Порядок лилейные (Liliales)
Семейство мелантиевые (Melanthiaceae) (А. Л. Тахтаджян)
Это относительно самое примитивное семейство в порядке лилейных. Оно объединяет 39 родов и около 350 видов, большинство которых встречается во внетропических областях северного полушария и в Африке (главным образом в Южной Африке); лишь немногие представители мелантиевых произрастают в тропической Азии, Австралии и Южной Америке.
Мелантиевые - типичные геофиты, обычно с хорошо развитыми запасающими подземными органами - корневищами, луковицами или клубнелуковицами. Листья у них расположены или по всему стеблю или же сосредоточены только у его основания (так называемые приземные или базальные листья). Устьица мелантиевых не имеют побочных клеток (аномоцитные), а сосуды обычно встречаются только в корнях и характеризуются лестничной перфорацией.
Цветки собраны в различного рода соцветия, реже одиночные, обычно обоеполые и лишь редко однополые. Сегменты околоцветника свободные или более или менее сросшиеся. Тычинок обычно 6, реже 3, редко 9; пыльники прикреплены к нити основанием или спинкой, большей частью вскрываются продольной щелью, реже верхушечной порой, экстрорзные или интрорзные. Пыльцевые зерна у большинства мелантиевых однобороздные, иногда двубороздные, но у некоторых наиболее подвинутых родов они 2-4-поровые. Гинецей состоит из 3 плодолистиков, которые у наиболее примитивных форм свободны до основания или почти до основания, но у большинства родов более или менее сросшиеся; завязь верхняя, с многочисленными или несколькими семязачатками. Семязачатки анатропные и, как правило, битегмальные. Плод представляет собой септицидную или локулицидную коробочку, но у некоторых примитивных родов он является многолистовкой или переходным типом от многолистовки к коробочке.
Среди ботаников имеются большие разногласия как в отношении таксономической самостоятельности семейства мелантиевые, так и его объема. Это семейство впервые установил еще в 1802 г. немецкий ботаник Август Бач. Через три года А. П. де Кандоллем (1805) было установлено семейство безвременниковые (Colcliicaceae), которое одними авторами рассматривалось как самостоятельное, а другими объединялось с мелантиевыми. В прошлом веке самостоятельность семейства мелантиевые признавали многие выдающиеся систематики, в том числе английские ботаники Роберт Браун (1810) и Джон Линдли (1836, 1846), русский ботаник П. Ф. Горянинов (1834), австрийский ботаник Стефан Эндлихер (1836, 1841), немецкий ботаник Адальберт Шницлейн (1843-1846), датский ботаник Йоханнес Варминг (1879) и швейцарский ботаник Пьер Эдмон Буасье (1884) (Горянинов и последние два ботаника - под названием Colchicaceae). Но уже во второй половине прошлого века в системах А. Эйхлера (1875), Дж. Бентама и Дж. Хукера (1883), А. Энглера (1888) и их последователей мелантиевые включаются в качестве одного из подсемейств в обширное и в принятом ими объеме очень разнородное семейство лилейные (Liliaceae), установленное еще Антуаном Лораном Жюсьё (1789). Таково положение мелантиевых и таков или почти таков объем семейства лилейных в системах Веттштейна, Бесси и Халлира, почти во всех современных руководствах по систематике и во всех определителях и "флорах". Но уже в первой четверти нашего столетия делаются попытки подразделения этой довольно искусственной группы "лилейных" на ряд более естественных семейств. Наиболее решительный шаг в этом направлении был сделан известным голландским ботаником Яном Паулусом Лотси (1911) в третьем томе его "Лекций о ботанической родословной". Он восстанавливает целый ряд хороших, таксономически ясно очерченных семейств, установленных ранее ботаниками, и прибавляет к ним новые семейства. Но в то же время Лотси чрезмерно раздробил лилейные, выделив из них целый ряд сомнительных семейств. Именно, вероятно, поэтому его система не получила признания. И лишь значительно позднее, начиная с работ Дж. Хатчинсона (1934) и А. Л. Тахтаджяна (1954, 1959) и особенно немецкого ботаника X. Хубера (1969), начинается широкая ревизия традиционной энглеровской системы. Последними попытками в этом направлении являются системы датского ботаника Р. Дальгрена (1975, 1980) и А. Л. Тахтаджяна (1980). Следуя Хуберу, Дальгрен подразделяет мелантиевые на три самостоятельных семейства - безвременниковые (Colchicaceae), трициртисовые (Tricyrtidaceae) и собственно мелантиевые (Melanthiaceae), причем безвременниковые ставит в самом начале порядка лилейных (в принятом им объеме), в то время как мелантиевые располагает в конце. В системе Тахтаджяна (1980) все эти три группы вместе с родом петросавия (Petrosavia) рассматриваются как одно естественное семейство. Современные исследования лилейных, основанные на широком сравнительно-морфологическом изучении (включая цитологию, палинологию и эмбриологию) и данных химии и систематической серологии, показывают, что наиболее близок к истине тот объем семейства мелантиевых (безвременниковые), который был принят Линдли, Эндлихером и другими ботаниками прошлого века.
Семейство мелантиевые в принятом здесь объеме состоит из трех подсемейств - петросавиевые (Petrosavioideae), собственно мелантиевые (Melanthioideae) и безвременниковые (Colchicoideae). Мелантиевые (и, вероятно, также петросавиевые) характеризуются гелобиальным эндоспермом, в то время как у безвременниковых эндосперм всегда нуклеарный. Имеются также различия в строении цветка, в том числе гинецея. Учитывая значение мелантиевых для построения филогенетической системы порядка лилейных и для понимания путей его эволюции, остановимся на них несколько подробнее.
Подсемейство петросавиевые состоит из одной трибы петросавиевые (Petrosavieae) и одного рода петросавия (Petrosavia). Петросавия настолько своеобразна и во многом так сильно отличается от остальных представителей семейства и даже порядка, что иногда, например в системах Дж. Хатчинсона (1934) и А. Кронквиста (1968, 1981), выделяется в отдельное семейство петросавиевые (Petrosaviaceae). Это небольшой род, состоящий всего из 3 видов, встречающихся в Японии (очень редко в лесах на острове Хонсю), Южном Китае, на острове Тайвань, на полуострове Малакка и на острове Калимантан. Виды петросавии произрастают в тенистых местах на влажной лесной почве. В отличие от всех остальных представителей семейства петросавии - сапрофиты, небольшие бледно-желтые травы с тонким корневищем и тонким стеблем, несущим лишенные хлорофилла редуцированные чешуевидные листья. Мелкие обоеполые актиноморфные цветки петросавии собраны в верхушечные кисти, иногда щитковидные. Околоцветник из 5 у основания сросшихся сегментов; внутренние сегменты крупнее наружных и у некоторых видов снабжены у основания нектарной железкой. Таким образом, нектарники у петросавии тепальные (от лат. tepalum - сегмент околоцветника). Тычинок 6, прикрепленных к сегментам околоцветника; пыльники прикреплены спинкой, интрорзные. Пыльцевые зерна как у большинства мелантиевых, однобороздные. Гинецей из 3 плодолистиков, коротко сросшихся у основания между собой и с сегментами околоцветника; столбик короткий, с более или менее головчатым рыльцем (у калимантанской петросавии звездчатой - P. stellaris, рис. 24, 7-12,- рыльце низбегающее) и многочисленными семязачатками в каждом гнезде. Плод - многолистовка, отдельные плодики которой раскрываются на верхней стороне. Семена многочисленные, бороздчатые, с эндоспермом. Уже из этого описания следует, что петросавия представляет собой очень своеобразную боковую ветвь развития примитивных мелантиевых, характеризующаяся очень контрастным сочетанием ряда, несомненно, примитивных признаков с признаками высокой специализации.
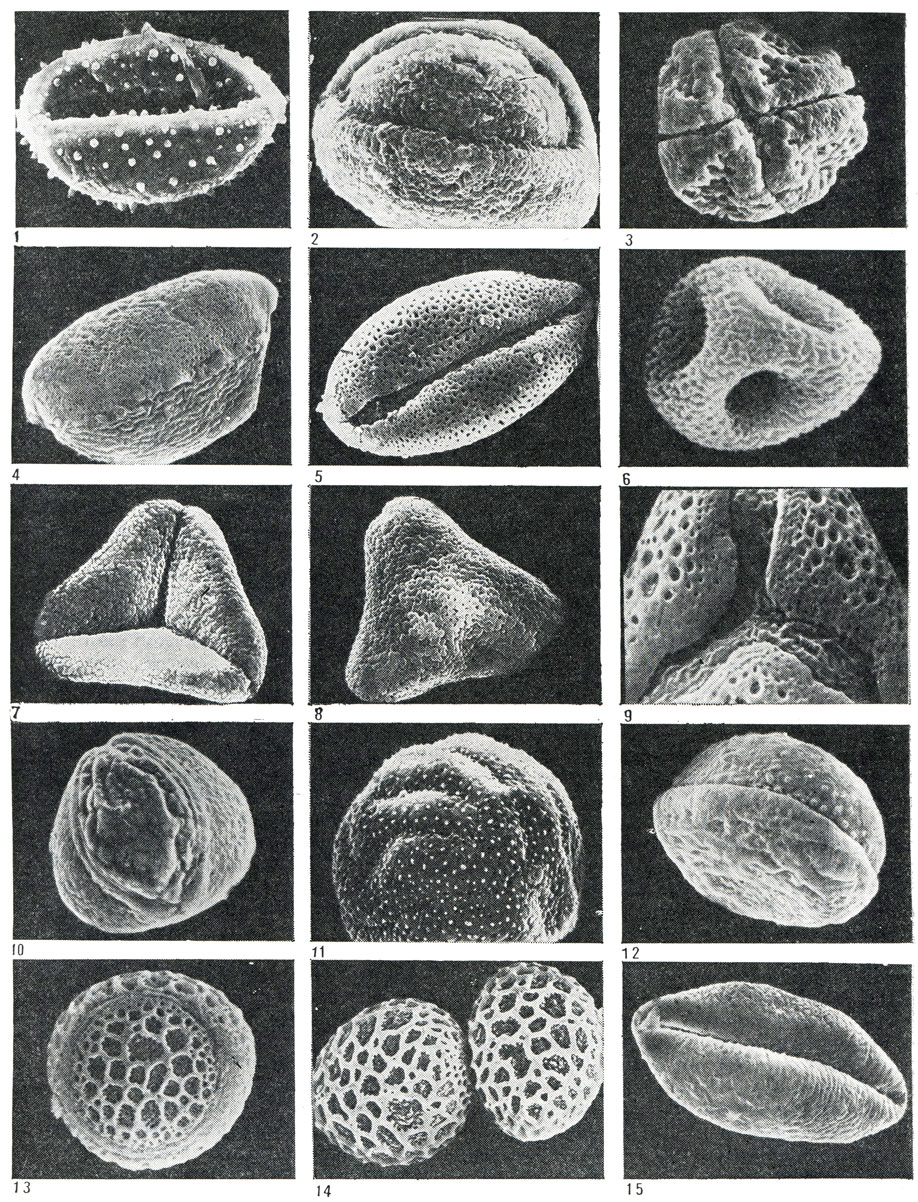
Рис. 22. Пыльцевые зерна некоторых представителей порядков лилейных и смилаксовых: 1 - гелониас пузырчатый (Helonias bullata, увел. 2000); 2 - пролеска двулистная (Scilla bifolia, увел. 2000); 3 - фуркрея Бедингхауза (Furcraea bedinghausii, увел. 620); 4 - алоэ переменчивое (Aloe commutata, увел. 1500); 5 - схенокаулок тонколистный (Schoenocaulon tenuifolia, увел. 2000); 6 - хионографис японский (Ghionographis japonica, увел. 4000); 7, 8 - гейтоноплезиум полузонтичный (Geitonoplesium cymosum, увел. 2200); 9 - формиум прочный, или новозеландский леи (Phormium tenax, увел. 3700); 10 - асфоделина либурнская (Asphodeline liburnica, увел. 780); 11 - шафран золотистоцветковый (Crocus chrysanthus, увел. 1100); - сансевьера цилиндрическая (Sansevieria cylindrica, увел. 1300); 13, 14 - агава оттянутая (Agave attenuata, увел, 1300); 15 - такка гребенчатая (Тасса cristata, увел. 1200)
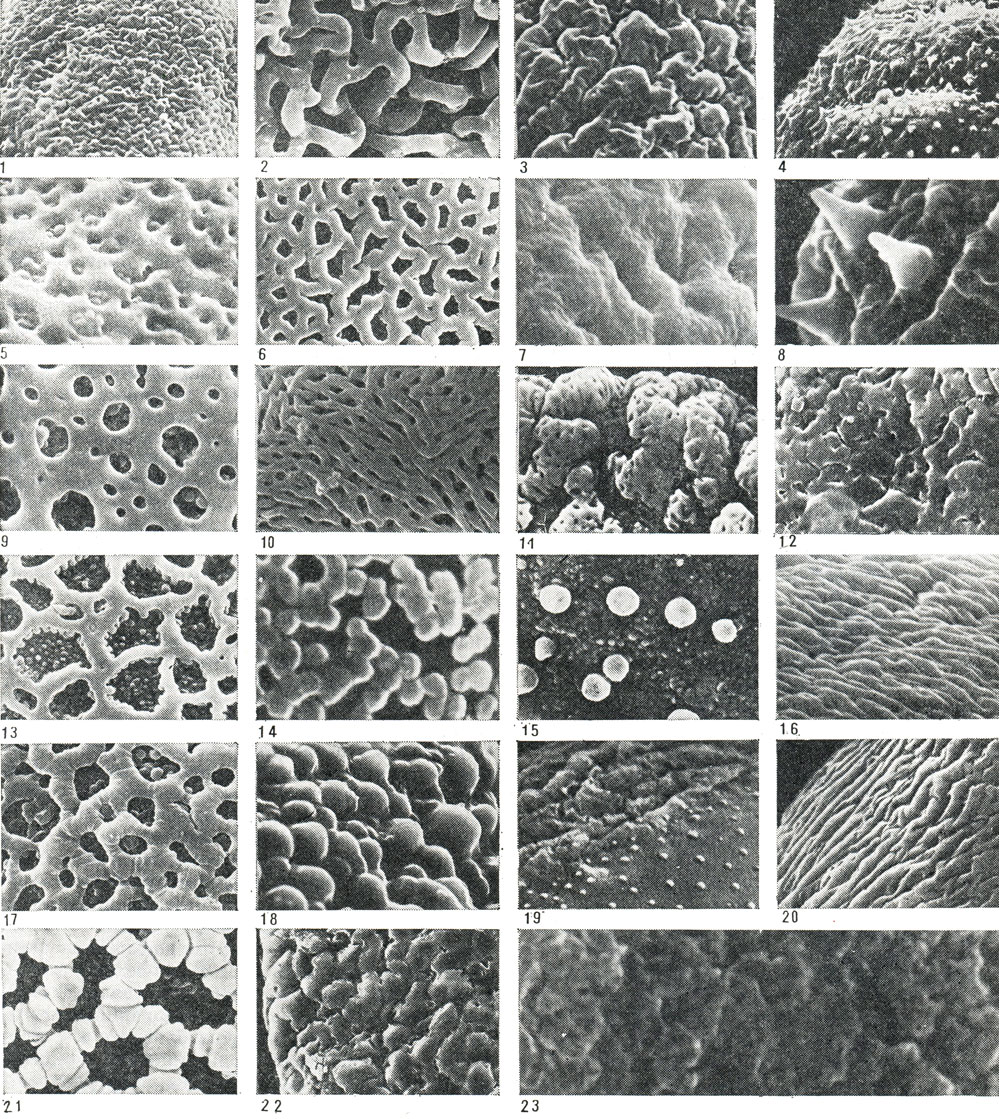
Рис. 23. Типы скульптуры пыльцевых зерен некоторых представителей порядков лилейных и смилаксовых: 1 - хлорофитум частухолистный (Chlorophytum alismaefolium, увел. 5000); 2 - гесперокаллис волнистолистный (Hesperocallis undulata, увел. 2700); 3 - бульбинелла хвостатая (Bulbinella caudata, увел. 5000); 4 - шафран золотистоцветковый (Crocus chrysanthus, увел. 5000); 5 - цезия полосатая (Caesia vittata, увел. 5000); 6 - зигаденус колорадский (Zigadenus coloradensis, увел. 5000); 7 - сансевьера цилиндрическая (Sansevieria cylindrica, увел. 5000); 8 - лапажерия розовая (Lapageria rosea, увел. 5000); 9 - леонтохир Овалье (Leontochir ovallei, увел. 5000); 10 - глориоза великолепная (Gloriosa superba, увел. 5000); 11 - сколиопус Бегелова (Scoliopus begelowii, увел. 5000); 12 - офиопогон пестролистный (Ophiopogon poecilophyllus, увел. 5000); 13 - крумия немногоцветковая (Croomia pauciflora, увел. 5000); 14 - хамелириум желтый (Chamaelirium luteum, увел. 5000); 15 - гелониас пузырчатый (Helonias bullata, увел. 5000); 16 - альстрёмерия открытоцветковая (Alstroemeria apertiflora, увел. 5000); 17 - красоднев, или гемерокаллис Миддендорфа (Hemerocallis middendorfii, увел. 5000); 18 - хоста ланцетолистная (Hosta lancifolia, увел. 5000); 19 - кринум повислый (Crinum flaccidum, увел. 2200); 20 - такка Шантрье (Тасса chanttrieri, увел. 5000); 21 - лилия леопардовая (Lilium pardalinum, увел. 5000); 22 - анигозантос желтоватый (Anigozanthos flavidus, увел. 2700); 23 - эвстрефус широколистный (Eustrephus latifolius, увел. 5000)
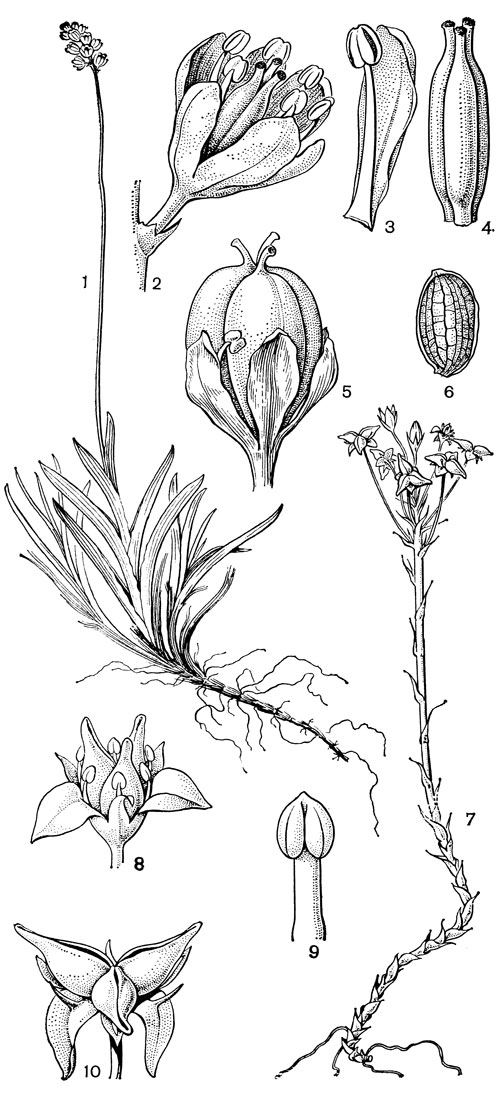
Рис. 24. Мелантиевые. Тофилдия крохотная (Tofieldia pusilla): 1 - общий вид; 2 - цветок; 3 - тычинка и сегмент околоцветника; 4 - гинецей; 5 - плод; 6 - семя. Петросавия звездчатая (Petrosavia stellaris): 7 - общий вид; 8 - цветок; 9 - тычинка; 10 - плод
Подсемейство мелантиевые - самое большое в семействе. В него входит 5 триб: тофилдиевые (Tofieldieae), нартециевые (Narthecieae), гелониевые (Helonieae), хионографовые (Chionographieae) и мелантиевые (Melanthieae). Самой примитивной является небольшая триба тофилдиевых, заключающая всего 2 очень близких рода - тофилдию (Tofieldia, рис. 24, 1-6) и плеею (Pleea). Именно с нее мы и начнем наше знакомство с подсемейством мелантиевых.
