Порядок бромелиевые (Bromeliales) (А. Л. Тахтаджян)
Семейство бромелиевые (Bromeliaceae)
Это одно из самых больших семейств однодольных, уступающее только орхидным, осоковым, злакам и пальмам. Оно насчитывает 46 родов и не менее 2100 видов. Почти все бромелиевые распространены в Новом Свете и лишь один вид - питкерния плодовитая (Pitcairnia feliciana) - встречается в тропической Западной Африке (Гвинея) совершенно изолированно от основного ареала семейства. В Америке бромелиевые приурочены главным образом к тропическим областям, особенно к бассейну Амазонки, где находится центр их разнообразия. Лишь немногие виды выходят за пределы тропиков, но и они ограничены рамкой тепло-умеренной зоны. Дальше всех на север продвинулась тилландсия уснеевидная (Tillandsia nsneoides), или так называемый "испанский мох" (рис. 155). Это похожее на лишайник уснею эпифитное бромелиевое доходит вдоль Атлантического побережья Северной Америки приблизительно до 38° с. ш. в Восточной Вирджинии. В Южной Америке несколько видов бромелиевых доходит до островов Хуан Фернандес и до 44° ю. ш. в Чили и Центральной Аргентине.

Таблица 42. Бромелиевые: 1 - эхмея прицветниковая (Aechmea bracteata), Куба; 2 - тилландсия уснеевидная (Tillandsia usneoides), Флорида; 3 - бромелия пингвин (Bromelia pinguin), Куба; 4 - акантостахис шишковидный (Acanthostachys strobilacea), оранжереи Ленинградского государственного университета
Бромелиевые встречаются в самых различных местообитаниях: от дождевых лесов до пустынь и от морских побережий до высокогорий. Они растут на плодородной почве, на песках, на скалах, деревьях и на засоленном субстрате, на регулярно затопляемых берегах рек и даже на телеграфных проводах. Большинство наземных бромелиевых является ксерофитами. Суккулентные гехтии (Hechtia) растут на сухих почвах Южного Техаса и Мексики, некоторые виды бромелии (Bromelia) встречаются на песках вдоль большей части побережья Восточной Южной Америки, а в прибрежных пустынях Перу виды тилландсии (Tillandsia) густо покрывают грунт при почти полном отсутствии других высших растений. Очень многие бромелиевые являются скальными растениями, и в Юго-Восточной Бразилии огромные пространства гранитных утесов покрыты видами тилландсии и фризеи (Vriesea). Энхолириум (Encholirium) и диккия (Dickia) в Бразилии и девтерохния (Denterochnia) и аброметиелла (Abrometiella) в Аргентине обычно растут на голых скалах под палящим солнцем. В Чили виды тилландсии вслед за лишайниками и мхами являются одними из первых пионеров при заселении обнаженных скал. Если лесное бромелиевое фостерелла (Fosterella), произрастающая в условиях высокой влажности и тепла, требует очень мало света, то виды пуйи (Puya), растущие на открытых вершинах Анд, получают максимум солнечного света и легко переносят очень резкие суточные колебания температуры. Многие бромелиевые приспособлены к местообитаниям, характеризующимся дефицитом азотистого питания. В пределах рода питкерния (Pitcairnia) можно наблюдать весь спектр приспособлений к самым различным местообитаниям, начиная от дождевого леса и кончая пустыней.
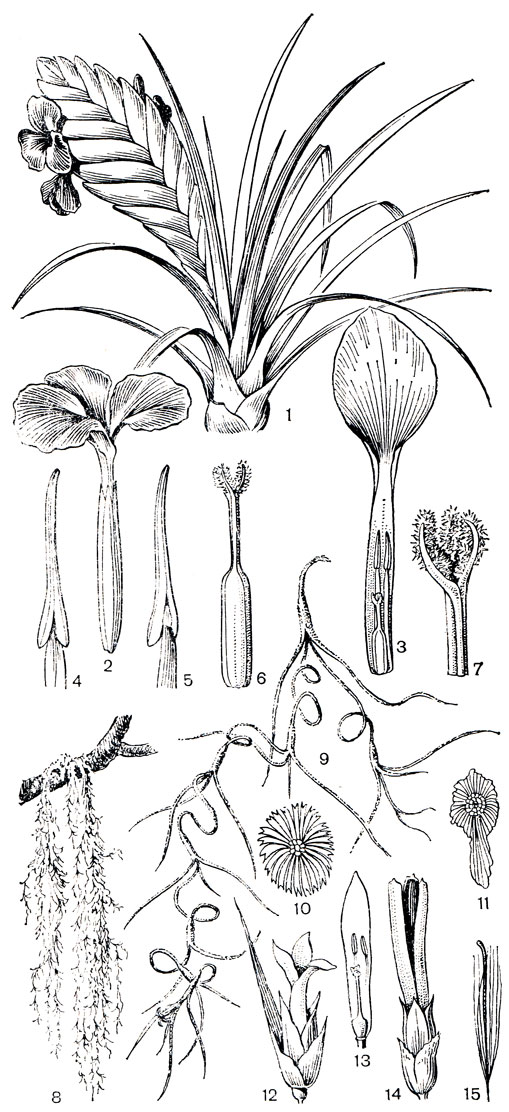
Рис. 155. Бромелиевые. Тилландсия синяя (Tillandsia cyanea): 1 - часть растения; 2 - цветок; 3 - лепесток с тычинками; 4, 5 - тычинки; 6 - гинецей; 7 - верхняя часть гинецея. Тилландсия уснеевидная (Т. usneoides): 8, 9 - общий вид; 10, 11 - чешуевидные волоски; 12 - цветок; 13 - часть цветка; 14 - коробочка; 15 - семя
Почти все бромелиевые - многолетние травы, очень редко - кустаршшовидные растения (девтерохния). Они обычно с очень укороченным стеблем, но многие тилландсии имеют удлиненный стебель, а виды рода пуйя имеют даже более или менее деревянистый стебель, который у пуйи Раймонда (Рuуа raimondii) достигает в высоту 9,5 м при толщине более 1 м и внешне несколько напоминает гигантские лобелии тропической Африки. Первичный корень бромелиевых отмирает очень рано, но у них развиваются придаточные корни. Строение корня зависит от того, служит ли он для прикрепления к субстрату и для питания, как у наземных видов, или же только для прикрепления к субстрату, как у эпифитов. У относительно самых примитивных бромелиевых (подсемейство питкерниевые - Pitcairnioideae) корни исполняют обычные для них функции поглощения воды и питательных веществ через корневые волоски. У большинства представителей этого подсемейства корни базальные, но у видов пуйи с простертыми стеблями они образуются в самых разных их частях. У представителей двух других подсемейств (тилландсиевых - Tillandsioideae и бромелиевых - Bromelioideae) корни утеряли большую часть своей функции поглощения и служат лишь для прикрепления к субстрату. У некоторых же видов тилландсии, включая "испанский мох", корни утеряли даже якорную функцию и атрофировались. У некоторых видов бромелии корни не имеют контакта с почвой и заканчиваются в пазухах листьев.
