Порядок орхидные (Orchxdales) (В. Н. Гладкова)
Семейство орхидные (Orchidaceae)
Это крупнейшее среди однодольных семейство, насчитывающее около 750 родов и от 20 ООО до 25 000 видов (Р. Л. Дресслер, 1981), а по некоторым данным, гораздо больше - до 800 родов и 35 000 видов. Природа щедро одарила это семейство необычайной красотой и разнообразием цветков (см. табл. 28-41), изумляющих людей с древнейших времен и до наших дней. Поэты посвящали орхидеям стихи, художники изображали их на своих полотнах, ботаники давали им имена богинь и красавиц.

Таблица 28. Орхидные: 1 - башмачок крапчатый (Cypripedium guttatum), Дальний Восток СССР; 2 - башмачок крупноцветковый (С. macranthon), там же; 3 - башмачок настоящий (С. calceolus), Ленинградская область; 4 - башмачок Ятабе (С. jatabeanum), окрестности Петропавловска-Камчатского; 5 - башмачок крапчатый, Алтай, Телецкое озеро
Орхидные - космополиты. Они встречаются почти во всех пригодных для обитания растений областях земли, от Швеции и Аляски на севере до Огненной Земли и субантарктического острова Маккуори на юге. Но большинство их сосредоточено в тропических широтах, особенно в тропической Америке и Юго-Восточной Азии. Здесь, в областях с коротким сухим сезоном и высоким уровнем осадков они находят наиболее благоприятные условия для своего роста. По данным P. Л. Дресслера (1981), в тропической Америке встречается 306 родов и 8266 видов орхидных, в тропической Азии - 250 родов и 6800 видов. Своеобразие флоры орхидных на разных континентах - характерная черта их распространения. К Азии приурочено большинство видов такого крупного рода, как дендробиум (Dendrobium, 1400 видов), родов целогина (Coelogyne, 200 видов), фаленопсис (Phalaenopsis, 35 видов), ванда (Vanda, 60 видов) и др. Тропическая Америка является родиной таких широко известных в культуре орхидных, как каттлея (Cattleya, 60 видов), эпидендрум (Epidendrum, 500 видов), одонтоглоссум (Odontoglossum, 200 видов) и др. Большое количество эндемичных родов и видов встречается в Африке, на Мадагаскаре, в Австралии и Новой Гвинее. И только относительно немногие роды, как булъбофиллум (Bulbophyllum, свыше 1000 видов), имеют очень широкое распространение на разных континентах. В умеренном поясе флора орхидных гораздо беднее, чем в тропических широтах. По данным Ф. Г. Бригера (1971), на умеренные широты северного полушария приходится только 75 родов (10% от общего количества) и 900 видов (4,5%). Еще меньше - 40 родов и 500 видов - встречаются в южном умеренном поясе. Во всей Европе насчитывается 120 видов орхидных, в Северной Америке к северу от Мексики - 170 видов, тогда как только на востоке Новой Гвинеи - свыше 1450 видов орхидных, на небольшой территории Малайи - 800 видов, а в Колумбии - 1300.
Растут орхидные повсюду: от склонов самых высоких гор до лесных просторов равнин, от болот и водоемов до сухих степей и пустынных оазисов. В Колумбии некоторые орхидные живут в нетающих снегах гор, в Австралии растут под землей. Большое разнообразие их видов наблюдается во влажных горных лесах, особенно в лесах пояса облаков и туманов. Большинство орхидных не поднимается выше 2000 м над уровнем моря, но некоторые встречаются у верхней границы леса и даже достигают высоты 5000 м над уровнем моря.
В умеренных областях орхидные - многолетние наземные травы с подземными корневищами или клубнями, обычно скромными и неброскими цветками. Порой они не сразу заметны на лесных опушках и полянках, в зарослях кустарников, лиственных и хвойных лесах, а иногда резко выделяются яркими свечками соцветий среди зелени равнинных и горных лугов и болот. Некоторые из видов орхидных умеренного и тропического пояса принадлежат к общим родам и сходны между собой по облику. Таковы герминиум (Herminium), лесняк, или липарис (Liparis), мякотница (Malaxis) и др. Но большинство тропических видов мало похожи на своих скромных собратьев из умеренной зоны прежде всего потому, что значительная часть их - эпифиты.
Стебли наземных и особенно эпифитных орхидей чаще всего характеризуются симподиальным ростом. Растение представляет собой систему многолетних побегов, плагиотропные (горизонтальные) части которых образуют корневище, покрытое обычно чешуевидными листьями, а ортотропные (вертикальные) части с нормальными и чешуевидными листьями возвышаются над субстратом. Верхние из чешуевидных листьев корневищ несут почки возобновления, вырастающие в новый побег, когда рост главного побега прекращается в результате формирования верхушечного соцветия или отмирания верхушечной почки.
Гораздо реже встречается у орхидных моноподиальный рост (упрощенная схема обоих типов роста изображена на рисунке 135). Некоторые виды с медленным моноподиальным ростом, например из рода фаленопсис (табл. 35, 2), имеют короткий стебель со сближенными листьями. У других же при неограниченном росте главной оси, характерном для моноподия, стебель может вырасти очень длинным. Такие растения неспособны сохранять вертикальное положение и переходят к лазающему образу жизни. Настоящие лазающие орхидеи характерны для тропических родов ванда (Vanda, табл. 38, 3), арахнис (Arachnis), онцидиум (Oncidium, табл. 37, 5, 41, 1) и др. Стебли этих растений вырастают вверх на значительную высоту, развивая большое число воздушных корней, укрепляющих растения на опоре с помощью корневых волосков, которые они образуют при соприкосновении с субстратом. Таким способом растения-эпифиты добираются до вершин деревьев и выносят на свет свои цветки. К корпелазающим орхидеям принадлежат и представители рода ваниль (Vanilla), в том числе и известная лиана - ваниль плосколистная (V. planifolia), выращиваемая в культуре (рис. 154). Междоузлия ванили сильно удлиняются раньше, чем развиваются листья. Развитые листья косо отстоят от стебля и как крючки закрепляют побег на дереве. Затем с противоположной листу стороны узла развивается в направлении к субстрату по одному быстрорастущему воздушному корню, которые все вместе, плотно цепляясь за опору, продвигают лиану в кроне дерева.
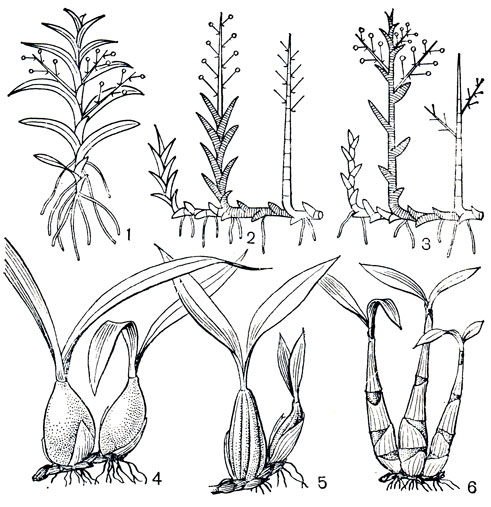
Рис. 135. Орхидные. Схема типов ветвления и псевдобульбы: 1 - моноподиальная форма роста; 2 - симподиальная форма роста с конечным соцветием; 3 - то же с боковым соцветием. Одночленные псевдобульбы: 4 - целогина гребенчатая (Coelogyne cristata); 5 - целогина повислая (С. flaccida). Многочленные псевдобульбы: 6, - дендробиум Кинга (Dendrobium kingianum)
Облик эпифитных орхидей, к которым принадлежит значительная часть тропических видов, очень разнообразен: от крошечных, едва различимых на ветвях деревьев растеньиц с невзрачными цветками, каковыми являются, например, виды азиатско-австралийского рода тениофиллум (Taeniophyllum) или центральноамериканская платистела юнгерманниевидная (Platystele jungermannioides), до пышных крупнолистных и крупноцветковых представителей американского рода онцидиум со свисающими соцветиями длиной несколько метров. Мимо одних из них путешественник проходит равнодушно или не заметив, другие заставляют его остановиться в изумлении. "Едва ли возможно вообразить что-либо более прекрасное,- писал о мадагаскарском эпифите ангрекуме полуторафутовом (Angraecum sesquipedale) немецкий ботаник В. Раух (1962), - чем дерево в девственном лесу, покрытое множеством этих величественных белых длинношпорцевых цветков, от которых к вечеру исходит сильный аромат ванили".
Эпифитные орхидеи можно встретить в районах как с ровным тропическим климатом без резких сезонных изменений, так и с муссонным климатом с выраженным сухим сезоном. Они поселяются на деревьях дождевого леса и саванны, в горных кустарниковых зарослях и в холодных лесах пояса облаков и туманов, при ярком солнечном освещении и в глубокой тени, одиночно и крупными колониями, повисая на ветвях деревьев над текучей водой. Некоторые из них вступают в союз с древесными муравьями и растут на их гнездах у вершин деревьев в "муравьиных садах", защищенные от всех посягательств (в том числе и ботаников) своими воинственными друзьями.
Семена этих растений содержат капли масла, а утолщенные стебли иногда бывают полыми с отверстиями у основания, через которые вползают муравьи.
Большинство эпифитных орхидей могут расти на разных деревьях, а кроме того, еще и на скалах, но у некоторых существует строгая приверженность к одному виду. Так, из 3000 видов деревьев на Филиппинах фаленопсис Шиллера (Phalaenopsis schillerana, табл. 35, 2) чаще всего выбирает дерево из семейства липовых - диплодискус метельчатый (Diplodiscns раniculata), а цимбидиелла Умбло (Cymbidiella humblotii) на Мадагаскаре всегда растет в заполненных гумусом щелях у основания листьев пальмы рафии муконосной (Raphia farinifera). Но сколь ни велик диапазон условий обитания эпифитных орхидных, они почти не выходят за пределы тропиков. Самой северной эпифитной орхидеей западного полушария считается эпидендрум комариный (Epidendrum conopsenm), встречающийся в буковых и магнолиевых лесах на юго-востоке Северной Америки, а в восточном полушарии, в Японии (о. Хонсю), на тех же широтах встречаются виды дендробиума. Еще севернее, на 39° с. ш., там же, в Японии, растет сакколабиум матсуран (Saccolabium matsuran).

