Порядок пандановые (Pandanales) (И. В. Грушвицкий)
Семейство пандановые (Pandanaceae)
В этом семействе, по данным известного специалиста Б. Стоуна, более 800 видов, относящихся к 3 родам. Большая часть видов, 600-700, относится к роду панданус (Pandanus), примерно 175 - к роду фрейсинетия (Freycinetia) и всего 2 вида представляют род сараранга (Sararanga).
Пандановые - вечнозеленые древовидные растения. Внешне многие из них несколько напоминают и драцены, и юкки, и пальмы, и даже некоторые двудольные. Стебли их простые или слабо ветвящиеся. Листья расположены на концах стеблей или их ветвей плотными пучками и снабжены стеблеохватывающими влагалищами, как бы вставлены один в другой. Они расположены на стебле тремя или четырьмя рядами и в то же время винтообразно, следуя закручиванию верхушки стебля (рис. 251). С этим связано употребляемое иногда название пандановых - "винтовые пальмы". Сами листья в большинстве случаев крупные, кожистые, цельные, колючие по краям и снизу вдоль средней жилки.
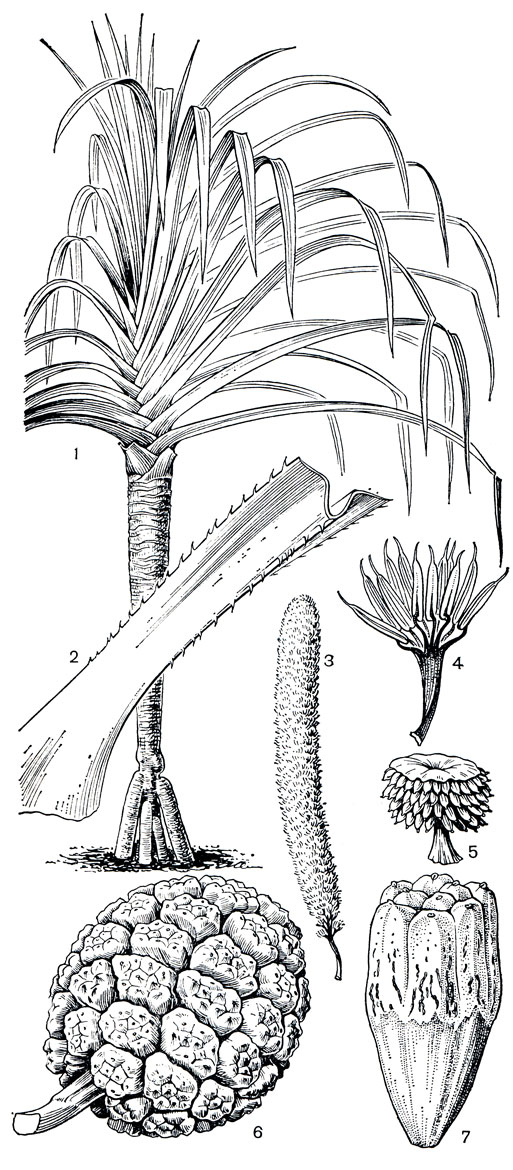
Рис. 251. Пандановые. Панданус вильчатый (Pandanus furcatus): 1 - общий вид; 2 - фрагмент листа; 3 - мужской початок; 4 - колонка с тычинками. Панданус крупноплодный (P. macrocarpus): 5 - щитовидная мужская фаланга. Панданус кровельный (P. tectorius): 6 - соплодие; 7 - женская фаланга из сросшихся костянок
В роде панданус у большой части видов имеются ходульные корни. Виды сараранги - панданусовидные "деревья", но без ходульных корней и с листьями, расположенными не тремя спиральными рядами, как у панданусов, а четырьмя. Виды фрейсинетии являются корнелазающими лианами, напоминающими тропические лиановидные аронниковые.
Пандановые - типичное палеотропическое семейство. Их ареал (карта 15) охватывает тропические области Африки, Мадагаскар, другие острова Индийского океана, Индию, Индокитай, все территории Малезийской области, север и северо-восток Австралии, а также тропические и некоторые субтропические острова Тихого океана. Наиболее обширным и объемлющим является ареал рода панданус, границы которого практически совпадают с границами ареала всего семейства. Значительно меньшие размеры имеет область распространения фрейсинетии, отсутствующей в Африке, Индии и западной части Индийского океана; фрейсинетия выходит за границы ареала панданусов лишь в одном месте, где один ее вид встречается в Новой Зеландии. Наконец, самый маленький ареал у сараранги, два вида которой распространены только в Новой Гвинее, на Филиппинах, Соломоновых островах и островах Адмиралтейства. Именно эта небольшая островная область является одновременно частью центра наибольшего видового богатства и панданусов, и фрейсинетии, и, очевидно, также "колыбелью" происхождения пандановых в целом, поскольку здесь сосредоточены наиболее примитивные представители семейства.
Род панданус, однако, имеет и второй центр видового богатства - на Мадагаскаре, где насчитывается до 90 его видов, причем все они являются эндемиками этого острова. В то же время в пределах во много раз большей по площади африканской части ареала рода распространено всего около 20 его видов.