Листья у бромелиевых очередные, обычно многорядные, но у некоторых тилландсий и у нескольких видов подсемейства бромелиевых - двурядные. Большей частью они расположены в базальных розетках, реже стеблевые, у основания более или менее расширенные во влагалище, цельнокрайные или по краям колючепильчатые, обычно более или менее ксероморфные, с толстой кутикулой и хорошо развитой водозапасающей паренхимой между эпидермой и хлоренхимой. Для листьев бромелиевых очень характерны особые пельтатные (щитковидные) чешуи, состоящие из ножки и однослойного щитка; в процессе развития клетки щитка в конце концов отмирают, но клетки ножки могут остаться живыми в течение всей жизни листа. Отдельные клетки щитка неравномерно утолщены. Наиболее специализированные чешуи характерны для тилландсиевых. У представителей подсемейства питкерниевые и у длинностебельных видов тилландсии влагалища листьев не бывают сильно расширены и тесно налегают друг на друга. Чешуи на листьях служат у них только для уменьшения транспирации. У большинства видов подсемейства бромелиевых листовые влагалища расширенные и краями плотно охватывают друг друга, образуя "чаши" или "вазы", в которых во время дождей накапливается часто довольно много воды. Цистерна у растения может быть одна общая, образованная всеми листьями (одноцистерновый тип, как у билъбергии - Billbergia) или же чаще цистерна образуется у основания каждого листа (многоцистерновый тип). В эти цистерны попадают и в конце концов растворяются в них обычно довольно значительные количества органического вещества в виде микроорганизмов, отмерших листьев и других частей растений, мертвых животных (особенно насекомых), а также выделений живых водных личинок. Вода вместе с растворенными в ней питательными веществами всасывается придаточными корнями, развивающимися между основаниями листьев, или же чешуями, развивающимися на внутренней стороне листовых оснований, и таким образом растение получает как воду, так и дополнительную азотистую пищу. Поглощению азота способствуют гнилостные бактерии, переводящие органические вещества в растворимую форму (аминокислоты, амиды, мочевина). У более специализированных форм этого цистернового типа листовые чаши крупнее и поглощение воды вместе с растворенными в ней веществами осуществляется не придаточными корнями, а чешуями. В таких цистернах иногда может накапливаться очень много воды, часто несколько литров. У фризеи гигантской (Vriesea gigantea) бывает более 5 л воды, а у гломеропиткернии (Glomeropitcairnia) было установлено даже около 20 л жидкости.
Многие виды тилландсии с длинным стеблем, в том числе и "испанский мох", не образуют листовых цистерн. Они поглощают воду из атмосферы посредством чешуй, которые через живые клетки их ножек осмотически передают ее внутрь листа. При высыхании чешуи сморщиваются, что не мешает газовому обмену через устьица, но уменьшает испарение с поверхности листа. Благодаря такому приспособлению растение может пережить сухое время года. Остальные представители тилландсиевых эволюционировали в другом направлении - у них образовались листовые цистерны и чешуи, абсорбирующие воду, сосредоточенные преимущественно на влагалищах листьев. Подсемейство собственно бромелиевых также эволюционировало в двух направлениях, но у них не наблюдается столь далеко зашедшей ксероморфности, как у некоторых тилландсиевых. У большинства родов решение проблемы водного режима достигается посредством развития листовых цистерн, варьирующих от широких чаш до тонких труб.
С развитием листовых цистерн связано возникновение очень своеобразных биологических связей, неизвестных у других однодольных. В цистернах постоянно обитает много бактерий, цианобактерий, водорослей, высших растений и особенно животных. Они являются постоянным местообитанием листостебельного мха хукерии бромелиефильной (Hookeria bromeliophila). В цистернах крупных тилландсий часто встречаются пузырчатка почковидная (Utricularia reniformis) и пузырчатка лотосолистная (U. nelumbifolia), где они образуют столоны (плети), достигающие чаш соседних тилландсий. Пузырчатка Гумбольдта (U. humboldtii) приурочена к цистернам броккинии кордилиновидной (Brocchinia cordylinioides). Но в цистернах гораздо более многочисленны (не менее 350 видов) и играют гораздо большую роль представители животного мира. В них встречаются различные простейшие, черви, моллюски, членистоногие, большое число разнообразных насекомых (особенно мух) и даже позвоночные, как саламандры, лягушки (большинство из рода квакша - Hyla), ящерицы и змеи. Земноводные не только размножаются в этих цистернах, но ночные формы находят здесь днем укрытия. Есть даже указание, что в полуаридной Северо-Восточной Бразилии один вид квакши находится в своего рода симбиотических отношениях с некоторыми видами бильбергии, в длинных трубчатых цистернах которых он живет. Эти квакши не только отсиживаются здесь в дневное время, но очень эффективно закрывают отверстие своей плоской головой и тем самым сохраняют влагу как для себя, так и для растения. Интересно также, что один вид квакши (квакша мелкожилковатая - Hyla venulosa) проводит все сухое время года в цистерне бильбергии зебровой (Billbergia zebrina). По мнению Б. Смита (1974), лягушки, возможно, являются важным фактором отбора в эволюции трубчатых цистерн некоторых видов бильбергии. В цистернах различных видов бромелиевых часто находят змей, а иногда также ящериц, проникающих туда в поисках пищи (насекомых, лягушек, саламандр).