Таблица 35. Орхидные: 1 - одонтоглоссум крупный (Odontoglossum grande), оранжереи Главного ботанического сада АН СССР; 2 - фаленопсис Шиллера (Phalaenopsis schillerana), там же
Все эпифиты развивают воздушные корни (рис. 136), одетые толстым слоем гигроскопической ткани из мертвых клеток, заполненных воздухом и способных впитывать не только дождевую воду, но и утреннюю росу и просто влагу из атмосферы, подобно промокательной бумаге. Иногда часть воздушных корней (у онцидиумов, например) образует подобие бороды, свисающей с деревьев, или густые сплетения, похожие на гнезда, в которых собирается гумус. Более мощные и толстые из них у лазающих орхидей - ванды или ангрекума - могут достигать земли и функционировать как нормальные корни. Для перенесения неблагоприятного периода эпифиты сбрасывают листья частично или полностью и, кроме того, запасают воду в сочных листьях и стеблях. Но главными водозапасающими органами служат у них утолщенные верхушечные части отрезков стебля, которые часто имеют форму луковиц и называются псевдобульбами (реже надземными или воздушными клубнями, туберидиями и пр.). В псевдобульбу превращается одно или несколько междоузлий, а иногда бывает утолщена вся ортотропная часть побега (одночленные и многочленные псевдобулъбы изображены на рисунке 135). По величине псевдобульбы варьируют от небольших, с булавочную головку (виды бульбофиллума), до шаровидных, с голову ребенка (у перистерии высокой - Peristeria elata). По форме они могут быть веретенообразными , округлыми, удлиненными плоскими, как лепешки, или цилиндрическими. Состоят они из мягких слизесодержащих тканей, а снаружи покрыты толстостенной эпидермой, уменьшающей испарение. У некоторых бульбофиллумов на верхушке псевдобульбы имеется полость с боковым отверстием, закрытым чешуевидным листом. В полости располагаются устьица, через которые осуществляется газообмен с атмосферой, и псевдобульба может функционировать как фотосинтезирующий орган. Псевдобульбы и скрытые в субстрате клубневидные утолщения стеблей развивают и многие наземные орхидные, растущие в муссонном климате с засушливым периодом, и даже некоторые орхидные умеренной зоны, как, например, калипсо (Calypso, рис. 137). У ряда других видов умеренной зоны и субтропиков (ятрышник - Orchis, офрис - Ophrys, любка - Platanthera, диурис - Diuris и др.) запасающую функцию выполняют аналогичные образования корневой природы - подземные корневые клубни. У австралийских диуриса, коризантеса (Corysanthes) эти клубни образуются на столонах, отходящих от верти-кального побега и растущих сначала горизонтально под землей, а затем, изгибаясь, выходящих на поверхность. В местах изгиба дифференцируется точка роста корня, который затем преобразуется в клубень. У ятрышника, любки, офриса под землей находятся 2 клубня (рис. 138). Один увядающий, мягкий, питательные вещества которого используются надземным побегом этого года и который к концу сезона темнеет и отмирает. Второй - молодой и твердый, с паренхимой, заполненной углеводами. Он погружается в почву с помощью столона и из него вырастает побег в следующем году. Согласно А. Кернеру фон Марилауну (1891), расположение клубней орхидных под землей характеризует в Средней Европе уровень промерзания почвы. У многих других орхидей запасающими органами могут также служить корневища, листья и утолщенные различным образом корни.
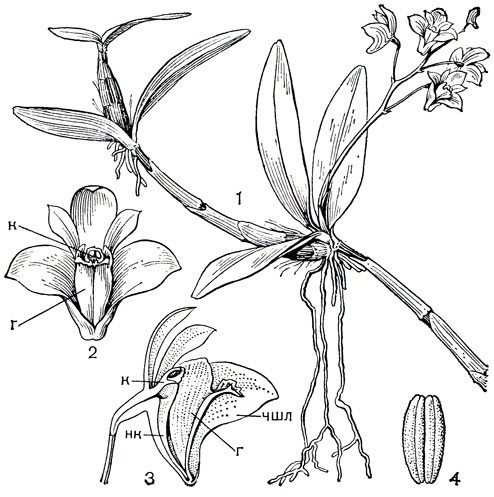
Рис. 136. Дендробиум Кинга (Dendrobium kingianum): 1 - общий вид; 2 - цветок; 3 - разрез цветка; 4 - поллинии (чшл - чашелистик, г - губа, к - колонка, нк - ножка колонки)
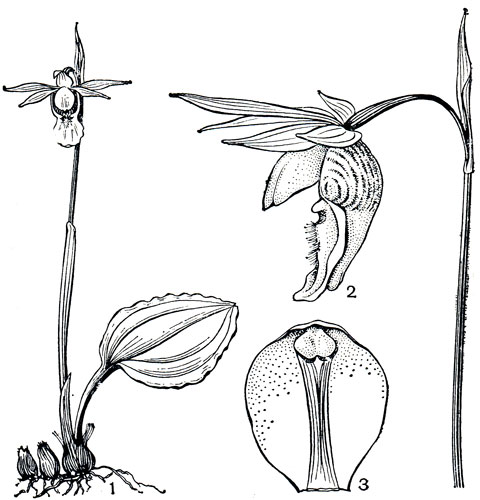
Рис. 137. Калипсо луковичная (Calypso bulbosa): 1 - общий вид; 2 - цветок; 3 - колонка
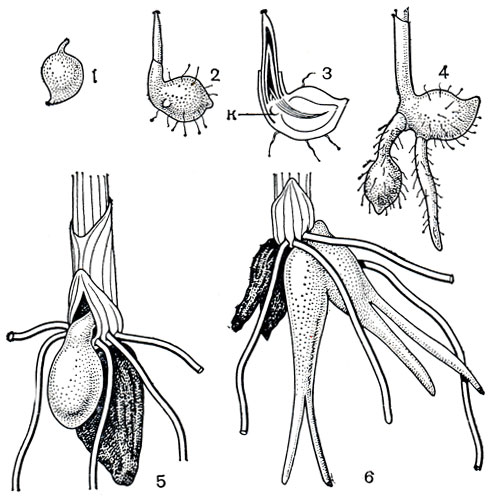
Рис. 138. Развитие подземных органов ятрышника шлемоносного (Orchis militaris) и корневые клубни пальчатокоренника широколистного (Dactylorhiza latifolia: 1 - зародышевый клубень; 2 - заложение первых листьев и первого корня; 3 - продольный разрез проростка, показывающий заложение первого корневого клубня (к); 4 - проросток следующей весной с корневым клубнем, погружающимся в почву; 5 - подземные органы взрослого растения: старый (темный) корневой клубень, несущий побег этого года, и молодой, из почки которого разовьется побег в следующем году; 6 - корневые клубни пальчатокоренника широколистного
Большинство наземных и эпифитных орхидей являются автотрофными растениями с зелеными листьями, в которых осуществляется фотосинтез. У некоторых эпифитных орхидей фотосинтезируют также и зеленые стебли, цветки и нередко воздушные корни. У видов фаленопсиса, безлистного тениофиллума и многих других корни являются главными фотосинтезирующими органами. Они плоские, зеленые и достигают, стелясь по субстрату, значительной длины. У многих орхидных с суккулентными листьями устьица открыты только ночью, когда влажность воздуха более высокая и через открытые устьица не происходит большой потери воды. Углекислый газ из насыщенной им атмосферы поступает в ткани растения и запасается там в виде молочной кислоты. Днем он освобождается и используется в фотосинтезе, происходящем при закрытых устьицах. Но среди эпифитных и наземных орхидей существует немалое количество видов, полностью утративших способность к фотосинтезу и ставших микотрофными. Симбиоз с эндофитными грибами свойствен всем орхидным на ранней стадии развития (речь об этом пойдет ниже), но только некоторые из них полностью зависят от грибов в течение всей своей жизни. Такие утратившие зеленые листья орхидеи поселяются на гниющих органических остатках, которые они, однако, не в состоянии усвоить сами и из которых извлекают необходимые для них соединения с помощью грибов. Гифы грибов проникают в живые клетки корней и корневищ и отчасти перевариваются растением, которое усваивает содержащиеся в них углеводы.
Большинство сапрофитных (или, точнее, микотрофных) орхидей - не очень крупные растения, как, например, виды леканорхиса (Lecanorchis) - обычного сапрофита в Юго-Восточной Азии, с тонкими и довольно хрупкими стеблями. Но некоторые бывают длиной 1 м и более, а виды галеолы (Galeola), лазающей сапрофитной орхидеи, распространенной от Восточных Гималаев до Австралии, достигают в длину 40 м и, подобно ванили, поднимают высоко на деревья и крыши домов свои коричневые стебли с красноватыми чешуевидными листьями.
К числу микотрофных орхидей принадлежат 2 замечательных австралийских вида: ризантелла Гарднера (Rhizanthella gardneri) и криптантемис Слетера (Cryptanthemis slateri). Обе эти орхидеи лишены листьев и корней и живут под землей. Они были открыты случайно (при расчистке и распашке фермерами участков своих земель) - ризантелла в 1928 г., а криптантемис три года спустя. Криптантемис - некрупная орхидея с белыми цветками, располагающимися под землей на глубине 2 см, с тех пор больше никем не была обнаружена и, возможно, уже исчезла, а колония ризантеллы после большого перерыва вновь была найдена в 1979 г. в 30 милях от Перта и взята под охрану. Эта колония состоит из очень маленьких растеньиц, несущих на верхушке хрупких и сочных подземных стеблей головки из крошечных суккулентных красных цветков, заключенных в обертку из кремоватых прицветников. Внешне соцветия напоминают соцветия маргаритки или другого сложноцветного. От корневищ отходят под землей боковые ветви с недоразвитыми цветками. Ризантелла цветет под землей, но близко к поверхности почвы. По мере того как прицветники вытягиваются, они выталкивают вверх тонкий слой опавших листьев и земли, образуя заметный холмик на поверхности почвы, и в конце концов цветок с завязавшимся плодом выносится на воздух, где коробочки вызревают и рассеивают семена. Цветки ризантеллы издают сладковатый запах, привлекающий каких-то живущих в почве опылителей, по предположению П. Бернгарда (1980), жуков. Жуки, по-видимому, вползают через отверстия между прицветниками в соцветие, и, пытаясь найти выход, прикрепляют к себе и уносят поллинии.
Как и другие бесхлорофилльные орхидеи, эти подземные растения живут за счет деятельности грибов, но особенность здесь состоит в том, что союз этот составлен не двумя, а тремя компонентами. Гифы грибов видов аспергиллa (Aspergillus) глубоко проникают в стебель ризантеллы через волоски на нем, а другим своим концом они связаны с гниющими пнями растения из семейства миртовых мелалеуки крючковатой (Melaleuca uncinata). Из этих пней они всасывают углеводы и минеральные соли, все еще содержащиеся там, и доставляют их в клетки орхидеи. Такой же сложный союз был отмечен и у второй подземной орхидеи - криптантемиса: гриб, который инфицирует эту орхидею, связан не только с ней, но и с корнями еще одного растения - автотрофной орхидеи диподиума точечного, или орхидеи-гиацинта (Dipodium punctatum). Возможно, здесь наблюдается непрямой (посредством гриба) паразитизм, отмеченный также и у некоторых эпифитных орхидей.
Микотрофные орхидеи в наших широтах также не редкость. В тенистых хвойных и лиственных лесах умеренной Евразии нередко можно встретить цветочные стрелки желтовато- бурых с медовым запахом цветков гнездовки обыкновенной (Neottia nidusavis, рис. 139, табл. 30, 2). Подземная часть ее состоит из корневища, обросшего множеством переплетающихся толстых корней, образующих подобие птичьего гнезда. Корневище служит запасающим органом и отчасти, как и корни, заражается грибом, поставляющим растению углеводы и азот из перегноя. Для формирования корневища требуется около 10 лет, только после этого оно выбрасывает недолго живущую цветочную стрелку. Соцветию не всегда удается пробиться сквозь почву и подстилку, и цветки на такой искривленной, обвившейся вокруг себя стрелке иногда развиваются прямо в слое перегноя. И семена иногда прорастают прямо в невыходящей на поверхность коробочке. У ладъяна трехнадрезного (Corallorhiza trifida, табл. 30, 2) и хрупкого желтостебельного надбородника безлистного (Epipogium aphyllum), редких лесных и болотных микотрофных орхидей северной умеренной зоны, подземная часть представляет собой разветвленное корневище, у ладьяна коралловидное, а у надбородника еще с отходящими от него столонами, из конечных почек которых развиваются новые растения (рис. 139).
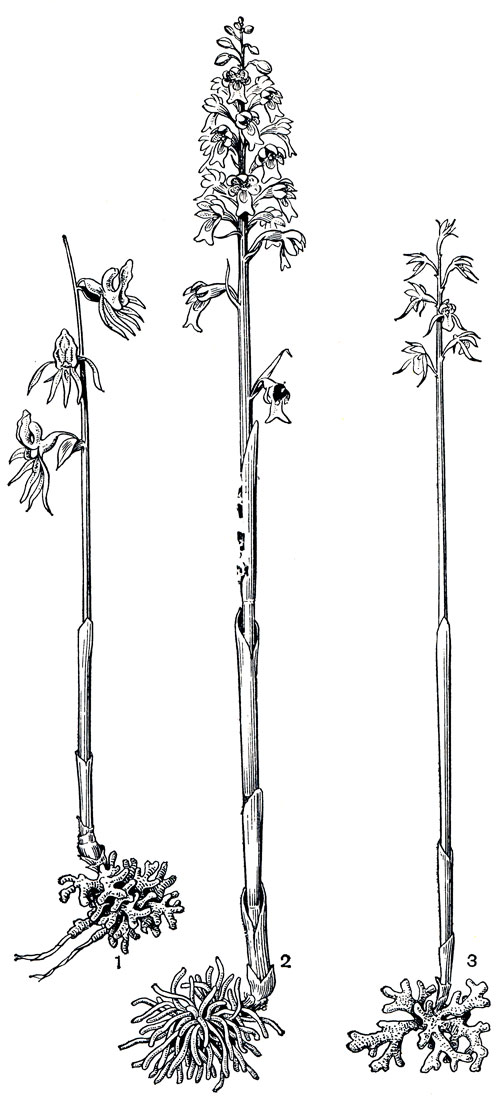
Рис. 139. Сапрофитные орхидные: 1 - надбородник безлистный (Epipogium aphyllum) с разветвленными корневищем и подземными столонами; 2 - гнездовка обыкновенная (Neottia nidusavis) с подземным корневищем, густо обросшим толстыми корнями; 3 - ладъян трехнадрезный (Corallorhiza trifida) с коралловидным корневищем

Таблица 30. Орхидные: 1 - гнездовка обыкновенная (Neottia nidus-afis), Алтай; 2 - ладьян трехнадрезный (Corallorhiza trifida), Ленинградская область; 3 - кокушник длиннорогий (Gymnadenia conopsea), там же; 4 - любка двулистная (Platanthera bifoiia), там же
Нередко грибами бывают заражены и растения с зелеными листьями. У гудайеры ползучей (Goodyera repens), например, часто вся подземная система бывает оплетена грибом. В чем состоит роль гриба в этом случае, остается недостаточно ясным. По предположению Э. Холтума (1953), гриб, возможно, разлагая гумус, снабжает растение минеральными солями, от недостатка которых особенно страдают эпифиты и без которых они не могут строить сложные органические соединения. И, возможно, "в обмен" растение делится с грибами частью углеводов, которые оно синтезирует.
Орхидеи не проявляют строгой специфичности по отношению к грибам, и один и тот же гриб может заражать несколько их видов. Но грибы неодинаково вирулентны по отношению к разным видам орхидных. Большинство грибов, поражающих их, относится к несовершенным грибам, главным образом, к роду ризоктония (Rhizoctonia). Некоторые микотрофные орхидные, как дидимоплексис (Didymoplexis) и гастродия (Gastrodia), бывают поражены высшими базидиомицетами, а японская гастродия высокая (G. elata) находится в плодотворном симбиозе с разрушительным паразитом деревьев опенком из рода клитоцибе (Clytocybe).

Таблица 31. Орхидные: 1 - офрис насекомоносный (Ophrys insectifera), Ленинградская область; 2 - офрис оводоносный (О. oestrifera), Крым; 3 - ремнелепестник козий (Himantoglossum caprinum), там же; 4 - траунштейнера шаровидная (Traunsteinera globosa), Ивано-Франковская область
На ранних стадиях развития все орхидные являются облигатно микотрофными. Их необычайно мелкие семена с недифференцированным зародышем не содержат никаких пищевых запасов, необходимых зародышу для роста, и неспособны прорастать без дополнительного источника питания. Почти все попытки вырастить орхидеи из семян заканчивались неудачей, пока французский ученый Н. Бернар (1899), изучавший микотрофную орхидею гнездовку, не обнаружил проросшие семена в ее подземных плодах и не предположил, что грибы играют какую-то роль в их прорастании. Позднее, в начале XX в. Н. Бернар и немецкий ученый Г. Бургефф подтвердили это экспериментально. Семя начинает разбухать и увеличиваться в размере, как только оно попадает на подходящий субстрат. Этому способствует легкость проникновения воды через рыхлую семенную оболочку. Возникает зародышевый клубень (рис. 138) с корневыми волосками на обращенной к субстрату поверхности. На этом развитие останавливается, так как ассимиляционной деятельности зародыша недостаточно для его дальнейшей дифференциации. Нити гриба проникают в зародыш со стороны суспензора, и, как в случае с сапрофитами, обеспечивают растущий зародыш органическим питанием (рис. 140). Иногда гриб становится паразитом и убивает зародыш, но в большинстве случаев между ними устанавливается физиологическое равновесие и начинается дифференциация зародышевого клубенька с формированием с верхней стороны конуса нарастания побега и заложением первого адвентивного корня. С развитием листьев, адвентивных корней и пазушных побегов завершается формирование проростка, а до развития взрослого цветущего растения проходят годы, иногда более 10 лет.
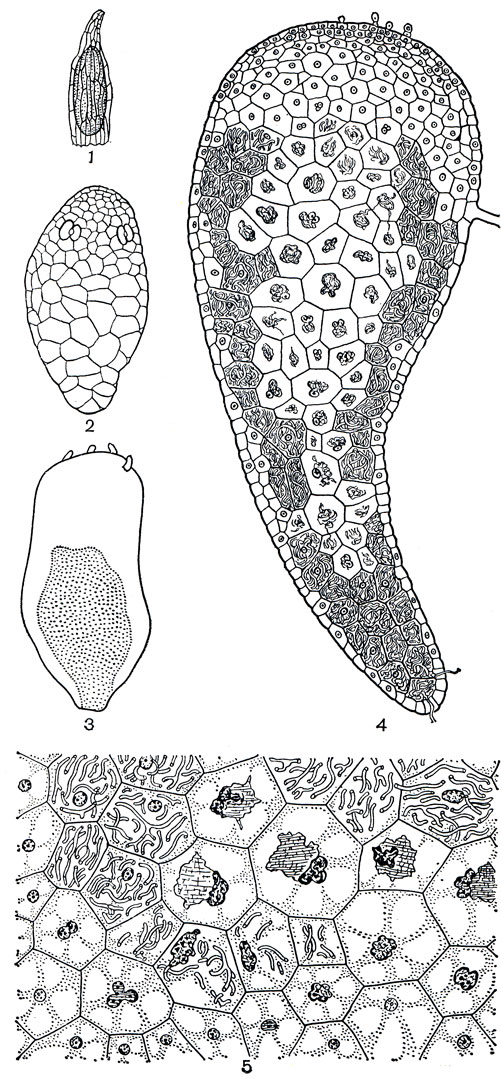
Рис. 140. Развитие орхидеи из рода фаленопсис (Phalaenopsis): 1 - семя; 2 - зародыш через 3 месяца после посева; 3 - схема разреза зародыша через 10 дней после заражения грибом ризоктонией плесневидной (Rhizoctonia mucoroides); 4 - продольный разрез зародыша через 50 дней после заражения грибом; видны здоровые клубочки нитей мицелия и тела вырождения, образовавшиеся в результате переваривания гриба; 5 - микроскопическая картина среза зараженного участка проростка; видны различные стадии переваривания клубков нитей мицелия
Гриб внедряется в периферическую зону зародыша или корней и корневищ (у взрослых орхидей), но не проникает за пределы этой зоны. Предполагают, что свободные от гриба зоны роста зародыша и клетки корневых клубней выделяют какие-то вещества с избирательным фунгицидным действием, не позволяющие нитям гриба проникать внутрь. Во внутренние участки коры корневища и корней они не проходят благодаря наличию слоев крупных крупноядерных, похожих на фагоциты, клеток, переваривающих грибные нити. В результате переваривания гриб теряет свою структуру, в клетках остается только бесформенная грибная масса, а освобождающиеся органические вещества используются растением. Не исключается возможность и прямого обмена веществами между двумя симбионтами через ненарушенные мембраны гриба.
По мнению Ж. Магру (1949), именно заражение грибом вызывает тот особый, характерный для корней и зародышей орхидных способ роста, приводящий к образованию клубней, и явление это есть не что иное, как скрытое грибковое заболевание, которое стало необходимым для их развития. По выражению У. Стерна (1960), орхидея состоит из двух растений и "носит прекрасные одежды наверху, но зависит от невидимого слуги внизу".

Таблица 32. Орхидные: 1 - комперия Компера (Comperia comperana), Крым; 2 - анакамптис пирамидальный (Anacamptis pyramidalis), Кавказ; 3 - ятрышник шлемоносный (Orchis militaris), Крым; 4 - ятрышник пурпурный (О. purpurea), там же
Семейство орхидных имеет много своеобразных черт, отличающих его от других семейств цветковых растений, как в строении репродуктивных, так и вегетативных органов. Как уже говорилось, корни орхидных придаточные, покрыты губчатой тканью, которая образуется в результате многократных клеточных делений эпидермиса, параллельных поверхности. Благодаря наличию воздуха в клетках, этот защитный покров (веламен) имеет белый или серый цвет. В веламене иногда поселяются сине-зеленые водоросли, вероятно, фиксирующие азот. В корнях развита механическая ткань, они трудно поддаются разрыву и удерживаются на опоре, даже если веламен разрушен. Корни и наземных и эпифитных орхидей иногда утолщаются и видоизменяются благодаря развитию в них водозапасающей ткани или уплощаются в связи с принятием на себя функции фотосинтеза.

Таблица 33. Орхидные: 1 - пальчатокоренник римский (Dactylorhiza romana), Крым; 2 - пальчатокоренник мясо-красный (D. incarnata), Ленинградская область; 3 - пальчатокоренник пятнистый (D. maculata), Московская область; 4 - пальчатокоренник Траунштейнера (D. traunsteineri), Эстонская ССР, остров Сааремаа
Листья орхидных простые, тонкие, или мясистые, влагалищные, или стеблеобъемлющие, очередные, двурядные, а иногда и супротивные, часто с выраженным сочленением пластинки и влагалища листа. Они очень различны по форме и величине: от довольно редко встречающихся крупных округлых (у австралийской нервилии - Nervilia), глубоколопастных (ацианта - Acianthe) или длинных ремневидных (у многих наземных орхидей с моноподиальным ростом) и даже цилиндрических (у брассаволы - Brassavola) до очень маленьких чешуевидных, едва различимых в лупу (у некоторых видов бульбофиллума, тениофиллума). У австралийской телимитры спиральной (Thelymitra spiralis) листья вьющиеся. В большинстве своем листья зеленые, но встречаются и пестролистные орхидеи (анектохилус - Anoectochilns, табл. 39, 2, виды фаленопсиса и др.). Листья располагаются либо на верхушке псевдобульбы по одному (у бульбофиллума) или по нескольку, либо отходят от боковых узлов стебля. На побеге обычно встречается два типа листьев: чешуевидные и нормальные. Иногда листья сближены на коротком стебле или даже образуют прикорневую розетку. Тонкие листья растение обычно сбрасывает на засушливый сезон, толстые и кожистые существуют по нескольку лет, у некоторых видов рода ванда (табл. 38, 3) - до 15. Как отголосок тропической природы орхидных можно рассматривать то, что в умеренной зоне листья некоторых орхидей уходят под снег зелеными (например, листья гудайеры ползучей или формирующиеся к осени единственный лист калипсо и розетки листьев видов ятрышника и офриса).