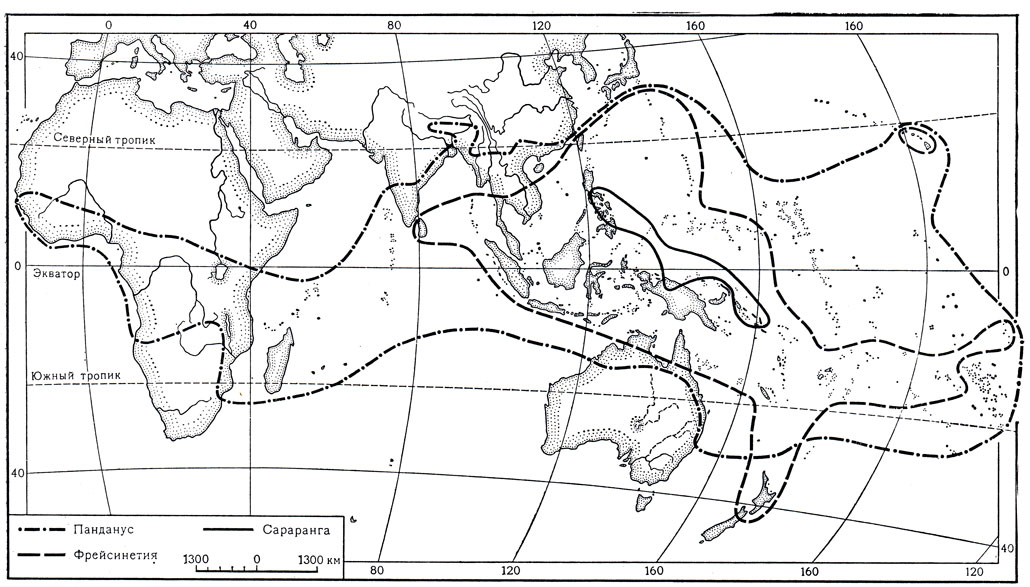
Карта 15. Ареалы родов панданус, фрейсинетия и сараранга
Род панданус наиболее разнообразен по приспособленности к условиям жизни. Многие его виды являются обитателями морских побережий, где на пологих берегах на песчаном или каменистом грунте, на прибрежных дюнах и утесах они нередко образуют целые заросли, подчас труднопроходимые. Таков, например, панданус кровельный (Pandanns tectorins, табл. 59, 1) - вид, широко распространенный на островах Индийского и Тихого океанов и от Южного Китая на севере до Австралии на юге. Этот сильно разветвленный "кустарник" с ходульными корнями образует заросли высотой от 3 до 7 м. В Индонезии и на берегах Новой Гвинеи такие заросли наряду с другими видами формирует низкорослый (высотой 2- 3 м) панданус многоглавый (P. polycephalus). Очень часто панданусы произрастают по краям тропических болот и на самих болотах. "Этот род,- отмечает автор обработки семейства во "Флоре Малезии", известный ботаник Ван Стеенис (1954),- проявляет почти универсальную приспособленность к различным условиям". Кроме побережий и болот, панданусы встречаются в заболоченных и более сухих низинных, горных и даже высокогорных лесах, на коралловых рифах и известняковых скалах, на склонах вулканов и по краю их кратеров, в саваннах, в целом же от уровня моря до высоты 3225 м. Нередки заросли панданусов по берегам рек, зарастают ими изолированные старицы, а также сами речные русла. Местами такие заросли тянутся на километры и настолько густы, что передвижение по реке на лодках становится невозможным. На Малакке и на Суматре причиной этого является, в частности, панданус винтовой (P. helicopus), чьи стебли с густыми тонкими ветвями лишь на одну треть возвышаются над водой. Чтобы расчистить водный путь, местным жителям приходится вырубать поверхностную часть этих зарослей. Подводным, настоящим гидрофитом является и панданус Пейръера (P. peyrierasii), образующий густые заросли в сублиторальной зоне пресноводных водоемов на юго-востоке Мадагаскара. Семена этого пандануса прорастают под водой, полностью под водой развиваются и молодые растения с погруженными листьями, и только у зрелых особей верхние листья и соцветия выступают из воды. В ходе приспособления к водному образу жизни здесь даже сложился листовой диморфизм. Погруженные листья очень узкие, с 32 или меньшим числом продольных жилок; надводные - линейно-эллиптические, шириной до 5 см и с 80- 82 жилками. Обширные сфагновые болота, которые можно назвать пандановыми, встречены на Северной Суматре и на острове Сулавеси на высоте 1700-1900 м.

Таблица 59. Пандановые: 1 - панданус кровельный (Pandanus tectorius), о. Мадагаскар; 2 - соплодие пандануса кровельного; 3 - панданус болотный (P. palustris), о. Маврикий; 4 - фрейсинетии Уокера (Freycinetia walkeri), Шри-Ланка; 5 - фрейсинетия крупноколосая (F. macrostachya), соцветия, Новая Гвинея; 6 - сараранга глубоковыемчатая (Sararanga sinuosa), соцветие женского растения, Соломоновы острова
Многочисленные виды пандануса входят в состав подлеска горных и высокогорных тропических лесов. Отмечается даже тенденция к увеличению их обилия по мере подъема в горы. Наряду с древовидными папоротниками панданусы часты в горных и моховых лесах, а также близ границы между последними и вышерасположенным поясом эрикоидных кустарников. В Индонезии, столь богатой вулканическими вершинами, отмечается частая встречаемость панданусов на склонах вулканов, по краям кратеров и вокруг сольфатар, где их стебли и листья бывают покрыты вулканическим пеплом.
Виды рода панданус из столь различных местообитаний разнообразны и по внешнему виду. Наиболее заметной чертой габитуса панданусов являются ходульные корни (рис. 251, табл. 59). Во всем мире цветковых не найти другого рода, где эта своеобразная структура была бы выражена у столь большого числа видов - не у единичных, даже не у десятков, а у сотен представителей. Не удивительно, что панданусы с ходульными корнями, к тому же первыми встречавшие на океанических побережьях исследователей тропической природы, уже давно, более 300 лет назад, стали объектом внимания ботаников. Ходульные корни прибрежных панданусов справедливо характеризовались как важное приспособление в условиях периодически сильных, а иногда и ураганных ветров. Способность этих корней-якорей противостоять нагрузкам как на разрыв, так и на сжатие (на излом) объясняли наличием в них механических тканей, с одной стороны, в кольце периферических проводящих пучков, с другой - в пучках, расположенных в центральной части корня. Позднее было показано, каким образом якорная функция ходульных корней проявляется и у панданусов, обитающих в местах, защищенных от сильных ветров. Дело в том, что для видов этого рода характерно не только отсутствие стержневого корня, но нередко также отмирание нижней надземной, всегда наиболее тонкой части ствола. В результате взрослый панданус держится на ходульных корнях действительно как на ходулях, а сами ходульные корни не только поддерживают ствол с листвой, но и монопольно обеспечивают растение водой и необходимыми питательными веществами.
