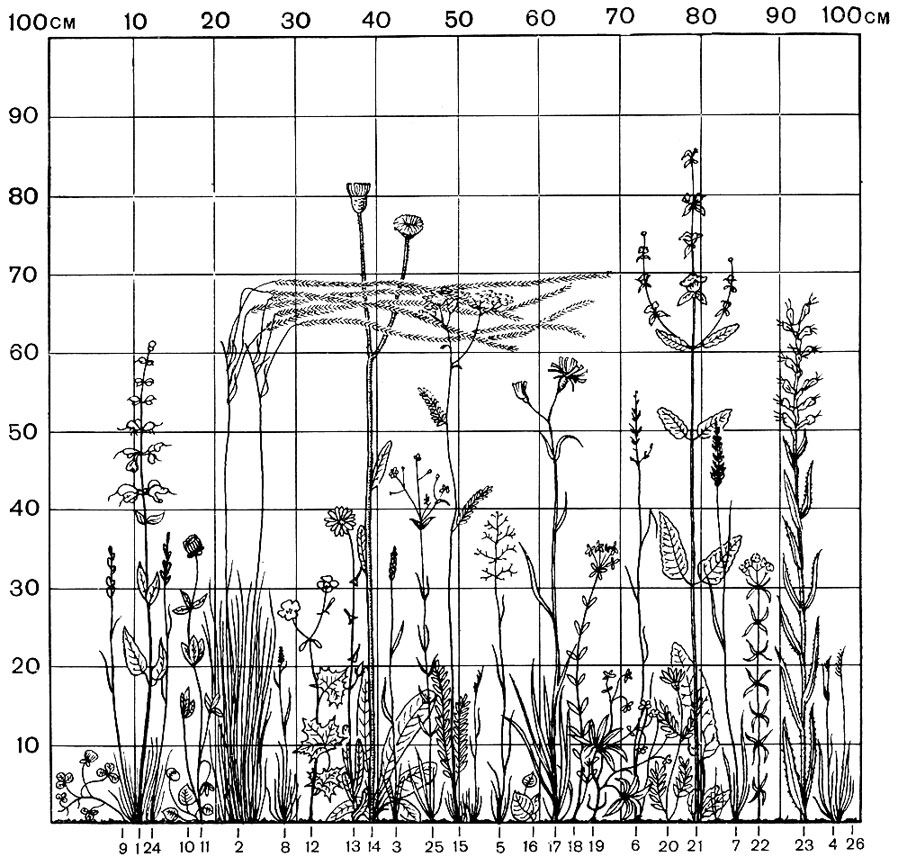
Рис. 77. Вертикальная проекция травостоя остепненного луга (Бобрпнская степь в Орловской области). Деюновинные злаки и осоки: 1 - типчак; 2 - ковыль Иоанна; 3 - душистый колосок; 4 - горная осока. Корневищные злаки и осоки: 5 - полевица Сырейщикова; 6 - красная овсяница; 7 - келерия Делявиня; 8 - русская осока. Разнотравье (включая бобовые): 9 - ползучий клевер; 10 - песчаная фиалка; 11 - горный клевер; 12 - многоцветковый лютик; 13 - обыкновенный нивяник; 14 - крапчатый пазник; 15 - шестилепестная таволга; 16 - собачья фиалка; 17 - пурпуровый козелец; 18 - стройный молочай; 19 - семилисточковая лапчатка; 20 - датский астрагал; 21 - клубненосный зопник; 22 - северный подмаренник; 23 - красная румянка; 24 - луговой шалфеи; 25 - злаколистная песчанка. Мхи: 26 - зеленый мох (сплошной покров)
Легче всего обнаружить различия в использовании среды различными видами и группами растений в надземной части растительных сообществ, поскольку в большинстве случаев в их состав входят растения разной высоты (рис. 77, 78). Это особенно хорошо заметно в лесах, где растительные сообщества образованы не только деревьями, но и кустарниками, травами, а нередко еще мхами и лишайниками. Во многих лесных сообществах, особенно в умеренных широтах, растения какого-либо вида или группы видов располагают свои листья в определенном интервале высоты над уровнем почвы, образуя достаточно четко оформленные и достаточно сомкнутые ярусы. Примером может служить расчленение еловых лесов на древесный, травяно-кустарничковыи и моховой ярусы.