Вегетативные органы бромелиевых имеют и целый ряд других приспособлений. Так, например, у некоторых видов тилландсии листья образуют крючки, посредством которых прикрепляются к ветвям поддерживающего растения, а у других видов превращаются в усики, которые обвиваются вокруг опоры. Многие наземные бромелиевые образуют ветвистые столоны, посредством которых они часто размножаются столь интенсивно, что цветут редко и еще реже плодоносят, как, например, род криптантус (Cryptanthus).
Сосуды у одних бромелиевых имеются во всех органах, у других - только в корнях и стеблях или только в корнях. Перфорация сосудов лестничная или реже простая.
Разнообразие вегетативных органов бромелиевых тесно связано с характером минерального питания и водного режима. На этом основании еще в прошлом веке известный немецкий ботаник А. Ф. В. Шимпер (1888) различал три биологических типа бромелиевых: 1) наземный, 2) эпифитный, накапливающий воду в цистернах, и 3) эпифитный, перешедший к поглощению атмосферной влаги через свои листья. Позднее американский ботаник Колин Питтендриг (1948), основываясь на своих исследованиях на острове Тринидад, переработал классификацию Шимпера и развил ее. Питтендриг, а вслед за ним Д. Бензинг (1980) в своей книге "Биология бромелиевых" различают следующие четыре биологических типа бромелиевых.
К первому типу относятся бромелиевые, добывающие воду и минеральные вещества непосредственно из почвы, для чего им служит хорошо развитая корневая система. Они не имеют цистерн и их листья играют лишь ничтожную роль или даже не играют никакой роли в добывании как влаги, так и солей. Чешуи на их листьях не обладают еще абсорбирующей способностью и служат лишь для уменьшения транспирации. К этому почвенно-корневому типу относится большинство представителей подсемейства питкерииевых, включая питкернию гехтию и фостереллу, и ряд представителей подсемейства собственно бромелиевых, например, некоторые виды бромелии.
Второй биологический тип бромелиевых, который можно назвать цистерново-корневым, отличается от почвенно-корневого типа прежде всего умеренно расширенными основаниями листьев, образующих цистерны. Чешуи на листьях бромелиевых этого типа хотя и могут поглощать воду и минеральные вещества, но лишь в небольших количествах. У некоторых цистерново-корневых бромелиевых корни не достигают почвы, но вместо этого растут вверх по направлению к основаниям листьев, где они внедряются в цистерну. Этот тип характерен для многих представителей подсемейства бромелиевых, включая бромелию и ананас (Ananas), и для очень немногих питкерииевых, как род броккиния (Brocchinia), произрастающий на торфяных болотах Гайанского нагорья. Кстати, есть подозрение, что это оригинальное бромелиевое - насекомоядное растение (Бензинг, 1980).
Следующий цистерново-бескорневой тип, характерный для многих тилландсиевых и ряда представителей подсемейства бромелиевых, имеет хорошо развитые цистерны, содержимое которых служит главным источником влаги и минеральных веществ в течение всей жизни растения, за исключением лишь ранних стадий развития. В отличие от предыдущего типа, пельтатные чешуи цистерново-бескорневых бромелиевых являются абсорбирующими структурами. Многие из представителей этого типа являются облигатными эпифитами, но довольно много также и факультативных эпифитов. Некоторые из них, как фризея клейкая (Vriesea gliitinosa) растут на скалах. Имеется очень интересная мирмекофильная разновидность этого цистерново-бескорпевого типа. У некоторых эпифитных видов эхмеи (Aechmea), например эхмеи Мертенса (A. mertensii), цистерны не являются вполне надежным источником влаги и питательных веществ. Поэтому эти виды поселяются на древесных муравейниках, из которых они извлекают значительное количество питательных веществ и влаги. При этом на одном и том же муравейнике могут расти разные виды эпифитов. К сожалению, биология этих "муравьиных садов" все еще недостаточно изучена и детали взаимоотношений компонентов этих своеобразных экосистем не вполне ясны.
Наконец совершенно особый биологический тип представляют так называемые "атмосферные" бромелиевые, или просто "атмосферики". Карл Мец (1935) назвал их "атмосферными", так как все необходимое они получают из атмосферы. Все атмосферики - суккулентные ксерофиты. К ним относятся несколько сот видов рода тилландсия и несколько видов очень близкого рода фризея. Атмосферики растут на деревьях, на скалах и даже на сухом песке. Они густо покрыты высокоспециализированными абсорбирующими чешуями, не имеют расширенных листовых оснований и корневая система у них обычно рудиментарная или иногда полностью отсутствует (за исключением проростков). Когда же корни имеются, то они служат только в качестве своеобразных крюков, прикрепляющих растение к субстрату. Влагу атмосферики поглощают непосредственно из воздуха, а минеральное питание они получают исключительно из воздушной пыли и дождевой воды. Одним из наиболее типичных атмосфериком является пустынная перуанская тилландсия пурпурная (Tillandsia purpurea), которая вполне довольствуется минеральными веществами, находящимися в приносимой ветром пыли, и влагой, доставляемой густым туманом, регулярно приходящим со стороны Тихого океана. Она вполне благополучно растет в жаркой пустыне, где, не имея корня, просто лежит на сухом песке под палящими лучами тропического солнца.