Ходульные корни у панданусов прямые и неветвистые и лишь после внедрения в почву сильно разветвляются. Некоторые из придаточных корней, обычно из числа отходящих от ствола выше других, не достигают почвы и остаются воздушными с кончиком, расширенным за счет мощного корневого чехлика. По свидетельству Ван Стеениса, у высоких панданусов такие корневые окончания, покрытые наслоениями пробки, могут быть размером с кулак, образуя с поселяющимися на них папоротниками и другими эпифитами "висячие цветочные горшки".
У некоторых видов пандануса придаточные корни развиваются не от ствола, а от его ветвей; достигая почвы и укореняясь в ней, они образуют столбовидные корни. Так, у пандануса лабиринтного (P. labirinticus) в лесах Суматры, по словам монографа пандановых Отто Варбурга (1900), одна особь образует иногда "форменный лес". У других видов сочетаются и ходульные и столбовидные корни. Наконец, некоторые виды с типично пальмовидным габитусом, как, например, панданус превосходный (P. princeps), не образуют ни ходульных, ни столбовидных корней. Таким образом, первоначальное представление о ходульных корнях как общем признаке всех панданусов оказалось превратным.
В пределах обширного рода панданус мы находим, наряду с наиболее обычными небольшой высоты (2-7 м) "деревцами" и "кустарниками" различных архитектурных типов, отдельные виды со стволом высотой до 30 м и больше. Самым крупным, очевидно, является новогвинейский панданус Джулъянетти (Р. jiulianettii), даже средние по высоте экземпляры достигают 30 м. Разветвленную верхушку ствола с кроной из 3-метровых листьев у этого пандануса поддерживают толстые и необычайно длинные, до 13-14 м, ходульные корни.
Совершенно уникальный габитус (рис. 252) характерен для небольшой группы мадагаскарских панданусов, около 12 представителей которой составляют особую секцию рода (секция акантостила - Acanthostyla). На ранних этапах своего развития эти панданусы формируются как это свойственно остальным представителям рода: 3-4-метровый неветвящийся ствол несет в верхней части крупные, длиной до 9-10 м, листья типично панданового строения. Позднее, в ходе моноподиального нарастания ствола, на нем образуются тонкие боковые ветви, с каждым годом все более короткие и со все более мелкими листьями. Взрослое растение с прямым стволом высотой до 20 м несет пирамидальную крону, напоминая издали некоторые хвойные. Панданусы такого строения и получили название "конифероидных". Такой тип развития и строения уникален не только в пределах семейства пандановых, но и среди однодольных в целом. С другой стороны, известны и бесстебельные, можно сказать травянистые панданусы - виды особой секции фузиформа (Fusiforma), распространенные в низкогорных лесах Малайи, Таиланда и Калимантана. Прямые листья этих видов длиной до 7-8 м пучком отходят от уровня почвы непосредственно от горизонтальных подземных побегов. Листья эти необычайно твердые; отсечь такой лист можно только несколькими энергичными ударами очень острым малайским ножом парангом. По краям листья усажены столь же твердыми и острыми колючками. Бесстебельные панданусы образуют труднопроходимые заросли за счет обильного образования вокруг материнского растения корневищных отпрысков.
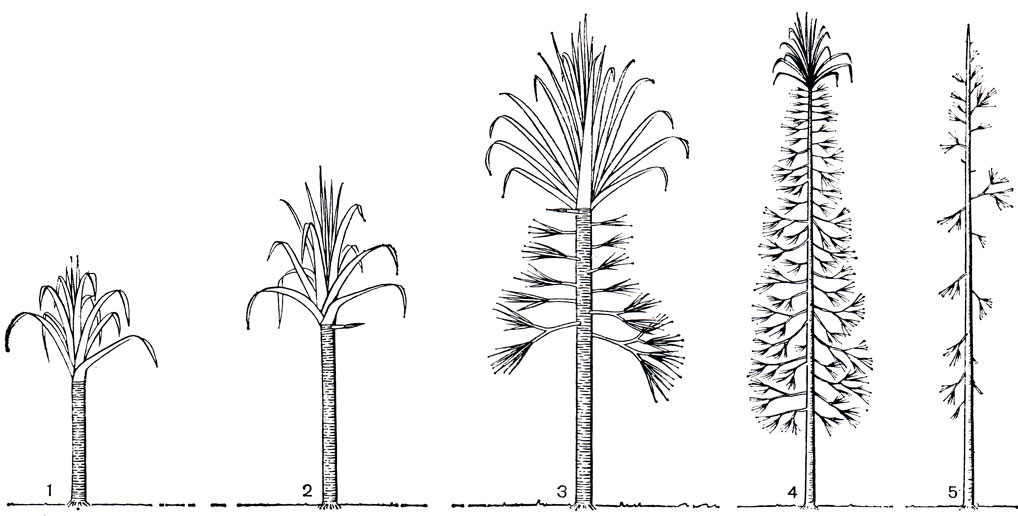
Рис. 252. Возрастные изменения (1-5) у 'конифероидных' панданусов из мадагаскарскои секции акантостила (по Б. Стоуну, 1970)
Попутно следует отметить и другой своеобразный способ вегетативного размножения, обнаруженный недавно у трех мадагаскарских панданусов (Ж. Л. Гийоме, 1972). Это вивипария, или живорождение. На стеблях этих растений, редко приносящих семена, развиваются выводковые почки, а сформировавшиеся из них миниатюрные растеньица-детки, опадая на землю, укореняются и образуют вокруг материнского растения разновозрастную колонию.