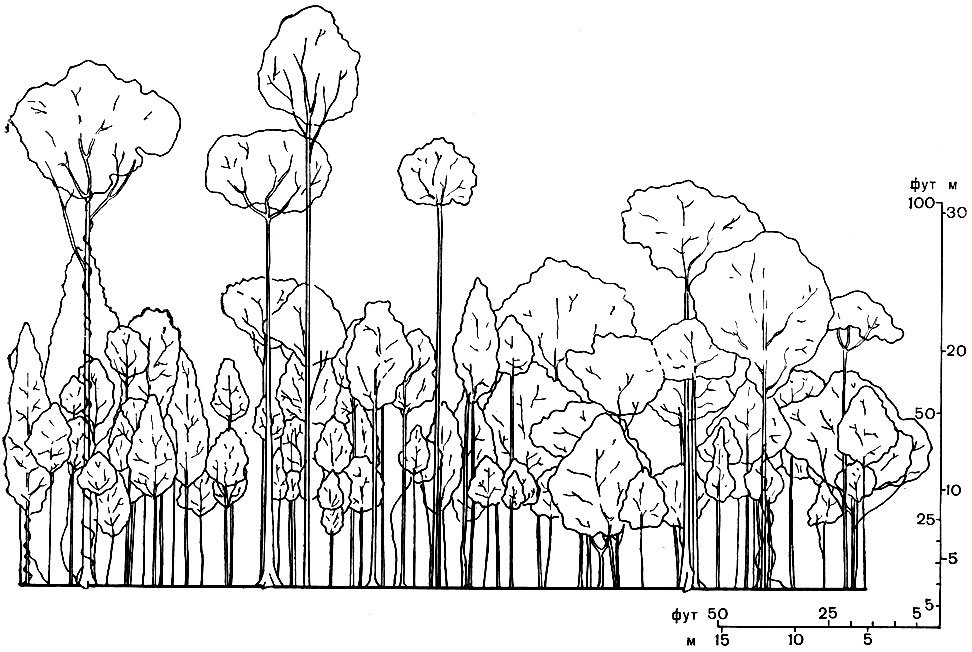
Рис. 78. Профильная диаграмма тропического леса на острове Калимантан (Борнео). Изображена полоса леса длиной около 60 м и шириной около 8 м. Показаны деревья высотой более 7 м
Ярусность наиболее хорошо выражена, в сообществах, образованных растениями, относящимися к разным жизненным формам, имеющим различную форму роста. Например, это бывает в лесах при совместном произрастании деревьев, кустарников, трав и других растений. Ярусность слабо выражена или вовсе не выражена в сообществах, где все растения относятся к одной жизненной форме, например во многих травяных сообществах (степях, лугах).