Таблица 39. Орхидные: 1 - лелия киноварно-красная (Laelia cinnabarina), оранжереи Главного ботанического сада АН СССР; 2 - анектохилус Даусона (Anoectochilus dawsonianus), там же; 3 - туния Маршалла (Thunia marschalliana), оранжереи Ботанического института АН СССР в Ленинграде
Соцветия орхидных развиваются либо после формирования листьев и псевдобульб, либо опережают их, либо появляются одновременно с ними. Располагаются они верхушечно на побеге или формируются в пазухах листьев главного побега, причем большинство высших орхидных имеют боковые соцветия. У моноподиальных орхидных соцветия всегда боковые. Основной тип соцветия, характерный для орхидных,- кисть, с цветками в пазухах прицветников, с цветоножками, часто незаметно переходящими в завязь. Кисть претерпевает различные изменения. В результате сильного укорочения ее оси возникает ложный зонтик (у цирропеталума украшенного - Cirrhopetalum оrnatissimum, табл. 37, 2, 41, 5). При укорочении цветоносов без укорочения оси возникают колосовидные соцветия ятрышников, гнездовки и др. При уплощении и утолщении оси развиваются головковидные соцветия некоторых тропических видов малаксиса (Malaxis) или других видов цирропеталума. При винтообразном скручивании оси появляются спиралевидные соцветия спирантеса (Spiranthes). Нередко соцветие метельчато ветвится и достигает больших размеров. Рекордную величину соцветия - до 5 м - имеет американский онцидиум волъвоксовый (Oncidium volvox). Иногда в результате редукции в соцветии развивается только одна боковая ветвь с единственным цветком или только один цветок в кисти - тогда говорят об "одиночных" цветках орхидей.
В основе строения цветка орхидных лежит трехчленный лилейный тип, претерпевший редукцию числа частей и значительные изменения в их структуре. Основной план строения цветков разных групп орхидей, демонстрирующий постепенные изменения от лилейного типа до однотычинкового цветка, представлен на рисунке 141. Цветки имеют 3 чашелистика, часто лепестковидные, обычно сходные между собой, но иногда 2 латеральных или 1 дорсальный отличаются от других по форме; изредка 2 или все 3 чашелистика срастаются между собой. Из 3 лепестков у подавляющего большинства орхидных средний сильно отличается от остальных, имеет специфическую функцию в опылении и называется губой (лабеллум). Губа, которая обычно крупнее других членов околоцветника, может быть цельной, лопастной, бахромчатой, рассеченной на множество тонких нитей, разделенной на 3 различающиеся по форме части (эпихилий, мезохилий, гипохилий). Поверхность ее может быть гладкой или с разнооб-разными выростами, папиллами, бородавками, гребнями. Кроме того, губа нередко несет нектарник в виде шпорца или продольной бороздки, ямки. Строение губы столь своеобразно, что некоторые ученые приписывают ей сложное происхождение в результате срастания разных элементов цветка. Ч. Дарвин (1862) считал ее возникшей путем страстания одного лепестка и двух лепестковидных тычинок наружного круга, а некоторые современные авторы, как Э. Нельсон (1967), полагают, что медианный лепесток выпал, а губа - результат срастания трех членов андроцея, принадлежащих разным кругам.
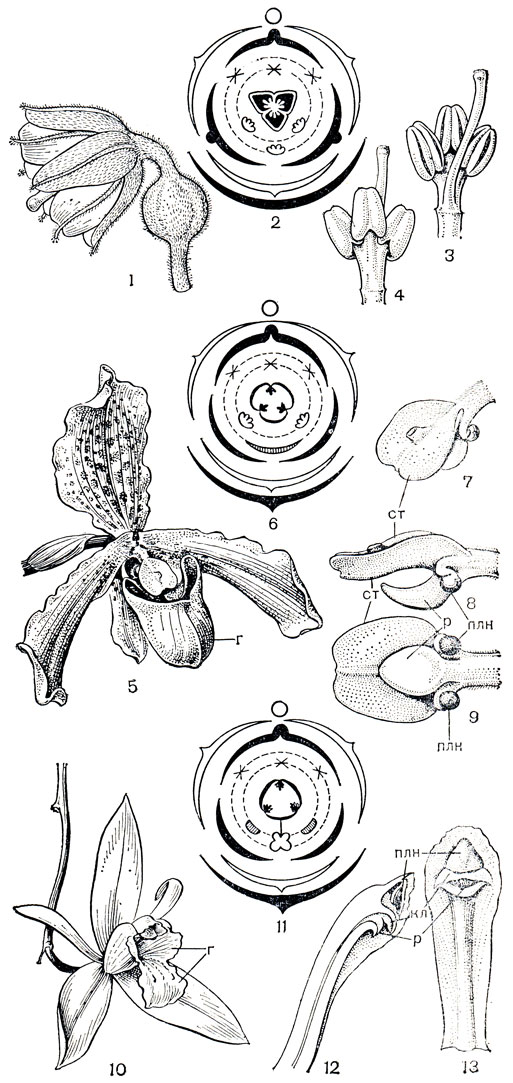
Рис. 141. Строение цветков в семействе орхидных. Подсемейство апостасиевые (Apostasioideae), неувидия Ины (Neuwiedia inae): 1 - цветок (чашелистики и лепестки с выраженным килем); 2 - диаграмма цветка; 3, 4 - колонка в двух положениях. Подсемейство циприпедиевые (Cypripedioideae), пафиопедилум замечательный (Paphiopedilum insigne): 5 - цветок с мешковидной губой; 6 - диаграмма цветка; 7, 8, 9 - колонка в разных положениях. Подсемейство однотычинковых орхидных (Orchidoideae), целогина повислая (Coelogyne flaccida): 10 - цветок; 11 - диаграмма цветка; 12 - разрез колонки; 13 - колонка; ст - стаминодий, р - рыльце, кл - клювик, плп - пыльник, г - губа
Из 6 двухкруговых тычинок, характерных для лилейных, у немногих примитивных орхидных еще сохраняются 3, из них 1 тычинка внешнего круга и 2 тычинки внутреннего круга (неувидия - Neuwiedia), или только 2 фертильные тычинки внутреннего круга (пафиопедилум - Paphiopedilnm), у всех же остальных орхидных функционирует только одна тычинка внешнего круга. У однотычинковых орхидей единственная тычинка объединена со столбиком и рыльцем в единую структуру, называемую колонкой или гиностемием. У более примитивных орхидных, как неувидия, апостасия (Apostasia), тычинки и столбики срастаются еще не полностью и имеют отчетливо выраженные свободные участки наверху. У циприпедиума и близких к нему родов также имеются свободные участки, но они более короткие, а над рыльцем нависает сильно развитый стаминодий, возникший из той тычинки внешнего круга, которая фертильна у однотычинковых орхидей.
Колонка - характернейший диагностический признак орхидных. Другим важнейшим признаком является строение их пыльцы. У подавляющего большинства однотычинковых орхидей тетрады пыльцевых зерен объединены в более или менее плотные массы - поллинии, которые бывают мучнистой, восковой или совершенно твердой роговой консистенции. Число поллиниев варьирует от 2 до 4, 6 и 8 и является систематическим признаком. Каждый поллиний содержит сотни тысяч пыльцевых зерен.
Завязь орхидных нижняя и у однотычинковых 1-гнездная с париетальной плацентацией, у большинства остальных - 3-гнездная с угловой плацецтацией. Особенностью завязи орхидных является ее скручивание (ресупинация) в процессе развития. В результате к концу бутонизации цветок разворачивается на 180° так, что губа, в бутоне обращенная коси соцветия, оказывается расположенной внизу и снаружи в удобном для насекомых положении (рис. 142). Разворот цветка губой вниз может осуществляться также в результате скручивания цветоноса или его поникания. Не всем орхидным нужна ресупинация: у тех из них, что имеют длинные, свешивающиеся вниз цветоносы, как у видов стангопеи (Stanhopea, табл. 38, 2, 2), или повисающие вниз соцветия, губа изначально находится в нужном положении. Но если привязать соцветие осью вверх, цветки в течение 24 часов поворачиваются вниз губой. Однако у немалого числа видов губа в течение всего времени цветения обращена вверх, ресупинация не происходит, и это связано со своеобразными способами их опыления. Случается также, что губа занимает верхнее положение в результате перекручивания завязи на 360°. У некоторых орхидных процесс скручивания захватывает и колонку. По мере созревания плода завязь постепенно раскручивается в обратную сторону.
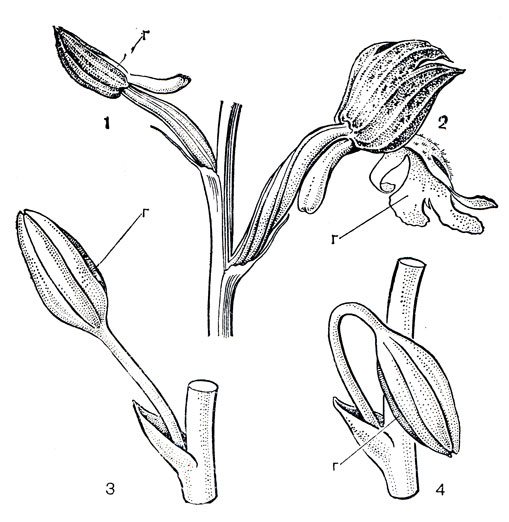
Рис. 142. Ресупинация цветков у орхидных: 1, 2 - вследствие перекручивания завязи (ятрышник пурпурный - Orchis purpurea); 3, 4 - вследствие поникания цветоноса (пафиопедилум - Paphiopedilum) (г - губа)
Плоды орхидных - коробочки, разнообразные по величине: от длинной мясистой стручковидной у ванили до маленьких сухих у большинства других родов. В большинстве случаев коробочки раскрываются тремя или шестью продольными щелями, створки при этом остаются соединенными наверху или расходятся. У некоторых видов коробочки раскрываются только двумя или даже одной щелью, а иногда семена освобождаются при сгнивании коробочки.
Для созревания семян необходимо от 2 до 18 месяцев. Необычайно мелкие и многочисленные семена с недифференцированным зародышем - еще одна характерная особенность орхидных. У большинства родов они снабжены сетчатой рыхлой заполненной воздухом оболочкой (рис. 143), но у представителей рода ваниль и близких родов оболочка плотная, жесткая, прилегающая к зародышу. Словно тончайшая пыль, семена орхидных уносятся ветром на большие расстояния. Большинство их гибнет, так как необходимая для прорастания встреча с грибом-симбионтом происходит не очень часто. Только у некоторых видов (блетилла гиацинтовая - Bletilla hyacintha, собралия крупноцветковая - Sobralia macrantha), имеющих дифференцированный зародыш, прорастание может происходить и без участия гриба.
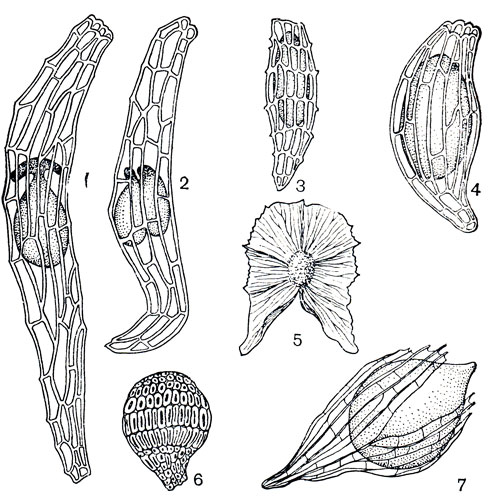
Рис. 143. Семена орхидных: 1, 2 - гибридная форма цимбидиума (Cymbidium) с одним и двумя зародышами; 3 - пафиопедилум замечательный (Paphiopedilum insigne); 4 - туния Бенсон (Thunia bensonieae); 5 - галеола высочайшая (Galeola altissima); 6 - ваниль плосколистная (Vanilla planifolia); 7 - ванда (Vanda sp.) в стадии начала прорастания
Классификация орхидных основана главным образом на строении их колонки. Новейшую систему орхидных разработал недавно американский ученый P. Л. Дресслер (1981) в своей книге "Орхидные. Естественная история и классификация". Семейство орхидных он разделяет на 6 подсемейств: апостасиевые (Apostasioideae), циприпедиевые (Cypripedioideae), собственно орхидные (Orchidoideae), спирантесовые (Spiranthoideae), эпидендровые (Epidendroideae) и вандовые (Vandoideae). Большинство современных авторов предпочитают разделение орхидных на 3 более четко очерченные подсемейства - апостасиевые, циприпедиевые (двутычинковые) и собственно орхидные (однотычинковые), с подразделением последнего подсемейства на более мелкие единицы.
Самое примитивное и самое маленькое подсемейство апостасиевых состоит всего из двух родов: апостасия (Apostasia, 7 видов) и неувидия (Neuwiedia, 9 видов). Это наземные лесные корневищные или бескорневищные травы, с очередными цельными влагалищными листьями и воздушными корнями. Цветки в верхушечных кистях, желтоватые или белые, только слегка зигоморфные, так как губа немного шире остальных лепестков или лепестки почти одинаковые. Чашелистики и лепестки с выраженным килем, переходящим в остроконечие. Колонка, прямая или искривленная в верхней части, образована неполным слиянием со столбиком оснований нитей трех (неувидия) или двух (апостасия) тычинок или двух тычинок и стаминодия (часть видов апостасии). Пыльца в отдельных зернах. Завязь у апостасии нересупинирована, а у неувидии ресупинация вызвана перекручиванием цветоноса. Как и у других орхидных, у апостасиевых развивается большое количество очень мелких семян в каждом из трех гнезд коробочки. Семена снабжены длинной или короткой ножкой и апикальным придатком, оболочка их ячеистая (рис. 144, 6, 9). Ареал апостасиевых охватывает Гималаи, Шри-Ланку, Бирму, Таиланд, Индокитай, Южный Китай, остров Хайнань, Южную Японию, Малайский архипелаг, архипелаг Луизиада и Северный Квинсленд (карта 8).
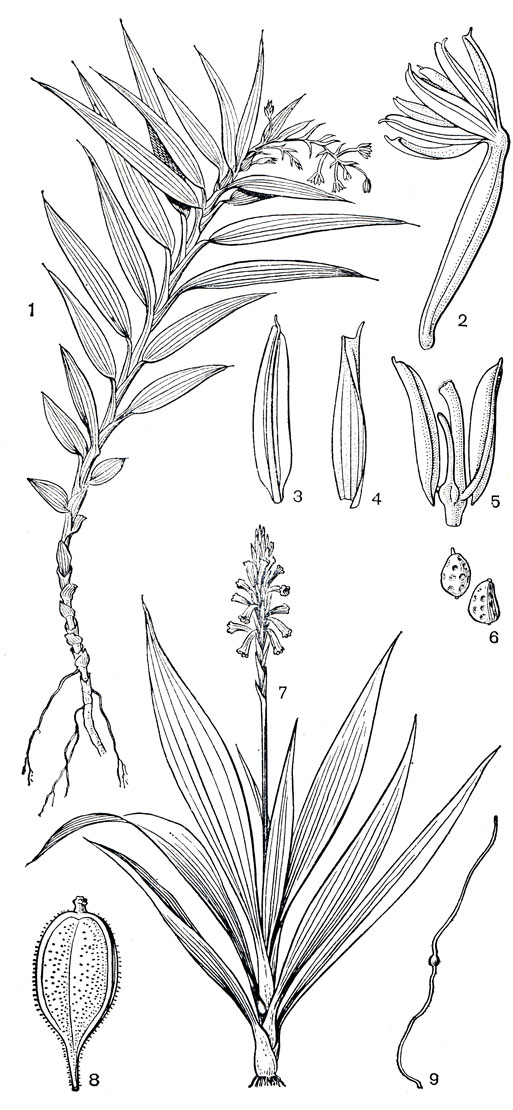
Рис. 144. Подсемейство адостасиевые. Апостасия душистая (Apostasia odorata): 1 - общий вид растения с воздушными корнями; 2 - цветок; 3 - лепесток; 4 - чашелистик; 5 - колонка с не полностью сросшимися двумя тычинками, стаминодием и столбиком; 6 - семена. Неувидия чемерицелистная (Neuwiedia veratrifolia): 7 - общий вид; 8 - плод; 9 - семя на длинной ношке и с волосовидным придатком сверху
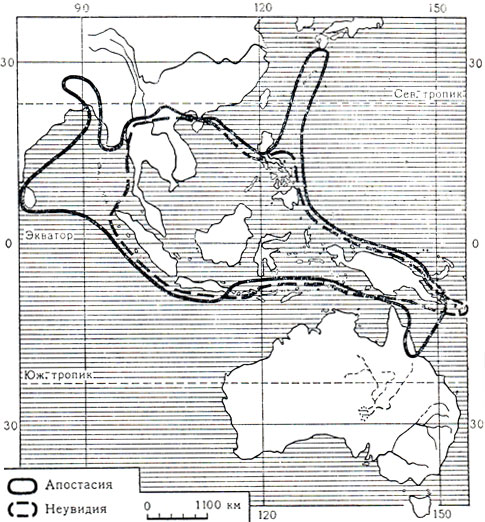
Карта 8. Ареал подсемейства апостасиевых
Соединительным звеном между настоящими орхидными и апостасиевыми является подсемейство циприпедиевых. Как и в предыдущем подсемействе, здесь развиты две боковые тычинки внутреннего круга и одна стахминоидальная тычинка внешнего круга, и в этом существенном плане два подсемейства обнаруживают сходство. Однако в отличие от апостасиевых циприпедиевые имеют зигоморфный цветок с мешковидной губой, скрученную у большинства родов завязь, 1-гнездную у циприпедиума и пафиопедилума и 3-гнездную у остальных родов, клейкую пыльцу в тетрадах или монадах, а у рода фрагмипедиум (Phragmipedinm) в поллиниях, как у настоящих орхидных.
Циприпедиевые включают 4 рода: селенипедиум (Selenipedium) с 4 видами и фрагмипедиум (табл. 40, 2, 2) с 11 видами в тропической Америке, пафиопедилум (Paphiopedilum, табл. 40, 5, 4) с 66 видами в тропической Азии и циприпедиум или башмачок (Cypripedium, табл. 28, табл. 40, 5) с 36 видами в умеренной и субтропической зонах северного полушария, особенно в горах Центральной и Восточной Азии. Это наземные или эпифитные травы с двурядно облиственными надземными побегами (циприпедиум, селенипедиум), являющимися конечными членами симподиального подземного корневища, или с укороченными побегами и розетками листьев на поверхности субстрата. К роду селенипедиум принадлежат крупнейшие наземные орхидеи Америки - стебли их могут достигать в высоту 5 м. Среди циприпедиумов, растущих в светлых лесах и на горных лугах, известны самые высокогорные виды, поднимающиеся в горах Восточной Азии до 5000 м над уровнем моря.