К низкорослым представителям рода относится и удивительный, также мадагаскарский, вид панданус карликовый (P. pygmaeus). Это как бы миниатюрная копия пандануса лабиринтного. На невысоком (до 60 см) стебле образуются расходящиеся во все стороны разветвленные, почти горизонтальные ветви, увенчанные на концах пучками тонких листьев. От этих ветвей вниз во множестве отходят придаточные воздушные корни. Ближайшие к стеблю, внедряясь в почву, образуют подобие столбовидных корней; отдаленные от него остаются воздушными.
Среди панданусов есть и эпифиты. Это представители особой секции рода, состоящей всего из 5-6 видов,- секции эпифитика (Epiphytica), распространенные в тропических дождевых лесах примерно в той же небольшой области, что и бесстебельные панданусы. Поселяясь вместе с эпифитными папоротниками и орхидными на деревьях (реже на скалах), эти панданусы развивают на коротком толстом стволике листья длиной до 3 м. Стволик весь лохматый от сохраняющихся оснований листьев, в пазухах которых накапливаются вода и перегной.
Два вида сараранги (Sararanga) не только по внешнему виду, но и по экологии сходны с типичными панданусами. Одиночными растениями или небольшими группами они встречаются по берегам рек, на осветленных участках в дождевых лесах, на болотах, а также в более сухих местах, в частности на пожарищах, что свойственно и многим горным панданусам. В горы сараранги поднимаются до 600 м. Сараранги относительно высокие пандановые (рис. 253): сараранга глубоковыемчатая (S. sinuosa) достигает 20 м, второй вид - сараранга филиппинская (S. philippinensis) - ниже, до 9 м. Это древовидные растения, в целом сходные с панданусами, но без ходульных корней. Зато основание ствола у них несколько расширено и от него, как у многих пальм, в почву отходят многочисленные тонкие при-даточные корни. Ствол гладкий, прямой и лишь в верхней части разветвляющийся на одном уровне на 2 или 3 ветви, каждая из которых может, в свою очередь, вильчато разветвиться. Как и у других пандановых, это ветвление в действительности является ложновильчатым и обязано пробуждению пазушных почек, когда верхушечная почка исчерпает себя на образование соцветия или отомрет при повреждении. Каждая конечная ветвь увенчана пучком четырехрядно и спирально расположенных листьев, достигающих 3-метровой длины.
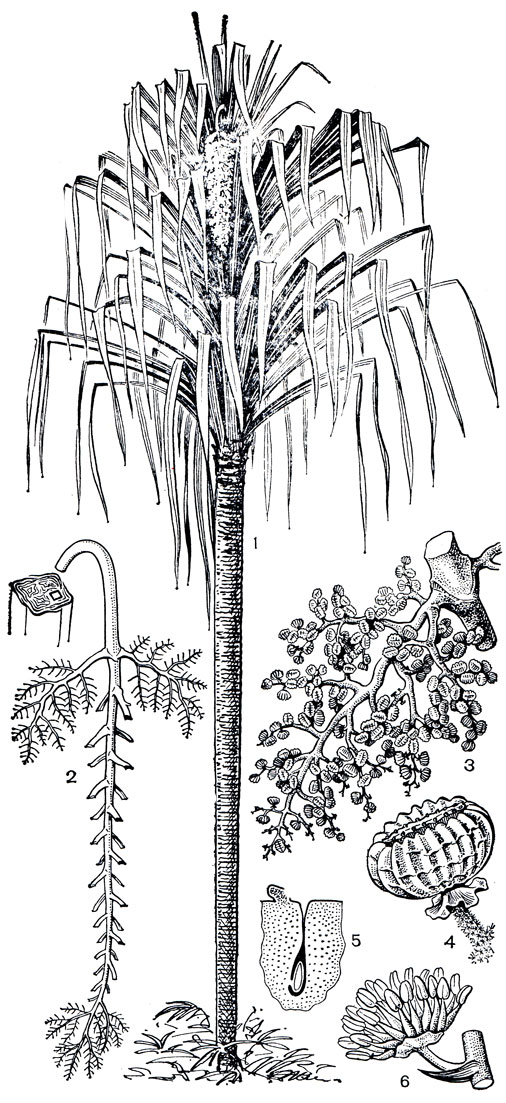
Рис. 253. Сараранга глубоковыемчатая (Sararanga sinuosa): 1 - общий вид; 2 - соцветие женского растения; 3 - плодоносящая ветвь этого соцветия; 4 - плод; 5 - гнездо завязи с семязачатком; 6 - мужской цветок
Виды фрейсинетии (Freycinetia) значительно однообразнее панданусов в своем приспособлении к условиям существования. Эти лианы в большинстве случаев являются обитателями тропических дождевых лесов, как низинных, так и горных, встречаясь от уровня моря до высоты 2500 м. Подобно плющам из семейства аралиевых, побеги фрейсинетий взбираются по стволам деревьев, иногда на большую высоту (некоторые виды до верхушек высоких деревьев), прикрепляясь к их коре своими придаточными корнями. У большинства видов листья скучены на концах тонких (диаметром до 1-2 см) побегов, но у некоторых они покрывают лиану почти от основания. И опять же, как плющи, фрейсинетии могут расти не только как корнелазающие лианы, но и как растения наскальные и даже почвопокровные; в последнем случае они заменяют местами травяной покров, что характерно, в частности, для единственного вида Новой Зеландии - обычно высоко взбирающейся лианы - фрейсинетии Бауэра (F. baneriana).