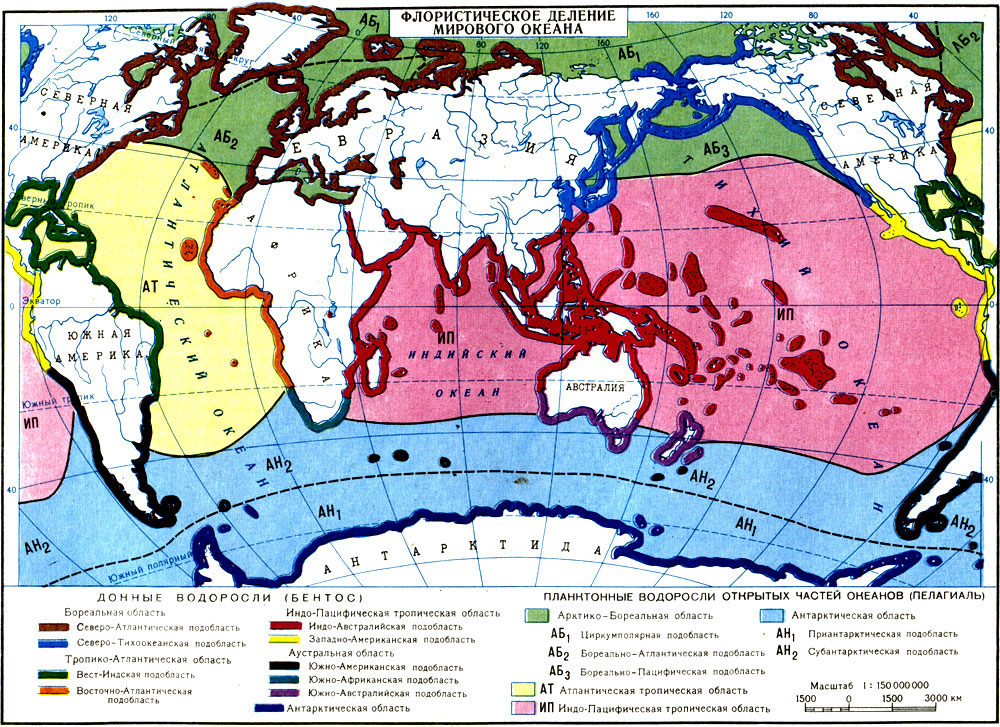
Флористическое деление Мирового океана
Во всех растительных сообществах (за редким исключением), даже там, где нет четкого расчленения на ярусы, всегда заметны различия в высоте надземных органов растений и горизонты, различающиеся по насыщенности листовой поверхности. Этим обусловливается более полное использование солнечной энергии.

Таблица 21. Дикие полезные растения: вверху слева - дикий лук; вверху справа - лимонник; внизу слева - женьшень; внизу справа - белена

Таблица 22. Культурные растения: вверху слева - слива; вверху справа - какао; внизу персик
В надземной части растительных сообществ ют верхнего уровня по направлению к поверхности почвы происходит изменение среды: снижается интенсивность света и изменяется его состав в связи с перехватом его выше расположенными листьями; повышается содержание CO2, так как основным источником углекислоты является выделение ее почвой (дыхание почвы); увеличивается влажность воздуха; изменяется температура. В горизонтах максимального скопления листьев объем воздуха на единицу площади листьев меньше, чем там, где насыщенность листьями незначительна. В соответствии с этим листья растений различной высоты находятся в разных условиях по обеспеченности их светом, углекислотой и т. п. Естественно, растения, располагающие листья вблизи поверхности почвы, характеризуются большей теневыносливостью и влаголюбивостью. Их листья обычно содержат больше хлорофилла, что в сочетании с лучшей обеспеченностью углекислотой дает им возможность существования в условиях слабого освещения. Высокие растения оказывают влияние на низкорослые, перехватывая свет. Однако и растения низкорослые влияют на высокорослые, используя углекислоту, воду и элементы минерального питания. Это можно наблюдать в лесах, где удаление травяно-кустарничкового покрова приводит к резкому увеличению продуктивности деревьев.

Таблица 23. Культурные растения: вверху слева - фисташка; вверху справа - яблоня; внизу - лимон

Таблица 24. Культурные растения: вверху (слева направо) - хлопчатник, подсолнечник, красный перец; внизу - кукуруза
Растения разных видов, входящих в состав растительных сообществ, обычно значительно различаются по глубине укоренения. С этим иногда связано совместное произрастание особей, отличающихся по их отношению к влажности почвы. Например, у ксерофитов ковылей корневые системы располагаются в верхнем слое почвы; у гигрофита тростника корни проникают вглубь до уровня почвенно-грунтовых вод. Однако у всех растений, вне зависимости от глубины их укоренения, часть их корней (иногда функционирующих непродолжительное время) располагается в самом верхнем горизонте почвы, наиболее богатом элементами минерального питания растений, в особенности азотом.

Таблица 25. Культурные растения: слева вверху - масличная пальма; слева в середине - сахарный тростник; слева внизу длиннолистная агава; справа вверху - кофейное дерево; справа внизу - дынное дерево

Таблица 26. Леса: вверху слева - березовая роща; вверху справа - широколиственный лес; внизу - сосновый бор
Наряду с вертикальным расчленением сообществ существует расчлененность их в горизонтальном направлении на пятна, отличающиеся по количественному соотношению разных видов растений, по сомкнутости, продуктивности и другим свойствам. Такая неоднородность сообществ получила название мозаичности. Она в основном обусловлена неоднородностью в экотопе, деятельностью животных, средообразующим влияниом растений, воздействием человека.