Особой разновидностью биологического типа атмосферных бромелиевых являются "мирмекофильные атмосферики". Сюда относятся тилландсия "голова Медузы" (Т. caput-medusae), тилландсия Бутца (Т. butzii), тилландсия луковичная (Т. bulbosa) и другие тилландсии с луковицеобразно расширенными основаниями листьев. В камерах этих "луковиц" живут муравьи, которые, в свою очередь, приносят растению двоякую пользу: они защищают его от различных вредителей и, кроме того, снабжают дополнительными питательными веществами. Питательными веществами служат продукты разложения различных вредителей, приносимых муравьями в камеры, и муравьиные экскременты.
Значительно больше половины всех бромелиевых являются эпифитами, факультативными пли облигатными. По мнению А. Ф. В. Шимпера (1888), эволюция эпифитных бромелиевых происходила первоначально внутри тропического дождевого леса. Через ряд промежуточных форм примитивные наземные бромелиевые дождевого леса перешли сначала на нижние ветви деревьев, а затем некоторые из них в результате дальнейшей специализации их чешуй постепенно приспособились к относительно сухому верхнему пологу. Большинство бромелиевых цистернового типа являются факультативными эпифитами. Как отмечает Э. Мак Виллиамс (1974), нет резкой границы между эпифитной и неэпифитной средой, а в дождевом лесу один и тот же вид может иногда произрастать на земле, на тенистом стволе дерева и в верхнем ярусе. Интересно, что, как указывает тот же автор, в Перу некоторые виды тилландсии растут в качестве эпифитов на кактусах. Но даже и в дождевом лесу условия произрастания эпифитных бромелиевых могут быть периодически крайне сухими. По мнению Э. Мак Виллиамса, успех бромелиевых в проникновении в полог дождевого леса и в другие ксерические местообитания связан с эффективностью таких адаптивных механизмов, как: 1) абсорбирующие чешуи; 2) накопление воды в цистернах, образованных листовыми основаниями; 3) суккулентность; 4) темновая фиксация углекислоты; 5) опадающие листья некоторых бромелиевых (некоторые питкернии и род айенсуа - Ayensua на Гайанском нагорье); 6) неотения и гетерофиллия и 7) некоторые особенности семени и его способность к прорастанию в условиях недостатка влаги. В разных группах бромелиевых преобладающее значение приобретают те или иные из этих приспособлений. Так, развитие абсорбирующих чешуй было ключевым фактором в успешной колонизации ксерических местообитаний тилландсиями а у большинства представителей подсемейства бромелиевых таким фактором было возникновение специальных резервуаров для воды.
Цветки бромелиевых собраны в простые или сложные кисти, колосья, головки или метелки, редко одиночные, обычно с хорошо развитым ярко окрашенным прицветником, который почти всегда более ярко окрашен, чем цветки. У некоторых бромелиевых цветущий стебель очень укорочен и соцветие сидит в центре розетки базальных листьев и обычно окружено покрывалом из ярко окрашенных прицветников. Удлиненные цветущие стебли могут нести листовидные стерильные прицветники, отличающиеся от обычных листьев только своей окраской, как у ананаса или у бильбергии, или же они могут образовать настоящий цветонос, несущий только несколько чешуевидных прицветников, расположенных непосредственно под цветками, как у тилландсий. В некоторых случаях цветущий стебель сохраняется в тече-ние нескольких лет, одревесневает и каждый новый сезон производит новое соцветие. Фертильные прицветники морфологически соответствуют листовым основаниям. Цветки обоеполые или иногда функционально однополые (некоторые виды гехтии и катопсиса - Catopsis), большей частью актиноморфные, почти всегда трехчленные, но в некоторых однобоких соцветиях, особенно висячих, наблюдается тенденция к зигоморфности, как, например, у питкернии. Околоцветник обычно с ясно различающимися между собой чашечкой и венчиком, каждый из которых состоит из 3 сегментов. Интересно почкосложение околоцветника, расположение его сегментов в цветочной почке: чашелистики скручены налево, а лепестки направо. Чашелистики зеленые и травянистые или более или менее лепестковидные, свободные или сросшиеся у основания. Лепестки почти всегда одинаковые, от линейных до яйцевидных, свободные или сросшиеся в короткую трубку, часто ярко окрашенные (белые, красные, желтые, зеленые или синие), как правило, снабжены на внутренней стороне и у основания парой язычковидных придатков, различающихся по форме и размерам у разных родов. К сожалению, функция этих придатков не вполне ясна. Тычинок 6 в двух кругах. В цветках со сростнолепестным венчиком нити тычинок более или менее сросшиеся с лепестками. Нити тычинок обычно узколентовидные, а пыльники линейные, подвижные, прикрепленные основанием или спинкой, вскрывающиеся интрорзно продольной щелью. Пыльцевые зерна однобороздные (подсемейство питкерниевые и тилландсиевые) или как однобороздные, так и 2 многопоровые (подсемейство бромелиевые). Гинецей из 3 плодолистиков, синкарпный; столбик обычно длинный, тонкий, на верхушке более или менее 3-лопастный; завязь верхняя, полунижняя или нижняя, 3-гнездная, обычно с более или менее многочисленными анатропными или редко кампилотропными семязачатками в каждом гнезде. Характерно наличие септальных нектарников, приуроченных к перегородкам, разделяющим соседние плодолистики, и занимающих места вдоль линий срастания; нектар выделяется через отверстие у основания столбика и задерживается чешуями у основания цветка. Плод обычно септицидная коробочка или чаще (подсемейство бромелиевые) ягода. У ананаса и близкого к нему рода псевдананас (Psendananas) отдельные ягоды срастаются в соплодие. Семена мелкие, с обильным мучнистым эндоспермом и большей частью маленьким, но иногда и довольно крупным зародышем. Семена, развивающиеся в плодах типа коробочка, обычно крылатые или с хохолковидным пучком волосков, образованным в результате расщепления удлиненного внешнего интегумента и верхушки семяножки, как у тилландсий.