Таблица 40. Цветки орхидных подсемейства циприпедиевых: 1 - фрагмипедиум хвостатый (Phragmipedium caudatum) с лентовидными лепестками, достигающими в длину 75 см; 2 - фрагмипедиум Седена (P. sedenii), гибрид, полученный в культуре; 3 - пафиопедилум сиамский (Paphiopedilum siamense); 4 - пафиопедилум Файера (P. fairieanum); 5 - башмачок крупноцветковый (Cypripedium macranthon)

Таблица 41. Цветки орхидных подсемейства орхидных: 1 - онцидиум Крамера (Oncidium krameranum), южноамериканская орхидея-бабочка; 2 - каттлея Трианы (Cattleya trianae), одна из красивых зимнецветущих американских орхидей; 3 - эпидендрум желточно-желтый (Epidendrum viteliinum), американская орхидея горных лесов пояса облаков и туманов; 4 - неотропическая орхидея энциклия темно-пурпурная (Encyclia atropurpurea); 5 - цирропеталум украшенный (Cirrhopetalum ornatissimum), эпифит на деревьях в горных лесах Азии; 6 - бульбофиллум Лобба (Bulbophyllum lobbii) из Юго-Восточной Азии; 7 - масдевалия багряная (Masdevallia coccinea), характерное растение неотрописа, крупные наружные сегменты околоцветника срастаются, скрывая маленькие внутренние сегменты
Подсемейство собственно орхидных отличается от двух вышеописанных наличием одной фертильной тычинки, которая является медианной тычинкой внешнего круга. Боковые, тычинки внутреннего круга здесь либо вообще отсутствуют, либо превратились в различные стаминодии: ушковидные, чешуевидные, в виде желёзок, крыловидные или даже лепестковидные, расположенные по бокам или на вершине колонки и приросшие к ней или свободные до основания (у диуриса). Однородные по вышеуказанному признаку однотычинковые орхидные обнаруживают большое разнообразие в деталях строения колонки и поллиниев и на основании этих признаков разделяются на трибы. Большинство современных авторов выделяет 4 трибы (с подтрибами): неоттиевые (Neottieae), ятрышниковые (Orchideae), эпидендровые (Epidendreae) и вандовые (Vandeae). Некоторые современные орхидологи (Ф. Г. Бригер, 1958, 1971) рассматривают все 4 трибы в качестве подсемейств, а P. Л. Дресслер считает неоттиевые разнородной группой и часть их родов причисляет к ятрышниковым, часть выделяет в подсемейство спирантесовых, а некоторые роды относит к эпидендровым.
Рассмотрим более подробно удивительно разнообразное строение репродуктивных органов однотычинковых орхидных.
Пыльник может располагаться как на вершине колонки, прямо или наклонно, так и на ее дорсальной или вентральной стороне. Взаимное расположение пыльника и рыльца, характеризующее систематические группы, представлено на схеме (рис. 145), заимствованной из работы P. Л. Дресслера и К. Додеона (1960). Рыльце только у некоторых родов недифференцированное и полностью фертильное, а у большинства однотычинковых орхидных его медианная лопасть вся или частично стерильна и превращена в особый орган - клювик или ростеллум, отделяющий пыльник от обычно углубленной в колонку полости фертильного рыльца. Часть клювика, обычно краевая, ослизняется и при малейшем прикосновении выделяет липкое вещество, служащее для приклеивания поллиниев к опылителю. У некоторых орхидных (дендробиум - Dendrobium, табл. 34, 7, 2, целогина - Coelogyne, табл. 36,1) контакт свободно лежащих поллиниев с клювиком и осуществляется только с помощью насекомых. У других пол линии соединяются с клювиком при помощи особого хвостика, или каудикулы. Этот хвостик образован путем стерилизации части поллиния и состоит из клейкого эластичного вещества. Он может быть общим для нескольких поллиниев или отдельным для каждого. Хвостик у многих орхидных связан с клювиком как раз в том месте, где от клювика отделяется мягкий и липкий; участок ткани с перепонкой сверху, легко снимающийся при прикосновении и увлекающий за собой не только хвостик, но и сам поллиний. Этот липкий кусочек ткани клювика, разнообразный по форме, называют прилипальцем или висцидием, а иногда липкой желёзкой, липким диском или тельцем. У более высокоорганизованных орхидных прилипальце соединяется с поллиниями еще через промежуточную не липкую лентовидную полоску ткани, называемую ножкой, также отделяющуюся от клювика. Вся эта сложная структура, состоящая из трех частей (поллинии - каудикула - прилипальце) или четырех частей (поллинии - слабовыраженная каудикула - ножка - прилипальце), называется поллинарием. Варианты строения колонок и поллиниев подсемейства собственно орхидных изображены на рисунке 146.

Таблица 36. Орхидные: 1 - целогина гребенчатая (Coelogyne cristata), оранжереи Ботанического института АН СССР в Ленинграде; 2 - каланта Вича, садовая форма (Calanthe veitchii), оранжереи Ленинградского государственного университета; 3 - каланта одетая (С. vestita), оранжереи Главного ботанического сада АН СССР
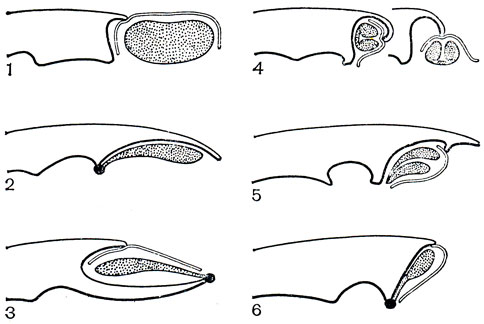
Рис. 145. Схематические продольные разрезы колонок однотычинковых орхидных, показывающие расположение пыльников, ноллиниев и рылец: 1 - примитивная орхидея с терминальным прямым пыльником и недифференцированным рыльцем (пыльцеголовник - Cephalanthera); 2 - орхидея с прямым пыльником, широко соединенным с колонкой, висцидий прикреплен к основанию поллиния (ятрышник - Orchis); 3 - орхидея с дорсальным пыльником, ростеллум вытянутый и прямой, висцидий прикреплен к верхушке поллиния (спирантес - Spiranthes); 4 - орхидея с крышечковидным пыльником (каттлея - Cattleya); 5 - орхидея с вентральным пыльником, перевершиниваемым колонкой (целогина - Coelogyne); 6 - орхидея с крышечковидным пыльником и ножкой, отделяющейся от ростеллума и прикрепляющей поллиний к висцидию (онцидиум - Oncidium)

Таблица 34. Орхидные: 1 - дендробиум непахучий (Dendrobium anosmum), Новая Гвинея, Ботанический сад в г. Лаэ; 2 - дендробиум благородный (D. nobile), оранжереи Ботанического института АН СССР в Ленинграде; 3 - дендрохилум пленчатый (Dendrochilum glumaceum), там же; 4 - одонтоглоссум красивый (Odontoglossum pulchellum), оранжереи Главного ботанического сада АН СССР
В трибе неоттиевых, к которой относятся роды пыльцеголовник (Cephalanthera, табл. 29, 2, 4), дремлик (Epipactis, табл. 29, 5), тайник (Listera), гудайера, анектохилус, спирантес, австралийские эндемики диурис, те ли митр а (Thelymitra), криптостилис (Cryptostylis), птеростилис (Pterostylis) и др., пыльник терминальный или дорсальный, прямостоячий или наклоненный. Пол линии мягкие, не имеют каудикулы, но могут быть снабжены прилипальцем. Контакт поллиниев с клювиком осуществляется через их удлиненную верхушку (пыльники акротонные). Пыльцевые гнезда сближены между собой, так как отсутствует выраженный связник. Типичное для трибы строение имеет колонка изображенного на рисунке 146, 1 тайника яйцевидного (Listera ovata). Здесь хорошо виден выступающий вперед клювик, образующий свод над рыльцевой ямкой и содержащий внутри с силой выталкивающуюся даже при легком прикосновении липкую жидкость, которая прикрепляет к опылителю свободно лежащие поллинии.