О возможности существования видов фрейсинетии, относящихся к другим жизненным формам, свидетельствуют некоторые интересные единичные находки. Так, один из 24 видов рода, обитающих на Соломоновых островах, обычно растущий как лиана, был встречен в виде приютившегося на стволе диллении (Dillenia) эпифита со свисающими побегами. В форме полуэпифита в лесах Новой Каледонии была обнаружена эндемичная здесь лиана фрейсинетия гидра (F. hydra).
Общей, бросающейся в глаза особенностью всех пандановых, как указывалось выше, является винтовое расположение листьев: трехрядное у видов пандануса и фрейсинетии и четырехрядное в роде сараранга; у последнего молодые ветви отчетливо четырехгранные. В виде исключения у одной из культурных форм пандануса полезного (Pandanus utilis) листья оказались расположенными в два ряда. Обычно спирали листьев на всем протяжении ствола однонаправленные, но у одного из южновьетнамских панданусов было обнаружено периодическое изменение в направлении листовых спиралей, приуроченное каждый раз к образованию нового соцветия.
Листья пандановых в большинстве случаев крупные, длиной 3-5 м, а у отдельных видов пандануса и до 9 м. Такие листья могут иметь ширину до 20 см, а в основании даже до 40 см. С другой стороны, некоторым видам пандануса и фрейсинетии, например фрейсинетии узколистной (F. angustifolia, табл. 60, 2), свойственны листья узкие, похожие на листья злаков. По форме листья цельные, большей частью продолговато-ланцетные или линейные, гораздо реже (у ряда фрейсинетий) эллиптические, как у новогвинейской фрейсинетии эллиптической (F. elliptica), листовая пластинка которой имеет длину 8-10 см и ширину 4-4,5 см. Типичный лист пандановых характеризуется весьма распространенным среди однодольных параллельным жилкованием без резко выраженной средней жилки, но лишь немногие из однодольных могут иметь столь большое число жилок, доходящее здесь у некоторых видов до сотни.

Таблица 60. Пандановые: 1 - фрейсинетия замечательная (Freycinetia insignis); 2 - фрейсинетия узколистная (F. angustifolia)
У всех видов семейства листья голые, но характерной их чертой являются колючки (шипики), расположенные по краям листа, а также снизу вдоль средней жилки (рис. 251). Безколючковые листья только у некоторых культивируемых панданусов. С другой стороны, в одной из гербарных коллекций был обнаружен образец пандануса ароматнейшего (Pandanus odoratissimus) с совершенно уникальной колючестью; здесь шипики не только покрывали всю нижнюю поверхность листа вдоль жилок (и располагались по его краям), но и были на верхней стороне листа.
У видов всех трех родов основание листа представляет собой более или менее выраженное стеблеохватывающее влагалище, в пазухе которого накапливается вода. Здесь у эпифитных панданусов и у многих наземных пандановых, зажатая между стволом и влагалищем, развивается и растет кверху сплющенная корневая система из разветвляющегося корня, отрастающего от стебля выше места прикрепления листа. Помимо всасывания накапливающейся в пазухах листьев воды с питательными веществами, окончания этих отрицательно геотропических корней, как и бородавковидных выростов (адвентивных корней) на стеблях и ходульных корнях многих панданусов, по мнению ряда авторов, выполняют функцию пневматофоров. Последние обеспечивают воздухом ткани сквозь мантию воды, стекающей по надземным частям растения в условиях постоянно высокой влажности под пологом дождевого леса. Так функционируют эти корни недолго; вскоре они отмирают и твердеют, превращаясь в торчащие вверх шипы.
Интересно отметить, что вода, накапливающаяся в пазухах листьев, очевидно, служит или местом выплода, или своеобразным водопоем для москитов, которыми кишат тенистые тропические леса. По наблюдениям Б. Стоуна (1967), на Марианских островах эти микроводоемы на стволах распространенного здесь пандануса сомнительного (P. dubius) привлекают к себе местный вид москита - аэдеса панданового (Aedes pandani), летающего и атакующего людей в дневное время.
Листья пандановых в большинстве случаев плотные и кожистые. У видов с крупными листьями их прочность обеспечивается сильным развитием механических тканей. Так, у сараранги глубоковыемчатой вдоль всего листа между жилками тянутся, как контрафорсы, подпирающие нижнюю и верхнюю гиподермы, мощные тяжи механической ткани, у многих других видов пандановых в прочный футляр из склеренхимы заключены сами проводящие пучки.
Из других признаков ксероморфной структуры, свойственной обеим сарарангам, многим видам пандануса и лишь немногим представителям рода фрейсинетия, следует назвать развитие толстой кутикулы, во многих случаях образующей выросты на поверхности листа,
появление на эпидерме простых или ветвистых сосочков, подчас совершенно перекрывающих устьица (.устьица у пандановых всегда с 4 побочными клетками), расположенные большей частью и с нижней и с верхней (в меньшем числе) стороны листа. Нередко устьица погруженные. Наконец, под верхней и под нижней эпидермой развита многослойная гиподерма.
Очевидно, именно ксероморфностью листьев сараранг и многих панданусов объясняется еще одна особенность их произрастания в природе, отмеченная рядом путешественников. Под сенью их крон почва обычно голая, лишенная растительности. Опадающие крупные плотные листья этих видов очень медленно перегнивают и, постепенно накапливаясь, образуют на почве слой толщиной до полуметра, создающий непреодолимую преграду для любых проростков.
Более нежные, менее плотные листья у пандановых тенистых тропических лесов - у большинства видов фрейсинетии и у некоторых панданусов.