Таблица 27. Пойма и степь: вверху - пойма горной реки (истоки Колымы); внизу - разнотравная степь (видны березовые колки)
Примером мозаичности, обусловленной неоднородностью в экотопе, могут быть изменения в пределах сообществ, связанные с наличием небольших понижений и повышений. То же самое происходит в результате деятельности животных-землероев, например кротов.
При заселении кротовин растениями создаются пятна, отличающиеся от окружающей их растительности, и только постепенно, спустя длительный период времени, эти различия стираются.
Любое растение в своей жизнедеятельности изменяет условия произрастания других растений. При этом мощные растения оказывают большее средообразующее влияние, чем маломощные. Группы особей одного и того же вида влияют на другие растения интенсивнее, чем единичные особи. Поэтому мозаичность, связанная с жизнедеятельностью растений, особенно ярко проявляется там, где в состав сообществ входят деревья или травы, образующие куртины, возникающие при вегетативном размножении или неравномерном обсеменении. В пределах лесных сообществ количественные соотношения видов нижних ярусов в непосредственной близости стволов деревьев иные, нежели в пространстве между стволами. То же наблюдается в пределах куртин травянистых растений и вне их. Мозаичность может также возникать в результате деятельности человека (например, при выборочной рубке леса).
Растительные сообщества подвержены непрерывным изменениям, зависящим от разных причин: особенностей сезонного ритма и жизненного цикла растений, воздействия человека и животных, изменения условий произрастания. При этом наряду с изменением состояния растительных сообществ может происходить смена одного растительного сообщества другим (сукцессия).
Различают смену состояний растительных сообществ по сезонам года (сезонную изменчивость) и в течение ряда лет, в связи с различиями в метеорологических и прочих условиях отдельных лет (разно-годичную, или погодичную, изменчивость - флюктуацию).
На большей части земного шара (а в пределах нашей страны повсюду) в течение года происходят существенные изменения в световом, температурном и гидрологическом режимах и в соответствии с этим изменяются условия обитания растений в растительных сообществах, а животных в тех биоценозах, в состав которых входят определенные растительные сообщества. Это было и в прошлом. Поэтому в процессе формирования растительных сообществ происходил подбор видов, способных совместно существовать в изменяющихся по сезонам года условиях произрастания, отличающихся по особенностям ритма сезонной вегетации, по времени прохождения соответствующих фенологических фаз.
Подбор видов с различным ритмом сезонной вегетации определяет возможность совместного существования в сообществах большого числа видов и более полного использования растениями среды. Помимо того, это ведет к снижению интенсивности конкуренции между видами в сообществах. Особенности ритма сезонной вегетации отдельных видов растений проявляются в различиях в смене фенологического состояния - фенологических фаз - в течение вегетационного сезона. Благодаря этому в сообществах, образованных видами с различным ритмом сезонной вегетации, в течение года происходят изменения их внешнего вида - смена аспектов.
Аспект - внешний вид сообществ -определяется как вегетативными, так и генеративными органами растений. Например, в лесах аспект в основном определяется листвой деревьев; на моховых болотах - особенностями внешнего вида мхов; в тростниковых зарослях - внешним видом высоких надземных побегов тростника и в меньшей мере его соцветиями. Смена аспектов наиболее четко проявляется там, где в течение вегетационного сезона происходит смена массово цветущих видов растений. Это особенно заметно на многих типах лугов и степей, где в течение вегетационного сезона можно различать до 8-10 и более аспектов - фенологических стадий. В таких сообществах, где преобладающие виды имеют многолетние надземные органы, внешний вид некоторых не изменяется в течение вегетационного сезона, смена аспектов не выражена. Примером может служить сосновый лес с напочвенным покровом из кустистых лишайников (кладоний).