Таблица 29. Орхидные: 1 - пыльцеголовник красный (Cephalanthera rubra), Кавказ, окрестности озера Рица; 2 - гаммарбия болотная (Hammarbia paludosa), Ленинградская область; 3 - дремлик широколистный (Epipactis helleborine), Кавказ, окрестности озера Рица; 4 - пыльцеголовник крупноцветковый (Cephalanthera damasonium), Хмельницкая область
Другие представители трибы могут иметь упрощенное или более сложное строение. Примитивное строение колонки характерно для пыльцеголовника и близких к нему родов, у которых нет клювика, пыльник стоит прямо на верхушке колонки, а под ним располагается рыльцевая ямка. Рассыпающаяся пыльца беспрепятственно попадает на рыльце в бутоне, а в раскрывшихся цветках прикрепляется к опылителю, когда он испачкается рыльцевой жидкостью.
У рода спирантес (Spiranthes) пыльник дорсальный. У вышеперечисленных австралийских родов пыльник, отделенный от рыльца клювиком, также смещен на дорсальную сторону колонки, а сама колонка имеет выдающиеся крылья или окрашенные стаминодии и придатки. У этих родов иногда наблюдается смещение места соединения пыльника с ростеллумом от верхушки к основанию.
Акротонными пыльниками без связников,3 поллиниями без каудикул триба неоттиевых отличается от следующей трибы - ятрышниковых, колонки представителей которой - ятрышника, любки (Platanthera), кокушника (Gymnadenia) изображены на рисунках 146, 3 и 148, 5). В трибе ятрышниковых фертильная тычинка, сидящая широким основанием на колонке, несет прямостоячий пыльник, основания гнезд которого оттянуты в длинные придатки, соединяющиеся с двумя висцидиями (базитонный пыльник). Гнезда отделены друг от друга широким связником. Пыльники не опадающие, сохраняются на колонке. Поллиниев в этой трибе 2 или 4, они имеют мозаичное строение: состоят из отдельных пакетиков пыльцы. Пакетики соединены между собой эластичными нитями, сконцентрированными в виде тяжа по средней линии поллиния и выходящими в виде стерильного хвостика в отростки пыльцевых гнезд, где они касаются клювика. Мозаичное строение поллиниев имеет преимущество в том, что одним поллинием могут быть опылены несколько цветков, так как поллиний легко разрывается на части, которые по одной или по нескольку прилипают к рыльцам.
У ятрышника (рис. 146, 2) отростки пыльцевых мешкав сближены и концы их скрыты в общем карманчике - бурсикуле, образованном отгибающимся клапаном, в который превращена верхушка клювика; каждый поллиний имеет там собственное прилипальце. При прикосновении происходит разрыв клювика, клапан легко отгибается, но затем снова захлопывается, так что если удален только один поллиний, второй останется надежно защищенным от высыхания. У любки зеленоцветковой (Platanthera chlorantha, рис. 146, 3) базальные части пыльцевых мешков, заключающие в себе хвостики поллиниев, напротив, сильно расставлены в стороны и располагаются по бокам выпуклых лопастей фертильного рыльца. У кокушника длиннорогого (Gymnadenia conopsea, табл. 39, 5, рис. 148) клювик имеет вид вросшего в колонку удлиненного перепончатого колпачка, лежащего между основаниями пыльцевых гнезд. На нем вплотную друг к другу лежат 2 удлиненных (в виде полосок) прилипальца. Базальные части пыльцевых гнезд здесь поэтому сближены, а вследствие такого смещения фертильные лопасти рыльца отделены друг от друга и занимают боковое положение.
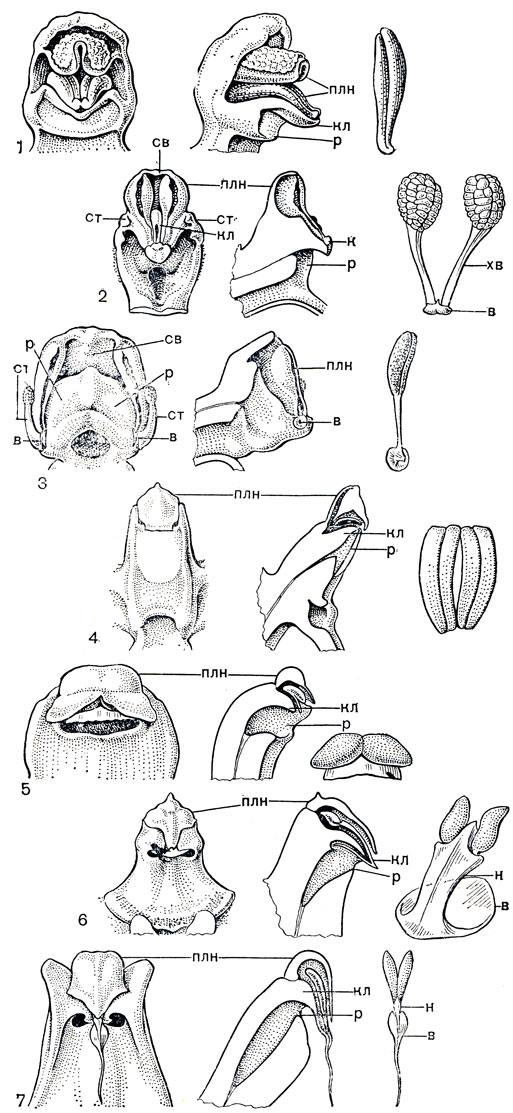
Рис. 146. Некоторые типы гиностемия (вид спереди и сбоку), поллинариев и поллиниев у однотычинковых орхидных: 1 - тайник яйцевидный (Listera ovata); 2 - ятрышник пурпурный (Orchis purpurea); 3 - любка зеленоцветковая (Platanthera chlorantha); 4 - дендробиум золотистый (Dendrobium chrysotoxum); 5 - цимбидиум повислый (Cymbidium pendulum); 6 - ванда трехцветная (Vanda tricolor); 7 - стангопея тигровая (Stanhopea tigrina): или - пыльник, кл - клювик, или ростеллум, р - рыльце, cm - стаминодий, к - карманчик, или бурсикула, св - связник, в - прилипальце, или висцидий; хв - хвостик, или каудикула, н - ножка у сложного поллинария
У других родов трибы тычинка может быть отклонена назад под углом 90° (диза - Disa и близкие роды) или даже на 180° и тогда свисает вниз (сатириум - Satyrium). Рыльце при этом располагается выше пыльника. Роды трибы ятрышниковых характеризуются наличием корневых клубней. Они широко распространены в Африке, в умеренных областях северного полушария и слабо представлены в тропиках. Наиболее крупным родом этой трибы является хабенария (Habenaria).
В двух следующих трибах - эпидендровых и вандовых, к которым принадлежит большинство тропических эпифитных орхидных, характеризующихся образованием псевдобульб, пыльник лежит на вершине колонки, как шапочка, или иногда занимает вентральное положение. У эпидендровых это достигается постепенным, по мере роста пыльника, изгибом тычиночной нити, а у вандовых - сильным укорочением нити пыльника (М. Хирмер, 1920, P. Л. Дресслер, 1981). У вандовых пыльник по сути прямой, но Хирмер допускал, что тычиночная нить у них изгибается на очень ранней стадии развития. Щитковидно расширенный связник с выступающей верхушкой опоясывает пыльцевые мешки сверху, препятствуя выпадению поллиниев. Но у некоторых родов пыльник может занимать и иное положение. Пыльники этих орхидных опадающие (у вандовых) либо качающиеся благодаря подвижному сочленению с тычиночной нитью. Рас-полагаются они обычно в особом имеющемся и у некоторых других орхидных углублении - клинандрии на вершине колонки. Углубление образовано основанием тычинки (сзади) и стаминодиями (с боков). Пыльцевые мешки акротонные или (нередко) выпячены в средней части (плевротонные). У эпидендровых поллиниев 4 либо, в результате полной или неполной, продольной или поперечной перешнуровки гнезд, 6 или 8. Они твердые, маслянистые или восковые и часто с хорошо выраженными каудикулами, которые либо соединяют каждую пару поллиниев, либо располагаются в виде тяжа параллельно поллинию, либо образуют общий для 4 или даже 8 поллиниев тяж. Иногда поллинии голые, как у дендробиума (рис. 136), бульбофиллума. Прилипальце есть у немногих родов, у большинства же оно редуцировано, и прикрепление к опылителю происходит с помощью каудикулы и липкого вещества клювика, стерильного обычно только по краю. Клювик может иметь разнообразную форму, от длинного, выступающего вперед подобно клюву птицы до подковообразного.
У большинства вандовых всего 2 очень твердых поллиния, уплощенных параллельно поверхности клинандрия. Каудикулы у них отсутствуют либо слабо выражены (видны только при искусственном оттягивании поллиния от прилипальца). Поллинии соединяются с прилипальцем при помощи особой ножки ростеллярного происхождения - полоски ткани, снимающейся с клювика. Иногда эта ножка расщепляется на 2, как у рода диподиум (Dipodium). У изображенного на рисунке 146, 4 дендробиума золотистого (Dendrobium chrysotoxnm, эпидендровые) пол линии лишены каудикулы, прилипальца приводятся в контакт с клювиком благодаря качающемуся пыльнику. У цимбидиума повислого (Cymbidinm pendulum, вандовые) развито прилипальце, а каудикула в рудиментарном состоянии (видна только при растягивании поллинария). У ванды трехцветной (Vanda tricolor) и стангопеи тигровой (Stanhopea tigrina) поллинарий состоит из трехвышеописанных частей, а воспринимающая поверхность рыльцевой полости сообщается с внешним пространством узкой щелью, в которую захватываются и иногда даже там замуровываются пол линии, принесенные насекомыми.
Со времен Ч. Дарвина до наших дней опыление орхидных не перестает привлекать внимание исследователей. И. И. Мечников в своей книге "Этюды о природе человека" (1903) называет механизм опыления орхидей одним из наиболее удивительных примеров среди гармонических явлений природы. Совершенные, иногда почти фантастические приспособления орхидей к специфическим опылителям поражают воображение. Словно изобретательный художник-конструктор забавлялся, выдумывая все новые и новые, иногда очень сложные механизмы, ярко и красочно исполненные и точно подогнанные к потребностям насекомых. Остроумные, часто обманные приспособления растений к опылению заставляют некоторых авторов говорить о "цветковой дипломатии", "сообразительности" и "безнравственности" орхидей. Опылению орхидных посвящена необозримая литература, основу которой заложил Ч. Дарвин (1862) замечательной книгой "Приспособления орхидных к оплодотворению насекомыми".
Опылителями орхидей подсемейства циприпедиевых являются двукрылые и пчелиные. Цветки одной из наиболее декоративных за пределами тропиков орхидей венерина башмачка (Cypripedium calceolus, рис. 147, табл. 28, 3) представляют собой как бы заранее расставленную и тонко продуманную ловушку, заставляющую насекомое произвести опыление. Крупная желтая губа, имеющая форму туфельки с завернутыми внутрь краями, сверху закрыта щитковидным стаминодием. Под ним лежит рыльце с обращенной вниз воспринимающей поверхностью, а по обе стороны от рыльца - по одной фертильной тычинке. Одиночные пчелы, особенно самки из рода андрена (Andraena), вползают и соскальзывают в губу цветка и, обследовав ее, начинают искать из нее выход. Ограниченное пространство не позволяет им взлететь, а завернутые края губы, ее гладкая внутренняя поверхность и заслон из стаминодия не позволяют не только выползти кратчайшим путем, но и рассмотреть прямой выход со дна губы. В задней стенке губы в направлении к гиностемию имеются, однако, "окна" - участки ткани, образованные клетками без пигментации, пропускающими свет. Принимая их за выходы, насекомое ползет в указанном ему направлении и отсюда уже видит настоящие выходы наружу: два узких отверстия слева и справа от колонки вплотную к пыльникам. Выбираясь из цветка, пчела сначала встречает на своем пути первую преграду - выпуклое рыльце. Грудь насекомого проходит над одним из выпуклых изгибов рыльца, которые счищают с нее принесенную пыльцу. Когда пчела пробирается все выше и ближе к свету, вторая помеха - пыльник суживает проход. А когда насекомое протискивается мимо пыльника, большая порция липкой пыльцы, как подушечка, приклеивается к груди пчелы (рис. 147).
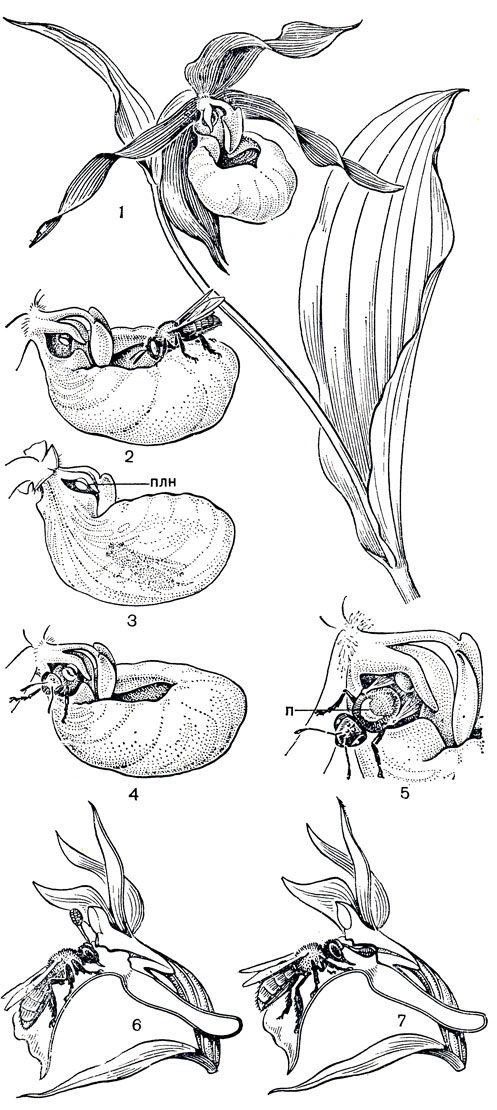
Рис. 147. Опыление орхидных. Башмачок настоящий (Cypripedium calceolus): 1 - верхушка побега; 2 - насекомое садится на край губы и соскальзывает внутрь ее; 3 - насекомое ползет в направлении 'окон'; 4 - голова насекомого показалась из узкого отверстия, расположенного вплотную к пыльнику; 5 - насекомое с комком липкой пыльцы (n) на спинке. Пальчатокоренник остистый (Dactylorhiza aristata): 6 - насекомое извлекает поллиний; 7 - насекомое вводит поллиний, наклонившийся во время полета, в рыльцевую ямку
Мелкие и ослабленные насекомые иногда так приклеиваются к пыльнику, что остаются там навсегда, закупоривая выход. А все, кому удается выбраться, уставшие и нередко мокрые после вынужденного купания в дождевой воде, которая скапливается на дне губы, на некоторое время даже теряют способность к координированному движению, но, отдохнув, снова летят на другие цветки. Что привлекает их в этих цветках? Дарвин считал, что насекомых может интересовать жидкость, выделяемая волосками, расположенными полосками в передней части губы. Е. Дауман (1969) показал, что, вопреки предположениям разных авторов, здесь не происходит отделения нектара или каких- либо иных питательных веществ, а маслянистая жидкость, содержащаяся в волосках, не служит питательным веществом для насекомых. Весь механизм опыления здесь построен на обмане: насекомых привлекает, по-видимому, запах и цвет, а яркий стаминодий служит для них указателем несуществующего нектара.
У подсемейства собственно орхидных бесчисленные вариации колонки определяют специфичность агентов опыления. На примере ятрышника мужского (Orchis mascula) -довольно обычной в Европе орхидеи-рассмотрим основную схему опыления у орхидных с базитонными пыльниками. Завязь этого вида ресупинирована, и губа занимает удобное для насекомых нижнее положение, имеет короткий шпорец. Два верхних лепестка и верхний чашелистик образуют колпачок, защищающий колонку от непогоды. В каждом пыльцевом мешке, открывающемся продольной щелью, располагается по одному поллинию с каудикулами и прилипальцами, скрытыми в карманчике на краю клювика (строение колонки ятрышника показано на рис. 146). Когда насекомое, севшее на губу, просовывает голову в полость, ведущую в шпорец, оно неизбежно натыкается на выступающий в этот проход клювик, нависающий над рыльцем. Конец клювика от прикосновения мгновенно расщепляется, и эластичный карманчик отгибается, обнажая 2 прилипальца, которые касаются головы насекомого, и быстро твердея, накрепко приклеиваются к нему. Если насекомое извлекает голову из цветка, оно вытаскивает за хвостик поллиний (один или оба), эластичный карманчик при этом захлопывается снова, защищая прилипальце оставшегося поллиния. Прикрепившиеся поллинарии сначала стоят вертикально на голове насекомого, как 2 рожка (рис. 147, 6 и рис. 148, 6).. Но в таком положении при посещении следующего цветка они попали бы снова на гнезда пыльника. И здесь вступает в действие замечательная способность поллинария к движению. Во время полета (примерно через 30 с) поллинарий благодаря несимметричному высыханию хвостика и сокращению прилипальца изгибается вперед примерно под углом 90°, и при посещении пчелой другого цветка поллиний точно попадает на его рыльце. Поверхность рыльца липкая, и весь поллиний или отдельные пакетики пыльцы остаются на нем, а пчела улетает с остатками хвостиков на голове, как свидетельством успешно выполненной работы.
Большинство ятрышников не имеет нектара в шпорце, и чем они привлекают насекомых, остается неизвестным. Ч. Дарвин полагал, что насекомые способны пробивать своим хоботком внутреннюю поверхность шпорца и высасывать обильную жидкость из его ткани, но более поздними наблюдениями это не было подтверждено. Ятрышник опыляют преимущественно коротко- хоботковые пчелиные, а также бабочки и мухи.
У других орхидных с базитонными пыльниками процесс опыления отличается только небольшими деталями. Например, у кокушника (рис. 148) удлиненные прилипальца образуют сводчатую кровлю над входом в нектарник - длинный шпорец, а лопасти рыльца занимают боковое положение с двух сторон от входа в нектарник. Когда бабочки просовывают хоботок в шпорец, сводчатые прилипальца приклеиваются к хоботку по бокам его и, как и у ятрышника, сначала торчат вертикально, а затем наклоняются вперед по обе стороны хоботка и, точно соответствуя боковому расположению рылец, попадают на них при последующем посещении цветка. Цветки кокушника привлекают насекомых запахом, усиливающимся к вечеру, и предлагают им нектар, обильно выделяемый эпидермальными клетками с папиллами в длинном изогнутом шпорце.
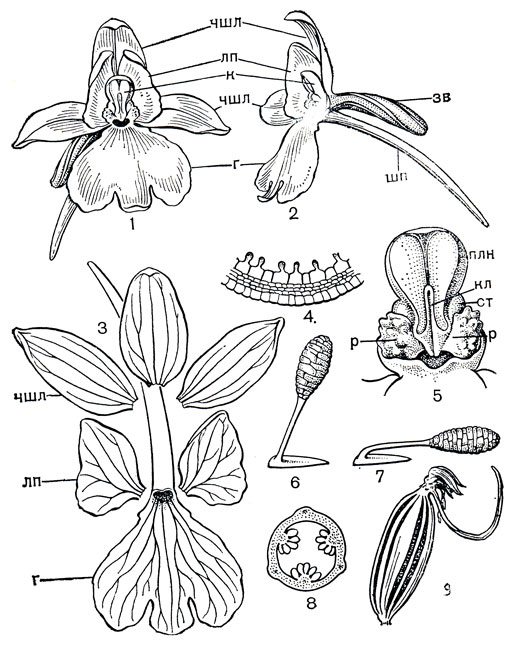
Рис. 148. Кокушник длиннорогий (Gymnadenia соnорsea): 1 - цветок; 2 - его вид сбоку (один лепесток и один чашелистик удалены); 3 - части цветка: 3 чашелистика, 2 боковых лепестка и медианный лепесток - губа с длинным шпорцем; 4 - поперечный разрез шпорца, видны эпидермальные клетки с папиллами, отделяющие нектар; 5 - колонка; 6 - поллинарий, состоящий из поллиния, его хвостика и длинного прилипальца; 7 - извлеченный из цветка поллиний изменяет свое положение, наклоняясь на 90° в соответствии с положением рыльца; 8 - поперечный разрез завязи; 9 - коробочка (чшл - чашелистик, лп - лепесток, г - губа, эв - завязь)
Опыление любки двулистной производят ночные бабочки, особенно бражники, летящие на сильный запах, который испускают к вечеру белые цветки. Отделенные друг от друга и расположенные по бокам клювика прилипальца (рис. 146) приклеиваются сбоку головки бабочки или к основанию хоботка либо даже на один из ее глаз, и во время полета их хвостик совершает движение вниз к центру так, чтобы поллиний мог попасть затем на поверхность рыльца, расположенную в середине колонки. А у близкого к ятрышнику рода анакамптис (Аnacamptis) прилипальце имеет седловидную форму, и во время полета бабочки края этого "седла" мгновенно закручиваются внутрь, охватывая хоботок с двух сторон, что приводит к расхождению поллиниев в стороны, затем они изгибаются, как у кокушника, параллельно хоботку, и такое их положение соответствует боковому положению лопастей рыльца. Разнообразные выросты, бороздки на губе ориентируют опылителя в нужном направлении. У анакамптиса, например, на губе имеются суживающие валики, направляющие движение хоботка насекомого к клювику. Иногда и сам клювик определяет действие насекомых. У хабенарии прямо вперед торчащий клювик заставляет пчел просовывать хоботок в нектарник со стороны так, чтобы хоботок коснулся рыльца. А у спирантеса (Spiranthes) клювик в молодых цветках закрывает вход в нектарник и одновременно доступ к рыльцу. Насекомое может только удалить поллинии, но не оставить их нарыльце, но у более старых цветков клювик сморщивается и отодвигается. Тогда освобождается проход к рыльцу. Этим обеспечивается перекрестное опыление.
Среди орхидных трибы ятрышниковых наиболее замечательный способ опыления наблюдается у офрисов. Цветки этих красивых и редких растений, растущих в Европе, Западной Азии и Северной Африке, часто напоминают насекомых (мух, пчел, шмелей) или пауков. От этого сходства и происходят их видовые названия - офрис насекомоносный (Ophrys insectifera), офрис пауковидный (О. sphegodes), офрис шмелецветковый (О. bombyliflora) и т. д.
У цветков офриса насекомоносного (рис. 149), произрастающего на карбонатной почве на лугах и лесных опушках от Средиземноморья и Малой Азии до Скандинавии, губа похожа на тело насекомого со сложенными крыльями, блеск которых имитируют серо-голубые полоски на губе, боковые лепестки имеют вид антенн, а расположенные в основании губы ложные нектарники напоминают глаза насекомого. Это загадочное сходство привлекало внимание многих ученых. Каким образом оно возникло и зачем оно растениям? Какие только догадки не высказывались по этому поводу! Не "сфотографировались" ли насекомые, кружащиеся над офрисами, на их цветках? Не существует ли это сходство только в восприятии человека, а насекомые его не замечают? Загадка разрешилась после того, как в 1916 г. французский ученый М. Пуйян высказал сенсационное предположение: цветки офриса воздействуют на половые инстинкты насекомых-самцов, имитируя их самок. Гипотеза Пуйяна была принята многими учеными. В России она пропагандировалась Б. М. Козо-Полянским (1938, 1939) в его блестящих работах о мимикрии у растений, в которых он также высказал предположение, что в привлечении самцов насекомых имеют значение какие-то неуловимые для нас запахи. Все это позднее было подтверждено экспериментально, а совсем недавно также наблюдениями в природе шведских ученых Б. Кулленберга и Г. Бергстрема (1976). Б. Кулленберг снял документальный фильм об опылении этих удивительных цветков, который был показан на XII Международном ботаническом конгрессе в Ленинграде в 1975 г.