Род тофилдия представляет для нас особый интерес, так как в некоторых отношениях он занимает едва ли не самое низкое положение в эволюционной системе семейства, а следовательно, и всего порядка. Известно около 25 видов тофилдии, распространенных во внетропических областях северного полушария (включая Арктику), а также в Венесуэле, Гайане и Андах. Виды тофилдии - небольшие растения с тонкими корневищами, простым или ветвистым прямостоячим симподиальным стеблем и линейными, большей частью базальными листьями, сложенными вдоль середины. Мелкие, белые, зеленовато-белые, желтоватые или коричневато-красные цветки тофилдии образуют короткую, густую, верхушечную кисть или колос. Тычинки в числе 6 прикреплены к основаниям сегментов околоцветника. У очень близкого к тофилдии и в последние годы иногда объединяемого с ней монотипного рода плеея, распространенного на юго-востоке США, тычинок 9 (к сегментам наружного круга околоцветника прикреплено по 2 тычинки). Пыльники тофилдиевых прикреплены к нити основаниями и интрорзные. Интересно, что пыльцевые зерна обоих родов обычно двубороздные, что по сравнению с однобороздными зернами является признаком некоторой подвинутости. Но самой замечательной особенностью этих двух родов является строение их гинецея. Дело в том, что плодолистики у обоих этих родов хотя и прилегают плотно друг к другу, но не срастаются, а столбики у них совершенно свободные. Кроме того, каждый плодолистик сидит на отдельной ножке, и таким образом весь гинецей как бы возвышается на трех ножках. Наличие у плодолистика ножки - признак очень примитивный (такие плодолистики встречаются только у примитивных представителей двудольных, например у дегенерии). Наконец, что не менее интересно, плодолистики тофилдиевых открыты, т. е. еще не срослись краями, что мы опять-таки встречаем только у наиболее примитивных цветковых растений. На поперечном разрезе гинецей тофилдиевых производит впечатление паракарпного гинецея, но это еще самая начальная стадия возникновения паракарпного гинецея (настоящая паракарпия предполагает гистологическое срастание соседних плодолистиков). Но наряду с этими архаическими особенностями в морфологии плодолистиков тофилдиевых есть и такая прогрессивная черта, как верхушечное расположение рылец. Вдоль обычно несколько загнутых назад краев плодолистиков сидят многочисленные семязачатки.
У тофилдии и плееи наблюдаются самые начальные стадии возникновения так называемых септальных нектарников, т. е. нектарников, расположенных на перегородках завязи (от лат. septum - перегородка). Эти своеобразные нектарники характерны для некоторых групп однодольных, в том числе для большинства представителей порядка лилейных. Они встречаются при неполном боковом срастании плодолистиков, когда несросшиеся участки становятся секреторными и превращаются в нектарники. Но у тофилдиевых, так же как и у частухи (Alisma), о которой речь была раньше, мы наблюдаем лишь самое начало формирования септальных нектарников. Уже на самых ранних стадиях развития плодолистиков тофилдиевых их боковые (а первоначально также внутренние) поверхности бывают покрыты секреторными сосочками. Любопытно, что сосочки первоначально закладываются не на самих плодолистиках, а под ними, т. е. фактически в верхней части цветоножки, что совершенно не наблюдается у других однодольных. Эти сосочки как бы скрепляют боковые стенки соседних плодолистиков и в то же время выделяют нектар в продольные межплодолистиковые ложбинки. Так как септы (перегородки) у тофилдиевых отсутствуют, то в сущности эти сосочки нельзя еще считать настоящими септальными нектарниками. Выделяемый ими нектар стекает вниз и собирается в виде крошечных капелек между основанием завязи и тычиночными нитями. Он привлекает пчел, мух и маленьких жуков (в том числе видов антобиума - Anthobium - из семейства стафилинид), которые и являются опылителями тофилдиевых. Для тофилдиевых характерна ясно выраженная протогиния. При отсутствии перекрестного опыления иногда возможно самоопыление.
Плод тофилдиевых представляет собой нечто среднее между многолистовкой и септицидной коробочкой, но стоит все же ближе к многолистовке. Так как перегородки в гинецее тофилдиевых отсутствуют, то здесь еще нет настоящего септицидного раскрывания, как у настоящих коробочек, а скорее лишь ложносептицидное отделение друг от друга зрелых плодиков.
Эллипсоидальные семена тофилдиевых интересны тем, что на обоих концах они бывают обычно снабжены более или менее длинными (иногда, как у тофилдии клейкой - Tofieldia glutinosa, очень длинными) придатками. Эти придатки имеют, вероятно, определенное значение при распространении семян.
С трибой тофилдиевых тесно связана триба нартециевых (Narthecieae), состоящая из 5 родов. Главным отличием нартециевых от предыдущей трибы является синкарпный гинецей и отсутствие нектарников. Пыльцевые зерна у них однобороздные, что характерно и для всех остальных триб подсемейства собственно мелантиевых. Вероятно, самым примитивным представителем нартециевых является монотипный японский род японолирион (Japonolirion, рис. 25), произрастающий на альпийских лугах островов Хоккайдо и Хонсю. Корневище у него тонкое, короткое, а листья базальные, двурядные, линейные, по краям сильно шероховатые. Цветонос с немногими мелкими чешуевидными листьями, заканчивается кистью. Цветки мелкие, обоеполые, актиноморфные, зеленоватые, на коротких цветоножках, без прицветничков, но с пленчатыми прицветниками. Сегментов околоцветников 6, пленчатых, широколанцетных. Тычинок 6, прикрепленных к основаниям сегментов околоцветника; пыльники прикреплены к нити основаниями, интрорзные, с параллельными гнездами. Гинецей из 3 сросшихся плодолистиков, столбики свободные, отогнутые наружу, с рыльцевой поверхностью на внутренней стороне (примитивный признак). Плод - септицидная коробочка. Японолирион имеет много общего с тофилдией, но отличается синкарпным гинецеем, низбегающими рыльцами и числом хромосом (у тофилдии 2n = 30или реже 60, а у японолириона 2n = 26) и их морфологией. Хотя Дж. Хатчинсон (1934) включает, правда со знаком вопроса, японолирион в тофилдию, в действительности эти два рода настолько отличимы, что их нельзя даже относить к одной трибе. По некоторым своим особенностям, в частности по кариотипу, японолирион стоит гораздо ближе к нартециевым.
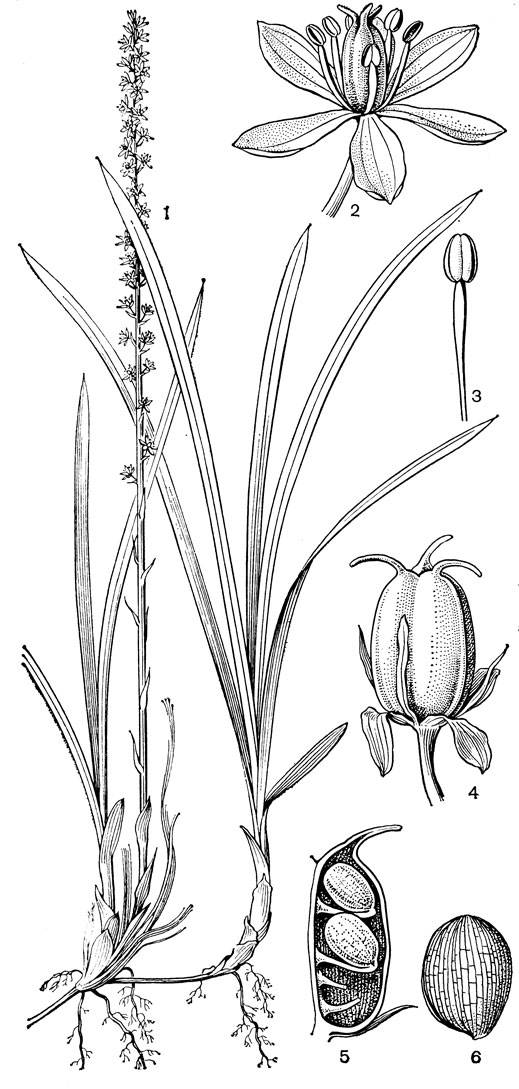
Рис. 25. Мелантиевые. Японолирион осенский (Japonolirion osense): 1 - общий вид; 2 - цветок; 3 - тычинка; 4 - плод; 5 - продольный разрез плода; 6 - семя
Относительно примитивен также род ксерофиллум (Xerophyllum, рис. 26, 1-4), состоящий из 2 видов, распространенных в умеренных и теплых областях Северной Америки на сухих склонах и в сухих сосновых лесах. От толстого клубневидного деревянистого корневища отходит пучок длинных (до 50-80 см), узколинейных, жестких, заостренных, по краям шероховатых, ксероморфных листьев. Стеблевые листья более редкие и значительно более короткие. Цветки в густых кистях, беловатые или кремовые. Гинецей паракарпный, в поперечном разрезе трехлопастный, со свободными столбиками с низбегающими рыльцами. У обоих видов эпидерма плодолистика, включая внутреннюю поверхность их сросшихся краев, густо покрыта сосочковидными клетками. В каждом плодолистике 2 или 4 семязачатка. Коробочка локулицидная. Семена продолговатые, трехгранные, без придатков. Соматическое число хромосом 2n = 30, т. е. такое же, как у тофилдии, причем даже размеры хромосом у них одинаковы.

Рис. 26. Мелантиевые. Ксерофиллум прочный (Xerophyllum tenax): l - общий вид; 2 - соцветие; 3 - цветок; 4 - плод. Алетрис желто-зеленый (Aletris luteo-viridis): 5 - общий вид; 6 - цветок; 7 - тычинка с сегментом околоцветника; 8 тычинка; 9 - гинецей; 10 - семя
Наиболее известным представителем трибы нартециевых является нартециум (Narthecium) - небольшой род, насчитывающий всего 7 видов. Ареал его очень разорван. Три вида встречаются в Европе, в том числе нартециум костоломный (N. ossifragum, рис. 27), распространенный в Северной, Западной и Средней Европе (на востоке до Юго-Восточной Швеции). На Кавказе встречается лишь один вид - нартециум Балансы (N. balansae), произрастающий в альпийском поясе Западного Закавказья. Еще более оторван ареал нартециума азиатского (N. asiaticum), эндемичного для Японии (острова Хоккайдо и Хонсю). Наконец, два остальных вида нартециума распространены в Северной Америке, где нартециум калифорнийский (N. californicum) встречается от Юго-Западного Орегона до Центральной Калифорнии, а нартециум американский (N. americanum) - на юго-востоке США. Такой разорванный ареал типичен для многих родов, которые в третичное время были широко распространены в умеренной зоне северного полушария. Виды нартециума растут на влажных горных лугах, на болотах, вдоль ручьев и на влажных местах. От ползучего ветвистого корневища нартециума отходят расположенные двурядно линейные объемлющие приземные листья. Стеблевые листья короткие, немногочисленные. Цветки мелкие, желтые или желтовато-зеленые, снабженные линейными прицветничками, в кистях, в нижней их части иногда ветвящихся. Тычинки несколько короче околоцветника, с густо беловойлочно опушенными нитями. Гинецей синкарпный, постепенно вытянут в короткий, колонновпдныи столбик, заканчивающийся маленьким слегка трехлопастным рыльцем. Продолговатая многосемянная коробочка нартециума раскрывается локулицидно. Веретеновидные семена снабжены у обоих концов длинными нитевидными придатками. Наличие придатков на семенах сближает нартециум с тофилдией, а соматическое число хромосом (26 или 52) - с японолирионом. Пыльники, созревающие одновременно с рыльцем, превышают его примерно на 3 мм. Однако они находятся на таком расстоянии от рыльца, что спонтанное самоопыление сильно затруднено. Пахучие цветки нартециума посещают шмели, медоносные пчелы, одиночные пчелы из рода галиктус (Halictus) и мухи. Некоторые виды нартециума используют в декоративном садоводстве.
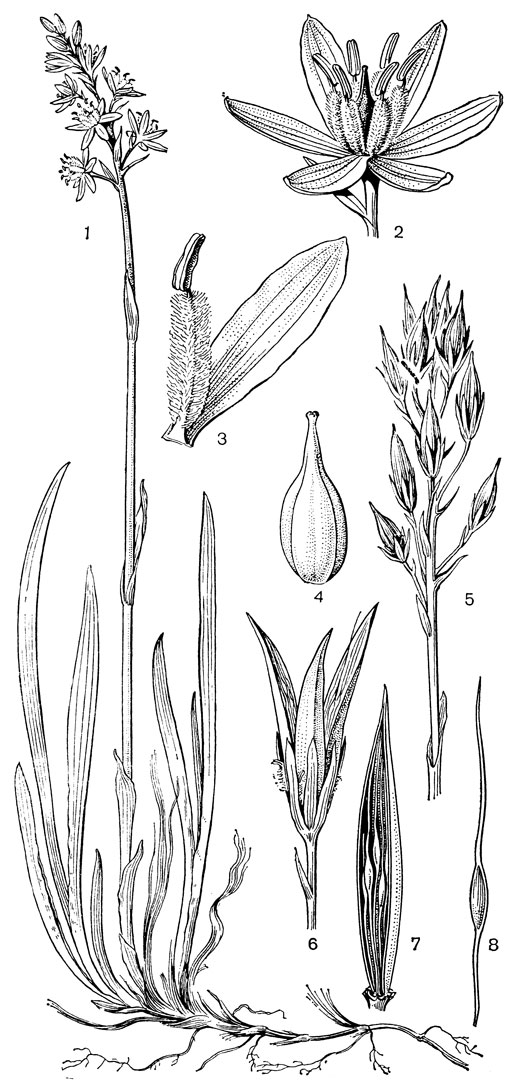
Рис. 27. Нартециум костоломный (Narthecium ossifragum): 1 - общий вид; 2 - цветок; 3 - тычинка и сегмент околоцветника; 4 - гинецей; 5 - часть стебля с плодами; 6 - плод; 7 - часть плода с семенами; 8 - семя
К нартециуму близок монотипный южноамериканский род нитнерия (Nietneria), встречающийся в Венесуэле и Гайане. От нартециума он отличается сегментами околоцветника, сросшимися у основания в трубку, которая, в свою очередь, приросла к основанию завязи (завязь, таким образом, нижняя) и тычинками, приросшими к трубке околоцветника. Рыльце маленькое, головчатое.
По числу и морфологии хромосом, как и по некоторым своим особенностям, к нартециуму близок род алетрис (Aletris, рис. 26, 5-10), насчитывающий около 16 видов. Алетрисы распространены в Гималаях, в Ассаме (Индия), Тибете, континентальном Китае, на острове Тайвань, на островах Рюкю, в Японии, в Западной Малезии и в Северной Америке. Корневище алетрисов короткое, толстое, а приземные листья линейные или ланцетные. Цветки собраны в колосовидные кисти, белые, желтые или желто-зеленые на очень коротких цветоножках или почти сидячие, с мелкими прицветниками и прицветничками. Сегменты околоцветника более или менее сросшиеся в колокольчатую или цилиндрическую трубку, редко почти свободные (японский алетрис желто-зеленый - А. luteo-viridis, рис. 26, 5-10, иногда выделяемый в отдельный род метанартециум - Metanarthecium). Тычинки прикреплены к околоцветнику, обычно с короткими нитями. Завязь чаще полунижняя, редко лишь у самого основания приросшая к околоцветнику (алетрис желто-зеленый), с колончатым столбиком, заканчивающимся 3-лопастным рыльцем. Семязачатки многочисленные. Коробочка локулицидная. Семена мелкие, в отличие от семян нартециума лишены придатков. Алетрисы чаще всего растут на горных лугах. Корневища их иногда употребляют для лечебных целей, а некоторые виды используют в декоративном садоводстве.
С трибой тофилдиевых связана также триба гелониевых (Helonieae). Все они корневищные травы с приземными листьями, маленькими обоеполыми или редко однополыми цветками, собранными в кисти или в колосья, и с септицидными или чаще локулицидными коробочками. Одним из наиболее примитивных родов этой трибы является монотипный род гелониас (Helonias), произрастающий на болотах приатлантических областей Северной Америки. Для гелониаса характерны очень короткое клубневидное корневище, длинные приземные листья, высокий, полый стебель, заканчивающийся густой колосовидной кистью, пахучие, розовые цветки с узкопродолговатыми сегментами, длинные тычиночные нити, синие экстрорзные пыльники, свободные столбики, локулицидная коробочка и линейные семена, у обоих концов снабженные белыми придатками. Соматическое число хромосом 34.
По многим своим особенностям, в том числе по числу и морфологии хромосом, к гелониасу очень близок небольшой восточноазиатский род гелониопсис (Heloniopsis, рис. 28). К нему относятся 2 вида, встречающиеся на Сахалине, в Японии, на полуострове Корея и на острове Тайвань. В отличие от гелониаса столбики гелониопсиса сросшиеся, красные, с головчатым пурпурным рыльцем. Между каждой тычинкой и сегментом околоцветника находится глубокий кармашек, в глубине которого у основания сегмента расположены нектарники. Семена, как и у гелониаса, линейные, у обоих концов с придатками. Гелониопсисы растут в кустарниках и на лугах, иногда достигая высокогорного пояса. Они отличаются высокими декоративными качествами и нередко культивируются на каменных горках. Упомянем также очень близкий к гелониопсису род ипсиландра (Ipsilandra) с 5 видами, распространенными в Тибете, Западном и Юго-Западном Китае и Северной Бирме. От гелониопсиса отличается главным образом подковообразными пыльниками (у гелониопсиса они стреловидные).
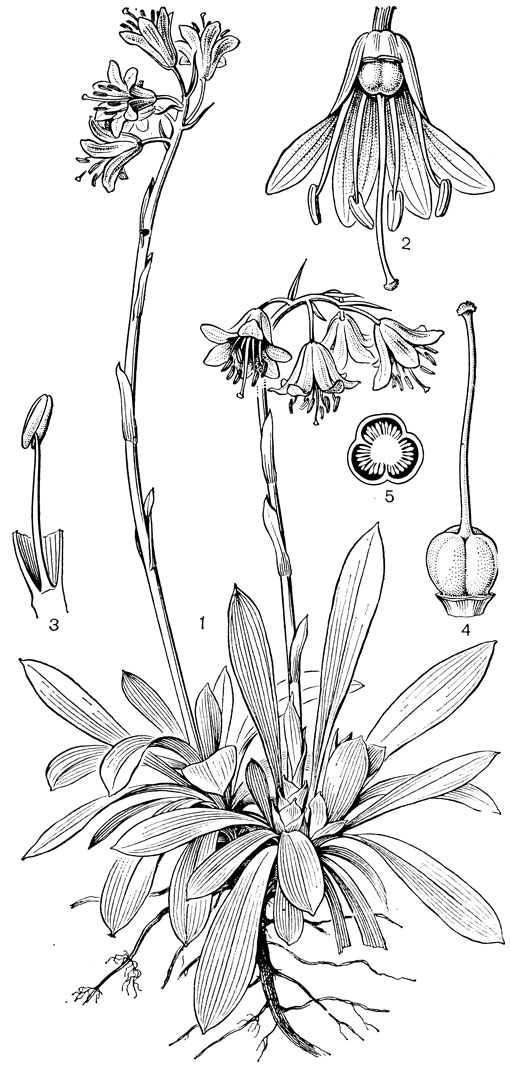
Рис. 28. Гелониопсис японский (Heloniopsis japonica): 1 - общий вид; 2 - цветок; 3 - тычинка; 4 - гинецей; 5 - поперечный разрез завязи
К гелониевым близка небольшая триба хионографисовых (Chionographideae), заключающая всего 2 рода - хионографис (Chionographis, рис.29) ихамелириум (Chamaelirium). Род хионографис (4 вида) встречается в Японии, на острове Чеджудо (расположен к югу от полуострова Корея) и в Южном Китае. Виды хионографиса чаще всего встречаются во влажных тенистых лесах и в горах вдоль ручьев. От короткого толстого корневища этих растений отходит розетка продолговато-яйцевидных или ланцетных листьев; стеблевые листья гораздо мельче и уже. Цветки собраны в колосья, мелкие, белые, слегка пахучие, лишенные прицветников, полигамные (обоеполые и мужские, иногда только мужские), зигоморфные. Сегментов околоцветника 6-3, линейных, однонервных, очень неравных; верхние 4 или 3 значительно длиннее, а нижние 3 или 2 очень короткие или даже отсутствуют. Тычинок 6, с нитями, приросшими к основаниям сегментов околоцветника; пыльники прикрепленные основаниями, раскрываются по бокам. Характерны специализированные пыльцевые зерна, снабженные 4 поровидными апертурами. Завязь верхняя, округло-трехгранная, столбики свободные, линейные, с рыльцевой поверхностью на внутренней стороне. В каждом гнезде завязи по 2 семязачатка. Коробочка локулицидная. Семена веретеновидные, с коротким придатком на одном из концов. Соматическое число хромосом 24 или 42.