Почти все бромелиевые - поликарпики, и лишь некоторые тилландсиевые и виды пуйи являются монокарпиками. Типичным монокарпиком является, в частности, гигантская пуйя Раймонда.
Бромелиевые протандричны. В то время, когда пыльники уже открыты и из них освобождается пыльца, рыльца спирально закручены в головку и таким образом еще не могут воспринимать пыльцу. Переносчиками пыльцы являются колибри, летучие мыши, различные насекомые (особенно пчелы, шмели, мотыльки и бабочки), а в некоторых случаях, по-видимому, также ветер. Опылителей привлекает обильный нектар, выделяемый септальными желёзками. Как количество, так и качество нектара (особенно содержание сахара) очень сильно варьирует у разных представителей семейства. Для птиц и летучих мышей обильный нектар в цветках многих бромелиевых имеет двоякое значение. Он является не только пищей, но и важным источником воды в сухое время года. Таково, несомненно, значение обильного, но сравнительно бедного сахаром нектара в цветках видов пуйи. Как отмечает М. Персиваль (1965), наблюдается замечательный параллелизм между уровнем нектара в цветочной трубке и длиной языка опылителя. Уровень нектара определяется чешуями на лепестках, представляющих очень важный таксономический признак для представителей подсемейства тилландсиевых и собственно бромелиевых. Очень многие бромелиевые, особенно виды с красными цветками, опыляются колибри, а некоторые из них, как виды фризеи и текофиллума (Thecophyllum), а также, вероятно, виды питкернии, раскрывающие свои цветки ночью и имеющие специфический запах, опыляются летучими мышами (Порш, 1932: С. Фогель, 1969).
В семействе бромелиевых преобладает перекрестное опыление, но немало видов перешло частично или даже полностью к самоопылению. Цветки некоторых видов гузмании (Guzmania) не раскрываются - они клейстогамны.
Семена бромелиевых обычно распространяются ветром, но у некоторых видов, произрастающих вдоль рек или в глубине дождевого леса, они распространяются, по всей вероятности, водой. Навия (Navia), семена которой лишены придатков, также является гидрохорным растением. Виды с семенами, лишенными придатков, но со съедобными сочными плодами являются преимущественно орнитохорами, некоторые из них распространяются также летучими мышами.
Многие виды бромелиевых размножаются главным образом вегетативно. У видов с розеточными листьями регулярно образуются вегетативные отпрыски. Виды с длинными стеблями вытягиваются, ветвятся и распадаются на отдельные особи, что особенно хорошо выражено у "испанского мха". Имеется много видов, включая произрастающую на прибрежных пустынях Перу тилландсию широколистную (Tillandsia latifolia), которые являются живородящими - вместо цвётков у них образуются маленькие растеньица.
Семейство бромелиевые состоит из 3 подсемейств - питкерниевые (Pitcairnioideae), тилландсиевые (Tillandsioideae) и собственно бромелиевые (Bromelioideae).
Питкерниевые - самое примитивное подсемейство бромелиевых. Это почти всегда наземные травы, редко кустарниковые растения (девтерохния), обычно с корнями, большей частью функционирующими. Листья обычно колючепильчатые. Завязь верхняя или почти верхняя и лишь у немногих видов питкернии и навии - нижняя. Плод всегда сухой, коробочка (обычно септицидная) или редко нераскрывающийся. Семена мелкие и легкие, почти всегда снабжены цельными придатками, которые отсутствуют лишь у питкернии афеландрицветковой (Pitcairnia aphelandriflora) и навии. В подсемействе 13 родов, из которых важнейшими являются пуйя, питкерния, навин и диккия.
В роде пуйя (рис. 156) около 170 видов, распространенных преимущественно в сухих областях, главным образом в Андах. Это наземные бесстебельные или чаще длинностебельные, часто гигантские травы; стебли простые или ветвистые, иногда вместе с соцветиями достигающие в высоту нескольких метров. Листья в густых розетках, кожистые, по краям колючие, обычно с широкими влагалищами. Чашелистики и лепестки свободные, после цветения закручивающиеся спирально. Тычинки свободные, несколько короче лепестков. Коробочка локулицидная и обычно также запоздало септицидная. Семена крупные, с дорсиапикальным крылом. Виды пуйи растут преимущественно на скалистых склонах в горах. Некоторые виды пуйи играют очень заметную роль в растительном покрове и нередко являются ландшафтными растениями. Многие виды имеют декоративное значение.
В близком к пуйе роде питкерния (рис. 156) около 260 видов. Они широко распространены в тропических областях Южной и Центральной Америки. Небольшие или крупные, большей частью надземные травы, бесстебельные или реже с более или менее длинными стеблями, очень разного облика. Некоторые виды питкернии имеют декоративное значение.

Рис. 156. Бромелиевые. Питкерния удивительная (Pitcairnia mirabilis) 1 - общий вид растения; 2 - лист; 3 - цветок; 4 - тычинка 5 - гинецей; 6 - верхняя часть гинецея; 7 - плод; 8 - семя Пуйя Бертеро (Puya berteroniana): 9 - общий вид растения; 10 - цветок; 11 - тычинка; 12 - гинецей; 13 - поперечный разрез завязи
Род навия (рис. 157), насчитывающий около 75 видов, распространен в Южной Америке в областях к северу от Амазонки. Он представлен травами очень разного облика и размера, от высоких раскидистых растений, достигающих в высоту 4-5 м, до низких, нередко подушковидных форм. Интересно, что один из низких, бесстебельных и, вероятно, однолетних видов навии (навия скальная - Navia rupestris) был описан известным американским ботаником Глисоном (1929) как вид тофилдии, т. е. был отнесен к совершенно другому семейству и даже другому порядку.