Пандановые до недавних пор считались строго двудомными растениями. Лишь в самое последнее время был установлен ряд исключений. Б. Стоун (1968) обобщил сведения о наличии в однополых цветках ряда видов пандануса и фрейсинетии рудиментарных элементов другого пола, а в 1981 г. американский ботаник Пол Кокс подробно описал проявление истинной однодомности у фрейсинетии с островов Самоа и сослался на аналогичное открытие Б. Стоуна в отношении другого вида этого рода с Филиппинских островов.
Цветки пандановых собраны в соцветия. Наиболее примитивными из них являются соцветия видов сараранги. Это крупные, сильно разветвленные верхушечные повислые метелки с многочисленными цветками на коротких цветоножках и с прицветниками (рис. 253). Каких размеров могут достигать такие соцветия, показывает следующий пример. У высокого (до 20 м) женского экземпляра сараранги глубоковыемчатой, встреченного Б. Стоуном на одном из Соломоновых островов, метелка имела длину 2 м при цветоносе в 90 см. У мужских растений соцветия короче. Соцветия панданусов и фрейсинетий также однополые и в большинстве случаев верхушечные (реже пазушные), но, в отличие от рыхлых раскидистых метелок сараранги, это початки; сидячие и лишенные прицветников цветки здесь очень плотно расположены на более или менее утолщенной оси. У одних видов початки одиночные; у других соцветия сложные, с двумя, тремя и большим числом початков на разветвленной главной оси, с кистевидным или колосовидным их расположением. Сами початки при этом могут быть или вытянутыми, или шаровидными - головчатыми. У многих видов фрейсинетии кисть початков имеет сильно укороченную главную ось. При этом початки на более или менее длинных цветоносах кажутся отходящими от верхушек листоносных ветвей пучками по 2, 3, реже больше, т. е. образуют ложный зонтик. Наконец, в самом основании соцветий панданусов (рис. 254) и фрейсинетий (табл. 59, 60) всегда имеются скученные прицветники и среди них крупные кроющие листья, нередко ярко окрашенные.
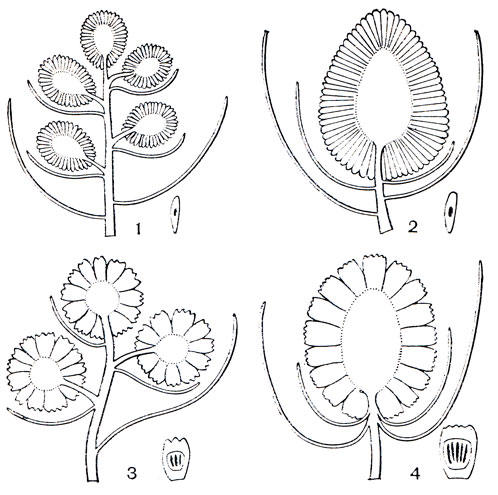
Рис. 254. Основные типы плодовых структур в роде панданус: 1 - кисть початков из свободных односемянных костянок; 2 - одиночный початок из свободных односемянных костянок; 3 - кисть початков, несущих фаланги - группы сросшихся костянок; 4 - одиночный початок, несущий фаланги (по Б. Стоуну, 1968)
Цветки в роде сараранга отличаются наличием короткой купулы, которую одни авторы рассматривают как рудиментарный околоцветник, другие - как преобразованные в эволюции и сросшиеся прицветнички. Андроцей мужских цветков сараранги представлен многочисленными тычинками. Гинецей в женских цветках паракарпный, из большого числа плодолистиков; у сараранги филиппинской их от 10 до 30, а у более мощной сараранги глубоковыемчатой - от 30 до 80. Каждому плодолистику соответствует сидячее рыльце, а в каждом гнезде развивается один семязачаток.
Цветки панданусов и фрейсинетий всегда голые и настолько сближены, что границу между отдельными цветками в ряде случаев установить крайне трудно. Более того, у многих видов обоих родов соседние цветки срастаются группами в обособленное образование, которое в литературе принято называть фалангами (греч. falanx - сустав, а также тесно сомкнутые линейные построения пехоты и конницы в Древней Греции и Македонии). Мужские цветки состоят из одних только тычинок, свободных или сидящих на особом выросте - колонке; на колонке кистевидно или в виде зонтика располагаются пыльники на укороченных нитях (рис. 251). Совершенно своеобразно расположение тычинок у пандануса крупноплодного (P. macrocarpus); верхняя часть колонки в его мужских цветках щитовидно расширена и несет на внутренней стороне щитка многочисленные пыльники, подобно тому как располагаются спорангии на щитках стробилов хвощей. Гинецей женских цветков или паракарпный, образованный несколькими сросшимися в нижней части плодолистиками, или мономерный (точнее, псевдомономерный). Рыльца на коротких столбиках или сидячие. Женские соцветия у панданусов, как правило, значительно более крупные, чем у фрейсинетий. Отличаются эти два рода и по числу формирующихся семязачатков; у видов пандануса на плаценте каждого плодолистика развивается, как правило, один семязачаток, у фрейсинетий - всегда от нескольких до многих.
Недавние исследования показали, что у некоторых пандановых, в частности у ряда панданусов, в строении гинецея проявляются необычайно примитивные черты: плодолистики в верхней части остаются несросшимися и в этом месте по их краям развивается диффузная рыльцевая поверхность с массой переплетающихся железистых волосков; при прорастании пыльцевого зерна пыльцевая трубка проходит сквозь этот слой волосков и оказывается в расположенной над семязачатком полости, заполненной в период цветения студенистой массой. Такое строение, как известно, характерно для ряда весьма примитивных двудольных. Наконец, как отмечено выше, у некоторых видов того и другого родов в цветках иногда наблюдаются слабо дифференцированные элементы другого пола. Анализируя данные о строении репродуктивных органов у пандановых, Б. Стоун (1968) приходит к предположению, что предковые формы пандановых должны были иметь метельчатые соцветия с обоеполыми цветками.