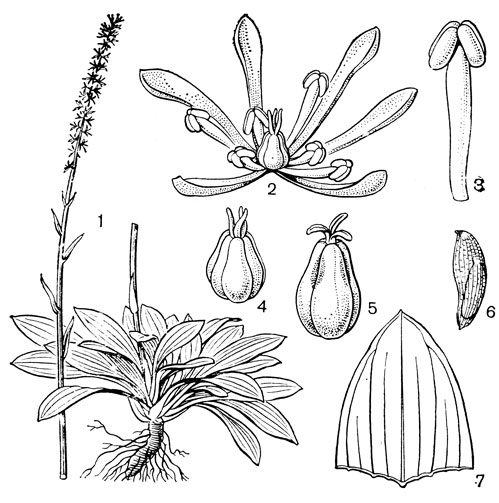
Рис. 29. Хионографис японский (Chionographis japo- nica): 1 - общий вид; 2 - цветок; 3 - тычинка; 4 - гинецей; 5 - плод; 6 - семя; 7 - часть листа
К хионографису близок монотипный род хамелириум (Chamaelirium), в своем распространении приуроченный к восточной части Северной Америки, где произрастает во влажных лесах и на болотах. Растение с клубневидным корневищем, многочисленными базальными листьями, несколькими стеблевыми листьями и рыхлой верхушечной колосовидной кистью однополых цветков. Цветки у хамелириума двудомные, светло-желтые, почти белые, без прицветников. Сегментов околоцветника 6, узколинейно-лопатчатых, одинаковых. Тычинок 6, с тонкими плоскими нитями и белыми экстрорзными пыльниками. Пыльцевые зерна с 4 округлыми апертурами. Завязь 3-гнездная, с многочисленными семязачатками; столбики свободные. Плод - локулицидная коробочка. Семена веретеновидные, окружены косым белым крылом. Корневище хамелириума используют для медицинских целей, а сам хамелириум имеет некоторое декоративное значение.
Последняя триба в подсемействе - собственно мелантиевые (Melanthieae), известная также под названием трибы чемерицевых (Veratreae). Все относящиеся к ней растения - многолетние травы с короткими корневищами или луковицами. В процессе эволюции луковица произошла из укороченного корневища, и поэтому луковичные мелантиевые являются (по крайней мере по этому признаку) эволюционно более подвинутыми. Стебли у мелантиевых облиственные, или же листья базальные. Цветки собраны в густые колосовидные соцветия, кисти или метелки, обоеполые или полигамные. Сегменты околоцветника свободные или почти свободные. Тычинок 6, свободных или прикрепленных к основаниям сегментов околоцветника. Пыльники прикреплены основаниями, экстрорзные. Пыльцевые зерна однобороздные, с сетчатой экзиной. Завязь у них верхняя, а столбики свободные или сросшиеся. Плод - септицидная коробочка. Семена узкие или крылатые. Триба собственно мелантиевых распространена в северной умеренной зоне. Это довольно примитивная группа, которая, вероятно, про-исходит от ближайших предков современных тофилдиевых.
Род чемерица (Veratrum, рис. 30) - самый большой и наиболее примитивный в трибе мелантиевых. В него входит около 45 видов, распространенных главным образом в умеренных, реже в холодных областях северного полушария. Это высокие травы (иногда достигающие в высоту 2 м) с коротким, толстым, прямостоячим, подземным корневищем, от которого отходят кругами шнуровидные поперечно-морщинистые корни. Некоторые авторы, например немецкий ботаник А. Мейер (1882), подземный побег чемерицы считают промежуточным образованием между корневищем, клубнем и луковицей. На верхушке корневища ежегодно развивается крупная трехгранная верхушечная почка, окруженная мясистыми и перегнивающими влагалищами листьев. От верхней части корневища отходит несколько низовых колпачковидных листьев и значительно большее число зеленых листьев. Листья чемерицы складчатые (гофрированные), с сильно развитыми дуговидными жилками, с замкнутыми влагалищными основаниями. Тесно налегающие друг на друга, как бы вставленные друг в друга, листовые влагалища создают впечатление стебля, но это ложный стебель. Чемерица зацветает только на 16-30-й год жизни (М. П. Бахматова, 1980).. Цветонос однолетний, полый, вверху опушенный. Соцветие чемерицы метельчатое, длиной до 60 см. Цветки на коротких цветоножках, мелкие, от беловатых, зеленоватых и зеленовато-желтых до желтых и темновато- или черновато-красных, колокольчатые или колесовидные, полигамные (нормальные обоеполые и более мелкие мужские на одном и том же растении). Сегменты околоцветника свободные, у основания снабженные нектарниками в виде 2 выступающих зеленых мозолей. Тычинки свободные, короче околоцветника, с сердцевидно-яйцевидными пыльниками. Основания тычиночных нитей и сегментов околоцветника приросли к основаниям плодолистиков и таким образом у чемерицы мы наблюдаем начальные стадии возникновения нижней завязи. Но вместе с тем гинецей у чемерицы довольно примитивный. Плодолистики, которые сидят здесь на коротких ножках, как у тофилдии, срастаются между собой только нижней частью, а верхние их части свободные и расходящиеся. Края плодолистиков лишь прижаты друг к другу и гистологически не сросшиеся, а столбики еще очень неясно отграничены от остальной части плодолистика и в верхней части с брюшной стороны открытые (поэтому на поперечном срезе они подковообразные). Плод представляет собой полусинкарпную многолистовку, раскрывающуюся септицидно (точнее, септицидно-вентрально). Семена чемерицы продолговатые, сплющенные, крылатые, разносятся ветром. Характерна четко выраженная протандрия. Сразу же после полного распускания цветков вскрываются пыльники наружного круга, а на следующий день также пыльники внутренних тычинок. На третий день после распускания цветка созревают рыльца, и столбики, прежде торчащие вверх, сгибаются наружу и вниз. Нектар и сильный (но малоприятный для человека) запах привлекают разных представителей отряда двукрылых, главным образом падальных и черных мясных мух (видов родов муха -Musca, каллифора - Calliphora, луцилия - Lucilia, саркофага - Sarcophaga и др.), а также маленьких жуков. Насекомые, посещающие цветки, находящиеся в мужской фазе, загружаются пыльцой, а когда они затем садятся на цветки со зрелыми рыльцами, расположенными теперь на том же самом месте, где прежде находились пыльники, то они неизбежно производят опыление. Виды чемерицы произрастают на пойменных, субальпийских и альпийских лугах, на лесных опушках и полянах, на берегах рек и озер, в кустарниковых зарослях, в разреженных лесах, а также в тундре.
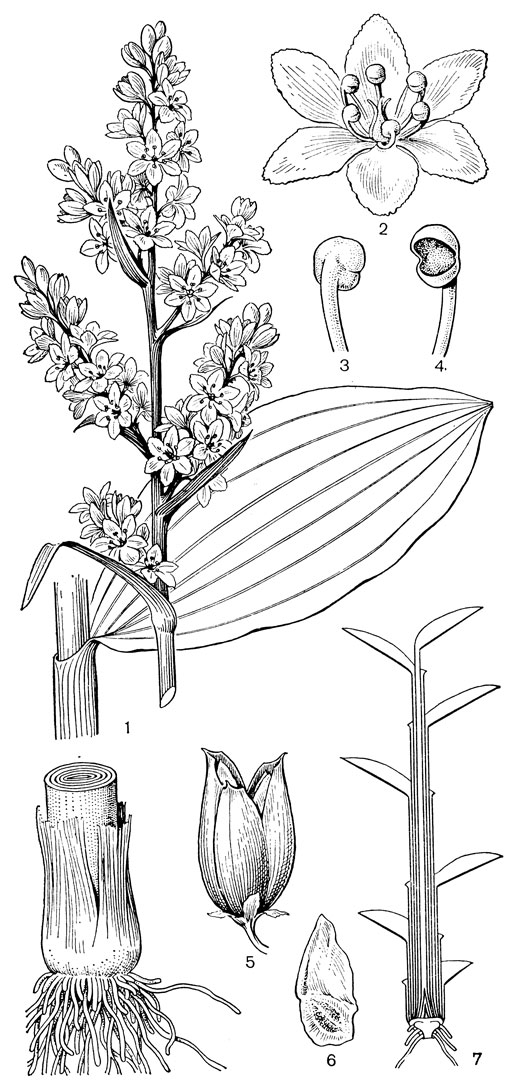
Рис. 30. Чемерица белая (Veratrum album): 1 - соцветие, лист и часть стебля; 2 - цветок; 3, 4 - тычинки; 5 - плод; 6 - семя; 7 - схема продольного разреза растения
Чемерицы - ядовитые растения, содержащие во всех своих частях, главным образом в корнях и корневищах, различные алкалоиды. Они вызывают тяжелые отравления и нередко гибель домашних животных, включая лошадей и птиц. Виды чемерицы относятся к числу самых ядовитых растений сенокосов и пастбищ. Настойка из чемерицы обладает инсектицидным действием и применяется в ветеринарии.
К чемерице близок род мелантиум (Melanthium), по имени которого названо все семейство. Он состоит из 2 видов, распространенных на востоке и юго-востоке Северной Америки. От чемерицы мелантиум отличается главным образом ноготковыми сегментами околоцветника и семенами, окруженными со всех сторон широким крылом. Цветки мелантиума, собранные в большую верхушечную метелку, полигамны (андромоноэция) и протандричны. У основания отгиба имеется плоская ложбинка с двумя желтыми нектарными железами. Цветки посещаются различными насекомыми, преимущественно мухами и жуками.
В отличие от чемерицы и мелантиума, являющихся корневищными растениями, все остальные мелантиевые характеризуются туникатными луковицами. Так как мы впервые в этой книге встречаемся с луковицами, то уместно сказать о них несколько слов. Как корневище, так и луковица представляют собой видоизмененные подземные побеги, служащие для запасания питательных веществ и воды, а также для вегетативного размножения. Но в то время как в корневище запасающей частью является утолщенный стебель, а листья сильно редуцированы, в луковице, напротив, редуцирован стебель, а запасные вещества и вода накапливаются или в разросшихся мясистых влагалищных основаниях обыкновенных зеленых листьев, как у мелантиевых, или же в специализированных мясистых чешуях, представляющих собой более или менее сильно видоизмененные низовые листья, как у многих других представителей порядка лилейных, например у видов рода лилия (Lilium). Луковицы трибы мелантиевых относятся, таким образом, к примитивному типу луковиц, из которого в процессе эволюции возникли другие, более специализированные его типы. Редуцированный стебель луковицы превращен в более или менее плоское, обычно дисковидное образование, называемое донцем. На верхушке донца расположена почка возобновления, а в пазухах листьев - боковые почки, из которых образуются дочерние луковички.
К числу луковичных представителей трибы мелантиевые относятся следующие роды: амиантиум (Amianthium монотипный род, встречающийся на востоке и юго-востоке США), стенантиум (Stenanthium, 4 вида на Сахалине, в Канаде, США и Мексике), схенокаулон (Schoenocaulon, рис. 31, около 9 видов в Северной и Центральной Америке, в Южной Америке до Перу) и зигаденус (Zigadenus, рис. 31), около 15 видов, из которых 1 вид распространен на юге-востоке европейской части СССР, в Сибири, на Дальнем Востоке (кроме Камчатки, Сахалина и Курильских островов), на острове Рисири близ Хоккайдо, в Северном Китае, а все остальные - в Канаде, США и Мексике. В отличие от остальных родов зигаденус характеризуется наличием 1 или 2 зеленоватых нектарных железок близ основания каждого сегмента околоцветника (отсюда название рода от греч. zygon - пара и aden - железа). Почти все части зигаденуса ядовиты. Ряд видов, например североамериканский зигаденус изящный (Z. elegans), имеет декоративное значение.
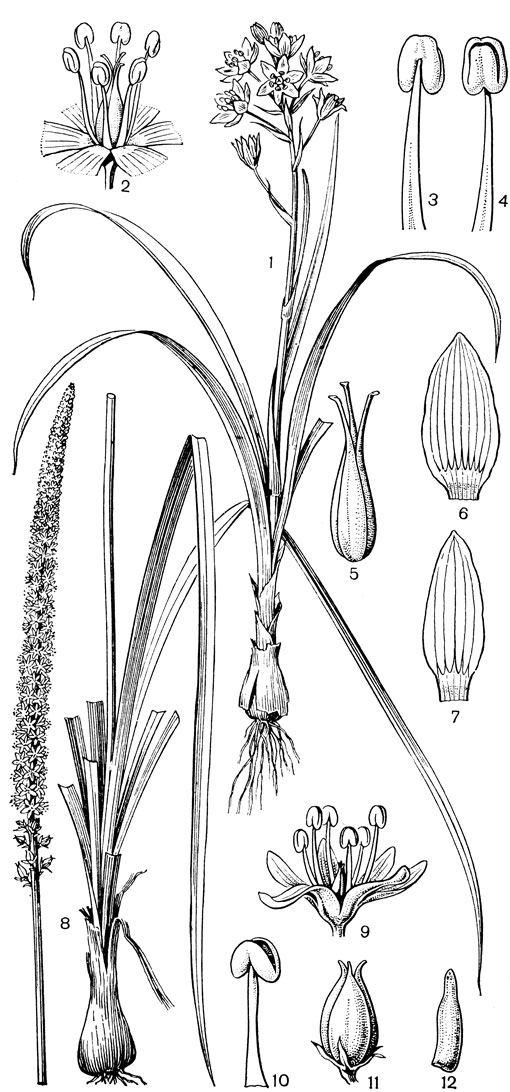
Рис. 31. Мелантиевые. Зигаденус Фремонта (Zigadenus fremontii): 1 - общий вид; 2 - цветок; 3, 4 - тычинки; 5 - гинецей; 6, 7 - сегменты околоцветника. Схенокаулон лекарственный (Schoenocaulon officinarum): 8 - общий вид; 9 - цветок; 10 - тычинка; 11 - плод; 12 - семя
Следующее подсемейство безвременниковые у или колхиковые, гораздо более разнообразно и в целом эволюционно более подвинуто, чем подсемейство мелантиевые. Оно состоит из 5 триб, из которых относительно наиболее примитивна триба увулариевых (Vvnlarieae), обнаруживающая некоторые связи с тофилдиевыми. Это корневищные травы с облиственными стеблями и одиночными или немногочисленными верхушечными цветками. Околоцветник из 6 свободных или сросшихся сегментов. Тычинок 6, свободных; пыльники прикреплены основанием или спинкой, экстрорзные или интрорзные. Гинецей синкарпный, завязь верхняя, а столбики свободные или частично сросшиеся. Плод - локулицидная коробочка. Для некоторых родов характерно наличие трополоновых алкалоидов, из которых наиболее известен колхицин.
Основным родом трибы увулариевых является увулария (Uvularia, рис. 32, 1-4), состоящая из 5 видов, произрастающих в лесах на востоке Северной Америки, от полуострова Новая Шотландия до Флориды. Увуларии - изящные растения обычно с 1, реже 2 цветками, висящими на длинных цветоножках. Так как ветвление стебля увуларии симподиальное, то цветки кажутся боковыми или пазушными, но в действительности они верхушечные. Околоцветник узкоколокольчатый, желтый или зеленовато- желтый, с нектарными ямками у основания свободных сегментов. Завязь 3-лопастная, у увуларии сидячелистной (U. sessilifolia) - на ножке (примитивный признак). Столбики свободные или наполовину сросшиеся. Коробочка треугольная или трехкрылая, с полушаровидными семенами. Растут увуларии в широколиственных лесах и среди кустарников. Они опыляются пчелами и шмелями, а также жуками. Для увуларий характерно также самоопыление, особенно детально изученное у увуларии крупноцветковой (U. grandiflora, рис. 32, 1-4). Расположение пыльников по отношению к рыльцам таково, что при их раскрывании пыльца той половины пыльника, которая направлена внутрь, может пойти на самоопыление, в то время как пыльца наружу направленной половины может быть взята насекомыми даже после того, как совершилось самоопыление. Виды увуларии, особенно увулария крупноцветковая, очень красивы и разводятся как декоративные растения в тенистых местах. К этой же трибе относятся монотипные восточноавстралийские роды крейсигия (Creysigia) и шелхаммера (Schelhammera), а также, возможно, род бурхардия (Burchardia), состоящий из 3 видов, распространенных в Австралии и Тасмании.
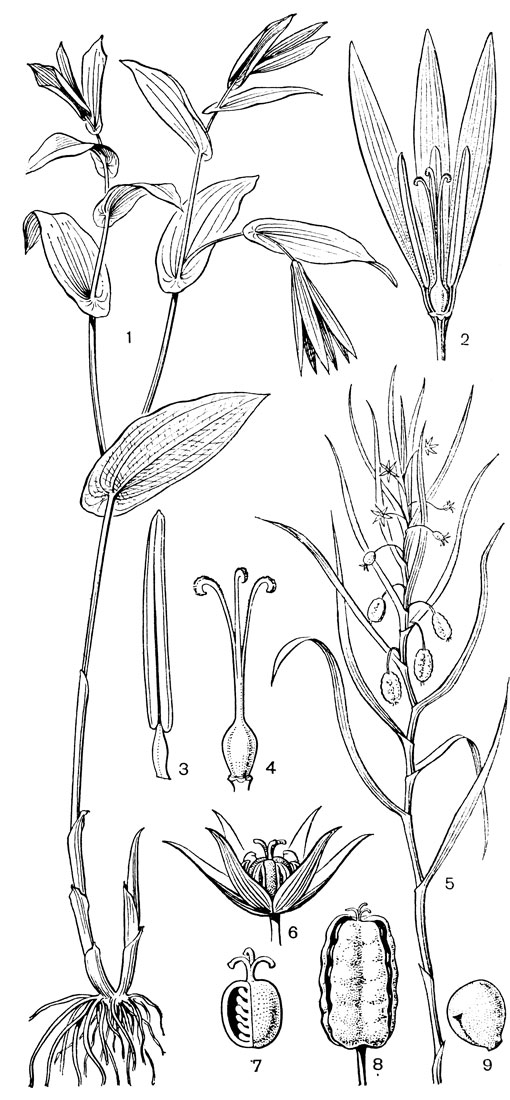
Рис. 32. Мелантиевые. Увулария крупноцветковая (Uvularia grandiflorа): 1 - общий вид; 2 - часть цветка; 3 - тычинка; 4 - гинецей. Ифигения Оливера (Iphigenia oliveri): 5 - верхняя часть растения; 6 - цветок; 7 - продольный разрез завязи; 8 - плод; 9 - семя
Близкородственна увулариевым, но заметно более подвинута триба глориозовых (Glorioseae), к которой относится несколько очень известных декоративных растений. Корневища у глориозовых клубневидные, а листья часто снабжены усиковидными окончаниями. Цветки пазушные, обычно на длинных цветоножках, со свободными или сросшимися в трубку сегментами околоцветника. Тычинок 6, свободных, с экстрорзными пыльниками. Коробочка септицидная или локулицидная. У всех исследованных химиками представителей трибы обнаружены трополоновые алкалоиды, в том числе колхицин.
Наиболее известным родом этой трибы является глориоза (Gloriosa, от лат. gloria - слава), заключающая 5 очень близких видов, которые распространены в тропической и Южной Африке и в тропической Азии. Это высокие лазающие или низкие прямостоячие травы. Карликовые прямостоячие формы, встречающиеся в аридных областях Кении, Сомали и Эфиопии, лишь редко превышают в высоту 30 см, в то время как лазающие формы, произрастающие в менее засушливых условиях, иногда достигают более 5 м. У глориозы коленчатые клубневидные корневища и простые стебли. Листья все стеблевые, сидячие или почти сидячие, очередные (но иногда супротивные или мутовчатые на том же стебле), яйцевидные, обычно (особенно в самой верхней части высоких растений) заканчиваются усиковидным заострением, посредством которого глориоза может прикрепляться к стеблям и ветвям других растений. Цветки сосредоточены в верхней части стебля, довольно крупные, на длинных цветоножках. Сегменты околоцветника свободные, красные, оранжевые или желтые, зеленовато-желтые или чаще двуцветные, с базальным V-образным желтым пятном, а в остальной части оранжевые, красные или пурпуровые; в некоторых цветках желтая окраска бывает как у основания, так и по краям сегментов. В процессе цветения окраска околоцветника подвергается значительным изменениям, что делает глориозу особенно декоративной. По краям сегменты обычно волнистые или даже курчавые, каждый с продольной складкой, в которой укрыт нектарник. Тычинки расходящиеся, с подвижными пыльниками, прикрепленными к нитям спинной стороной. Завязь с многочисленными семязачатками. Своеобразно устроен столбик глориозы. Он нитевидный, длинный и у основания согнут по направлению к оси завязи под прямым или даже почти острым углом. Столбики сросшиеся, заканчиваются тремя обычно короткими рыльцевыми ветвями, редко почти свободные. Коробочка глориозы кожистая, локулицидная, а семена шаровидные, мясистые, ярко-красные. Самым распространенным видом является глиориоза великолепная (G. superba), произрастающая в тропиках Африки и Азии.
По данным К. Краузе (1930) и С. Фогеля (1954), виды глориозы опыляются бабочками. Сильно удлиненные тычинки, снабженные качающимися пыльниками, и столбик, согнутый в сторону пыльников, задевают летящих бабочек, которые на лету высасывают нектар. О. Порш (1931) считал глориозу орнитофильной, но С. Фогель высказывает некоторые сомнения. Виды глориозы, особенно тропическая африканская глориоза Ротшильда (G. rothschildianа) и глориоза великолепная, относятся к числу наиболее красивых декоративных растений. Однако как корневища, так и надземные части глориозы очень ядовиты.