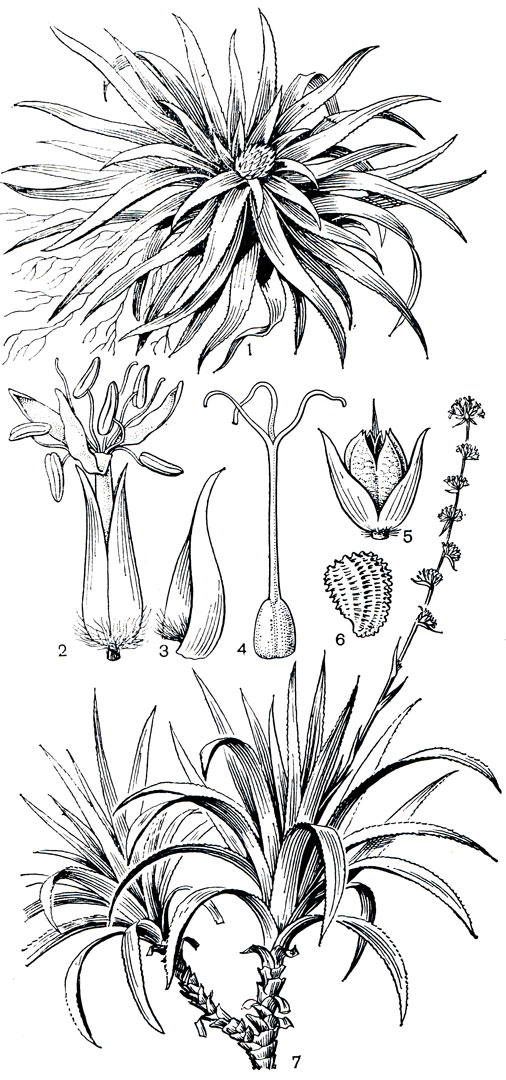
Рис. 157. Бромелиевые. Навия бесстебельная (Navia acaulis): 1 - часть растения; 2 - цветок; 3 - прицгетник; 4 - гинецей; 5 - плод; 6 - семя. Навия стеблевая (N. caulescens): 7 - часть растения
В роде диккия более 100 видов, распространенных исключительно в Южной Америке, главным образом в восточных и юго-восточных ее частях. Это мелкие или крупные травы с толстыми, часто ползучими корневищами. Листья в густых розетках, обычно с широкими, довольно мясистыми влагалищами.
Подсемейство тилландсиевые состоит из травянистых растений, ведущих большей частью эпифитный образ жизни. Корни часто функционируют только для прикрепления к субстрату и иногда полностью отсутствуют. Листья в розетках или в пучках, или же расположены вдоль стебля, цельнокрайные. Завязь верхняя или почти верхняя (за исключением рода гломеропиткерния - Glomeropitcairnia). Плод - септицидная коробочка. Семена с хохолковидными придатками. Важнейшие роды - тилландсия, фризея, гузмания.
Род тилландсия (рис. 155), насчитывающий более 400 видов, распространен по всему ареалу семейства. Он выходит относительно далеко за пределы тропической зоны, достигая тепло-умеренных областей Северной и Южной Америки. Габитуальные и биологические различия в роде тилландсия очень велики. За исключением "испанского мха", все виды тилландсии - прямостоячие растения. Одни из них имеют более или менее хорошо развитый стебель, другие же бесстебельные. Листья в розетках или в пучках или же распределены вдоль стебля. Как форма пластинки листьев, так и тип соцветия очень варьируют. Иногда все соцветие редуцировано до одного цветка. Чашелистики свободные или более или менее сросшиеся. Лепестки свободные. Наиболее известным представителем рода является "испанский мох" (рис. 155), который благодаря своим очень тонким, длинным (нередко достигающим 8 м), ветвистым побегам, свисающим с ветвей деревьев, действительно напоминает всем известный лишайник уснею. Своим обликом он сильно отличается от всех остальных тилландсий, особенно от розеточных "цистерновых" эпифитов. Как показал П. В. Томлинсон (1970), своеобразная жизненная форма тилландсии уснеевидной объясняется ее неотеническим происхождением. Сравнивая "испанский мох" с ювенильной стадией развития некоторых других тилландсий, Томлинсон пришел к выводу, что взрослые особи этого вида сходны с их проростками. Но, как заключает П. В. Томлинсон, "испанский мох" не просто перманентная ювенильная форма исходных "цистерновых" тилландсий, но ювенильная фаза, специализировавшаяся в совершенно своеобразном направлении. Эти признаки специализации включают такие изменения, как удлинение междоузлий, что вместе с утерей способности к геотропической реакции и отсутствием механической поддержки определило висячий образ жизни. Взрослые особи рано или поздно теряют корни, хотя у проростков корни обычно имеются. В отличие от более крупных тилландсий с соцветиями в виде метелки цветки "испанского мха" одиночные, расположенные на верхушках побегов.
К тилландсии стоит очень близко род фризея, включающий около 250 видов, распространенных от Центральной Америки и Вест-Индии до Аргентины и крайнего юга Бразилии. Это бесстебельные и обычно эпифитные многолетние травы с розетками жестких цельнокрайных листьев. Соцветия разного типа, большей частью колосья. Чашелистики свободные или почти свободные, лепестки свободные или сросшиеся в трубку, которая значительно короче чашечки. В отличие от тилландсии, у основания каждого лепестка имеются 2 чешуи. Завязь верхняя или почти верхняя. Семена с придатком. Ряд видов фризеи имеет декоративное значение. Одним из наиболее декоративных видов является фризея красивая (Vriesea speciosa).
С тилландсией имеет много общего род гузмания (рис. 158). В нем около 130 видов, распространенных в Южной и Центральной Америке и в Вест-Индии, а гузмания одноколосковая (Guzmania monostachya) достигает Южной Флориды.