Приспособления к опылению у пандановых еще мало исследованы. Считается, что виды панданусов, образующие заросли на открытых местах (на побережьях, болотах), опыляются ветром, чему благоприятствуют большие количества продуцируемой пыльцы. Иначе обстоит дело у большинства представителей всех трех родов, обитающих в лесах. Вечнозеленые тропические леса, отмечает известный знаток тропической природы Э. Дж. Корнер (1964), кишат насекомыми, и ветроопыляемые растения здесь столь редки, что их трудно даже найти. Опылителями пандановых в этих условиях являются скорее всего насекомые. Яркий цвет кроющих листьев, неровная бородавчатая поверхность пыльцевых зерен, наконец, аромат, испускаемый соцветиями, особенно мужскими,- все это приспособления к насекомоопылению. Благоухающими являются соцветия пандануса кровельного и микронезийского пандануса душистого (P. fragrans); резким, неприятным для человека запахом отличаются соцветия пандануса вонючего (P. foetidus) и т. д. Некоторые виды фрейсинетии приспособлены к опылению птицами и летучими мышами. Тех и других привлекают как источник питания расположенные в основании соцветий этих видов прицветники, превращенные в мясистые сахаристые придатки.
Опыление летучими мышами из рода циноптерус (Cynopterus) было установлено еще в конце прошлого века в тропическом ботаническом саду Бьютензорг (остров Ява) у одной из самых мощных в роде лиан - фрейсинетии замечательной (Freycinetia insignis). Соцветия ее (табл. 60, 1) раскрываются вечером. При этом неяркие кроющие листья и наружные при-цветники отгибаются и открывают початки и расположенные под ними внутренние прицветники, превращенные в розовые питательные придатки. Раскрывающиеся соцветия начинают испускать характерный аромат с примесью запаха плесневеющих плодов. В течение ночи вблизи соцветий наблюдается активность летучих мышей; утром на соцветиях обнаруживаются следы когтей этих посетителей, а мясистые придатки оказываются в большей или меньшей степени отгрызенными.
Опыление птицами изучено у другого, также малезийского вида - фрейсинетии веревковидной (F. funicularis), соцветия которой раскрываются утром на дневные часы и лишены аромата. Птиц привлекают здесь необычайно яркие огненно-красные прицветники. В целом и мужские и женские соцветия с пучками из 2-3 (золотисто-желтых мужских или зеленых женских) початков в центре и с пламенным окружением из прицветников создают впечатление причудливых одиночных цветков. В Бьютензорге, где проводились наблюдения, постоянным посетителем соцветий этой фрейсинетии оказалась небольшая, размером с зяблика, птица из отряда воробьиных бюль-бюль золотобрюхий (Pycnonotus aurigaster). Птица садится на прочные наружные прицветники, стараясь добраться до мясистых внутренних при цветников - придатков, погружает свою головку в углубление между початками и их защитным обрамлением и при этом собирает на перьях лба, щек или шеи пыльцу, а при последующем посещении женских растений опыляет женские цветки. В отношении гавайской фрейсинетии древовидной (F. arborеа) высказано мнение об опылении не только птицами, но и крысами; во всяком случае, усы и мех крыс - постоянных посетителей этой фрейсинетии - часто бывают опушены ее пыльцой.
Плоды у панданусов и сараранги - костянки, у фрейсинетии - ягоды. У сараранги костянки свободные, мелкие (не крупнее 2 см), многокосточковые. При этом многочисленные дисковидные косточки образуют в мясистом мезокарпии изогнутые четковидные ряды. Экзокарпий костянок яркий: оранжевый у сараранги глубоковыемчатой, леденцово-красный у второго вида.
Из плотного женского соцветия панданусов формируется ананасовидное соплодие из прижатых друг к другу костянок. У видов со сросшимися завязями последние выделяются даже на поверхности соплодия в виде очерченных отдельностей (рис. 254); при распадении соплодия они целиком отделяются от его оси, а при прорастании заключенных в них семян образуют в ряде случаев целый пучок проростков. Утолщенная ось соплодия у одних видов деревенеет, у других остается мясистой. Сами костянки, особенно при головчатом характере соплодия, имеют обратнопирамидальную форму и у многих видов несут на расширенной наружной поверхности отвердевший столбик. В костянке панданусов только одна-единственная косточка, даже у видов, у которых женские цветки срастаются в фалангу.
Размеры и масса соплодий в роде панданус варьируют от маленьких (диаметром около 2 см) головок до огромных и тяжелых початков, как, например, у пандануса простого (Pandanus simplex). Одиночный початок этого растения достигает в длину 60 см при ширине до 20 см и массы в 25 кг. Значительно меньше и размеры и масса початков у фрейсинетии. Их гибкие и тонкие плодоносные побеги не смогли бы удержать крупные и тяжелые соплодия. Початки фрейсинетий состоят из плотно расположенных на утолщенной оси многосемянных ягод.
Семена пандановых мелкие, с маленьким Зародышем и обильным эндоспермом.
Плоды пандановых распространяются у одних представителей водой, у других - животными. Водами океана разносятся плоды панданусов, обитающих на океанических побережьях. Такие плоды с воздухоносной тканью (аэренхимой) в наружной части мезокарния, богатые и волокнистой тканью, и с семенами, защищенными прочным, почти водонепроницаемым эндокарпием, способны держаться на плаву месяцы, возможно и до года. С этим связана распространенность многих из таких видов не на одном, а на группе иногда значительно удаленных друг от друга островов. Распространением плодов океаническими течениями, по-видимому, можно объяснить и широкое распространение таких из названных уже прибрежных видов, как панданус кровельный и панданус многоглавый.