По вегетационному сезону изменяется не только внешний вид сообществ, но и (что более существенно) количественные соотношения между видами, воздействие отдельных видов и в целом сообщества на среду, продуктивность сообщества. В зависимости от состава растительного сообщества количественные соотношения его компонентов изменяются в течение вегетационного сезона более или менее заметно. Особенно велики изменения в количественном соотношении видов растений в травяных сообществах и в травяных ярусах лесных сообществ. В некоторых из них по вегетационному сезону происходит смена доминирующих растений. Особенно хорошо это заметно в таких сообществах, в состав которых входят виды с коротким периодом вегетации, приуроченным к весне: либо однолетние - эфемеры, либо многолетние - эфемероиды.
Эфемеры и эфемероиды преобладают там, где от весны к лету происходит резкое изменение условий произрастания, связанное с обеспечением растений водой или светом. Примером может служить пышное развитие ранней весной таких растений, как мятлик луковичный и осока толстостолбиковая в некоторых районах Средней Азии, когда условия увлажнения и температуры складываются благоприятно для их жизни. По аспекту такие сообщества в это время напоминают луга. Но как только наступает засуха с высокой температурой, от эфемеров остаются лишь семена, успевшие осыпаться на поверхность почвы, а у эфемероидов сохраняются лишь подземные органы, пребывающие в состоянии покоя до наступления благоприятных условий для их вегетации. То, что ранее напоминало луг, приобретает вид пустыни.
Аналогичные изменения, но вызванные изменением других условий обитания, происходят в некоторых типах коренных листопадных лесов, например в дубовых. Здесь ранней весной (до развертывания листьев дуба) в травяном покрове преобладают эфемероиды (хохлатка, ветреница, пролеска), успевающие закончить сезонный цикл развития к началу резкого изменения в световом режиме после полного развертывания листьев дуба, после чего в травяном ярусе начинают преобладать виды летней вегетации (сныть обыкновенная и др.). По вегетационному сезону происходит возникновение и отмирание особей, а также побегов и отдельных органов растений. В связи с различиями в новообразовании и отмирании по сезонам меняется масса надземных и подземных органов отдельных видов и в целом по сообществу. Наиболее заметные изменения происходят в массе недолговечных надземных органов растений: листьев деревьев с ежегодно опадающей листвой, надземных побегов травянистых растений.
В травяных сообществах от весны к осени вначале происходит более или менее быстрое увеличение массы надземных побегов, а затем то постепенное, то быстрое снижение продуктивности травостоев в связи с отмиранием листьев и целых побегов, осыпанием семян и т. д. Сезонные изменения в продуктивности травяных сообществ имеют большое значение для выбора сроков использования их как кормовых угодий.
Большое значение в жизни сообществ имеют сезонные изменения в появлениях всходов, в их отмирании, в переходе особей растений из одного возрастного состояния в другое.
Любой год своеобразен по метеорологическим и гидрологическим условиям. Периодически повторяются годы с особенно обильными атмосферными осадками либо особо засушливые или с малоснежной морозной зимой и т. д. В поймах рек год от года изменяется длительность заливания полыми водами и мощность отлагаемого наилка. Все это определяет изменения по годам условий произрастания растений и влияет на условия обитания животных, входящих в состав биоценозов, на изменение численности животных. Периодически численность некоторых видов животных резко возрастает, что оказывает большое непосредственное (при поедании животными растений) и косвенное (через изменение условий произрастания, например в результате деятельности землероев) влияние на растения.
Погодичные изменения растительных сообществ (флюктуации) могут быть то более, то менее значительными. Для них характерны устойчивость флористического состава, отсутствие выпадения одних видов и внедрение других, что происходит при смене одного сообщества другим. Количественные соотношения видов травянистых растений по годам колеблются то в большей, то в меньшей степени, так же как и продуктивность надземных и подземных органов.
Флюктуации (погодичные изменения) в отличие от сукцессий характеризуются ненаправленностью своих изменений.