Близок к глориозе род литтония (Littonia), характеризующийся интересным географичесским распространением. В роде 7 видов, распространенных в тропической и Южной Африке и на Аравийском полуострове. Это вьющиеся травы с маленьким клубневидным корневищем, снабженным короткими боковыми лопастями. Стебли обычно простые, прямостоячие или вьющиеся. Листья очередные и мутовчатые, с кончиками, оттянутыми в более или менее усиковидное образование. В отличие от глориозы пыльники прикреплены основаниями, столбик короткий и прямой, коробочка септицидная (по этим признакам литтония несколько менее подвинута, чем глориоза). Семена шаровидные, мясистые, на длинных семяножках. Как декоративное растение давно культивируется южноафриканская литтония скромная (L. modesta), на родине произрастающая среди кустарников и на лесных опушках.
Заслуживает упоминания также монотипный южноафриканский род сандерсония (Sandersonia), распространенный в Трансваале и Натале до востока Капской провинции и в Свазиленде. Растения этого рода - травы с маленьким клубневидным лопастным корневищем и простыми, облиственными стеблями. Цветки у них пазушные, висящие на длинных цветоножках. Сегменты околоцветника сросшиеся в кувшинчатую, короткую, 6-лопастную трубку, у основания с нектарной ямкой. Пыльники прикреплены основаниями, интрорзные. Завязь со многими семязачатками. Столбики свободные, с верхушечными рыльцами. Коробочка локулицидная. Семена шаровидные, с большой широкой семяножкой. Сандерсония нередко культивируется в оранжереях как декоративное растение.
К глориозовым близка триба трициртисовые (Tricyrtideae), состоящая из одного рода трициртис (Tricyrtis), объединяющего около 10-15 видов, распространенных в Восточных Гималаях, континентальном Китае, на Тайване и в Японии. Это короткокорневищные растения с прямым, обычно простым, облиственным стеблем и сидячими, иногда стеблеобъемлющими листьями. Цветки у них одиночные или в верхушечных или пазушных полузонтиках или пучках, довольно крупные, белые, желтые или кремовые, воронковидные или колокольчатые. Сегменты околоцветника свободные, на внутренней стороне обычно пятнистые, у основания внешних сегментов расположены нектарники в виде мешочка или реже короткой шпоры. Нити тычинок более или менее сросшиеся; пыльники прикреплены спинной стороной, экстрорзные. Завязь с многочисленными семязачатками; столбик колончатый, с 3-2-лопастными ветвями. Коробочка септицидная. Семена бурые или черные, плоские. Виды трициртиса произрастают в тенистых лесах. Иногда культивируются как декоративные растения.
Триба ангвиллариевых (Anguillarieae) включает 5 родов, распространенных исключительно в Южной и тропической Африке и Австралии (вместе с Тасманией), т. е. почти исключительно в южном полушарии. Это растение с облиственными стеблями, подземный запасающий орган которых представляет собой клубнелуковицу. Клубнелуковица - это укороченный и клубневидно утолщенный стебель, покрытый чешуями - остатками оснований прошлогодних листьев. На верхушке или вблизи основания клубнелуковицы образуется почка, из которой развиваются листья и цветки, а на нижней части формируются придаточные корни. В пазухах чешуй возникают новые клубнелуковицы или побеги. Ежегодно клубнелуковица заменяется новой. Цветки в кистях или колосьях, лишенные прицветников (большинство родов) или с прицветниками (беометра - Beometra), обычно обоеполые, редко однополые (ангвиллария двудомная - Anguillaria dioica). Сегменты околоцветника свободные или коротко сросшиеся, часто выше основания железистые. Тычинки с экстрорзными пыльниками. Завязь 3-гнездная, с многочисленными семязачатками. Столбики свободные или коротко сросшиеся. Коробочка септицидная или реже локулицидная (ангвиллария). На семенах вдоль рубчика имеется довольно хорошо развитый вырост (типа так называемой строфиолы). Семена обычно более или менее шаровидные. Характерно наличие колхицина. Роды дипидакс (Dipidax, 2 вида), неодрегея (Neodregea, 1 вид) и беометра (1 вид) распространены в Капской области; род вюрмбея (Wurmbea, около 25 видов) встречается в тропической и особенно в Южной Африке, а также в Западной Австралии, а ангвиллария (3 вида) - в Австралии и Тасмании. Представители трибы ангвиллариевых культивируются очень редко.
К ангвиллариевым очень близка триба ифигениевых (Iphigenieae), состоящая из 3 родов. Это также клубнелуковичные травы с облиственными стеблями. Цветки на длинных цветоножках, собраны в кисти и, в отличие от ангвиллариевых, снабжены прицветниками. Сегменты околоцветника свободные. Тычинки с короткими нитями и экстрорзными пыльниками. Завязь обычно 3-гнездная, столбик с головчатым рыльцем или с 3 отогнутыми рыльцевыми ветвями (ифигения - Iphigenia, рис. 32, 5-9). Коробочка локулицидная. Семена шаровидные, с хорошо развитым придатком типа строфиолы, что также сближает эту трибу с ангвиллариевыми.
Самым большим и, по-видимому, относительно самым примитивным является род ифигения, включающий около 12 видов, распространенных в Южной и тропической Африке, на Мадагаскаре, в Индии, Южном Китае, на Филиппинах, в Новой Гвинее и на острове Южном Новой Зеландии. Стебель ифигении простой. Нижние 1-3 листа с трубчатыми влагалищными основаниями или нитевидными пластинками, верхние же листья сидячие, низбегающие, расположенные близ цветка. Цветков 3-10, на согнутых вниз цветоножках. У очень редуцированной ифигении новозеландской (I. novae-zelandiae), высотой от 3 до 8, редко 10 см, цветки одиночные и верхушечные. Но этот вид вообще очень обособлен внутри рода ифигения. Сегменты околоцветника линейные, опадающие (у ифигении новозеландской, однако, остаются и вместе с тычинками сохраняются в засохшем состоянии у основания зрелой коробочки). Пыльники прикреплены к тычиночной нити основанием. Семязачатки многочисленные, и лишь у ифигении новозеландской всего около 8 семязачатков в каждом гнезде завязи, причем завязь чаще всего 2-гнездная. Столбики свободные до основания, с рыльцевой поверхностью на внутренней стороне. Насколько известно, цветки ифигении опыляются пчелиными. На семенах ифигении вдоль рубчика имеется довольно хорошо развитый вырост, который, возможно, служит для привлечения муравьев. Семена встречающейся в Индии ифигении звездчатой (I. stellata) содержат от 1,2 до 2% колхицина, т. е. в несколько раз больше, чем семена безвременника (Colchicum), что делает это растение исключительно важным источником этого ценного алкалоида.
К ифигении очень близка кампториза (Саmptorrhiza), состоящая из 4 видов, распространенных в более северных районах Южной Африки, в Намибии, Ботсване, Зимбабве и Мозамбике. В отличие от ифигении сегменты околоцветника продолговатые, вогнутые, с завернутыми краями; короткие нити тычинок в середине шаровидно вздутые, пыльники подвижные, во время цветения - с горизонтально расположенными гнездами; столбики сросшиеся, с маленьким верхушечным рыльцем.
Но наиболее интересен третий род трибы - орнитоглоссум (Ornithoglossum), состоящий из 2 видов, произрастающих в засушливых областях Южной, Юго-Западной и тропической Африки. Для орнитоглоссума характерен остающийся околоцветник и свободные столбики, заканчивающиеся очень маленькими головчатыми рыльцами. Они опыляются пчелами (орнитоглоссум зеленый - О. viride) или мухами (орнитоглоссум сизый - О. glaucum). Оба вида протандричны. Сегменты околоцветника немного поникающих цветков орнитоглоссума зеленого загнуты кверху и назад, но иногда один сегмент остается направленным вперед. Сегменты цветков белые, но их края и капюшоновидные верхушки красно-фиолетового цвета. Тычинки, прикрепленные к основаниям сегментов околоцветника, загнуты книзу и вперед и до некоторой степени играют роль нижней губы цветка. После выпадения пыльцы из пыльников тычинки откидываются назад, а 3 рыльца занимают их место. Нектар находится в мешковидных складках в нижней части сегментов околоцветника и доступен только насекомым с удлиненным хоботком. Цветки орнитоглоссума сизого также поникающие, но, в отличие от предыдущего вида, они радиально-симметричные. Сегменты околоцветника все загнуты кверху. У основания они бледно-зеленые, а вытянутая в остроконечие верхушка коричневая. Нектарники в виде плоских ямочек с открыто расположенными каплями нектара. Расположение нектара, пыльников и рыльца приспособлено к опылению мухами (С. Фогель, 1954).
Наконец, последней трибой в подсемействе является триба безвременниковых, или колхиковых (Colchiceae). Подземные органы этих растений представляют собой клубнелуковицу, а листья собраны в розетку. Цветки пазушные или верхушечные. Сегменты околоцветника свободные до основания и с длинными ноготками, или же околоцветник сростнолистный, воронковидно-колокольчатый, с длинной цилиндрической трубкой и 6-раздельным отгибом. Тычинки прикреплены к околоцветнику, с интрорзными пыльниками. Пыльцевые зерна специализированного типа, 2-, 3- или 4-поровые. Нектарники расположены на тычиночных нитях (стаминальные). Гинецей синкарпный, большей частью со свободными, нитевидными столбиками, реже столбики сросшиеся и лишь наверху 3-раздельные (брандушка - Bulbocodium). Коробочка септицидная, с шаровидными семенами. На семенах имеются придатки такого же типа, как у ифигении.
Самый примитивный род этой трибы - андроцимбиум (Androcymbium, рис. 33) - занимает промежуточное положение между трибами безвременниковых и ифигениевых. Он заключает около 35 видов, распространенных главным образом в Южной и Восточной тропической Африке; 3-4 вида андроцимбиума встречаются в Макаронезии и в Средиземноморье (от Марокко и Юго-Восточной Италии до Синайского полуострова и Южной Сирии и Палестины). На территории Европы произрастает лишь один эндемичный вид - андроцимбиум европейский (А. europaeum), приуроченный к сухим скалистым местам между Альмерией и мысом Гата на крайнем юго-востоке Испании. Андроцимбиумы - травы с коротким, преимущественно подземным стеблем и глубокосидячей, асимметричной клубнелуковицей, покрытой твердой, кожистой, темно-коричневой туникой. Листья (2 или больше) с широким влагалищным основанием образуют розетку, которая обволакивает в виде покрывала соцветие. Цветков 1-6, собранных в верхушечное головчатое соцветие, напоминающее один цветок. Околоцветник белый или розовый, иногда с пурпурными жилками или пятнами. Сегменты околоцветника свободные, с длинным узким ноготком и эллиптическим или ланцетным отгибом. Тычинки прикреплены к основанию отгиба. Не полностью сросшиеся плодолистики постепенно вытянуты в столбики, заканчивающиеся маленьким рыльцем. Виды андроцимбиума приурочены к сухим местообитаниям. Опыляются они пчелиными или мухами. Цветки видов, опыляемых пчелами, издают приятный медовый запах, в то время как цветки видов, опыляемых мухами, пахнут неприятно. Нектар выделяется небольшой темно-красной железой, расположенной в нижней части или у основания тычиночной нити.
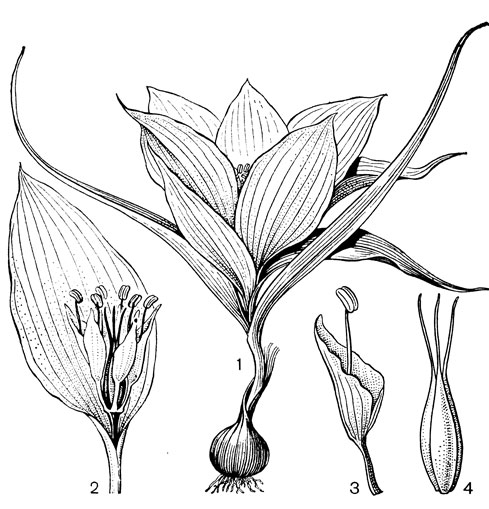
Рис. 33. Андроцимбиум мелантиевидный (Androcymbium melanthioides): 1 - общий вид; 2 - цветок; 3 - тычинка; 4 - гинецей
С родом андроцимбиум очень тесно связан род мерендера (Merendera, рис. 34), в котором около 15 или более видов, распространенных в Южной и Юго-Западной Европе и в Средиземноморской и Ирано-Туранской флористических областях. На востоке виды мерендеры достигают Пакистана и Пенджаба (Индия). Клубнелуковица мерендеры защищена одной или несколькими кожистыми или перепончатыми чешуями, вытянутыми на верхушке в более или менее длинную шейку, охватывающую нижнюю часть влагалища. Самая внутренняя чешуя представляет собой замкнутую тунику, облегающую всю клубнелуковицу, остальные представляют собой разорванные туники прошлых лет. Если снять все чешуи, то на верхушке прошлогодней клубнелуковицы можно видеть засохшие остатки оснований прошлогодних листьев, а снаружи сбоку место, от которого отходили прошлогодние корни. Новая клубнелуковица (клубнелуковица этого года, несущая зеленые листья и цветки) возникла у основания старой клубнелуковицы но в результате асимметричного роста этой последней новая клубнелуковица оказывается втянутой глубже в почву. Рассматривая внимательно основание молодой клубнелуковицы этого года, можно заметить зачаток клубнелуковицы будущего года. Листья с замкнутыми влагалищами, которые полностью заключены в подземное влагалище видоизмененного низового листа. Цветки обычно появляются одновременно с листьями (реже раньше) и вместе с нижними частями листьев на ранних стадиях развития бывают заключены во влагалище низового листа. Цветки одиночные или собраны по нескольку в пучках, почти сидячие, распускаются сразу после таяния снега и поэтому на юге обычно называются "подснежниками". Околоцветник розовый или белый. Сегменты его свободные, с длинными узкими ноготками. В зеве иногда имеются зубчатые придатки, которые служат для скрепления соседних сегментов. Наличие длинных ноготков объясняется тем, что у этого типичного геофита вся нижняя часть цветка вместе с завязью и нижней частью столбиков находится в земле, иногда довольно глубоко. Только благодаря длинным ноготкам окрашенные отгибы сегментов околоцветника выносятся на поверхность, Тычинки прикреплены к основаниям отгиба.
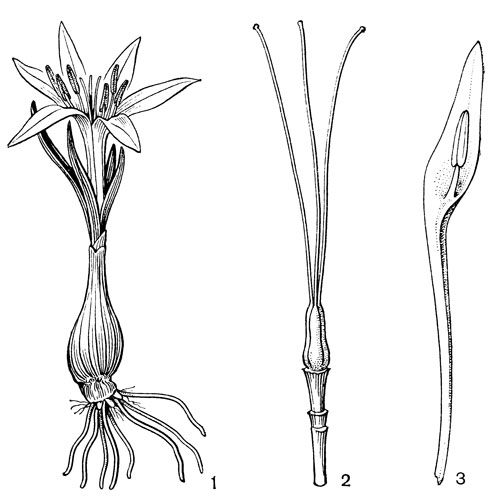
Рис. 34. Мерендера Эйтчисона (Merendera aitchisonii): 1 - общий вид; 2 - гинецей; 3 - тычинка
От подземной завязи отходят свободные, длинные, нитевидные столбики, выносящие на поверхность мелкие головчатые рыльца. При плодах подземный стебель немного возвышается над землей, и тем самым развивающийся плод оказывается также над землей. Зрелая коробочка раскрывается у верхушки, а не по всей длине, что объясняется низким ее положением на поверхности почвы.
Как и другие представители трибы, виды мерендеры содержат алкалоиды, в том числе колхицин, а также мерендерин. Они локализованы главным образом в листьях, цветках и семенах. Особенно богаты ими семена. Количество алкалоидов в клубнелуковицах относительно небольшое, но к моменту плодоношения их содержание заметно увеличивается. Наибольшее количество колхицина содержится в незрелых семенах.
Одни виды мерендеры произрастают на сухих каменистых, песчаных или глинистых местах; другие предпочитают влажные горные склоны или альпийские луга. Большинство видов цветет весной или (высокогорные виды) в начале лета, но некоторые виды, как, например, эндемичная для Центральных Пиренеев мерендера горная (М. montana), цветут осенью.
Опыляется мерендера главным образом пчелами. По наблюдениям Ц. Р. Тонян (1949), у кавказско-переднеазиатской мерендеры трехстолбиковой (М. trigyna) наблюдается тенденция к однополости. Имеются две формы, связанные между собой переходами. Первая из них характеризуется двумя - четырьмя цветками, обычно широколанцетными сегментами околоцветника, нормально развитым гинецеем и нормально развитыми семязачатками, но относительно более мелкими пыльцевыми зернами с несколько более низким процентом прорастаемости (81%). Вторая форма характеризуется более низкими растениями с одиночными, редко двойными цветками, большей частью узколанцетными лепестками, обычно редуцированной завязью (длиной 2 мм) с очень короткими столбиками (длиной 0,5 мм), рудиментарными семязачатками и несколько более крупными пыльцевыми зернами, характеризующимися 100-процентной прорастаемостью. Очевидно, мы имеем здесь дело с самыми начальными стадиями дифференциации на женские и мужские растения.
Несколько более специализирован небольшой род брандушка (Bulbocodium), состоящий из 2 видов, распространенных в Средиземноморье и в Европе (за исключением северных районов). От мерендеры брандушка отличается главным образом сросшимся, лишь наверху 3-раздельным столбиком и ноготками сегментов околоцветника, сложенными трубкой и связанными между собой при помощи зубцов, имеющихся с каждой стороны при основании отгиба. Виды брандушки растут на горных лугах (эндемичная европейская брандушка весенняя - В. vernum) или же в степях, среди кустарников, на сырых полянах, заливных лугах (брандушка разноцветная - В. versicolor). Характерна протогиния. Когда цветок только начинает распускаться, кончики сегментов околоцветника расходятся лишь на расстояние 15-20 мм друг от друга. На концах расходящихся лопастей столбика развиваются рыльцевые сосочки. Закрытые еще пыльники, расположенные плотно в 2 ряда, окружают тесным кольцом столбик ниже его свободных частей. Цветки издают в это время сильный медовый запах. Насекомые-опылители (преимущественно пчелы и шмели), имеющие свободный доступ к нектару, расположенному между тычиночными нитями и сегментами околоцветника, при посещении цветка легко касаются рылец. Если до этого они уже были на растениях, находящихся на более поздней мужской стадии, то производят перекрестное опыление. Позднее сегменты околоцветника расходятся дальше друг от друга, пыльники вскрываются и бывают покрыты пыльцой. Тем временем тычинки еще больше выставляются наружу. Сегменты околоцветника ниже места прикрепления к ним тычинок удлиняются, и пыльники тем самым приподнимаются. Так наступает мужская стадия.
Род безвременник, или колхикум (Colchicum, рис. 35), по которому названо подсемейство, один из самых больших в семействе, насчитывающий около 45 или более видов, распространенных в Средиземноморье, Европе (за исключением северных районов), Западной, Средней и Центральной Азии, Гималаях, Тибете. В отличие от трех остальных родов трибы околоцветник у безвременника сростнолистный, воронковидно-колокольчатый, с длинной цилиндрической трубкой. Листья и цветки заключены в перепончатое влагалище. Цветки одиночные или в пучках, снабжены маленьким прицветником и сидят на очень короткой ножке, пурпуровые, розовые, желтые или белые. Тычинки прикреплены в зеве околоцветника, три из них более короткие и три (внутренние) более длинные. Столбики свободные, нитевидные, полые, с булавовидными рыльцами. Завязь подземная. Клубнелуковица покрыта пленчатыми или кожистыми бурыми чешуями, которые обычно продолжены в более или менее длинную шейку, охватывающую нижнюю часть растения. Цветки и листья появляются более или менее одновременно весной - в первой половине лета (у высокогорных видов), или цветки появляются в конце лета или осенью, а листья и плоды - весной следующего года. Семена видов безвременника снабжены большим мясистым разрастанием, проходящим от халазы до микропиле вдоль рубчика. В зрелом семени оно сильно увеличивается и занимает большую часть поверхности семени. Это ариллоид, который, в отличие от настоящей элайосомы, лишен масла, но содержит крахмал и в зрелом состоянии выделяет на своей поверхности сахара в виде капель. Предполагают, что этот придаток семени служит для привлечения муравьев. Все органы растения очень ядовиты, так как содержат ряд алкалоидов, в том числе колхицин.