Рис. 149. Опыление офриса насекомоносного (Ophrys insectifera): 1 - общий вид растения; 2 - цветок; 3, 4 - оса в процессе псевдокопуляции; 5 - голова осы с прикрепившимися к ней поллинариями
Цветки офрисов не "предлагают" насекомым нектара, пыльцы или иных питательных веществ. Но губа их выделяет летучие вещества из группы феромонов, главным образом производные жирных кислот и циклические сесквитерпены, набор которых специфичен для каждого вида или группы видов. Некоторые из этих соединений идентичны секрету желёз насекомых и воздействуют на них как ключевой раздражитель, стимулирующий цепь поведенческих реакций, характерных для спаривания. Внешний облик губы действует как визуальный стимул, а различные эпидермальные выросты на ней и определенным образом расположенные волоски воздействуют как тактильный раздражитель при попытках спаривания. Самцы жалящих перепончатокрылых из родов андрена (Andrena), эвцера (Еnсеrа), кампсосколия (Campsoscolia), антофора (Anthophora) и др. появляются после перезимовки несколько раньше самок и, побуждаемые половым инстинктом, летят на цветки офрисов. На разных их видах насекомые, в зависимости от структуры губы, ориентируются либо головой, либо брюшком к гиностемию и ведут себя так, как при копуляции с самками (рис. 149). В результате их импульсивных движений поллинии прилипают либо к их голове, либо к брюшку. Сексуальный инстинкт насекомых не удовлетворяется на одном цветке, и в результате нескольких посещений они производят перекрестное опыление.
Такая приспособленность офрисов к специфическим опылителям могла возникнуть только в результате длительной синхронной эволюции насекомых и растений. В этой цепи развития приспособлений наиболее специализирован офрис зеркальный (Ophrys speculum), опыляемый только осой из семейства сколиидовых кампсосколией реснитчатой (Campsoscolia ciliata), а наименее - офрис пчелоносный (О. apifera), который частично опыляется пчелами рода эвцера, а частично самоопыляется. При этом в строении цветка его имеются и совершенные приспособления к самоопылению. Они состоят в том, что гнезда пыльников самопроизвольно вскрываются, и поллинии под влиянием собственной тяжести выпадают из них, опускаются в направлении рыльцевой ямки, оставаясь прикрепленными к фиксированным хвостикам, имеющим как раз такую длину, которая необходима, чтобы при колебаниях воздуха они коснулись рыльца.
Процент завязывания семян у опыляющихся описанным способом офрисов чрезвычайно низок вследствие их очень узкой специализации по отношению к определенным видам насекомых. Особенно это заметно по контрасту с самоопыляющимся офрисом пчелоносным, у которого, как отмечено Ч. Дарвином, каждый цветок дает коробочку с семенами.
Привлечение опылителей запахами, воздействующими на их сексуальные инстинкты, характерно и для ряда орхидных из других триб, в частности для видов спикулеи (Spiculaea), дракеи (Drakaea), криптостилиса (Cryptostylis). Губа этих растений, с точки зрения человека, не всегда имеет сходство с самкой насекомого, тем не менее самцы пытаются спариваться с ней и уносят поллинии, которые, изменив во время полета свое положение на наклонное, при следующем "спаривании" точно попадают на рыльце.
Рассмотрим теперь опыление нескольких представителей трибы неоттиевых - листеры, или тайника, неоттии, или гнездовки, и эпипактиса, или дремлика. Принципиальное отличие их от представителей трибы орхидных состоит в том, что пол линии их лишены каудикул и контактируют с клювиком своей верхушкой (рис. 146, 1). У листеры в начале цветения клювик, как ширма нависающий над рыльцем, не имеет связи с поллиниями. Внутри клювик подразделен на ряд камер, содержащих липкую жидкость. Пыльник вскрывается еще в бутоне, и поллинии свободно лежат на клювике, защищенные вытянутой верхушкой тычиночной нити. Удлиненная раздвоенная на конце губа имеет по центру продольную бороздку, в которой скапливается нектар. Мелкие двукрылые и перепончатокрылые садятся на губу и, слизывая нектар, постепенно двигаются снизу вверх. Заканчивая трапезу, они поднимают головку вверх и касаются клювика. Из него тотчас выступает капля клейкой жидкости, которая приклеивается одновременно и к головке насекомого, и к поллиниям. Через несколько секунд эта жидкость затвердевает, прочно цементируя поллинии на голове насекомого. Поскольку поллинии из-за отсутствия хвостиков не способны здесь к движению, это компенсируется движением клювика и явлением протандрии. У более старых цветков, из которых извлечены поллинии, клювик поднимается кверху, освобождая широкий доступ к рыльцу. И множество разнообразных насекомых, для которых невзрачные цветки листеры с открытым нектаром очень привлекательны, успешно совершают перекрестное опыление.
Сходный механизм перекрестного опыления характерен для гнездовки, но пыльцевые массы здесь гораздо более рассыпчаты, и пыльца часто самопроизвольно падает на рыльце или переносится мелкими ползающими насекомыми в пределах одного цветка. Это вызывает самоопыление, которое является здесь, по-видимому, нормой. У неоттии бывают и клейстогамные цветки, развивающиеся под землей или в моховом покрове и образующие нормальные семена. Есть сообщения о клейстогамии и у других неодтиевых.
Опыление дремлика болотного (Epipactis раlustris, рис. 150) производят главным образом одиночные осы и, как выяснено Л. Нильсоном (1978), строение и размеры частей губы являются идеальной адаптацией к строению и размерам их тела. Губа дремлика разделена на 2 подвижно сочлененные половины: дистальную (эпихилий) и базальную (гипохилий). Эпихилий у своего основания утолщен, снабжен посредине глубокой бороздкой и краями обращен вверх, причем расстояние между краями очень мало (1,7 мм), так что там может разместиться только мелкое насекомое, у которого верхний сегмент брюшка меньше этой величины. Из жалящих перепончатокрылых со стебельчатым брюшком наиболее подходящие размеры имеют осы рода эвменес (Eumenes), которые и являются самыми эффективными опылителями этих растений (по наблюдениям Л. Нильсона в Швеции). Первый сегмент их брюшка имеет своеобразную колокольчатую форму, идеально соответствующую величине и форме эпихилия. Когда оса (рис. 150) садится на эпихилий, он, будучи подвижно сочленен с гипохилием, прогибается под ее тяжестью вниз и при кратковременной потере равновесия оса делает инстинктивное балансирующее движение своим брюшком, приводящее ее в контакт с клювиком и поллиниями. Нектар дремлика, отделяемый в чашевидной части губы, обладает наркотическим действием, что, возможно, зависит от сбраживающих процессов, вызываемых в нем дрожжевыми грибками, которые там обнаружены. Оса после посещения одного цветка еще способна сбросить с себя поллинии своими сильными ножками, но после посещения нескольких цветков она, отравленная нектаром, падает на землю и уже не может лететь и переползает от цветка к цветку, от соцветия к соцветию, нагруженная гроздьями поллиниев. А цветки дремлика долго не увядают и после опыления, продолжая выделять дурманящий нектар, неудержимо влекущий к ним насекомых во все время цветения. Кроме эвменеса, цветки дремлика опыляют и другие насекомые, другие одиночные осы, шмели, муравьи.
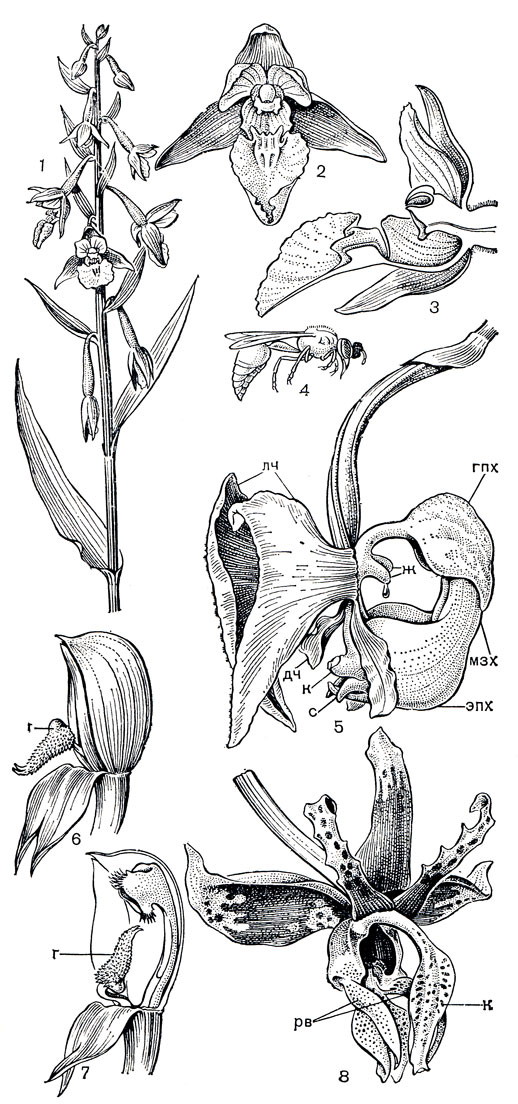
Рис. 150. Опыление орхидных. Дремлик болотный (Epipactis palustris): 1 - верхняя часть растения; 2 - цветок; 3 - разрез цветка, показывающий соответствие формы сложной губы форме тела опылителя (4). Кориантес пятнистый (Coryanthes maculata): 5 - цветок (гпх - гипохилий, мзх - мезохилий, эпх - эпихилий, ж - желёзки, выделяющие капли жидкости, лч - латеральные чашелистики, дч - дорсальный чашелистик, к - колонка, с - устройство для стока жидкости, через которое выползает насекомое). Птеростилис длиннолистный (Pterostylis longifolia): 6 - цветок с губой (г) в нормальном положении; 7 - цветок с поднявшейся губой (часть околоцветника удалена). Стангопея тигровая (Stanhopea tigrina): 8 - цветок (к - колонка, рв - роговидные выросты губы, направляющие насекомое по ее скользкой вогнутой средней части к верхушке колонки)
Характерное строение колонки эпидендровых и вандовых (рис. 146) с пыльником, расположенным на ней сверху, как шапочка, обусловливает особенности опыления этих растений. Поллинии приклеиваются к насекомым в момент, когда опылители покидают цветок и при этом приподнимают крышку пыльника. Пыльник, если он опадающий, приклеивается к насекомому вместе с поллиниями, а если он качающийся (сидит на тонкой нити с подвижным сочленением), то при прикосновении он отскакивает назад, выбрасывая поллинии (рис. 151). Сохранение крышки пыльника, снимающейся вместе с поллиниями, препятствует опылению того же цветка и, кроме того, действует как раздражающий фактор для насекомого, заставляющий его взлететь, чтобы избавиться от помехи. Сбросив с себя пыльник, насекомое с прикрепившимися поллиниями летит на следующий цветок, производя перекрестное опыление. У дейдробиума золотистоцветкового (Dendrobium chrysanthum), по наблюдению Ч. Дарвина, после посещения насекомого включается еще дополнительный механизм, гарантирующий самоопыление. Поллинии здесь, если не успели приклеиться к опылителю после скачкообразного движения пыльника, провисают на эластичной нити до самой губы венчика. Губа же под действием толчка от взлетающего насекомого подбрасывает поллиний так, что он попадает на рыльце собственного цветка, где и прикрепляется.
В опылении вандовых большое значение имеет характерная для их поллиниев ножка, которая выполняет здесь ту же роль, что и каудикула в трибе ятрышниковых. Ножка вследствие высыхания на открытом воздухе и сокращения способна к движению, может свертываться, наклоняться, складываться, благодаря чему поллинии занимают положение, необходимое для точного попадания на рыльце; длина ее в сложенном виде обычно соответствует глубине рыльцевой полости. От движения ножки происходит также сбрасывание с поллиниев крышечки пыльника. Ножки вандовых бывают очень разной формы. Каудикулы, как правило, скрыты в поллиниях, тем не менее они прочно прикрепляют их к ножке и благодаря их прочности поллинии могут быть удержаны и оторваны только очень липким рыльцем, что соответствует состоянию его зрелости. Тем самым также обеспечивается перекрестное опыление. Насекомых привлекает нектар, выделяемый в глубоком нектарнике, расположенном на внутренней стенке рыльцевой полости ниже рыльца или в шпорце губы. Опыление производят крупные насекомые, нередко пчелиные и бабочки, вынужденные глубоко просовывать в узкую рыльцевую щель свои хоботки, чтобы добыть нектар. Во многих случаях эта щель так узка, что способ введения в нее поллиниев остается не вполне ясным.
Успеху перекрестного опыления нередко способствует особая структура губы. У прекрасной орхидеи азиатских тропиков фаленопсиса (Phalaenopsis) губа снабжена у своего основания разной формы придатками. У фаленопсиса Шиллера (P. schillerana) этот придаток имеет форму кресла (рис. 151,2-8), на котором удобно устраивается муха, просовывающая свою голову в круглое отверстие рыльцевой полости, где имеется нектар. Прямо над этим отверстием нависает клювик, действительно имеющий здесь форму птичьего клюва, верхушка которого легко прилипает к голове мухи. Когда муха закончит свой обед, она вытягивает из пыльника оба поллиния, соединенные с прилипальцем клювика, и летит с ними на другой цветок. За краткое время перелета ножка поллиниев изгибается, по выражению А. Кернера фон Марилауна (1891), подобно лебединой шее,, и в другом цветке направленные вперед поллинии входят в рыльцевую полость и приклеиваются к рыльцевой поверхности. У некоторых других видов фаленопсиса придатки губы захлопываются как щипцы вокруг насекомого заставляя его извлечь поллинии вместе с пыльниками.
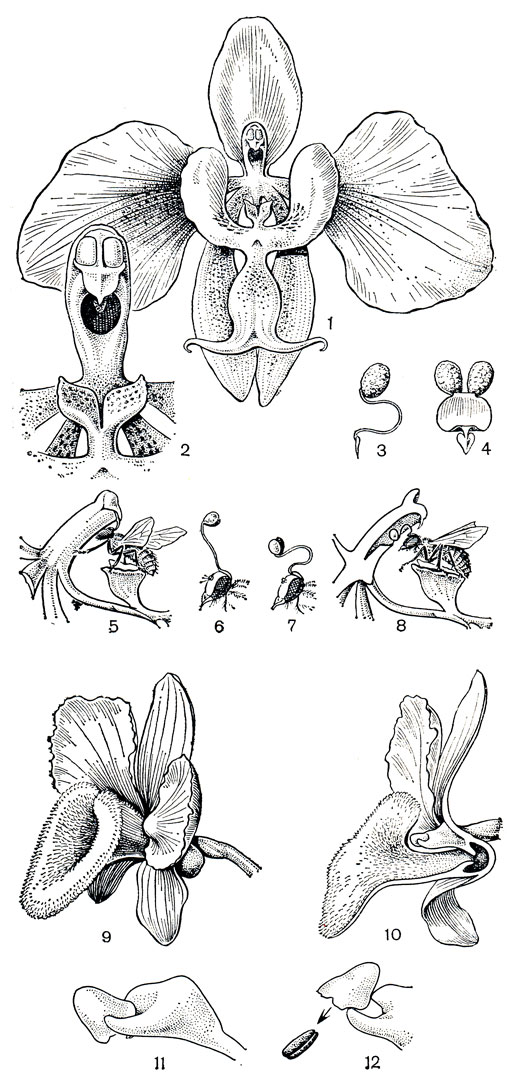
Рис. 151. Опыление орхидных. Фаленопсис Шиллера (Phalaenopsis schillerana): 1 - цветок, губа которого имеет придаток в виде 'кресла'; 2 - колонка; 3, 4 - поллинарий спереди и сбоку; 5 - муха, севшая на 'кресло', извлекает поллинии; 6 - голова мухи с приклеившимся поллинарием; 7 - ножка извлеченного поллинария изогнулась как лебединая шея; 8 - поллиний вводится в рыльцевую полость следующего цветка. Дендробиум бахромчатый (Dendrobium fimbriatum): 9 - цветок; 10 - разрез цветка; 11 - верхушка колонки; 12 - качающийся пыльник при прикосновении откидывается назад, и поллинии выбрасываются на насекомое
Но наиболее интересный механизм опыления среди вандовых имеет, пожалуй, представитель неотропической подтрибы катасетовых (Catasetinae) род катасетум (Gatasetum, рис. 152). Для видов этого рода характерны однополые цветки, причем мужские и женские резко различаются по внешнему облику. Иногда образуются еще и обоеполые цветки, причем все три варианта могут встречаться в одном соцветии. Собранные отдельно растения с мужскими и женскими цветками раньше принимали за особые роды, пока не выяснилось, что одно и то же растение может производить женские цветки в один сезон, мужские - в другой и смешанное соцветие - в третий. Факторы, вызывающие такое чередование, недостаточно изучены. По мнению К. Додеона (1962), изучавшего опыление этой группы орхидных, это обусловлено главным образом изменением экологических условий. В мужских цветках видов катасетума колонка имеет длинные роговидные выросты- антенны, отходящие от клювика по обеим сторонам пыльника к центру цветка или к полости губы. Раздражимые антенны при прикосновении к ним насекомого срабатывают как спусковой механизм, освобождая натянутую как тетива ножку поллинария, который при этом выстреливает вперед прилипальцем на грудь насекомого. Опыление катасетума крупноязычкового (С. macroglossum) представлено на схеме (рис. 153, из К. Додсона, 1962). Цветки его с губой, занимающей верхнее положение, начинают испускать сильный мускусный запах через 2-3 дня после расцветания. Этот запах неудержимо влечет самцов пчел из рода эвлема (Eulaema). Они охватывают губу, вползают в цветок в перевернутом положении и начинают передними ножками скрести внутреннюю поверхность губы. Выделяющийся при этом пахучий секрет воздействует на их хеморецепторы, расположенные на подушечках передних ножек. Во время этой работы пчела слегка задевает кончик антенны, расположенной как раз над углублением в губе, наиболее привлекательным местом, откуда исходит самый сильный запах. И спущенные стрелы - поллинарии вместе с крышечкой пыльника прикрепляются к ее тораксу. Клейкое вещество прилипалец вмиг затвердевает, поллинарии слегка отклоняются вниз и назад вдоль средней линии брюшка, и в таком положении пчела вносит их в женский, также нересупинированный цветок, где ведет себя так же, как в мужском. В женском цветке провисающие в направлении рыльцевой щели поллинарии захватываются липкой рыльцевой поверхностью при выходе пчелы из цветка. Женские цветки катасетума испускают сильный запах до тех пор, пока не будут опылены, иногда в течение месяца, но через несколько часов после опыления запах исчезает.
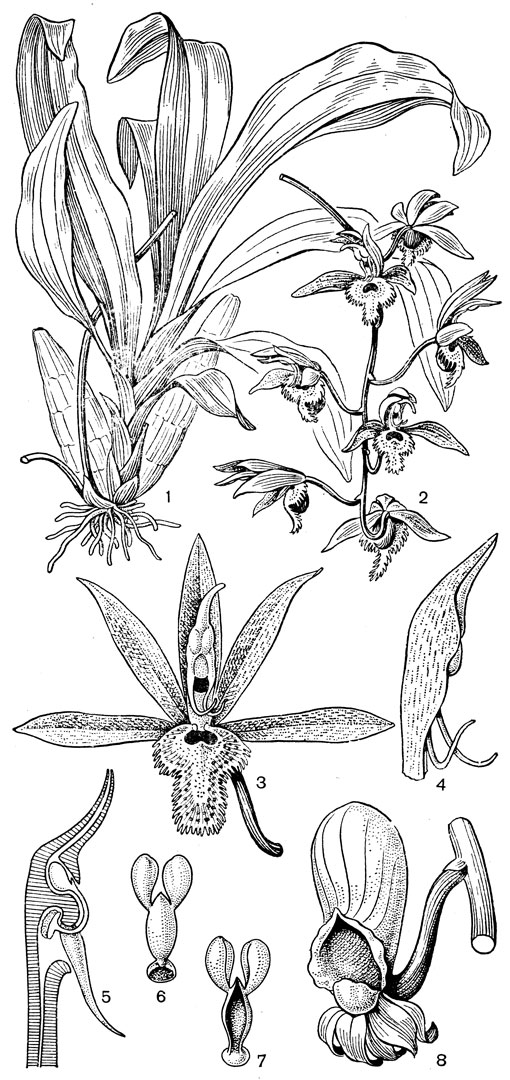
Рис. 152. Катасетум мешковидный (Catasetum saccatum): 1 - общий вид; 2 - соцветие из мужских ресупинированных цветков; 3 - мужской цветок; 4 - колонка с раздражимыми антеннами; 5 - продольный разрез колонки; 6, 7 - поллинарий в разных положениях; 8 - женский цветок
Другие виды подтрибы катасетовых, включающей еще 2 рода - мормодес (Mormodes) и цикнохес (Cycnoches), также способны выбрасывать поллинарий на опылителя, отличаясь деталями строения спускового механизма и способами прикрепления поллинариев. Опыление производят исключительно самцы пчел подсемейства эвглоссовых (Euglossinae), особенно из родов эвлема и эвглосса. Прежние сообщения о том, что пчелы обгрызают различные выросты на губе, не подтверждаются современными наблюдениями. Цветки не отделяют и нектара, так что единственным привлекающим фактором являются вещества, выделяемые осмофорами на губе растений. Эти пахучие вещества оказывают на насекомых наркотическое действие, пчелы совершенно теряют осторожность и обнаруживают острое пристрастие к "наркотику", по нескольку раз возвращаясь в один и тот же цветок. По некоторым предположениям (С. Фогель, 1966), самцы пчел, собирающиеся в скопления, используют этот цветочный запах, чтобы привлекать самок. Такие же "наркотические" вещества, сходным образом воздействующие на самцов пчел, выделяют и некоторые другие орхидеи из вандовых, в частности эпифитный род стангопея, цветок одного из видов которого - стангопеи тигровой (Stanhopea tigrina) - изображен на рисунке 150,S и на таблице 38,2. Очень крупные, пятнистые, сильно пахнущие цветки этой орхидеи парами свисают с ветвей деревьев. Занимающие самое нижнее положение в цветке губа и колонка располагаются напротив друг друга. Губа отчетливо разделена на 3 части (эпихилий, мезохилий, гипохилий), из них средняя часть имеет 2 крупных, направленных к колонке выроста, похожих на рога. "Голова быка" - называют это растение на его родине в Мексике. Привлекаемые запахом самцы эвглоссы скребут передними ножками углубленную поверхность верхней части губы, "пьянеют" и сползают вниз по гладкой, как каток, поверхности, "рога" же точно направляют их к колонке. При этом эвглосса задевает острие клювика со стороны прилипальца и уносит с собой поллинии.