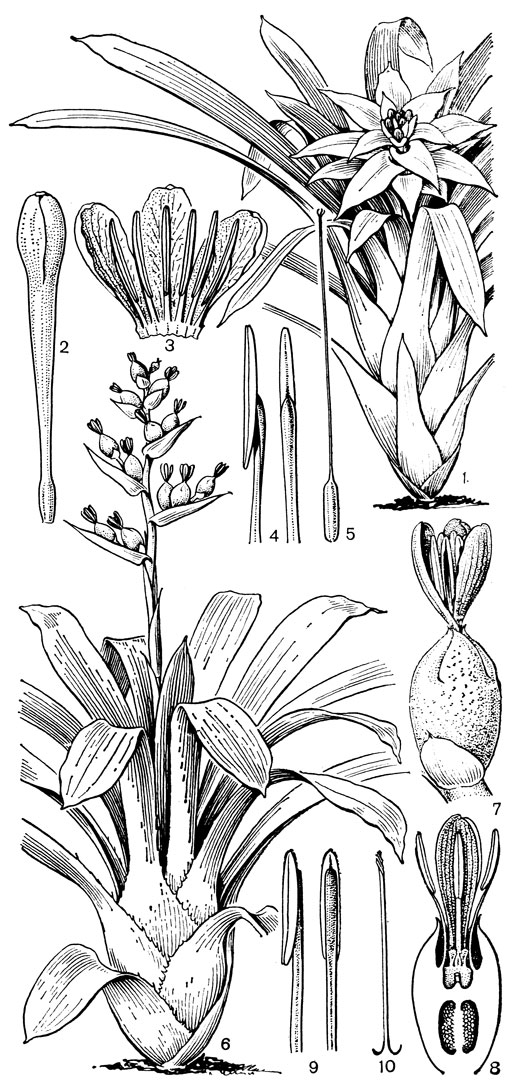
Рис. 158. Бромелиевые. Гузмания меньшая (Guzmania minor): 1 - общий вид; 2 - цветок; 3 - верхняя часть цветка в развернутом виде; 4 - тычинки; 5 - гинецей. Эхмея Вайльбаха (Aechmea weilbachii var. leodienchis): 6 - общий вид; 7 - цветок; 8 - продольный разрез цветка; 9 - тычинки; 10 - верхняя часть столбика с рыльцем
Подсемейство собственно бромелиевые - самое большое в семействе и по числу видов приблизительно равно двум первым, вместе взятым. В него входят травы, часто эпифитные, обычно бесстебельные. Корни в основном функционируют только для прикрепления к субстрату. Листья в розетках или пучках, обычно по краям колючепильчатые. Завязь нижняя. Плоды ягодовидные, но часто сухие. Семена, как правило, без придатков, редко с придатками. Главные роды - неорегелия (Neoregelia), нидулариум (Nidnlarium), бромелия, бильбергия (рис. 159), эхмея и ананас. Центром разнообразия подсемейства бромелиевых является Восточная Бразилия.
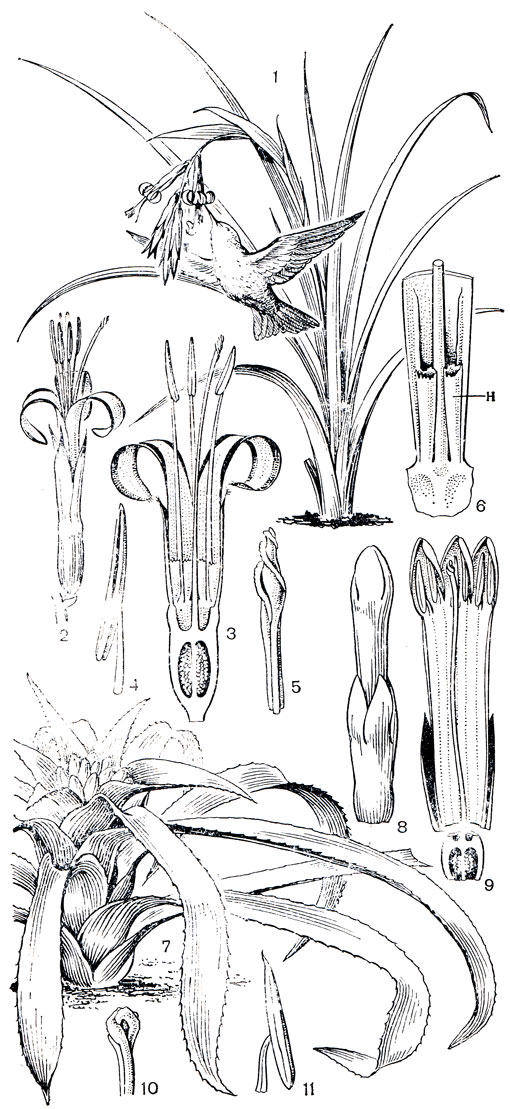
Рис. 159. Бромелиевые. Бильбергия поникшая (Billbergia nutans): 1 - общий вид растения; 2 - цветок; 3 - продольный разрез цветка; 4 - тычинка; 5 - верхняя часть гинецея; 6 - нижняя часть лепестка и тычинки с нектарниками. Нидулариум Шереметьева (Nidularium scheremetiewii): 7 - общий вид растения; 8 - цветок; 9 - продольный разрез цветка; 10 - верхняя часть гинецея; 11 - часть тычинки с пыльником
Род неорегелия содержит более 50 видов, распространенных в тропиках Южной Америки. Листья в густых розетках, обычно колючепильчатые, а цветонос короткий, полностью заключенный в розетку. Среди видов неорегелии имеется ряд очень декоративных растений. К неорегелии очень близок род нидулариум (23 вида).
Род бромелия (около 50 видов) представлен наземными травами с розеткой жестких листьев и цветками, собранными в головки или метелки.
К роду эхмея относится более 170 видов эпифитных трав, распространенных в Вест-Индии и Южной Америке. Листья у них в розетках или пучках, жесткие. Некоторые виды эхмеи характеризуются бросающимися в глаза крупными, ярко окрашенными прицветниками, что придает им особую декоративность. Одним из наиболее декоративных видов является эхмея королевы Марии (Aechmea mariae-reginae) из Коста-Рики.