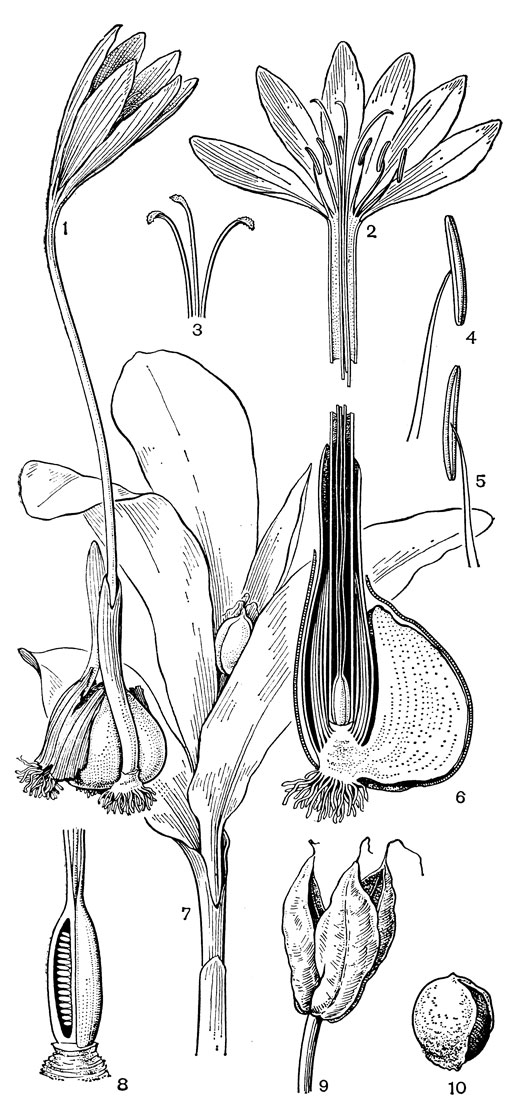
Рис. 35. Безвременник Борнмюллера (Colchicum bornmuelleri): 1 - общий вид; 2 - цветок в развернутом виде; 3 - трехлопастное рыльце; 4, 5 - тычинки; 6 - продольный разрез клубнелуковицы; 7 - часть стебля с плодом; 8 - продольный разрез завязи; 9 - плод; 10 - семя
Виды безвременника произрастают в самых разных местообитаниях: на лугах и в степях, на открытых каменистых склонах, под скалами, среди кустарников, в лесах (преимущественно горных). Цветки безвременника осеннего (С. autumnale) и других видов протогиничны и опыляются пчелами и, менее эффективно, многочисленными мухами. Оранжевый нектарник, который помещается немного выше места срастания тычиночной нити с сегментом околоцветника, выделяет нектар, заполняющий короткие желобки сегмента околоцветника, и защищен шерстистыми волосками. Так же как виды брандушки, безвременниковые очень декоративны и являются хорошим объектом для озеленения садов, парков и скверов весной и осенью.
Семейство калохортовые (Calochortaceae) (А. Л. Тахтаджян)
К мелантиевым, особенно к подсемейству безвременниковых, стоит близко семейство калохортовые, включающие один род - калохортус (Calochortus, рис. 36). В этом роде около 60 видов, распространенных от южной Британской Колумбии (Канада) до Гватемалы, с наибольшей концентрацией видов в Калифорнии; в Северной Америке виды калохортуса распространены в западных областях, доходя на востоке до Западной Небраски и Дакоты.
Калохортусы - травы с туникатными луковицами, одетыми чешуями или реже с волокнисто-сетчатой оболочкой. У калохортуса наблюдается своеобразный способ погружения (заглубления) луковиц, обнаруженный М. В. Барановой (1981) у проростков изученных ею видов. По ее наблюдениям, почка возобновления проростка располагается на укороченном гипокотиле в полости влагалища семядоли. По мере развития проростка паренхимные клетки первичного корня и его верхней части под гипокотилем, так же как клетки влагалища семядоли, начинают постепенно разрушаться. Однако проводящий пучок, соединяющий влагалище семядоли с первичным корнем через гипокотиль, сохраняется и продолжает функционировать. Но самое замечательное заключается в том, что почка возобновления, которая, как на шнуре, закреплена в полости, образованной корнем и влагалищем семядоли, постепенно погружается в полость корня. При погружении почки пучок сверху натягивается, а снизу под нею ослабляется. В результате проводящий пучок принимает под почкой вид свободно сложенного петлями шнура. В дальнейшем, разрастаясь, почка возобновления рвет сухую оболочку влагалища и корня и освобождается от их покрова. Одновременно ссыхается и рвется проводящий пучок влагалища. Как показали наблюдения М. В. Барановой, в первый год развития сеянцы видов калохортуса погружаются в почву на глубину до 13 см. Подобное же явление погружения было отмечено у нескольких видов безвременника и ириса.
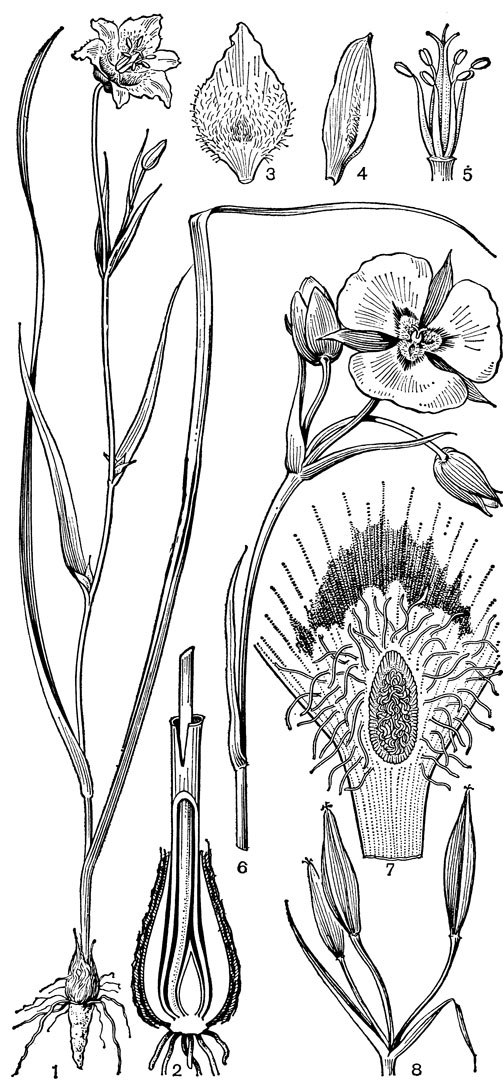
Рис. 36. Калохортус желтый (Calochortus luteus): 1 - общий вид; 2 - продольный разрез луковицы; 3, 4 - лепестки; 5 - андроцей и гинецей; 6 - часть стебля с бутонами и цветком; 7 - часть лепестка; 8 - плоды
Стебли калохортусов обычно ветвистые, высотой от 10 до 80 см, редко до 2 м. Листья немногочисленные, приземные и стеблевые, линейные. Устьица аномоцитные. Цветки крупные, верхушечные, одиночные или собраны в зонтиковидное соцветие. Околоцветник колокольчатый или звездчатый, из 6 свободных сегментов, обычно более или менее ясно дифференцированных на чашелистики и лепестки, редко все сегменты лепестковидные, как у мексиканского калохортуса Хартвега (С. hartwegii). Чашелистики от яйцевидных до узколанцетных, обычно более или менее зеленые, у основания обычно пятнистые; лепестки крупнее и шире, различной окраски, клиновидные или ноготковые, более или менее волосистые, с нектарной ямкой при основании, часто с 1 или 2 пятнами над ямкой. Тычинок 6, расположенных в 2 ряда, короче околоцветника; пыльники прикрепленные основаниями, раскрывающиеся латерально. Пыльцевые зерна 1-бороздные. Гинецей из 3 плодолистиков, синкарпный (по-видимому, вторичносинкарпный, возникший в результате срастания выпяченных внутрь постенных плацент); завязь верхняя, 3-гнездная, на поперечном сечении треугольная или трехкрылая (крылья развиваются вдоль средней жилки плодолистиков), с сидячим рыльцем. Семязачатки многочисленные, анатропные и битегмальные. Эндосперм нуклеарный. Плод - септицидная коробочка, трехгранная или трехкрылая, от линейной до шаровидной. Семена многочисленные, с линейным зародышем средних размеров.
Виды калохортуса встречаются в самых различных местообитаниях, от тепло-умеренной до тропической зоны. Они произрастают как в аридных условиях, так и в горных лесах и на лугах, поднимаются до субальпийского и реже альпийского пояса.
Еще известный бельгийский ботаник Б. Ш. Ж. Дюмортье (1829) выделил род калохортус в отдельное семейство, которое, однако, вплоть до последнего времени редко кем признавалось. Обычно калохортус включают в подсемейство лилейные (Lilioideae) семейства лилейные (Liliaceae). Однако от лилейных калохортус отличается многими существенными признаками. Как показала М. С. Кейв (1941, 1953), у калохортуса зародышевый мешок нормального типа, в то время как у всех родов подсемейства лилейных он типа фритилларии. Но, как указали еще Ф. Буксбаум (1937) и К. Шнарф (1949), калохортус отличается от типичных лилейных также способом раскрывания коробочки, строением семени, прорастанием, размером фуникулуса, формой краев плодолистиков в столбике, основным числом хромосом (n = 6, 7, 8, 9, 10) и анатомией листа, а также многоклеточной эпидермой верхушки нуцеллуса (Р. Берг, 1960). Как указывает Р. Берг, имеется гораздо больше общего между калохортусом и подсемейством мелантиевые (в нашем понимании семейство мелантиевые). Но и от мелантиевых он отличается достаточно большим числом признаков, включая целый ряд эмбриологических признаков (Р. Берг,. 1960). Поэтому, следуя Хуберу (1969), лучше всего рассматривать калохортус как представителя самостоятельного семейства калохортовые.
Ряд видов калохортуса представляет интерес в качестве бордюрных растений и особенно для каменных горок, а иногда также как комнатные растения.
Семейство эррериевые (Herreriaceae) (Т. В. Вельгорская)
Семейство эррериевые, состоящее из 2 родов и около 10 видов, распространено в южном полушарии и имеет разорванный ареал: род эррерия (Herreria) с 8-9 видами встречается в субтропической и тропической Южной Америке, а род эррериопсис (Herreriopsis) с единственным видом эррериопсисом изящным (Н. elegans, рис. 37) обитает на острове Мадагаскар.
Эррериевые - травянистые многолетники с прямостоячим или вьющимся стеблем, иногда снабженным небольшими шипами, и с клубневидным корневищем. Сосуды с лестничной перфорацией имеются в корнях и в стебле. Узкие, линейные или ланцетные кладодиевидные жесткие листья с частым жилкованием собраны в боковые пучки на сильно укороченных побегах. По данным М. А. Барановой (1982), устьица эррериевых аномоцитные. Соцветия расположены в пазухах мелких, почти чешуевидных прицветников и представляют собой редкие кисти или метелки. Цветки обоеполые, на более или менее длинных цветоножках с маленькими прицветничками. Сегментов околоцветника 6, свободных, у эррерии равных и отгибающихся, а у эррериопсиса внешние сегменты линейные и внизу коленчато изогнутые (рис. 37, 6), внутренние - обратноланцетные, в основании расширенные и опушенные ресничками, вверху голые. В основании сегментов околоцветника эррериопсиса расположены нектарники. Тычинок 6, свободных или сросшихся в основании; тычиночные нити тонкие, у эррерии они короткие, у эррериопсиса нити длиннее сегментов околоцветника, и прикрепленные спинкой интрорзные пыльники заметно возвышаются над околоцветником. Пыльники вскрываются продольной щелью. Пыльцевые зерна однобороздные. Гинецей синкарпный; завязь нижняя, 3-гнездная с 3-6 (у эррерии) или многочисленными (у эррериопсиса) анатропными %семязачатками в каждом гнезде. Столбик у эррерии короткий, с трехлопастным рыльцем (рис. 37,2); у эррериопсиса столбик длинный, с маленьким головчатым рыльцем. Плод эррериевых - локулицидная 3-гнездная коробочка, позднее вскрывающаяся септицидно, содержит от 3-6 до многочисленных (у эррериопсиса) семян. Семена черные, мелкие, плоские, с относительно большим крылом (рис. 37, 4, 8). Зародыш маленький, прямой, цилиндрический, окруженный обильным эндоспермом.
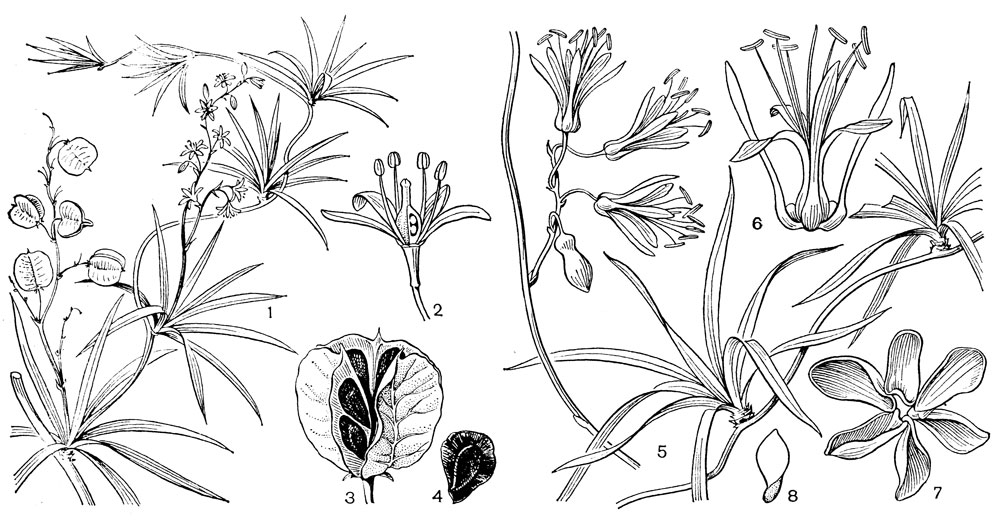
Рис. 37. Эррериевые. Эррерия звездчатая (Herreria stellata): 1 - часть растения с плодами и цветками; 2 - цветок (2 тычинки срезаны, видны семязачатки); 3 - плод; 4 - семя. Эррериопсис изящный (Herreriopsis elegans): 5 - часть растения с пучками листьев и соцветием; 6 - цветок; 7 - раскрытая коробочка; 8 - семя
Мелкие желтоватые цветки эррерии, хотя и невзрачны, необычайно ароматны. Обладают приятным запахом также и красные или желтые цветки эррериопсиса, к тому же они еще содержат и нектар. И хотя сведений об опылении эррериевых нет, можно с уверенностью предположить, что они опыляются насекомыми. Цветут эррерии в феврале - марте, а эррериопсис изящный начинает цвести во время сухого сезона на Мадагаскаре, когда на растении еще нет листьев, которые появляются в период сезона дождей.
Снабженные крылом легкие семена эррериевых распространяются ветром. Упавшие на землю семена могут вместе с комочками почвы разноситься на ногах животных или человека.
Род эррерия был назван в честь испанского ученого и агронома XVI в. Габриэля де Эрреры. Среди видов эррерии наиболее известна эррерия сарсапарилловая (Herreria salsaparilha), произрастающая в дождевых лесах Юго-Восточной Бразилии в штатах Баия и Минас-Жерайс. Местные жители используют эту лиану с шершавым колючим стеблем, достигающим 4-метровой длины, в народной медицине. Они называют ее "сарсапариллой". Следует отметить, что это название в Латинской Америке относится к растениям нескольких семейств, эти растения представляют собой более или менее колючие лианы и обладают сходными лечебными свойствами. В Перу и на севере Чили растет эррерия звездчатая (Н. stellata, рис. 37,1-4) - изящное растеньице с мелкими, звездчатообразно расположенными сегментами околоцветника. Широко распространена в Парагвае, Уругвае, Северной Аргентине и Восточной Бразилии эррерия монтевиденская (Н. mоntevidensis) - лиана с гладким стеблем и кистями прямостоячих цветков.
На юго-западе Мадагаскара по берегам рек, на сухих холмах и по побережью озера Кинкони довольно часто встречается эррериопсис изящный с красивыми алыми цветками или его разновидность - эррериопсис желтоцветковый (Herreriopsis elegans var. luteiflora). Он растет на сухих песчаных, известняковых или каменистых почвах. Клубни эррериопсиса сладковатого вкуса, но содержат токсические вещества, тем не менее их очень любят и безо всякого вреда для себя едят дикие кабаны. Местное население использует клубни эррериопсиса в народной медицине.
Семейство лилейные (Liliaceae) (Е. В. Мордак)
Это семейство содержит 45 родов и около 1300 видов, распространенных в умеренных областях Евразии, Африки и Северной Америки с немногочисленными представителями в горах тропической Африки и Южной Америки.
Лилейные - многолетние травянистые луковичные растения. Самое высокое из них - гималайский вид кардиокринум гигантский (Cardiocrinum giganteum) - достигает 4 м, а самое маленькое - южноафриканский литантус крохотный (Litanthus pusillus): его высота вместе с луковицей величиной с горошину составляет всего 25 мм. Среди лилейных известно два эпифита. Это родокодон ургинеевидный (Rhodocodon urgineoides) на острове Мадагаскар и лилия древесная (Lilium arboricola) в Восточной Азии. Но и у них имеется луковица, скрытая в моховом покрове ствола дерева.
Строение луковиц, способы образования замещающей луковицы и дочерних луковичек и способы их заглубления у лилейных очень разнообразны. Обычно заглубление луковиц происходит с помощью особых втягивающих (контрактильных) корней, толстых и сочных. По мере высыхания они сжимаются (укорачиваются) в вертикальном направлении, втягивая за собой луковицу, часто на значительную глубину. У тюльпанов (Tulipa) замещающая луковица втягивается с помощью столона заглубления, внутри которого она находится. Луковицы лилейных многолетние, состоящие из нескольких годовых циклов, или однолетние, ежегодно возобновляющиеся. И те и другие составлены одними листовыми или одними низовыми чешуями или сочетанием тех и других; чешуи могут быть широкими, замкнутыми или не замкнутыми, иногда узкими (черепитчатыми). Кроме сочных запасающих листовых и низовых чешуй, могут быть пленчатые влагалищные чешуи, чаще замкнутые, верхушка которых находится на поверхности и охватывает основание зеленых листьев. У однолетних луковиц к концу сезона вегетации все чешуи отмирают, а в старых оболочках остается новая замещающая луковица, заложившаяся ранее у основания цветоноса. У многолетних луковиц часть чешуй отмирает, поэтому в луковице накапливаются чешуи за несколько лет или годовых циклов. В годичном цикле у разных видов имеется определенное соотношение одних и других чешуй. Наиболее древним типом считаются крупные, почти наземные многочешуйчатые луковицы, составленные только листовыми чешуями; наиболее подвинутыми, специализированными - малочешуйчатые, ежегодно возобновляющиеся, глубоко залегающие. Луковица, представляющая собой специализированный побег, имеет два типа ветвления: моноподиальное и симподиальное. Определить тип ветвления можно лишь на ранних стадиях развития побега. У моноподиальной луковицы зачаток первого листа почки возобновления обращен к цветоносу брюшной стороной, а у симподиальной луковицы - спинной (рис. 38, 5, 6).
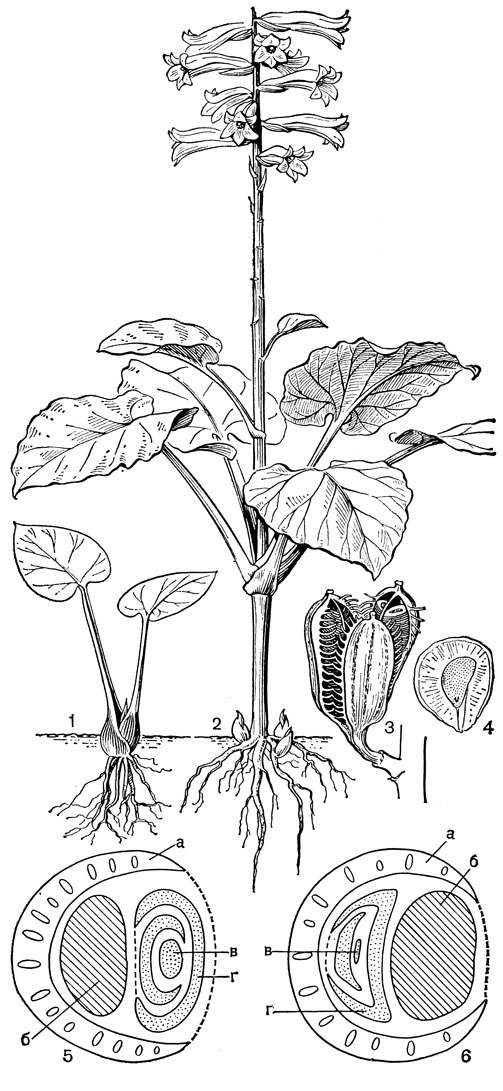
Рис. 38. Кардиокринум сердцевидный (Cardiocrinum cordatum) и два типа луковиц у лилейных: 1 - молодое (ювенильное) растение с луковицей; 2 - цветущее растение; 3 - плод; 4 - семя; 5 - моноподий (а - влагалище последнего внутреннего листа, б - цветонос, в - почка возобновления, г - зачаток первого листа почки); 6 - симподий (обозначения те же)
Надземные цветоносные стебли бывают облиственные и безлистные - стрелки, или цветоносы. В последнем случае все листья собраны в приземном пучке (базальные). Листья цельные, чаще ланцетные или линейные, иногда сердцевидные и на черешках (роды кардиокринум - Cardiocrinum и дримиопсис - Drimiopsis), обычно с параллельным жилкованием.