Интересно допущение английского ботаника Генри Ридли (1930) о возможности расселения водных (речных) панданусов из одной реки в другую рыбами. Так, соплодия упомянутого выше пандануса винтового, опадая в воду, вскоре распадаются в ней на отдельные плавучие тускло-зеленые костянки, столь мелкие (длиной около 6 мм), что их легко могут заглотнуть даже некрупные рыбы. В этом случае твердая косточка должна служить надежной защитой для заключенного в ней семени. Рыбы так любят эти плоды, что малайцы издавна используют их как приманку в рыболовных ловушках.
Лесные представители рода панданус, а также виды фрейсинетии и сараранги расселяются с помощью животных. С приспособлением к орнитохории, очевидно, связан более или менее яркий, контрастирующий с зеленью листвы цвет соплодий и плодов многих видов пандановых - красный, оранжевый, желтый и т. д. Иногда крупные соплодия даже разноцветны. Так, по наблюдениям Б. Стоуна на Марианских островах, зрелые соплодия пандануса сомнительного в нижней части сизовато-пурпурные, в средней - каштановые и на верхушке - оранжевые.
Птицы поедают ягоды видов фрейсинетии и при этом, несомненно, разносят их семена. Семена гавайской фрейсинетии древовидной, например, находили (иногда в изобилии) в желудках у ряда пернатых Гавайских островов - гавайской вороны (Corvus tropicus), местного гавайского медососа (Acrulocereus), попугайноклювой гавайской цветочницы (Psittacrostris). Ягодами новозеландской фрейсинетии Бауэра питаются новозеландский медосос (Prosthemadura novaezelandiae), попугай нестор (Nestor meridionalis) и филиппинский пастушок (Rallus philippensis). Опавшие на почву плоды некоторых панданусов поедают, а семена их при этом, по-видимому, частично разносят и четвероногие, в частности дикие свиньи, белки, крысы. В качестве возможных распространителей костянок и ягод пандановых называют также летучих мышей, сухопутных крабов и улиток.
Хозяйственное применение имеют многие виды пандануса, но в подавляющем большинстве случаев - это полезные растения лишь местного использования, основанного на жизненном опыте многих поколений аборигенов. При этом в той или иной степени находят применение практически все части этих растений: в одних случаях их мясистые и богатые протеинами, жирами и витаминами плоды и соплодия; в других - листья, цветочные почки или целые соцветия, семена и легкая "древесина" стеблей или их ходульные корни. Съедобные плоды используют в пищу, пищевым продуктом служат также листья некоторых панданусов (как овощ или ароматическая приправа), жесткие листья многих видов являются источником волокнистого материала для различных кустарных изделий (мешков, корзин, матов, циновок, веревок, шляп, поясов, парусов, сетей и т. д.) или целиком идут на покрытие кровель хижин.
Разнообразно используют части разных панданусов в народной медицине. Незрелые и зрелые плоды, мужские соцветия, листья, отвары из корней применяют местные лекари от разных болезней. В этой области народный опыт причудливо переплетается с древними обрядами и колдовством.
Не ограничиваясь сбором полезных частей панданусов непосредственно в местах их естественного произрастания, аборигены высаживают некоторые виды вблизи от мест проживания. По свидетельству Ван Стеениса, при расчистке участков тропического леса под посевы и посадки папуасы Новой Гвинеи не только бережно сохраняют заросли панданусов, остающиеся на полях или по их краям, но и высаживают здесь дополнительно их семена, так что иногда трудно определить, какие из панданусов представляют бывший здесь девственный лес, а какие высажены.
Большее экономическое значение имеет ряд видов пандануса, распространившихся в культуре в тропических областях за пределы своих ареалов и представленных в ряде случаев большим или меньшим числом культурных форм. Панданус кровельный - вид малезийско-тихо-океанского происхождения, но в настоящее время многие его культурные формы широко распространены в культуре от Аравийского полуострова на западе до далеких островов Океании на востоке в основном как источник волокнистого материала. Культивируется он и как пищевое растение, и на островах часто является единственной плодовой культурой. Растения этого вида полезны и в другом отношении: цветки, почки и листовые влагалища употребляются как овощ и как примесь к бетелю, сами листья - как приправа к рису, а также как лечебное средство при кожных заболеваниях и даже (в составных лекарствах с другими продуктами растительного происхождения) при проказе. Маслу из семян этого пандануса приписывают стимулирующее действие. Панданус полезный - вид мадагаскарского происхождения - возделывают как волокнистое растение уже более столетия, кроме тропических областей Африки и островов Индийского океана, также в Южной и Центральной Америке и на островах Карибского бассейна. Волокно листьев этого пандануса идет на изготовление тканей, плетений, бумаги, мешков для зерен кофе и для тростникового сахара. Из африканских видов наибольшее применение имеет панданус канделябровый (Pandanus candelabrum), листья которого используют для плетения корзин, кулей, для набивки матрацев, а "древесину" - на топливо и для изготовления плотов. Наконец, целый ряд видов пандануса, в меньшей степени фрейсинетии находят применение как
растения декоративные. Панданусы причудливого вида выращивают в странах с влажным тропическим климатом в садах, парках, в ряде случаев в виде живых изгородей по краям рисовых полей.
В областях умеренного климата наиболее часты в оранжереях панданусы полезный и кровельный, панданус тихоокеанский (P. pacificus), оригинальный панданус карликовый и др. На рисунке 251 изображен эффектный индийский вид панданус вильчатый (P. furcatus), выращенный в тропической оранжерее Ботанического института АН СССР в Ленинграде. Своеобразны пестролистные культурные формы, в частности панданус Вича (P. veitchii), привезенный в Европу в 1868 г. известным английским садоводом Джеймсом Вичем из Полинезии. Листья этого оригинального пандануса, являющегося, очевидно, одной из форм пандануса кровельного, по краям имеют белые или серебристо-белые полосы.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'