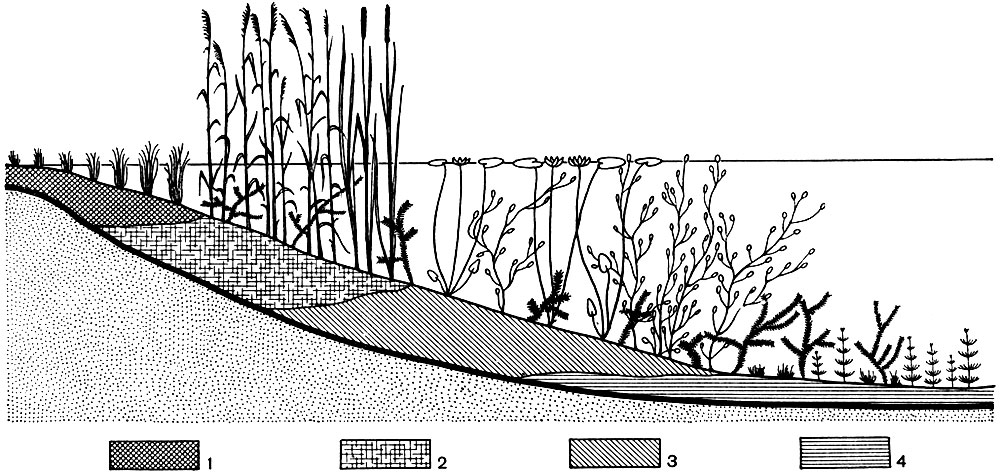
Рис. 79. Схема зарастания озера. Растительность (начиная от берега): осоки, тростник, камыш и рогоз (с примесью погруженных в воду растений); кувшинки; кубышки и другие растения с плавающими листьями; рдесты и другие погруженные в воду растения; донные мхи и водоросли (глубоководная часть озера без высших растений). 1 - осоковый торф; 2 - тростниковый и камышовый торф; 3 - сапропелевый торф; 4 - сапропелит
В разные годы, в зависимости от причин, вызывающих изменения, они могут быть направлены то в одну, то в другую сторону: в годы с большим количеством атмосферных осадков в сообществах увеличивается участие видов, более требовательных к влаге; в засушливые годы, наоборот, возрастает роль видов, устойчивых к недостатку воды, и т. д. После устранения причины, вызвавшей изменения, сообщество обычно довольно быстро возвращается к состоянию, близкому к исходному. Практическая обратимость погодичных изменений - важнейший признак, отличающий их от сукцессий. Погодичные изменения особенно отчетливо выражены в травяных сообществах (луга, степи), в местах с изменчивым по годам гидрологическим режимом (поймы, западины), в районах, где метеорологические условия достаточно резко изменяются от года к году. Наиболее полно эти изменения изучены на лугах, в степях и пустынях. Некоторые погодичные изменения малозаметны на глаз или выражаются в периодической взаимной смене преобладающих растений. В одни годы преобладает один вид (или одни виды), в другие - другой вид (или другие виды). Однако иногда, например при длительном застое воды в пойме, при суровой малоснежной зиме, последовавшей за очень засушливым летом, в результате резкого увеличения численности растительноядных животных происходит массовое отмирание преобладающих видов растений. Вместо них начинают преобладать виды, способные быстро занимать освободившиеся места, обычно размножающиеся вегетативно, а нередко к тому же имеющие большой запас всхожих семян в почве. Эти виды, названные Л. Г. Раменским эксплерентами (выполнителями), не обладают большой конкурентной способностью. Поэтому, как только причина, вызвавшая отмирание ранее преобладавших видов, перестает действовать, эти виды вновь начинают преобладать, а участие эксплерентов сильно снижается.
Погодичная изменчивость сообществ имеет большое значение для разработки рациональных приемов использования и улучшения кормовых угодий, поскольку по годам изменяются урожайность и качество урожая, а также эффективность отдельных приемов улучшения (внесения удобрений, подсева и пр.).