Цветки от мелких до крупных, одиночные или собраны в верхушечные соцветия, обычно кисти. Прицветники обычно мелкие, неокрашенные, иногда они в виде крупных зеленых листьев собраны на верхушке стебля (некоторые виды рябчика, или фритилларии - Fritillaria, род эвкомис - Eucomis). Цветки обоеполые, обычно актиноморфные, реже несколько зигоморфные, как, например, у кардиокринума и некоторых видов из родов камассия (Camassia), рябчик, белъвалия (Bellevalia), мускари (Muscari) и лашеналия (Lachenalia). Околоцветник венчиковидный, из 6 сегментов в 2 кругах; сегменты свободные или сросшиеся в трубку; сегменты наружного круга обычно незначительно отличаются от сегментов внутреннего круга. Нектарники примитивные, расположены в основании сегментов околоцветника (тепальные, рис.39,40) или септальные (рис.43).Тычинок 6, расположенных в 2 круга. Нити тычинок прикреплены к основанию сегментов или к трубке околоцветника, свободные или сросшиеся; пыльники прикреплены к тычиночным нитям спинкой или основанием и вскрываются обычно продольной щелью, интрорзные. Пыльцевые зерна однобороздные. Гинецей состоит из 3 сросшихся плодолистиков. Завязь верхняя, с многочисленными или несколькими обычно анатропными семязачатками. Цветки лилейных часто ароматные, с большим количеством нектара, опыляются различного рода насекомыми, а некоторые - птицами. Плод - локулицидная коробочка. Семена плоские или шаровидные.
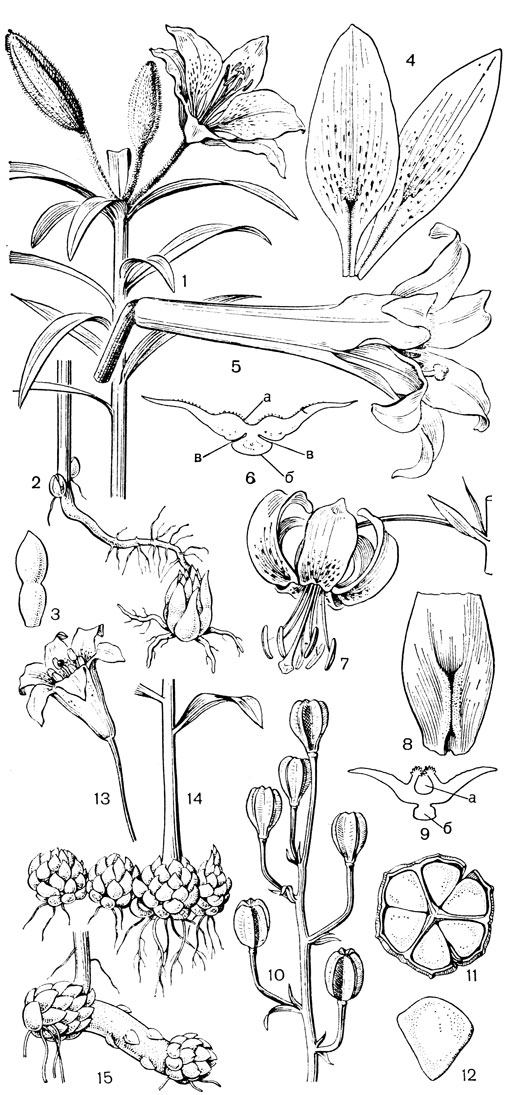
Рис. 39. Лилии. Лилия даурская (Lilium pensylvanicum): 1 - общий вид; 2 - подземная часть; 3 - чешуя луковицы; 4 - сегменты околоцветника. Лилия длинноцветковая (L. longiflorum): 5 - цветок; 6 - поперечный срез сегмента околоцветника в нижней части (а - нектарная канавка, б - вырост центральной жилки, в - выемки, куда плотно входят края наружных сегментов). Лилия кудреватая, или саранка (L. martagon): 7 - цветок; 8 - нижняя часть внутреннего сегмента с внутренней стороны, на которой находится нектарная канавка, прикрытая кожными складками; 9 - поперечный срез сегмента (обозначения те же); 10 - стебель с коробочками - 'канделябр'; 11 - поперечный разрез коробочки; 12 - семя. Лилия малая (L. parvum): 13 - цветок; 14 - плагиотропные луковицы. Лилия канадская (L. canadense): 15 - столон с замещающей луковицей
По способу распространения семян лилейные чаще относятся к баллистам, у которых семена распространяются посредством метания. Непременным условием для такого способа рассеивания семян является прямостоячий, упругий к моменту плодоношения стебель, который раскачивается от ударов ветра или каких-либо других агентов и выбрасывает семена, подобно снарядам из древних осадных орудий. Для многих лилейных - баллистов характерны резкие изменения положения цветоножек к моменту плодоношения, так называемые карпотропические изгибы, когда горизонтальные или загнутые вниз цветоножки изгибаются дугообразно или под прямым углом кверху, придавая коробочке строго вертикальное положение, что препятствует самопроизвольному высыпанию семян. Баллисты этого типа нередко имеют дисковидные семена с анемохорными приспособлениями. Они плоские, окружены крыловидной тонкой каймой и легко разносятся ветром (баллисты-анемохоры). У некоторых лилейных семена растаскивают муравьи. В стеблях или цветоносах у этих видов слабо развита механическая ткань. В отличие от баллистов, у которых стебли высыхают и деревенеют, стебли мирмекохоров при плодах слабеют и полегают. Семена этих растений имеют нежные и сочные придатки - элайосомы, образованные крупными паренхиматическими клетками, богатыми жирными маслами, которые легко отделяются от семян и поедаются муравьями.
Семейство лилейные состоит из двух резко очерченных подсемейств: собственно лилейные (Lilioideae) и пролесковые (Scilloideae). Пролесковые некоторые авторы, например Р. Дальгрен (1975, 1980), рассматривают как самостоятельное семейство гиацинтовые (Hyacinthaceae).
Подсемейство собственно лилейные (Lilioideae) имеет 10 родов (примерно 470 видов), относящихся к 4 трибам. Представители подсемейства встречаются только в северном полушарии. Луковицы у них составлены низовыми чешуями, так как базальные листья отсутствуют, за исключением рода кардиокринум и нескольких видов лилий. Цветоносный стебель облиственный. Сегменты околоцветника свободные, а нектарники тепальные. Пыльники прикреплены спинкой, качающиеся (триба лилейных) или прикреплены основанием, вращающиеся вокруг оси (трибы тюдьпановых и гейджиевых). Нити тычинок свободные. Семена обычно плоские. Из анатомических признаков характерно отсутствие угловых клеток эпидермы. Рафиды оксалата кальция отсутствуют. Характерно наличие алкалоидов, и по химическому составу эта группа гомогенная (К. Вильяме, 1975). Однородность группы подтверждается и серологическими данными (В. С. Чупов и Н. Г. Кутявина, 1980).
Триба лилейных (Lilieae) объединяет 5 родов, центром развития которых является Восточная Азия. Здесь целиком сосредоточены древние роды кардиокринум и номохарис (Nomocharis), а также наиболее древние представители родов лилия (Lilium), нотолирион (Notholirion) и рябчик. В Восточной Азии сохранились промежуточные звенья между родами - виды с признаками обоих родов. У представителей трибы лилейных луковицы составлены незамкнутыми низовыми (кроме родов кардиокринум и нотолирион) чешуями, специализированных покровных чешуй нет. Коробочка прямостоячая, семена плоские, дисковидные, с крыловидной каймой, распространяемые по способу баллистов.
Род кардиокринум (Cardiocrinum) насчитывает 3-4 вида, произрастающих в Гималаях, Китае и в Японии. Ранее его рассматривали как секцию или подрод рода лилия, однако совершенно отличный жизненный цикл, иное строение луковицы, черешковые сердцевидные листья с сетчатым жилкованием и несколько зигоморфные цветки подтверждают его родовую самостоятельность. Эти травы высотой 1,5-4 м являются монокарпиками, т. е. цветут и плодоносят единственный раз, после чего все растение отмирает. Луковица кардиокринума образована разросшимися основаниями черешков листьев; она не имеет покровных чешуй, находится почти на поверхности почвы и на свету приобретает темно-зеленый цвет. (рис. 38, 1). Когда начинает расти цветоносный стебель, чешуи луковицы, расходуя на него питательные вещества, постепенно высыхают, так что к моменту цветения луковицы уже не существует. У основания стебля остаются лишь дочерние луковички, образовавшиеся в пазухах чешуй материнской луковицы. Обыч-но листья у кардиокринумов только стеблевые, которые собраны в ложную мутовку; у кардиокринума гигантского имеется приземная розетка и стеблевые листья очередные. Листовые пластинки достигают в диаметре 30-45 см, длина черешков около 16 см. Цветки в числе от 4 до 24 на коротких цветоножках, горизонтальные, трубчатые, длиной 12-15 см, с опадающими прицветниками, белые, снаружи зеленоватые, ароматные. Коробочка по краям растрескавшихся гнезд имеет характерные для представителей трибы лилейных выросты эпидермы в виде зубчиков (рис. 38, 3). Семена снабжены широким (до 5 мм) крыловидным краем (рис. 38, 4). В роде, по мнению одних авторов, 3 вида, по мнению других - 4, так как кардиокринум Глена (С. glehnii), встречающийся на Сахалине и Курильских островах, часто присоединяют к японскому виду кардиокринум сердцевидный (С. cordatum). Аборигены острова Хоккайдо получают из луковиц этого растения пищевой крахмал. Все виды рода - высокодекоративные растения.
В роде лилия (Lilium) от 80 до 100 видов, широко распространенных в умеренной зоне северного полушария. Виды рода распространены на территории между 68° с. ш. (низовья реки Енисея) и 11° с. ш. (Южная Индия). Наиболее протяженный ареал (от Португалии до верховьев реки Лены в Сибири) у самого северного вида лилий - саранки, или лилии кудреватой (L. martagon), которая имеет несколько разновидностей с цветками различной окраски. Большинство видов встречается в предгорьях и горных районах, в лесах, в субальпийском высокотравье вдоль ручьев, на заболоченных местах или открытых травянистых склонах. Особенно богат видами лилий (около 40) Западный Китай, Юго-Восточный Тибет и Северная Бирма.
Луковицы лилий многолетние, черепитчатые (чешуи незамкнутые и узкие), составлены низовыми чешуями. Лишь у нескольких видов в луковице имеются еще и листовые чешуи. Чешуи цельные или членистые, обычно в числе 8-40, но иногда их может быть 100-120, как, например, в луковице лилии Кесселъринга (L.kesselringianum, табл. 6, 5), достигающей в диаметре 25 см и состоящей из 7 годичных циклов. По местонахождению почки возобновления и направлению роста побега у лилий различают ортотропные и плагиотропные луковицы (М. В. Баранова, 1981). У ортотропных луковиц почка возобновления расположена у основания цветоносного стебля, рост нового побега идет в вертикальном направлении и замещающая луковица формируется внутри материнской. У плагиотропных луковиц почка возобновления находится в пазухе нижележащих чешуй, рост нового побега идет в горизонтальном направлении и замещающая луковица располагается сбоку от материнской. Если плагиотропная луковица имеет несколько годичных по-бегов, она приобретает вид цепочки из нескольких луковиц, как, например, у лилии малой (L. parvum, рис. 39, 14) и некоторых других эндемичных видов западного побережья Северной Америки. Если же, кроме замещающей луковицы, имеются еще и дочерние, то образуется гнездо, которое с годами может увеличиваться до нескольких сотен луковиц, как, например, у лилии леопардовой (L. pardalinum). У некоторых видов замещающая луковица выносится за пределы материнской на толстом столоне, как у лилии канадской (L. canadense, рис. 39, 15) и других видов восточного побережья Северной Америки. От донца луковицы у лилий отходят многочисленные и сильные многолетние разветвленные корни. Некоторые из корней или их верхние участки являются втягивающими. Многие виды имеют корни на подземной части цветоносного стебля, над луковицей. Они дополнительно питают мощный цветущий стебель.

Таблица 6. Лилии и рябчики: 1 - лилия красивая (Lilium speciosum), оранжерея Ботанического института в Ленинграде; 2 - лилия двурядная (L. distichum), Дальний Восток; 3 - лилия Кессельринга (L. kesselringianum), хребет Кацихра, Абхазия; 4 - лилия однобратственная (L. monadelphum), парк Ботанического института АН СССР в Ленинграде; 5 - рябчик камчатский (Fritillaria camschatcensis); 6 - рябчик Северцова (F. sewerzowii), парк Ботанического института АН СССР в Ленинграде
Стебель у лилий обычно прямостоячий от основания, но у некоторых восточноазиатских видов он сначала как бы блуждает под землей и может появиться на поверхности на расстоянии 60 см от луковицы. Листья обычно стеблевые, в ложных мутовках или очередные, обычно без черешков, лишь у нескольких видов имеются базальные листья (лилия белоснежная - L. candidum и др.). Цветки горизонтальные и трубчатые (как у лилии длинноцветковой - L. logniflorum, рис. 39, 5), прямостоячие и кубковидные (как у лилии даурской, названной при первоописании по недоразумению лилией пенсильванской - L. pensylvanicum, рис. 39, 7), поникшие и чалмовидные (как у саранки, рис. 39, 7). На форме околоцветника и основана традиционная система рода. Сегменты околоцветника варьируют у различных видов по форме, размеру и окраске. У большинства лилий и наружные и внутренние сегменты имеют нектарники в виде желобка или канавки, которая может быть открытой и голой (лилия длинноцветковая), закрытой с двух сторон кожными складками (саранка, рис. 39, 8) или окаймленной сосочками (лилия Генри - L. henryi, табл. 4, 3). У лилии луковиценосной (L. bulbiferum), встречающейся в Центральной Европе, на концах листьев и у бутонов на кончиках внешних сегментов расположены примитивные экстрафлоральные нектарники, где весной выделяется сахаросодержащая жидкость, привлекающая муравьев (Е. Рицек, 1974). С наружной стороны сегментов у лилий имеются выросты центральной жилки, разной формы у различных видов (рис. 39, 6). В выемку между выростом жилки и спинкой внутренних сегментов плотно входят края наружных сегментов, в результате чего трубка околоцветника не распадается. Пыльники прикреплены к нитям спинками, качающиеся. Столбик значительно выше цилиндрической завязи. Рыльце головчатое, 3-лопастное.

Таблица 4. Лилейные. Ледебурия общественная (Ledebouria socialis): 1 - общий вид растения; 2 - цветок. Лилия Генри (Lilium henryi): 3 - фрагмент соцветия
Цветки лилий опыляются бабочками или мотыльками. Саранку, например, опыляют преимущественно ночные бабочки-бражники, а иногда также дневные бабочки. Приманкой для первых служит усиливающийся к ночи аромат цветков, для вторых - грязно-пурпуровый с темными пятнышками околоцветник. Нектарный желобок длиной 10-15 мм, с многочисленными мелкими выростами-сосочками имеет на наружном конце вход шириной 1 мм. Порхающий бражник, просовывая свой тонкий длинный хоботок в нектарник, касается брюшком, вымазанным пыльцой другого растения, сначала выдвинутого рыльца, а затем покрытых пыльцой пыльников. Так как во время цветения столбик у лилий изгибается вверх настолько, что рыльце соприкасается с двумя верхними тычинками, возможно и самоопыление цветков. У лилии белоснежной самоопыление исключено, так как столбик не изгибается и рыльце находится на 20-25 мм выше пыльников. Лилия малая, встречающаяся на альпийских лугах в горах Сьерра-Невады в Калифорнии, имеет характерные орнитофильные трубчатые, от оранжевых до темно-красных тонов цветки, опыляемые колибри. Однако колибри кормятся часто и на энтомофильных цветках, например лилии Гумбольдта (L. humboldtii). Стебель с коробочками у лилий имеет вид канделябра. Коробочки бескрылые или крылатые, с многочисленными дисковидными семенами в каждом гнезде.
Лилии хорошо размножаются и вегетативно: дочерними луковичками, которые образуются в пазухах чешуй материнской луковицы; чешуями или их члениками, а также луковичками-бульбочками, которые образуются на надземной части стебля в пазухах листьев (у лилии луковиценосной) или на подземной части стебля в зоне стеблевых корней.
Лилии - первоклассные декоративные растения, и начало их культуры восходит к далекому прошлому. Древнейшая из культивируемых лилий - лилия белоснежная, известная также как лилия Мадонны, лилия Благовещения, в диком состоянии встречается кое-где в Ливане, Палестине и Сирии на известняковых скалах, среди средиземноморской растительности. Ее разводили и до сих пор разводят из-за красивых и ароматных цветков и для получения эфирного масла и лекарственных препаратов. Первые изображения этой лилии встречаются на Критских вазах и фресках начиная с 1750 г. до н. э., а затем у древних ассирийцев, египтян, греков и римлян. Древние римляне считали, что после розы нет цветка более прекрасного, чем лилия. От древних римлян лилия попала в завоеванные ими страны Европы. В раннем средневековье ее выращивали монахи в монастырях сначала как лекарственное растение. Белизна ее цветков считалась символом правды, чистоты и непорочности, поэтому у ранних христиан цветки лилии белоснежной стали атрибутами девы Марии - богоматери. Начиная с эпохи итальянского Возрождения и ранней фламандской живописи в картинах на библейский сюжет "Благовещение" изображался архангел Гавриил, приносящий мадонне белую лилию. Карл Линней, давший ботаническое название этой лилии, следовал латинским поэтам, которые начиная с Вергилия (70- 19 гг. до н. э.) называли ее белоснежной. С XVI в. в культуру ввели и другие виды лилий. С конца XVIII в. начались работы по гибридизации. В начале XX в. с нахождением в Западном Китае лилии царственной (L. regale) значительно увеличивается число сортов лилий, достигшее теперь 2000. В Восточной Азии лилии разводят как овощную культуру, ради съедобных луковиц. Использовали в пищу и луковицы сибирских видов. В луковице лилии даурской обнаружены сердечные гликозиды. В народной медицине использовали и луковицы саранки, которая, согласно старой сибирской легенде, возникла из сердца казачьего атамана Ермака, погибшего в 1585 г. в бою с ханом Кучумом на реке Иртыш.
В роде нотолирион (Notholirion) 5 видов, которые встречаются в Восточной и Западной Азии на высокогорных лугах. Три из них приурочены к Восточной Азии. Нотолирион Томсона (N. thomsonianum) произрастает в сравнительно сухих областях, на солнечных склонах от Центральных Гималаев до Северо-Западного Афганистана. Самый западный вид - нотолирион Коэ (N. koeiei) - найден на горных болотах хребта Загрос в Западном Иране. Виды нотолириона - монокарпические растения, которые отличаются от лилий строением луковицы и отчетливо 3-раздельным рыльцем. Луковица нотолириона состоит из влагалищ базальных листьев и низовых чешуй, в пазухах которых образуются дочерние луковички, остающиеся в земле после отмирания растения. Покровными чешуями служат сухие основания листьев. Листья базальные и стеблевые. Цветки трубчатые или колокольчатые, иногда немного зигоморфные, протандричные (табл. 5,7). В основании сегментов околоцветника имеется нектарная ямка, как у видов рябчика. Семена мелкие, бескрылые.

Таблица 5. Лилейные. Нотолирион крупнолистный (Notholirion macrophyllum): 1 - соцветие. Номохарис Фаррера (Nomocharis farreri): 2 - соцветие; 3 - тычинка; 4 - наружный сегмент; 5 - внутренний сегмент (оба с внутренней стороны)
Представители рода номохарис (Nomocharis) встречаются на высокогорных лугах или пастбищах в Восточной Азии. В роде, по-видимому, около 6 видов. Ранее к роду номохарис присоединяли виды с признаками и лилий и рябчика, которые теперь относят к роду лилия (С. Лян, 1980). Луковицы у номохариса такого же строения, как и у лилий, но цветки совершенно плоские, блюдцевидные (табл. 5, 2). Сегменты околоцветника широко распростертые от основания, с пятнышками. Основания внутренних сегментов и тычиночные нити темноокрашенные и образуют заметный "глазок" в центре цветка. Наружные сегменты обычно без нектарников. В основании внутренних сегментов в середине темного пятна имеется короткий нектароносный желобок, от которого веерообразно расходятся складки ткани. Тычиночные нити с мясистым цилиндрическим или кеглевидным основанием и игловидной верхушкой. Столбик почти равен по высоте завяви.
Виды номохариса и нотолириона - декоративные растения.