К бромелии близок род ананас (рис. 160), заключающий 8-9 видов, распространенных в Южной Америке на юг до Аргентины и Парагвая. Это наземные травы с коротким стеблем, розеткой базальных суккулентных и по краям колючезубчатых листьев. После оплодотворения соцветие ананаса превращается в компактное соплодие, состоящее из сросшихся между собой плодов, прицветников и осей соцветия. Почти у всех видов ананаса, за исключением ананаса уродливого (Ananas monstrosus), главная ось соплодия продолжает расти и образует на верхушке соплодия розетку листьев (корону). У основания этой короны часто развиваются отпрыски, способствующие вегетативному размножению.

Рис. 160. Ананас посевной (Ananas sativus): 1 - общий вид растения с соцветием; 2 - цветок; 3 - тычинка с лепестком (видны нектарники); 4 - гинецей; 5 - соплодие
Культурный ананас относится к виду ананас крупнохохолковый (A. comosus), дико произрастающему в Центральной Бразилии. Сочные и душистые золотисто-желтые соплодия культурных сортов ананаса отличаются высокими вкусовыми качествами. Большинство культурных сортов характеризуется самонесовместимостью и обычно не образует семян, но при скрещивании с некоторыми другими сортами семена нормально развиваются.
Родиной культурного ананаса является, вероятно, Бразилия, откуда еще в доисторические времена его возделывание распространилось как к югу, так и к северу в Центральную Америку и в Вест-Индию. Уже сразу после открытия Америки культура ананаса проникла во многие страны мира, первоначально главным образом благодаря португальцам. Они интродуцировали ананас на остров Святой Елены вскоре после его открытия в 1502 г. Вслед за этим португальцы перенесли культуру ананаса в разные части Африки и на Мадагаскар и около 1550 г.- в Индию. До конца XVI в. культура ананаса успела распространиться в большинство тропических стран, включая некоторые острова Тихого океана. В Европу ананас был доставлен около 1650 г., где с тех пор его выращивают в оранжереях. В 1769 г. капитан Кук интродуцировал ананас на острове Таити. Относительно быстрое распространение культуры ананаса объясняется не только исключительно высокими вкусовыми качествами его соплодий, но и легкостью его размножения посредством быстро укореняющихся крон на верхушках соплодий. В настоящее время ананас широко культивируется во многих тропических и субтропических странах обоих полушарий. Особенно велики плантации ананаса на Гавайских островах, поставляющих около 30% всей мировой продукции. Большие площади ананас занимает также в Бразилии, Мексике, на Филиппинах, Кубе и на острове Тайвань. Довольно значительны плантации ананаса также на Азорских островах. Ананас употребляют в пищу как в сыром, так и в консервированном виде (в собственном соку). Из него получают очень приятный на вкус и ароматный сок и готовят также варенье. Ананас является хорошим источником витаминов А и В. Его разводят также для получения из листьев прядильного волокна, главным образом на Филиппинах и Тайване. В соплодиях близкого к ананасу крупнохохолковому бразильского вида ананаса прицветникового (А. brасteatus) плоды хотя и с семенами, но съедобны.
Кроме ананаса, в семействе бромелиевых имеются и другие полезные для человека растения, однако они не имеют столь большого значения, как ананас. Плоды некоторых видов бромелии употребляют в пищу местные жители, в Колумбии индейцы едят сердцевину и молодые листья пуйи, в Боливии и Аргентине используют как овощи молодые побеги некоторых тилландсий, а в Пуэрто-Рико - молодые цветоносы бромелии пингвин (Bromelia pinguin). Кроме того, из плодов этого последнего вида делают прохладительные напитки. Пуйю крючковатую (Puya hamata) из парамоса Южного Эквадора используют для приготовления сладкого напитка ("jugo de agnaronge"). Напиток изготовляют из мягких листовых оснований и верхушек стеблей.
Некоторые бромелиевые, например уже упомянутая выше бромелия пингвин, являются лекарственными растениями. Наибольшее медицинское значение имеет протеолитический фермент бромелаин, добываемый из плодов ананаса и некоторых других бромелиевых. Высокое содержание этого протеолитического фермента в плодах объясняется тем, что оно возникло как химическая защита против личинок насекомых, которые перевариваются прежде чем созреют семена (Дж. Л. Коллинз, 1958; Д. Бензинг, 1980). При консервировании ананаса бромелаин разрушается.
Из разных видов бромелиевых добывают волокна, используемые местными жителями для изготовления одежды и снастей, особенно волокна из ананаса на Филиппинах, эхмеи Магдалины (Aechmea magdalenae) в Мексике и Ко-лумбии и неоглазовии пестрой (Neoglazowia variegata) в Бразилии. Волокна ананаса исследовали также для производства бумаги. "Испанский мох", или тилландсию уснеевидную (Tillandsia usneoides) использовали как заменитель конских волос в обойном деле. Шипы по краям листьев пуйи чилийской (Рnуа chilensis) местные жители используют в качестве рыболовных крючков.
Очень велико чисто эстетическое значение бромелиевых. Многие из них разводят в теплых странах в открытом грунте, а в странах с умеренным или холодным климатом в оранжереях и комнатах как декоративные растения. У бромелиевых красивы даже и сами листья, как, например, пестролистные формы ананаса, листья некоторых видов бильбергии, розетки диккии, нидулариума и арегелии (Aregelia).
У ряда родов красивы соцветия, например у питкернии, бильбергии, эхмеи и фризеи. Культивируют также бромелии и неорегелию.
Но бромелиевые приносят не только пользу человеку. В некоторых сухих тропиках вода в цистернах местных эпифитных и наземных бромелиевых служит местом размножения малярийных анофелесовых комаров, что является определенной помехой в борьбе с малярией.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'