Опишем еще несколько хитроумных приспособлений к опылению у орхидных. Кориантес (Coryanthes), замечательный род американских орхидей, часто поселяется на гнездах древесных муравьев. Его крупные, у некоторых видов достигающие в диаметре 13 см, цветки парами свисают вниз на длинных ножках. Замечателен этот род оригинальной формой своей губы, состоящей, как и у стангопеи, из трех частей (рис. 150,5). Гипохилий имеет форму ножки, отклоняющейся под прямым углом от оси цветка и от основания колонки и расширенной пузыревидно в месте перехода в цилиндрический мезохилий. От мезохилия отходит горизонтально и параллельно "ножке" (гипохилию) нижняя часть губы (эпихилий), которая и определяет все "события" опыления. Эпихилий имеет форму ванночки с очень гладкими стенками. Когда цветок полностью открыт, в "ванночку" стекает капельками бесцветная жидкость, выделяемая двумя желёзками, расположенными у основания колонки. Избыток жидкости вытекает через "сточную трубочку" - трехрогий отросток, действующий как носик у чайника. Колонка свисает вниз, причем ее изогнутая верхушка располагается как раз над этой трубочкой. Цветение продолжается 3-4 дня, чашелистики увядают раньше, делая губу еще более заметной насекомым. Привлекаемые сильным запахом, самцы эвглоссовых садятся на губу и, как и в предыдущих случаях, скребут поверхность гипохилия. "Пьянея", а также в результате борьбы друг с другом за место у. "вкусного стола" они сваливаются в приготовленную для них ванну. Приняв "холодную ванну", они начинают барахтаться в поисках выхода и выползают через "сточную трубочку", тесно примыкающую к колонке, пол линии при этом приклеиваются к насекомому. По наблюдениям П. Аллена (1950), для того чтобы выбраться из ловушки, пчеле требуется около 45 мин. После удаления поллиниев цветок перестает издавать запах, но на следующее утро запах появляется вновь и снова привлекает насекомых, чем достигается перекрестное опыление.
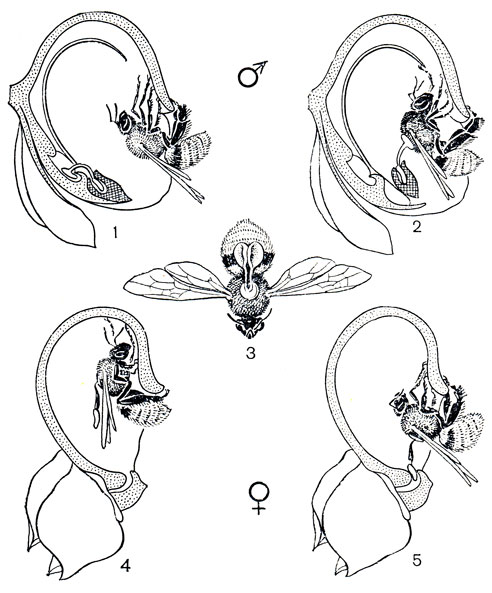
Рис. 153. Схема опыления катасетума крупноязычкового (Catasetum macroglossum) с нересупинированными цветками: 1 - пчела в перевернутом положении вползает в мужской цветок, охватывая губу; 2 - пчела касается антенны, и поллинарий вместе с пыльником выстреливает на насекомое; 3 - при перелете на следующий цветок крышка пыльника слетает, а поллиний изменяет свое положение; 4 - пчела вползает в женский цветок; 5 - поллинии захватываются в рыльцевую полость, когда пчела покидает женский цветок
Отметим еще несколько способов опыления, связанных с подвижностью губы и цветков. У многих орхидных части губы подвижно сочленены друг с другом или губа соединена эластичным сочленением с основанием или выростом (ножкой) колонки и совершает колебательные движения на ветру. Мелкие невзрачные цветки булъбофиллума бородконосного (Bulbophyllum barbigerum) из Западной Африки в результате дрожания губы и булавовидных волосков на ее верхушке становятся похожими на маленьких трепещущих насекомых и привлекают опылителей-мух. А у цирропеталума украшенного (Cirrliopetalum ornatissimum) эффект аналогичного колебания цветков еще усиливается благодаря постоянному мерцательному движению ресничек на их боковых лепестках.
Дрожание цветков видов южноамериканского рода онцидиум (Oncidium) предназначено для использования в опылении сложных непищевых инстинктов насекомых. Самцы некоторых перепончатокрылых, например рода центрис (Centris), покидают гнезда и сидят в засаде, охраняя свою территорию от вторжения пришельцев. И когда желто-коричневые пятнистые цветки онцидиума начинают при порыве ветра дрожать, насекомые принимают их за вторгшихся представителей другого вида, подвергают внезапной атаке с целью изгнания соперников со своей территории и производят опыление.
В ряде других случаев подвижно сочлененная губа действует как механическое приспособление для приведения насекомого в контакт с колонкой.У американской наземной орхидеи калопогона красивейшего (Calopogon pulchellus), цветки которого напоминают стайку расположившихся на стебле розовато-лиловых бабочек, губа обращена вверх и заканчивается желтой бородкой. Пчелы летят на яркую бородку, цепляются за нее, и под действием тяжести пчелы губа, обладающая подвижным сочленением, прогибается вниз, приводя спинку пчелы в контакт с колонкой. А у многих видов орхидных с перевернутыми цветками губа способна благодаря своей раздражимости или смещению центра тяжести к внезапному движению вверх в направлении колонки, опрокидывающему насекомое спиной на рыльце и пыльник. Такое замечательное устройство имеют многие виды бульбофиллума, виды плакоглоттиса (Placoglottis), чувствительной подвижной губой известны австралийские представители трибы неоттиевых калеана (Caleana), дракея (Drakaea), спикулея (Spicullaea), птеростилис (Pterostylis). Цветок рода птеростилис с губой в двух положениях изображен на рисунке 150,6,7. Два лепестка и один из чашелистиков этого цветка образуют колпачок над колонкой, и, когда взметнувшаяся вверх губа закроет вход в этот колпачок, насекомое оказывается заключенным во временной тюрьме, из которой оно выпускается после того, как губа снова откинется и станет опять чувствительной к прикосновению. За это время пытающееся освободиться насекомое успевает извлечь поллинии или произвести опыление.
Приведенные примеры не дают, конечно, сколько-нибудь полного представления о бесконечно разнообразных и удивительных способах опыления орхидных. Интересующегося читателя мы отсылаем к увлекательной, выше цитированной книге Дарвина, многочисленным работам К. Додеона, Л. ван дер Пейла, Л. Нильсона, Р. Бейтса, М. Проктора и др.
Рассмотрим лишь еще некоторые общие аспекты опыления орхидных. Среди множества приспособлений к перекрестному опылению следует отметить одно, универсальное для семейства,- длительность цветения. Только у некоторых родов (стангопея, собралия, некоторые виды дендробиума) цветки остаются свежими 1-5 суток. У большинства орхидных цветки в ожидании опыления не увядают неделями и месяцами, до 2-3 месяцев у ванды, одонтоглоссума, онцидиума. У многих орхидных цветки в соцветии распускаются неодновременно и все соцветие имеет свежий вид еще более длительный срок. Иногда цветение его растягивается на целый вегетационный период, а новые ветви могут образоваться и на следующий год. В большинстве случаев соцветия выносятся далеко в пространство, для того чтобы они были видны и удобны для посещения летающим насекомым, но иногда, наоборот, ось соцветия и цветоносы сильно укорачиваются и соцветия прижимаются к субстрату, для того чтобы опыление могли производить ползающие насекомые. Пыльца у большинства представителей семейства находится в связанном состоянии и не может разноситься ветром. Да и рыльца не приспособлены для улавливания пыльцы из воздуха. Орхидные поэтому опыляются только животными (насекомыми и птицами) или самоопыляются. Как установлено К. Додсоном (1966), основную часть (50%) опылителей семейства составляют перепончатокрылые (Hymenoptera), особенно те из них, для которых пищей является нектар. На втором месте (18%) - чешуекрылые (Lepidopterа), особенно ночные бабочки, на третьем (12%)-двукрылые (Diptera). Колибри как опылители составляют всего 3%, установлено опыление ими красных цветков дизы (Disa), масдеваллии (Masdevallia), эллеантуса (Elleanthus) и др. Некоторые крупные роды орхидных, например эпидендрум (Epidendrum), могут опыляться разными опылителями, но многие другие роды адаптированы к опылению преимущественно одной группой насекомых. Так, большинство видов цимбидиума и дендробиума опыляют пчелы, многие виды масдеваллии, бульбофиллума с губой, часто превращенной в ловчий аппарат, опыляют мясные мухи. Любку, ангрекум, хабенарию опыляют ночные бабочки, которые способны парить в воздухе и не нуждаются в посадочной площадке (губа у растений этих родов утратила свою функцию и либо обращена вверх, либо рассечена).
Бесконечно разнообразные форма и окраска околоцветника, различные вспомогательные устройства способствуют успеху перекрестного опыления орхидных. Они предлагают красные тона птицам, все оттенки красок - пчелам, грязно-фиолетовые или коричневато-красные - мясным мухам, снежно-белые - ночным бабочкам. А запахи их варьируют от запаха гниющего мяса до тончайшего аромата духов, исходящего часто от невзрачных и некрупных зеленовато-белых цветков.
Пыльца как пищевой фактор не имеет значения в опылении орхидных. Она экономично упакована в поллинии, не тратится впустую и у большинства видов вся целиком попадает на рыльце, что способствует оплодотворению сотен тысяч семязачатков, которые находятся в каждой завязи и из которых разовьются сотни тысяч семян. Основной "товар", который предлагают насекомым растения с помощью яркой "рекламы",- это нектар. Иногда он открытый и выделяется в бороздках губы, у основания колонки, или он скапливается в глубине рыльцевой полости или в шпорцах, отходящих от основания губы, иногда от чашелистиков. У некоторых орхидных нектарники осевые, они пронизывают на разную глубину стенки завязи и иногда пузыревидно вздуты. Разная длина шпорцев соответствует разной длине хоботков опылителей. Поразительной длины достигают шпорцы у некоторых видов ангрекума, аэридеса (Aerides), аэрангиса (Aerangis). У ангрекума полуторафутового (Angraecum sesquipedale) их длина составляет 21 - 30 см! Долго оставалось загадкой, какое же насекомое может достать нектар, скапливающийся на дне этого шпорца. И хотя Ч. Дарвин еще в 1877 г. предсказал существование гигантской бабочки с очень длинным хоботком, открыта она была на Мадагаскаре только в 1903 г. Бабочка ксантопан Моргана - предсказанная (Xanthopan morgani-praedicta) имеет свернутый в спираль хоботок длиной 22,5 см. Эта или подобные ей бабочки вполне могут достать нектар из длинного шпорца ангрекума.
Иногда насекомые отгрызают волоски и разнообразные выросты на губе растения. Известны случаи собирания ими воска с губы орнитидиума (Ornithidium). У эрии (Eria), полистахии (Polystachia) и максиллярии (Maxillaria) на губе образуется пыльцевидный порошок, поедаемый насекомыми. Но не всегда они находят в цветках то, что ищут: шпорцы оказываются пустыми, желтые, похожие на наполненный пыльцой пыльник пятна на губе - окрашенными волосками, а заманчивые резервуары с жидкостью - хитрыми ловушками, из которых не выбраться без "принудительной работы" по опылению. Обман - это еще один и весьма эффективный способ привлечения насекомых к опылению в этой группе растений. Но наиболее замечательным приспособлением можно считать использование ими в опылении непищевых инстинктов насекомых (сексуальных, защитных) и широкое привлечение к опылению самцов пчел и ос, т.е. группы насекомых, обычно не участвующей в опылении растений. Ф. Г. Бригер (1971), Р. Л. Дресслер (1981) объясняют такую своеобразную адаптацию тем, что семейство орхидных вступило на арену эволюции, когда уже существовало множество конкурентов, снабжавших опылителей пищей - нектаром и пыльцой, и орхидеи вынуждены были искать другие пути обеспечения опыления.
Казалось бы, при таком совершенстве приспособлений к опылению результатом должна быть 100-процентная завязываемость семян. На самом деле это сверхсовершенство оборачивается обратной стороной. Именно в силу очень узкой специализации цветки орхидных часто остаются неопыленными. Ч. Дарвин приводит следующие цифры и наблюдения: в тропических лесах Южной Бразилии у эпидендрума на 370 цветков образовалась только одна коробочка, 223 цветка опали неопыленными, а из 146 оставшихся только у 4 были удалены поллинии. Очень редко образует коробочки обильно цветущая в Бразилии ваниль, не более чем 1 цветок из 1000 дает коробочку у дендробиума красивого (Dendrobium speciosum) в Новом Южном Уэльсе. Только 5 коробочек на 200 цветков завязалось у кориантеса трехлопастного (Coryanthes triloba). Не очень плодовиты и орхидные в Европе. Многие из них прибегают к самоопылению для поддержания своего существования. Не являются ли в таком случае все достижения длительного приспособительного развития орхидных по отношению к определенным насекомым своего рода "ошибкой эволюции"? Ответом на этот вопрос служит тот факт, что орхидные - одно из двух крупнейших семейств цветковых растений, освоившее почти все экологические ниши на Земле. Ограниченные в силу узкой специализации возможности опыления компенсируются у них невероятно высоким количеством семян, которое образуется в каждой коробочке. Ч. Дарвин подсчитал, что если бы все семена палъчатокоренника пятнистого (Dactylorhiza maculata, табл. 33,5) благополучно прорастали, то правнуки одного растения "почти могли бы покрыть однообразным зеленым ковром всю поверхность суши на земном шаре". И того небольшого количества семян, которые превращаются во взрослые растения, орхидным вполне достаточно для продолжения рода в ненарушенных природных условиях.
Масса одного семени в коробочке орхидных составляет от десятых до тысячных долей миллиграмма. Около 76-96% объема семени занимает воздух. Ботаники XVI в., составители травников полагали даже, что орхидные вообще не имеют семян, а коробочки их содержат только пыль. Иероним Бок считал, что проростки орхидных появляются из животного семени птиц. И в XVII в. встречались еще утверждения, согласно которым орхидные вырастают не из семян, а из спермы животных.
Из раскрытых коробочек семена орхидных выдувает ветер. Помогают ему гигроскопические волоски, перемешанные с семенами в коробочках многих видов орхидных. Эти длинные волоски, сплетенные как войлок, при малейшем изменении влажности скручиваются и извиваются. Когда в сухую погоду коробочки открываются, волоски начинают свое интенсивное движение, перемещая семена ближе к поверхности, откуда их сдувает ветер. Сами коробочки также гигроскопичны и при увлажнении воздуха немедленно закрываются, чтобы раскрыться вновь при сухом ветре.
У наземных орхидных коробочки, как правило, направлены вверх, у эпифитных они расположены горизонтально или повисают. Поникающее положение коробочек у эпифитов способствует тому, что семена понемногу падают вниз, задерживаясь на многочисленных ветвях лесных деревьев, которые они встречают на пути и в коре которых они могут найти подходящее место для прорастания. Воздушные потоки выдувают семена наземных орхидных из вверх торчащих коробочек и относят их от материнского растения. Только у крупных наземных орхидных, растущих на открытых пространствах, коробочки могут быть поникающими.
Цветоносы в процессе развития коробочек часто вытягиваются в длину, в особенности если соцветия густые или если растения очень низкие, полуспрятанные в мох или лесную подстилку. Шлемовидные цветочки крошечной австралийской орхидеи корибаса расширенного (Corybas dilatatus), растущей колониями среди кустов или в расщелинах скал, едва приподнимаются над лежащим на почве ее единственным листочком. Коробочки же ее на сочных цветоносах выступают на несколько сантиметров вверх. У некоторых сапрофитных орхидных, растущих в густой тени лесов, цветонос при плодоношении удлиняется в 15-30 раз, увеличивая семенам возможность подальше отлететь от материнского растения.
Легкие, почти невесомые семена орхидных способны преодолевать большие расстояния. Достаточно сказать, что 4 вида орхидей были среди первых растений, поселившихся на острове Кракатау после катастрофического из-вержения вулкана.
У очень немногих видов орхидных в распространении семян участвуют животные. Млекопитающие и птицы поедают сочные стручковидные плоды ванили. Возможно, животные поедают и ягодовидные коробочки некоторых апостасиевых. И семена в этих сочных плодах имеют плотную, а не воздушную оболочку, характерную для распространяемых ветром семян.
У некоторых эпифитных орхидных, особенно растущих на муравьиных гнездах, в оболочке семян содержатся капельки масла, и такие семена растаскивают муравьи. У болотных орхидных семена могут распространяться водой. Имеет значение в жизни орхидных и вегетативное размножение. Многие из них завоевывают новые территории с помощью расползающихся корневищ и их отпрысков. А у маленькой торфяной орхидеи из Северной Европы гаммарбии болотной (Hammarbia paludosa) для размножения служат крошечные округлые выводковые почки, располагающиеся по краю листа. Псевдобульбы и куски корневищ орхидных иногда вымываются штормами на берег и, по крайней мере у некоторых видов, способны после путешествия в соленой воде утвердиться на новом месте.
Начало орхидологии восходит к древней Греции, когда Теофраст (IV - III вв. до н. э.) в своем "Исследовании о растениях" впервые употребил греческое слово orchis для обозначения одного из этих растений, а затем в I в. н. э. это же название Диоскорид дал двум растениям в своих сочинениях о лекарственных травах. Orchis - по-гречески означает "яичко" и указывает на сходство парных подземных клубней орхидных с яичками животных.
Интерес к орхидеям в Европе, где они не отличаются пышной красотой, долгое время определялся только их предполагаемыми лекарственными свойствами. Очень рано возник интерес к орхидным в Азии и Америке. В Китае они были введены в культуру более 1000 лет назад, а в научной литературе они упоминаются уже в III в. н. э. Во времена Сунской империи (960-1279) выпускались монографии по орхидным, в частности, по роду цимбидиум (Cymbidium, табл. 37,2), где описывались уже и методы его культуры. Орхидеи - излюбленный мотив в древнекитайской живописи, они изображались также в росписи по фарфору, в вышивках по шелку. В травнике ацтеков (Badianus manuscript, 1552) содержится самое раннее в Америке изображение орхидей и, кроме изображенного рода ваниль, упоминаются еще несколько орхидных.