Род рябчик, или фритиллария (Fritillaria), насчитывает около 100 видов, распространенных так же, как лилия, в северном полушарии. Часть видов, близких лилиям, встречается в лесах Восточной Азии. Большая часть наиболее подвинутых видов сосредоточена в Западной Азии. В отличие от остальных родов трибы, имеющих длительный период вегетации, рябчики - типичные эфемероиды. Луковицы их ежегодно возобновляются, составлены низовыми чешуями, покровных чешуй обычно не имеют. У представителей подрода лилиориза (Liliorhiza) луковицы черепитчатые, рыхлые, с многочисленными мелкими чешуями. У рябчика камчатского (Fritillaria camschatcensis, рис. 40, 7) и других лесных восточноазиатских видов этой секции чешуи сильно вздутые и похожи на зерна риса. Они легко отделяются от донца и укореняются, имитируя луковички-детки. На самом деле это разбухшие основания низовых чешуй, у которых верхняя часть осталась тонкой и отсохла, о чем свидетельствует рубец. Замещающая луковица у этих видов выносится наружу на толстом столоне. Втягивающих корней у них нет, и луковицы залегают у поверхности почвы. Подавляющее большинство рябчиков имеет луковицы, составленные 1-4 мясистыми широкими чешуями, причем у видов типового подрода луковицы полутуникатные, из 2-3 несросшихся или едва сросшихся чешуй, а у видов подрода петилиум (Petilium), подрода терезия (Theresia) и у рябчика Северцова (Fritillaria sewerzowii) луковица туникатная, из 1-4 массивных, сросшихся целиком или наполовину чешуй (М. В. Баранова, О. И. Захарьева, 1981). У этих растений аридных местообитаний луковица покрыта высохшими чешуями прошлых лет. Многочисленные втягивающие корни способны втянуть луковицу на глубину 25 см и таким образом защитить почку возобновления от высыхания. Замещающая луковица образуется внутри материнской. Стебли у рябчика с очередными или мутовчатыми листьями. Прицветные листья прямостоячие, иногда многочисленные (рябчик императорский - F. imperialis, табл. 7, 1), иногда спирально закрученные (рябчик русский - F. ruthenica, рис. 40, 8). Цветки одиночные или их несколько, иногда до 30 (рябчик персидский - F. persiса), собранные в кисть или зонтик (мутовка). Околоцветник актиноморфный или зигоморфный (подрод ринопеталум - Rhinopetalum), обычно колокольчатый, у некоторых видов с шахматным рисунком. На внутренней стороне сегментов в их основании имеются нектарные ямки (рис. 40, 9,10,14, 15), иногда у верхнего сегмента нектарник выпячивается наружу в виде особенно крупного шпорца (подрод ринопеталум, рис. 40, 13); встречаются и бороздчатые нектарники, как у лилий (рябчик камчатский, рис. 40, 2). Пыльники прикрепляются к нити основаниями или несколько выше основания.

Таблица 7. Рябчики и тюльпаны: 1 - рябчик императорский (Fritillaria imperialis), ботанический сад Ленинградского государственного университета; 2 - рябчик шахматный (F. meleagris), Закарпатье; 3 - рябчик кавказский (F. caucasica), г. Араилер, Армения; 4 - тюльпан Шренка (Tulipa schrenkii), Крым; 5 - тюльпан двухцветковый (Т. biflora), Крым; 6 - тюльпан Эйхлера (Т. eichleri), Азербайджан; 7 - тюльпан Биберштейна (Т. biebersteiniana), Воронежская область
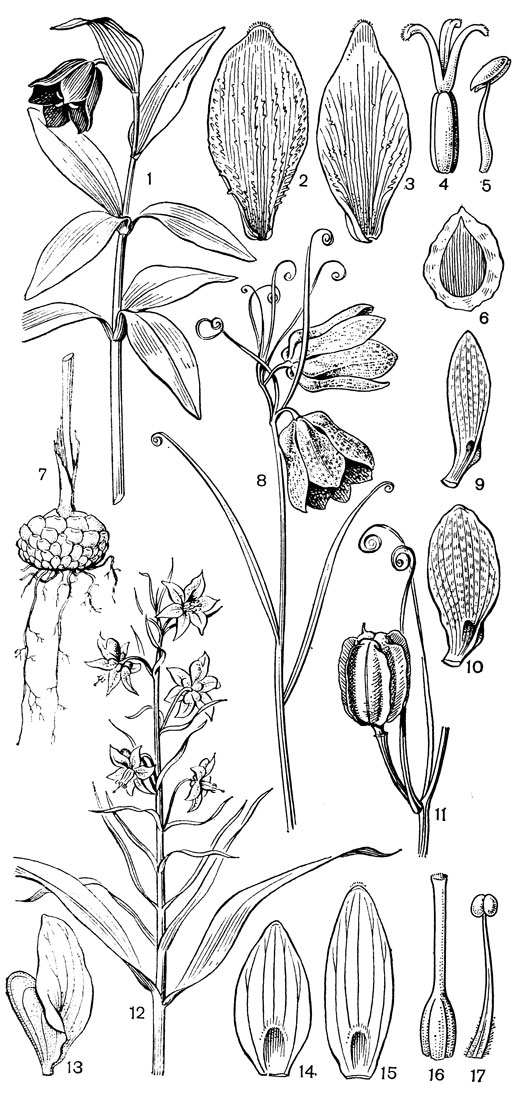
Рис. 40. Рябчики. Рябчик камчатский (Fritillaria camschatcensis): 1 - общий вид; 2 - внутренний сегмент; 3 - наружный сегмент; 4 - гинецей; 5 - тычинка; 6 - семя; 7 - луковица. Рябчик русский (F. ruthenica): 8 - соцветие с прицветными листьями; 9 - наружный сегмент; 10 - внутренний сегмент; 11 - коробочка. Рябчик Карелина (F. karelinii): 12 - соцветие; 13 - верхний наружный сегмент; 14 - боковой наружный сегмент; 15 - внутренний сегмент; 16 - гинецей; 17 - тычинка
Карта 1. Ареал рябчика камчатского
Цветки рябчика протогиничные. У рябчика камчатского и других американских видов цветки имеют неприятный запах и опыляются мухами. Восточносредиземноморские виды опыляются шмелями, осами и мелкими мухами. В Калифорнии наблюдается замечательный параллелизм в строении цветков рябчика отогнутого (F. recurva) и лилии малой. Оба вида имеют трубчатые багряно-красные цветки с большим количеством нектара, привлекающего опылителей-колибри. Рябчик императорский имеет признаки цветка, опыляемого птицами (Г. Куглер, 1970). У подавляющего большинства видов рода преобладает семенное размножение и лишь у столонообразующих восточноазиатских видов оно почти полностью подавлено и компенсируется вегетативным размножением с помощью большого числа (до 50) легко опадающих чешуй.
Рябчики используют как декоративные и лекарственные растения. Многие виды ядовиты, так как содержат алкалоиды. Несмотря на горечь, луковицы некоторых видов съедобны. Так, луковицы рябчика камчатского, встречающегося по обоим побережьям (азиатскому и американскому) Тихого океана, служили пищей камчадалам и индейцам. Камчадалы, называвшие растения "сараной", выкапывали луковицы мотыгой или забирали их из кладовых полевки-экономки (Microtus oeconomus). Сушеные чешуи луковиц, нанизанные на веревки, вывозили с Камчатки, где растения росли в изобилии, в Америку и продавали индейцам, которые называли их "северо-западным рисом".
Триба тюльпановых (Tulipeae), более подвинутая, чем триба лилейных, включает 2 рода, у представителей которых луковицы ежегодно возобновляются и составлены 1-5 низовыми замкнутыми чешуями.
Род тюльпан (Tulipa) насчитывает, вероятно, около 100 видов, но некоторые авторы доводят их число до 140 (З. M. Силина, 1977). Тюльпаны распространены в Евразии и Северной Африке, в областях с жарким и сухим летом и небольшим количеством осадков весной и осенью, в пустынях, полупустынях и степях, редко в лесу. Они растут на равнинах и во всех поясах гор, но редко вблизи ледников, на различных почвах: от плотного лесса до сыпучих песков, на нейтральных до засоленных, но не кислых почвах.
Луковицы тюльпанов залегают на глубине до 45 см и более. Они состоят из 2-5 мясистых запасающих чешуй, одетых специализированными покровными кожистыми или пленчатыми чешуями, которые с внутренней стороны бывают голыми, прижато-щетинистыми или шерстистовойлочными. У ювенильных (до первого цветения) растений ежегодно формирующиеся замещающие луковицы выносятся за пределы материнской луковицы и углубляются при помощи вертикальных (у некоторых видов горизонтальных) полых столонов, внутри которых они находятся (рис. 41, 5, 10). У некоторых видов сохраняется цепочка отмерших луковиц, которые иногда ошибочно принимают за "четковидные луковицы" (рис. 41, 8). Кроме замещающей луковицы, может образоваться 1-2 дочерних, при этом или одна ив них, или обе в столонах (рис. 41). Стебель тюльпанов с 2-4 (реже с 1 и до 12) листьями и с 1-2 (реже 3-15) прямостоячими цветками, хотя бутоны у некоторых видов бывают поникшие. Сегменты околоцветника без нектарников, опадают при плодах. С изменением температуры среды происходит движение сегментов (термонастии). При повышении температуры цветки широко раскрываются благодаря расширению клеток на внутренней стороне сегментов, при понижении температуры закрываются вследствие расширения клеток наружной стороны. Для тюльпанов характерна многоцветная окраска цветов (полихроизм). Так, например, даже в одной популяции степного тюльпана Шренка (Т. schrenkii) встречаются растения с цветками самой разной окраски - белой, желтой, розовой, красной, фиолетовой, с пятном при основании черным или желтым или без пятна. Тычинки часто с расширенными к основанию, иногда толстыми нитями, которые оканчиваются острием, входящим в углубление основания пыльника, отчего пыльники вращаются вокруг оси. Рыльце 3-лопастное, чаще сидячее; столбик более или менее длинный.
У тюльпанов ярко выражена протандрия. Цветки тюльпанов опыляются мелкими дчелами, мухами, собирающими пыльцу. Нектарники, подобные нектарникам безвременника, имеются лишь у тюльпана лесного (Т. sylvestris, Э. Дауманн, 1970). Весенние эфемероиды тюльпаны в период массового цветения покрывают открытые горные или степные склоны красными, желтыми, пестрыми коврами. Прямостоячие коробочки тюльпанов содержат многочисленные плоские, треугольные коричневые семена, распространяемые по способу баллистов, что характерно для растений открытых пространств. Некоторые виды размножаются вегетативно дочерними луковичками преимущественно в ювенильный период.
Тюльпаны - прекрасные декоративные растения. Первые культурные тюльпаны попали в Западную Европу в XVI в. из Константинополя. Их называли турецкими тюльпанами, прародителем их был тюльпан Шренка, привозимый из Кафы (нынешней Феодосии), входившей тогда в состав Османской империи (М. Хоог, 1973). Голландия стала страной промышленного выращивания культурных сортов тюльпанов. В начале XIX в. стало известно о существовании настоящего очага видового разнообразия тюльпанов в Средней Азии (около 75 видов). Заслуга в интродукции многих декоративных растений Средней Азии и тюльпанов в первую очередь принадлежит директору ботанического сада в Санкт-Петербурге Эдуарду Регелю. В настоящее время насчитывается 2900 промышленных сортов тюльпанов, объединенных в 15 классов; 3 класса из них составляют разновидности и гибриды замечательных среднеазиатских видов: тюльпана Кауфмана (Т. kanfmanniana), тюльпана Фостера (Т. fosteriana) и тюльпана Грейга (Т. greigii), который считается самым красивым тюльпаном в мире.
Род кандык, или эритрониум (Erythronium), насчитывает 24 вида, из которых 15 произрастают на западе, 5 - на востоке Северной Америки, а 4 вида - евразиатских. Эритрониумы - лесные горные растения, они встречаются от предгорий до альпийских лугов в хвойных, смешанных и лиственных лесах, реже на высокогорных лугах. Луковица кандыка сибирского (Е. sibiricum) и кандыка кавказского (Е. caucasicum) состоит из одной запасающей чешуи, образовавшейся из 3 сросшихся на ранних стадиях онтогенеза между собой и с цветоносом в одну замкнутую чешую (Н. И. Шорина и А. Г. Куклина, 1976). Покровные чешуи луковицы образуются из высохших запасающих чешуй. У основания луковицы кандыка сибирского сохраняются донца луковиц прошлых лет, образуя членистое (до 6 члеников) корневище (Г. В. Скакунов, 1974, рис. 41, 2). У ювенильных растений замещающая луковица углубляемся в почву при помощи шпорообразно удлиняющегося влагалища низового листа, обладающего положительным геотропизмом.
Эритрониумы - ранневесенние эфемероиды. Весной появляется стебель с 2-3 часто пятнистыми листьями и обычно одним поникшим розовым, лиловым, белым или желтым цветком. Сегменты околоцветника сближены в нижней части и образуют трубку, а в верхней отогнуты кверху, как у цикламена. В пасмурную погоду и с наступлением сумерек сегменты опускаются, предохраняя пыльцу от намокания. Нижняя часть сегментов обычно окрашена иначе, а на месте отгиба сегментов имеются различные метки: пятна, крапинки, штрихи. У некоторых видов в основании внутренних сегментов имеется поперечная, разделенная на 4 лопасти складка, которая продолжается с каждой стороны сегмента в виде ушковидных отростков (рис. 41, 4). Тычинки эритрониумов имеют подвижные пыльники, как у тюльпанов, и нити, иногда сильно уплощенные (кандык сибирский и кандык европейский, или собачий зуб,- Е. denscanis, рис. 41, 5); у некоторых видов тычинки обоих кругов разной длины (кандык японский - Е. japonicnm). Соотношение длины зрелого столбика и тычинок варьирует у разных видов.
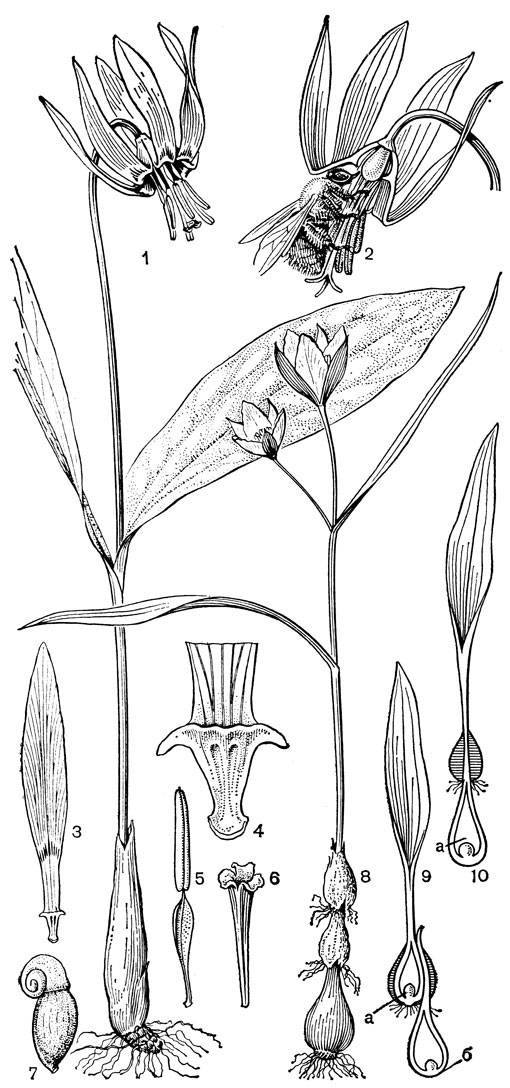
Рис. 41. Лилейные: триба тюльпановых. Кандык сибирский (Erythronium sibiricum): 1 - общий вид. Кандык японский (Е. japonicum): 2 - ксилокопа (Xylocopa appendiculata circumvolans), высасывающая нектар из цветка. Кандык сибирский: 3 - внутренний сегмент околоцветника; 4 - нижняя часть внутреннего сегмента с внутренней стороны с поперечной складкой, продолжающейся в ушковидные выросты; 5 - тычинка; 6 - столбик с рыльцем; 7 - семя с придатком. Тюльпан двуцветковый (Tulipa biflora): 8 - общий вид взрослого растения. Вегетативное размножение тюльпана (схема): 9, 10 - образование замещающей луковицы (а) и дочерней (б), разные варианты
Цветок эритрониумов имеет интересные приспособления к насекомоопылению, что особенно ярко выражено у близкого к кандыку сибирскому кандыка японского (Ф. Утех и С. Кавано, 1975). У этого растения нектар в большом количестве выделяется секреторной тканью в основании сегментов. Для того чтобы нектар не вытекал из опрокинутого околоцветника, существует система приспособлений, образующая как бы колпак вокруг завязи. Ниже лопастей складки, по краям основания сегментов имеются продольные вздутия, благодаря которым колпак вокруг завязи плотно прижат к ее стенкам и не позволяет вытекать нектару. Чтобы получить нектар, надо отстранить сегменты и нити тычинок от завязи, с чем успешно справляется ксилокопа - самая крупная пчела японской фауны перепончатокрылых. Лиловые отвернутые части сегментов околоцветника сильно отражают ультрафиолетовые лучи и разительно контрастируют с пурпурным зубчатым ободком в зеве трубки, таким заметным в видимом спектре. В сочетании эти свойства околоцветника указывают путь насекомым с трехцветным зрением. Действия пчелы точные и быстрые. Удерживаясь на тычинках и столбике, она головкой отстраняет сегменты околоцветника, сжимающие завязь, и открывает выход нектару, который высасывает своим хоботком (рис. 41, 2). В это время волосистая поверхность тела и лапок пчелы покрывается пыльцой. Эффект усиливается благодаря тому, что пыльники, открывающиеся двумя продольными щелями, вращаются на оси, а также благодаря различной длине тычиночных нитей, в результате чего образовавшаяся зона пыльников почти вдвое превышает длину одного пыльника. Цветки кандыка японского слабо протандричны. Лопасти рыльца раскрываются только тогда, когда длина рыльца превысит длину тычинок с созревшими пыльниками, что исключает самоопыление.
Коробочки прямостоячие, с небольшим числом семян, стебли при плодах полегающие. У евразиатских видов семена продолговато-яйцевидные, желтые, с хорошо выраженным бесцветным придатком в области халазы (рис. 41, 7). Придаток, содержащий жирные масла, поедают муравьи, которые способствуют распространению семян. По своей морфологической природе придаток представляет ариллоид, вырост наружного интегумента (Г. А. Комар, 1978). Основной способ размножения видов - семенной, некоторые размножаются вегетативно. Луковицы некоторых видов используют в пищу и как лекарственные средства. Все виды рода - ранневесенние декоративные растения.
Последней, наиболее подвинутой трибой подсемейства лилейных является триба гейджиевых (Gageeae). Это приоритетное название трибы (Г. Руи, 1910), ранее известной как триба ллойдиевых (Lloydieae, Ф. Вуксбаум, 1937). В трибе гейджиевых 2 рода, которые характеризуются ежегодно возобновляющейся луковицей, состоящей из одной замкнутой низовой чешуи, покрытой сухими влагалищами прошлогодних листьев.
Род гейджия, или гусиный лук (Gagea), насчитывает около 70 видов, распространенных в умеренных областях Евразии и Северной Африки, от лесотундры и ледников в горах до полупустынь. Гусиные луки - ранневесенние эфемероиды. Их желтые звездчатые цветки покрывают весной горные луга, щебнистые склоны и трещины скал, встречаются в степи, иногда на засоленной почве и на известняках, отдельными куртинами в лиственных лесах и на газонах в парках или, как сорняки, в посевах. Это самые мелкие растения в подсемействе лилейных - высотой от 3 до 35 см. Они имеют одну луковицу или 2-3 в том случае, если у материнской луковицы образуется 1 или 2 луковички-детки, сохраняющиеся при материнском растении. У основания материнской луковицы иногда за ряд лет скапливается множество мелких луковичек-деток, образующих гнездо разновозрастных луковиц. Покровные чешуи сетчато-волокнистые, пленчатые или кожистые. У некоторых видов подрода платиспермум (Рlаtyspermum), обитающих в сравнительно сухих местах, имеются корни двоякого рода: обычные нитевидные придаточные корни, идущие от середины донца луковицы вертикально вниз (положительный геотропизм), и жесткие склерифицированные корни по краям донца, идущие сначала вниз, а затем горизонтально и вверх (нулевой или отрицательный геотропизм, рис. 42, 1). Они оплетают луковицу и вместе с песчинками между ними образуют вокруг нее как бы капсулу, которая, возможно, защищает луковицу от высыхания в летний период. Базальные листья (1 или 2) обычно превышают соцветие, плоские, с килем или дудчатые (полые внутри). Подсоцветных листьев 1-3(5), реже стебель равномерно облиственный. Соцветия зонтиковидные, малоцветковые. Цветки часто на неравных цветоножках. Сегменты околоцветника снаружи зеленые, внутри желтые, в основании с нектарной ямкой. Тычинки так же, как у видов тюльпана и эритрониума, имеют пыльники, прикрепленные основаниями, подвижные (рис. 42, 2). Цветки слабопротогиничные, без запаха. Опылителей - маленьких мух, жуков и пчел - привлекает нектар, который скапливается в виде капель между основаниями сегментов и тычиночных нитей. Семена плоские, треугольные (подрод платиспермум) или яйцевидные (типовой подрод), причем у некоторых видов, например у гусиного лука желтого (G. lutea, табл. 8, 7) и гусиного лука сетчатого (G. reticulata), семена с элайосомами (рис. 42, 4). По своей морфологической природе это одна из разновидностей ариллоида - строфиола, т. е. разрастание ткани наружного интегумента семени вдоль семенного шва. Гусиные луки интенсивно размножаются и вегетативно с помощью луковичек, которые образуются на донце луковицы, в пазухах базальных или стеблевых листьев или иногда на месте бутонов. У одних видов усиленное вегетативное размножение происходит до первого цветения растения, у других способность к вегетативному размножению сохраняется и во взрослом состоянии.