Ни одно сообщество не существует вечно, рано или поздно оно сменяется другим сообществом. Это происходит при воздействии внешних причин либо в результате изменения среды в связи с жизнедеятельностью организмов, образующих биоценозы, в том числе при внедрении в сообщества новых видов. Среди многообразных форм смен сообществ различают первичные и вторичные сукцессии. Первичные сукцессии представляют собой естественную смену сообществ на территориях, ранее не занятых растительностью, например на песчаных отмелях в поймах рек, в местах, освободившихся после отступления ледников, и пр. В зависимости от субстрата (его физических и химических свойств) здесь вначале поселяются либо только бактерии, водоросли и лишайники, либо наряду с ними сосудистые растения. Постепенно создается сомкнутый растительный покров, начинает формироваться почва, в которой происходит накопление органического вещества и элементов минерального питания, в частности азота, создаются условия для формирования растительных сообществ, типичных для данной природной зоны и данных условий рельефа, увлажнения и т. п. (например, хвойных лесов для таежной зоны, степей для степной зоны и т. д.). При нарушении сообществ, возникших при первичных сукцессиях, происходит смена сообществ в направлении возврата к состоянию, близкому к исходному, - вторичные сукцессии.
В настоящее время первичные сукцессии происходят на относительно небольших площадях. Основное значение имеют вторичные сукцессии. Из причин, вызывающих вторичные сукцессии, за последние десятилетия особенно большое значение имеет деятельность человека (вырубка леса, осушение болот, выпас скота и т. д.). В зависимости от формы воздействия человека эти смены происходят либо быстро, катастрофически (вырубка леса), либо постепенно (при внесении удобрений на луга). Воздействие человека может выражаться в непосредственном воздействии на растительность (вырубка леса) без изменения экотопа или в изменении среды без непосредственного воздействия на растительность (осушение болот).
При изменении условий произрастания, если измененные условия впоследствии поддерживаются человеком (при ежегодном внесении удобрений на луга), формируются новые, достаточно устойчивые сообщества.
При уничтожении растительности без изменения почвенно-грунтовых условий происходит смена сообществ в направлении возврата к состоянию, характеризующему исходный коренной тип. Например, при уничтожении хвойных лесов на вырубках или гарях вначале разрастаются травы (вейники, иван-чай и др.), а затем постепенно формируются насаждения из мелколиственных пород (береза, осина), под пологом которых поселяются ель или другие хвойные, которые впоследствии выходят в первый ярус и образуют растительные сообщества, сходные с существовавшими до нарушения.
Изучение смен сообществ под влиянием воздействия человека имеет большое теоретическое и практическое значение для разработки мер охраны природы и рационального использования природных растительных ресурсов.
Классический пример смены сообществ под влиянием жизнедеятельности организмов - это процесс зарастания озер. В воде любого озера, в особенности если оно богато азотом и зольными элементами, обитает огромное количество микроскопических организмов (водоросли, простейшие и др.). Отмирая, они падают на дно вместе с мелкоземом, приносимым в озеро со склонов. Этот процесс, повторяясь из года в год, приводит к образованию на дне озера сапропеля, к уменьшению глубины озера, к проникновению солнечного света до дна озера. В результате создаются условия для поселения мхов и многоклеточных водорослей, что обусловливает ускорение накопления на дне озера органических остатков (сапропелевый торф) и приводит к еще большему обмелению водоема. А это сопровождается поселением сосудистых растений с погруженными в воду побегами или с плавающими на поверхности воды листьями (рдесты, кувшинки, кубышки и др.). Следующей стадией зарастания озера является поселение озерного камыша и обыкновенного тростника, развивающих огромную массу надземных побегов, из которых после их отмирания образуется камышовый или тростниковый торф. При дальнейшем заполнении озера отмершими остатками растений и его обмелении поселяются осоки. Озеро постепенно превращается в болото. Наблюдая отдельные пояса водной растительности на зарастающем озере, можно восстановить основные стадии его зарастания - превращение в болото.
В настоящее время человек на огромной площади (свыше 1,4 млрд. га) создал искусственные сообщества путем посева и посадки растений в целях получения зерна, овощей, фруктов, корма для скота, сырья для промышленности и т. д. В посевах и посадках, созданных человеком, есть основные признаки, характерные для природных растительных сообществ: влияние растений друг на друга и на среду, создание особой, свойственной этим сообществам фитосреды.
Таким образом, современный растительный покров Земли состоит не только из фитоценозов (природные сообщества), но и агроценозов (искусственные сообщества).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'