Таблица 37. Орхидные: 1 - цирропеталум украшенный (Cirrhopetalum ornatissimum), оранжереи Ботанического института АН СССР в Ленинграде; 2 - цимбидиум гигантский (Cymbidium giganteum), оранжереи Батумского ботанического сада; 3 - онцидиум тигровый (Oncidium tigrinum), оранжереи Главного ботанического сада АН СССР

Таблица 38. Орхидные: 1 - стангопея тигровая (Stanhopea tigrina), оранжереи Ботанического института АН СССР в Ленинграде; 2 - стангопея глазковая (S. oculata), там же; 3 - ванда трехцветная (Vanda tricolor), там же; 4 - ренантера Имшота (Renanthera imschootiana), оранжереи Главного ботанического сада АН СССР
Первые экзотические орхидные попали в Европу в конце XVI - начале XVII вв. Со слов путешественников было известно, что орхидеи растут на деревьях, и долгое время в Европе полагали, что они являются паразитами и на их культивирование смотрели как на безнадежное дело. Тем более ценились единичные экземпляры, привезенные из дальних путешествий и бережно сохранявшиеся в частных коллекциях. Интерес к орхидеям все возрастал, и люди отправлялись за ними в дальние страны. Охотников за орхидеями подстерегали в девственных лесах тропиков многочисленные опасности, немало из них поплатилось своей жизнью за добывание новых видов. Но игра стоила свеч! Цены на орхидеи в XVIII-XIX вв. были необычайно высоки. Цена некоторых особо редких орхидей превышала десятилетний заработок уэльского шахтера. Уникальность каждого экземпляра, невозможность вырастить орхидеи из семян (о необходимой для этого микоризе тогда еще не знали), трудности добычи и транспортировки оправдывали эти высокие цены. За выгодным товаром снаряжали дорогостоящие экспедиции. Одни добывали таким способом огромное состояние, других ждало разочарование.
В начале XX в. была открыта возможность выращивания орхидей из семян, заражая их эндофитным грибом от материнского растения. Были разработаны способы их выращивания на искусственных питательных средах с добавлением углеводов и других органических веществ. Такие открытия положили начало культуре орхидей в широких масштабах. Этому способствовало также усовершенствование оранжерей в связи с введением новой системы отопления (водой по трубам).
Одновременно с освоением семенного размножения открылась новая сторона в культуре орхидей - возможность получения гибридов, превосходящих красотой и грацией родительские формы. Первый межвидовой гибрид калапта Домини (Calanthe X dominii) зацвел в 1856 г. Он был выведен английским садоводом-селекционером Джоном Домини путем скрещивания С. masuca X С. furcata (Каланта масука X Каланта вильчатая). А первый меж-родовой гибрид - лелиокатлея (Laeliocattleya) - был выведен в 1863 г. Число гибридных орхидей быстро росло, в гибридизацию вовлекались все новые компоненты, были получены гибриды трех и даже четырех родов. Сейчас в некоторых родах искусственных гибридов насчитывается больше, чем чистых видов. Документированные списки тысяч новых гибридов публикуются ежемесячно специальными журналами, свидетельствуя о терпеливой и целеустремленной работе селекционеров-орхидологов, тратящих десятилетия на опыты по получению гибридных растений.
Способность к гибридизации выражена у орхидных и в природе. Впервые гибридные формы были открыты у наземных орхидей в европейской флоре. Это были гибриды кокушника длиннорогого и нигрителлы черной (Gymnadenia conopsea X Nigritella nigra), обнаруженные в Альпах вблизи Гренобля. Позднее были открыты многие другие естественные межвидовые и межродовые гибриды как у наземных, так и у эпифитных орхидей. Случалось, что после получения искусственного гибрида совершенно идентичный гибрид находили и в природе.
В настоящее время в совершенстве освоено разведение орхидей в оранжереях и комнатах, а выращивание этих растений стало одним из ведущих направлений в цветоводстве многих стран. Многочисленные любители и специалисты объединяются в общества (сейчас их в мире более 400), издают специальные журналы по орхидным. Существуют общества даже по отдельным родам орхидных. Созываются локальные и международные конференции, устраиваются выставки. За наиболее выдающиеся достижения в селекции орхидей учреждаются премии. Наиболее развита культура орхидей в США, особенно в Калифорнии, где известные фирмы выращивают огромное количество цимбидиумов, каттлеи, фаленопсиса и других орхидных. Фирмы продают также проростки, семена и все необходимое для выращивания орхидей.
Получение тропических орхидей больше не связано для европейцев с риском: многочисленные фирмы в тропических странах производят сбор орхидей в лесах и выращивают их на экспорт. Товар отправляется в заморские страны большими партиями, и случается, что европейцы открывают теперь новые виды не в тропических лесах, а в витринах магазинов. Так было, например, с открытием одного из видов пафиопедилума, который был замечен двумя любителями-орхидологами в витрине магазина в Золингене среди растений другого, известного уже вида - пафиопедилума мозолистого (Раphiopedilum callosum). После тщательного анализа, сделанного специалистами, выяснилось, что это неизвестный науке вид. Место его сбора и экологию уточнил по запросу управляющий фирмой-экспортером в Северо-Восточном Таиланде Сукхакул, в честь которого вид и был назван пафиопедилум Сукхакула (Р, sukhakulii).
Экспорт наиболее популярных видов орхидей в отдельных странах измеряется сотнями тысяч экземпляров в год. Значительную часть этого количества в прежние годы составляли орхидеи, добытые в природе. И хотя сейчас хищническое истребление орхидей сократилось в связи с хорошей организацией их культуры, примерно 1/3 количества их видов находится под угрозой вымирания. Этому способствуют также и интенсивная вырубка лесов, проведение сельскохозяйственных мероприятий, промышленное загрязнение и любые другие разрушения естественных местообитаний, к чему орхидные особенности чувствительны. Многие виды орхидных внесены в списки охраняемых растений. Во многих странах выработано специальное законодательство, ограничивающее сбор и экспорт орхидей. Организованы природные заповедники, идут работы по вегетативному размножению видов, находящихся под угрозой исчезновения.
Подлежат охране и многие из наших местных орхидей - офрис, ятрышник, циприпедиум,) пыльцеголовник, анакамптис (Anacamptis), комперия (Comperia) и многие другие. Выкапывание растений, сбор их на букеты, приводящий к истощению их подземных органов, наносят непоправимый вред природным популяциям этих редких и замечательных растений. И если вам встретится на пути это маленькое чудо природы - цветущая орхидея - и вы протянете руку, чтобы в один миг ее сорвать,) остановитесь и вспомните, что, может быть,, именно этому растению потребовалось около десяти лет, чтобы пройти длинный и сложный путь от семени до первого цветения!
Орхидные называют семейством "аристократов" среди растений. Они обогащают духовный мир человека, как это делают шедевры искусства. Орхидеи-птицы, орхидеи-бабочки, -гномы,, -ящерицы, -лягушки, -пауки, -медузы, -лебеди! Где еще найдутся такие необычайные формы, такое буйство красок, такие удивительные запахи? За красоту и уникальность многие страны выбрали местные орхидеи своими национальными символами. Орхидея-голубь, или орхидея - "святой дух" (перистерия высокая - Реristeria elata),- национальный цветок Панамы. В центре этого снежно-белого цветка как бы сидит кроткий голубь со слегка приподнятыми крыльями - такую форму имеет здесь колонка. Испанские монахи, впервые увидевшие перистерию в Мексике, сочли ее цветок за воплощение святого духа и использовали как иллюстрацию в своем учении. Индейцы и до сих пор поклоняются этому цветку. Виды каттлеи выбраны национальными цветками Коста-Рики и Венесуэлы, ликаста (Lycaste) - национальный цветок Гватемалы.
Орхидеи нередко изображаются на почтовых марках. Первая марка с изображением ванили плосколистной появилась в 1905 г. в Гваделупе. А к настоящему времени 86 стран выпустили уже свыше 400 разных марок с изображением орхидей.
Немногие орхидные имеют чисто практическое значение в жизни людей. Ваниль - первое американское орхидное, которое стало известно европейцам. Испанские конкистадоры, возглавляемые Кортесом, обнаружили эту дикорастущую лиану на юго-восточном побережье Мексики. Ваниль, или "черный цветок" на языке ацтеков, использовалась индейцами для ароматизации шоколада и приготовления напитка, который "придает телу силу гладиатора, уносит прочь усталость, прогоняет страх и укрепляет сердце". Уже в начале XVI в. была сделана первая малоуспешная попытка интродуцировать ваниль в Европу. Успех был достигнут только в 1807 г. в Англии, откуда затем ваниль, размноженная вегетативно, разошлась по садам других стран Европы. В условиях культуры ваниль не давала плодов, да и на родине она образует их в небольшом количестве. Попытки вместе с ванилью интродуцировать некоторые виды возможных ее опылителей не увенчались успехом. И только когда стали производить опыление ванили искусственным путем, она начала давать урожай. Теперь этот метод используется также и на родине растения.
Используемые как пряность плоды ванили плосколистной (Vanilla planifolia), известные в коммерции как "бобы ванили", представляют собой длинные сочные цилиндрические коробочки длиной 10-25 см и шириной 0,8-1,5 см. После сбора их подвергают длительному процессу ферментации и сушки, в результате чего содержащийся в плодах гликозид глюкованилин расщепляется на глюкозу и свободный альдегид ванилин, после чего плоды приобретают запах. Сморщенные бурые "палочки ванили" употребляют в пищевой промышленности и в медицине для ароматизации лекарств. Дикорастущая ваниль плосколистная встречается в Юго-Восточной Мексике, Вест-Индии, Центральной Америке и на севере и западе тропической Южной Америки. Она представляет собой крупную, забирающуюся до вершин деревьев лиану (рис. 154) с мясистыми, почти сидячими удлиненно-эллиптическими или ланцетными листьями и пазушными соцветиями из зеленовато-желтых цветков. Промышленная культура ванили развита на Мадагаскаре, в Мексике, на островах Малых Антильских, Индийского и Тихого океанов, Малайского архипелага и др. Кроме ванили плосколистной, для получения ванилина используют еще американскую ваниль помпонную (V. pompona) и растущую на острове Таити в Тихом океане ваниль таитянскую (V. tahitensis). Их плоды, однако, ценятся гораздо ниже. В настоящее время значительную часть ванилина получают синтетически. Синтетический ванилин дешевле, но обладает менее приятным ароматом.
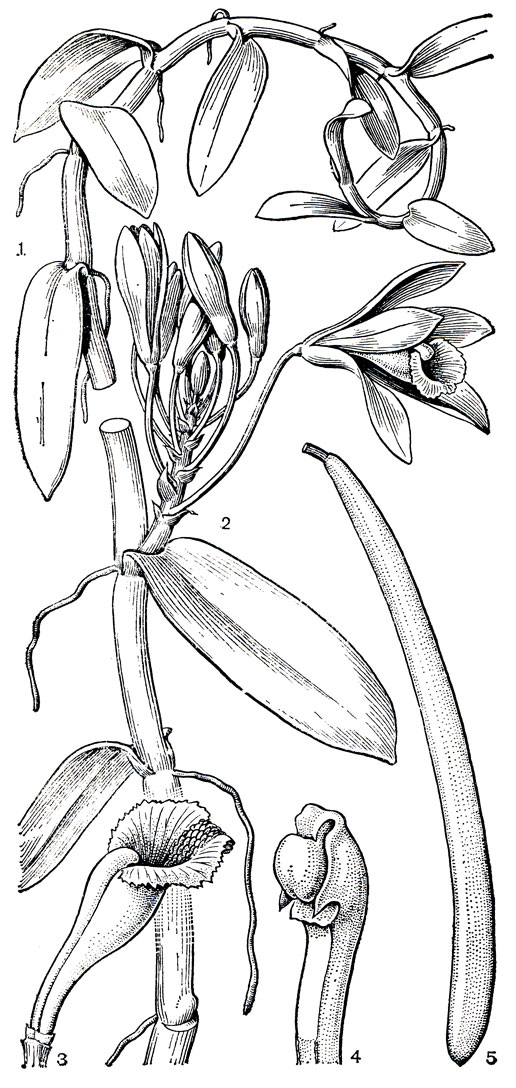
Рис. 154. Ваниль плосколистная (Vanilla planifolia): 1 - молодой стебель с воздушными корнями; 2 - часть стебля с соцветием; 3 - губа, свернутая в трубку и заключающая колонку; 4 - колонка; 5 - плод
Некоторые орхидные нужны человеку как лекарственные растения. В медицине применяют салеп - высушенные корневые клубни видов ятрышника, пальчатокоренника, любки, кокушника и др. Как лекарственные растения, а также для приготовления ароматного чая применяют и многие другие орхидеи. Стебли некоторых орхидей используют в плетении и для изготовления музыкальных инструментов.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'