Таблица 8. Лилейные: 1 - кандык сибирский (Erythronium sibiricum), парк Ботанического института АН СССР в Ленинграде; 2 - кандык кавказский (Е. caucasicum), там же; 3 - кандык европейский (Е. dens-canis), там же; 4 - тюльпан моголтавский (Tulipa mogoltavica); 5 - тюльпан Грейга (Т. greigii); 6 - тюльпан Фостера (Т. fosteriana), Узбекистан; 7 - гусиный лук желтый (Gagea lutea), Ленинградская область
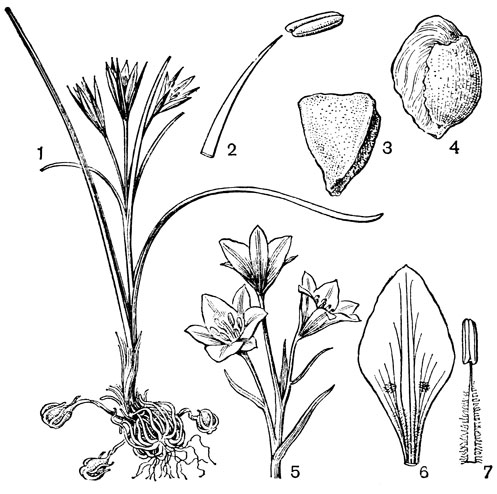
Рис. 42. Лилейные: триба гейджиевых. Гусиный лук переменчивый (Gagea commutata): 1 - общий вид растения, видны жесткие склерифицированные корни с отрицательным геотропизмом и луковички-детки ва столонах; 2 - тычинка, у которой остроконечие нити входит в углубление основания пыльника, в результате чего пыльник вращается вокруг оси; 3 - семя. Гусиный лук желтый (G. lutea): 4 - семя с придатком. Ллойдия тибетская (Lloydia tibetica): 5 - соцветие; 6 - внутренний сегмент околоцветника; 7 - тычинка
К роду гусиный лук близок род ллойдия (Lloydia), насчитывающий примерно 20 видов. Ллойдии - высокогорные растения (ореофиты). Большинство видов встречается в Восточной Азии (7 видов в Китае), несколько - в Восточном Средиземноморье. Ареал ллойдии поздней (L. serotina) охватывает территорию от гор Уэльса в Англии, горы Евразии до запада. Северной Америки и Арктическую провинцию. В. Грейтер (1975) склонен принимать род ллойдия как монотипный, с одним видом ллойдия поздняя. По облику ллойдии похожи на гусиные луки, но луковица у них продолговатая и окутана волокнистыми влагалищами прошлогодних листьев. У основания луковицы могут сохраняться донца прошлогодних луковиц, образуя членистое корневище. Цветки одиночные или по 2-3, воронковидные. Сегменты околоцветника беловатые, с пурпуровыми жилками и темными основаниями, опушенные по жилкам (ллойдия тибетская - L. tibetica, рис. 42, 5, 6), или с нектарниками (ллойдия поздняя), или без опушения и нектарников. Тычинки с подвижными пыльниками, как у гусиных луков. Нектарники ллойдии поздней представляют собой поперечную складку выше основания сегмента, над которой в углублении выделяется и задерживается нектар. Протандричные цветки ллойдии поздней со свободно лежащим нектаром опыляются короткохоботковыми насекомыми. Стебли при плодах не полегают. Коробочка растрескивается в верхней части гнезд, рассеивая плоские, полулунной формы семена по способу баллистов.
К подсемейству лилейных, возможно, относится и монотипная триба медеоловых (Medeoleae) с единственным родом и видом медеола вирджинская (Medeola virginiana). Медеола растет в лесах в восточной части Северной Америки и по облику похожа на триллиум (Trillium) или вороний глаз (Paris). Вот почему эти роды объединяли ранее в трибу парисовых (Parideae) или в семейство триллиевые (Trilliaceae), пока Р. Берг (1962) не установил, что медеолу гораздо правильнее отнести к подсемейству лилейных. У медеолы симподиальный, ежегодно замещающийся клубень (уникальный тип в пределах семейства). Надземный стебель имеет 2 ложные мутовки листьев и верхушечное соцветие - зонтик, обычно из 3 цветков на поникших цветоножках, которые становятся прямостоячими при плодах. Цветки мелкие, без запаха. Сегменты околоцветника одинаковые, зеленоватые, тычинки красноватые, завязь с 3 рыльцами. Пыльники находятся ниже широко раскинутых рылец и самоопыление не происходит. Плод - ягода; семена распространяются, по-видимому, птицами.
Подсемейство пролесковых (Scilloideae) включает 35 родов (около 825 видов), относящихся к 5 трибам. Представители подсемейства встречаются как в северном, так и в южном полушарии. Луковица у них составлена листовыми и низовыми чешуями с присутствием влагалищной чешуи у ежегодно возобновляющихся луковиц. Листья все базальные, и цветоносы, или стрелки, безлистные. Сегменты околоцветника свободные или сросшиеся в трубку. Нектарники септальные. Пыльники прикреплены спинкой, качающиеся. Нити тычинок свободные или сросшиеся. Семена обычно неплоские, за исключением родов дримия (Drimia) и дипкади (Dipcadi). В отличие от подсемейства лилейных имеются угловые клетки в эпидерме и рафиды. Наличие алкалоидов нехарактерно, и по химическому составу эта группа гетерогенная (К. Вильяме, 1975). Гетерогенность пролесковых подтверждается и серологическими данными (В. С. Чупов и Н. Г. Кутявина, 1981).
Довольно разнородная триба пролесковых (Scilleae) объединяет 11 родов и примерно 550 видов, распространенных в Африке и Евразии, преимущественно в Средиземноморье и Южной Африке. У видов трибы сегменты околоцветника свободные или сросшиеся у основания, реже сросшиеся в короткую трубку. Здесь представлены все формы луковичных лилейных: от круглогодично вегетирующих до эфемероидов, с луковицами многолетними до однолетних. Виды родов ледебурия (Ledebouria) и дримиопсис (Drimiopsis), встречающиеся в тропической области, вегетируют круглый год (т. е. листья появляются непрерывно в течение года), луковицы развиваются на корневищах, которые имеют многолетние ветвящиеся корни; контрактильные корни развиты слабо или не развиты совсем. В результате интенсивного вегетативного развития на разветвленном корневище образуется большое гнездо разновозрастных луковиц. Луковицы находятся у поверхности почвы и составлены только основаниями базальных листьев (М. В. Баранова, 1976). Виды рода дримия, радамантус (Rhadamanthus), некоторые виды альбуки (Albuca) и виды подрода просперо (Prospero) рода пролеска (Scilla) имеют продолжительную вегетацию с небольшим периодом покоя в наземном развитии, после которого у них сначала появляются цветоносы, а затем листья, причем цветонос принадлежит почке настоящего года, а листья - почке следующего года. Цветонос выходит из пазухи низовой или влагалищной чешуи и находится сбоку от появляющегося позднее пучка листьев. Среди видов родов пролески и птицемлечника (Ornithogalum), приуроченных к умеренным областям, встречаются зимнезеленые виды, гемиэфемероиды и эфемероиды. У последних цветки появляются одновременно с листьями; во всех трех случаях цветки и листья принадлежат одной почке и цветонос выходит из пазухи листовой чешуи, т. е. находится в центре пучка листьев. Луковицы обычно многолетние, составлены листовыми и низовыми чешуями. Всасывающие корни однолетние, не ветвящиеся. Втягивающие корни хорошо развиты (рис. 43, 6).
Род пролеска (Scilla) насчитывает, вероятно около 50 видов, распространенных в Евразии и Средиземноморье, а также 4 вида в Южной Африке. Около 20 видов, приводимых в различных тропических флорах Африки и Мадагаскара, возможно, относятся к роду ледебурия. Южноафриканские виды имеют многолетние луковицы с многолетними ветвящимися корнями, двойные прицветники, иногда фиолетовые снизу листья, сравнительно мелкие цветки. Средиземноморские виды с длительным периодом вегетации представляют собой обычно крупные многоцветковые растения, зацветающие в начале лета, после появления листьев" или в конце лета, перед появлением листьев (пролеска осенняя S. autumnalis). Евразиатские виды - преимущественно эфемероиды встречающиеся в лесах и на субальпийских лугах, где обычно доминируют в ранневесеннем аспекте. Сегменты околоцветника у пролесок свободные до основания, внизу часто сложены в трубку, а выше распростертые, обычно голубые, синие, фиолетовые или почти белые с темной центральной жилкой. Тычинки свободные, с узкими нитями. Цветки гомогамные или протогиничные. У пролески двулистной (S. bifolia) тычинки находятся на одном уровне с одновременно созревающим рыльцем, но достаточна далеко от него, так что самоопыления не происходит. В период цветения пролески интенсивно посещаются различными пчелиными.
У одних видов коробочки сухие, на прямостоячих цветоносах и рассеивание почти шаровидных или эллипсоидных семян происходит по способу баллистов. У эфемероидных видов подрода пролеска (Scilla) коробочки мясистые, на полетающих цветоносах, с семенами, имеющими масляные придатки - элайосомы. У видов пролески они отграничены от семени и легко отделяются. Известно около 14 видов, у которых семена имеют придатки, различные по происхождению, форме и цвету (Е. В. Мордак, 1970; Ф. Шпета, 1971,1980; Г. А. Комар, 1973, 1978). Придатки пролесок представляют собой ариллоид - разрастание ткани наружного интегумента. У пролески двулистной (табл. 9, 7) и у видов родства пролески сибирской (S. sibirica), в том числе и у пролески Розена (S. rosenii, табл. 9, 2), разрастание происходит в области семявхода и возникает одна из разновидностей ариллоида - карункула (рис. 43, 8). У атропатенского вида пролески Мищенко (S. mischtschenkoana, табл. 9, 3), встречающегося в трещинах и в нишах под навесом скал, это разрастание происходит в области семенного шва и возникает другая разновидность ариллоида - строфиоль (рис. 43, 9). Обычно придатки бесцветные, но иногда они окрашены, как семя. У турецкого вида пролески темной(S. melaina) - черная строфиоль (рис. 43,10), а у среднеазиатской пролески пушкиниевидной (S. puschkinioides) - черная карункула. Многие виды пролесок являются декоративными растениями. Некоторые виды имеют алкалоиды. Так, в луковицах средиземноморского вида пролески осенней содержатся вещества, подобные веществам дримии морской (Drimia maritima).
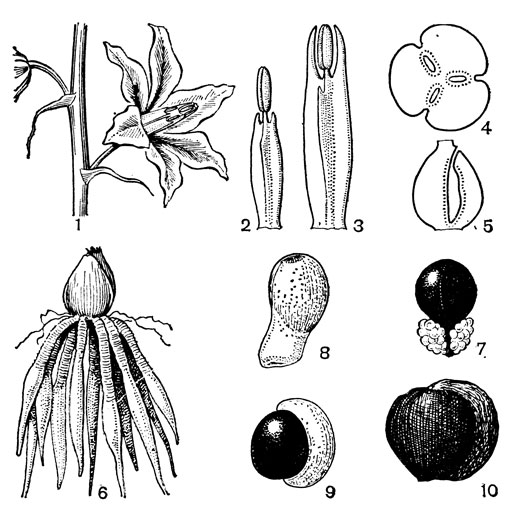
Рис. 43. Лилейные: триба пролесковых. Птицемлечник Буше (Ornithogalum boucheanum): 1 - цветок; 2 - тычинка наружного круга; 3 - тычинка внутреннего круга. Септальные нектарники (схема): 4 - поперечный срез завязи с нектарниками; 5 - продольный срез завязи через нектарник. Пролеска двулистная (Scilla bifolia): 6 - луковица со втягивающими (контрактильными) корнями; 7 - семя с придатком. Пролеска Розена (S. rosenii): 8 - семя с придатком. Пролеска Мищенко (S. mischtschenkoana): 9 - семя с придатком. Пролеска темная (S. melaina): 10 - семя с придатком

Таблица 9. Пролески и птицемлечники: 1 - птицемлечник понтийский (Ornithogalum ponticum),перевал Джубга, Западный Кавказ; 2 - пролеска Розена (Scilla rozenii), парк Ботанического института АН СССР в Ленинграде; 3 - пролеска Мищенко (S. mischtschenkoana), там же; 4 - пролеска двулистная (S. bifolia), Крым; 5 - птицемлечник Гуссона (О. gussonei), окрестности Еревана, Армения; 6 - пролеска сибирская (S. sibirica), парк Ботанического института АН СССР в Ленинграде; 7 - птицемлечник пирамидальный (О. thyrsoides), Крым, Никитский ботанический сад
Род ледебурия (Ledebouria), ранее считавшийся подродом пролески, восстановлен Дж. Джессопом (1970) и насчитывает около 30 видов, распространенных почти исключительно в Африке. Из них 15 видов приурочены к Южной Африке, где они встречаются в различных местообитаниях: лесных, травянистых, болотистых, сухих и среди скал. Из 15 видов, распространенных в тропической Африке, 1 вид - ледебурия гиацинтовая (L. hyancinthina) - встречается еще в Индии и на острове Шри-Ланка. В отличие от подавляющего большинства пролесок у ледебурий крупные луковицы, сочные, заметные, пятнистые, часто пурпуровые снизу листья и мелкие невзрачные цветки, как у капского вида ледебурии общественной (L. socialis, табл. 4, 1, 2). Сегменты околоцветника почти свободные до основания, где они образует мелкую чашу, концы их распростертые или отогнутые; тычинки свободные, завязь коническая, на ножке.
Близкий ледебурии род дримиопсис (Drimiopsis) включает около 15 видов, встречающихся в тропической и Южной Африке в разнообразных местообитаниях. Многие виды дримиопсиса имеют сердцевидные листья на длинных черешках, поэтому луковицы у них составлены черепитчатыми узкими чешуями - преобразованными влагалищами черешков. Цветки мелкие, многочисленные, на очень коротких цветоножках или сидячие, околоцветник зеленый, белый, розовый или пурпурный; сегменты свободные, сложены в трубку с немного отклоненными верхушками. Семена полушаровидные. Некоторые виды содержат алкалоиды типа наперстянки.
Род эвкомис (Eucomis) насчитывает 10 видов, распространенных от юга тропической Африки до Южной Африки. Эвкомисы встречаются во влажных местообитаниях. Это растения с розеткой крупных длительно вегетирующих листьев и многоцветковой цилиндрической кистью, увенчанной хохолком из прицветных зеленых листьев. Сегменты околоцветника сросшиеся у основания, обычно белые, кремовые или зеленые. Нити тычинок расширенные и сросшиеся у основания, образуют мелкую чашу. У эвкомиса двуцветного (Е. bicolor) околоцветник зеленоватый, а сросшиеся внизу нити тычинок пурпуровые, что создает впечатление яркого венчика на фоне зеленой чашечки. Цветки имеют зловонносладковатый запах и обильный нектар, выделяемый септальными нектарниками и скапливающийся у основания завязи. Цветки посещают мухи, прежде всего падальные, для которых приманкой на дальнем расстоянии служит запах, а на ближнем - окраска тычинок (А. Пашер, 1959). Коробочки сухие и жесткие или вздутые, пленчатые и легкие, как у велътгеймии (Veltheimia, см. рис. 45). Семена почти шаровидные.
Большой и гетерогенный род птицемлечник (Ornithogalum), состоящий из 6 подродов, насчитывает около 200 видов, распространенных в Африке, Евразии и Южной Америке (1 вид), но преимущественно в Средиземноморье и Южной Африке. По составу белков семян этот род близок родам эвкомис, гальтония (Galtonia) и дипкади. У птицемлечников цветки мелкие до крупных, без запаха, от нескольких до 100 и более, в цилиндрической или щитковидной кисти. Сегменты околоцветника белые, желтые, зеленоватые, свободные, редко сросшиеся внизу; тычинки свободные, нити их плоские или расширенные, с двумя зубцами наверху, как у птицемлечника Буше (О. boucheanum, рис. 43, 2, 3). Столбика нет, или он вальковатый, нитевидный; рыльце маленькое, головчатое. Семена обратнояйцевидные, угловатые или дисковидные. У некоторых видов в оболочке семени содержатся жирные масла. В отличие от элайосом пролески это недифференцированная элайосома, или саркотеста (Г. А. Комар, 1978). Многие виды птицемлечников ядовиты. У некоторых были обнаружены гликозиды. Птицемлечники очень декоративны. Срезанные цветоносы в бутонах южноафриканского птицемлечника пирамидального (О. thyrsoides, табл. 9, 7), у которого завязь черного цвета, отправляют в Англию.
Три вида, составляющие южноафриканский род неопатерсония (Neopatersonia), отличаются от птицемлечников в основном рыльцем с 3 расходящимися ветвями.
Род альбука (Albuca), имеющий примерно 100 видов, распространен в Африке и в Аравии, причем примерно половина видов встречается в Южной Африке. Это высокие (до 1 м) растения, у которых листья опадают или иногда развиваются после цветения. Сегменты околоцветника свободные, белые или желтые, с широкой зеленой или коричневой полоской посередине; 3 наружных сегмента распростертые, 3 внутренних - сближены и заключают тычинки и гинецей. Тычинки все или только 3 наружные фертильные; нити их внизу расширены и охватывают завязь (рис. 44, 5, 7). Цветки иногда приятно пахнут. Семена плоские, черные.
Род дримия (Drimia) с присоединенными к нему родами ургинея (Urginea), турантос (Thuranthos) и ургинеопсис (Urgineopsis) насчитывает около 120 видов, распространенных в Африке, Южной Европе и Азии (Дж. Джессоп, 1977; У. Стерн, 1978), но преимущественно, по-видимому, в Южной Африке. В Средиземноморье 6 видов, из которых дримия морская (D. maritima), известная с древнейших времен как лекарственное растение, имеет самое широкое распространение по морским побережьям всей Средиземноморской области. В Индии известны 3 вида. Один из них - дримия индийская (D. indica) - повсеместно встречается в Индии, а также в сосновых лесах Непала и диптерокарповых лесах Бирмы, в тропической и Южной Африке. В Южной Африке одни и те же виды встречаются в сухих и влажных местообитаниях. В тропической Западной Африке виды дримии растут в саваннах и полуаридных растительных зонах, часто вместе с видами близкого рода альбука. В Южной Африке и в Средиземноморье дримии встречаются вместе с видами птицемлечника.
У дримии луковицы наземные или подземные, иногда очень крупные, с диаметром до 20 см (дримия высочайшая - D. altissima), с рыхлыми, иногда черепитчатыми чешуями, у некоторых видов красноватыми. Дримия хавортиевидная (D. haworthioides), встречающаяся в узком районе восточной части Капской области, имеет наземную луковицу, подобную луковице радамантуса однобокого (Rhadamanthus secundus, рис. 44, 1). У листьев этих видов, образовавшихся осенью - зимой, к лету листовая пластинка отсыхает, их подземные части - влагалища - удлиняются, а верхушки разбухают и превращаются в запасающие органы. После того как листья отсохли, в наземном развитии дримий наступает период покоя, а летом - осенью появляется цветонос. У дримии высочайшей он достигает в высоту 1,5 м.
Цветки у дримии мелкие. Прицветники с хвостатыми отростками, иногда крупными и окрашенными, рано опадающими или остающимися. Сегменты околоцветника свободные и раскрытые или сросшиеся внизу в трубку и отогнутые, белые, розовато-лиловые, коричневые или зеленые, с темной центральной жилкой. Тычинки свободные. У некоторых видов цветки открываются вечером и ночью и имеют запах. Бледно-коричневые цветки дримии индийской открываются ночью и издают неприятный запах. Желто-коричневые цветки с белыми просвечивающими краями у дримии крупноцветной (D. macrantha), похожие по форме на цветки альбуки, открываются на одну ночь и имеют приятный запах. Коробочки округлоугловатые. Высокие цветоносы растений, встречающихся на больших открытых пространствах, плоские семена с крыловидной каймой - все указывает на то, что это - баллисты-анемохоры. Крупные луковицы дримии морской (народное название "морской лук"), диаметром до 16 см, часто находятся почти на поверхности и осенью образуют крупные ремневидные листья, которые к лету высыхают. К концу лета из сухой земли появляется 1,5-метровый цветонос с многочисленными белыми звездчатыми цветками. Это одно из самых древних медицинских и священных растений. В Древнем Египте его использовали во время жертвоприношений, а в Древней Греции считали, что его луковицы отгоняют злых духов. Со времен Гиппократа (460-377 гг. до н. э.) началось использование дримии морской как лекарственного растения, которое продолжается и по сей день. В ней содержатся сердечные гликозиды, и это было самое эффективное сердечное средство, пока во второй половине XVIII в. не была введена в употребление наперстянка.
Близкий роду дримия род радамантус (Rhadamanthus) насчитывает 9 видов, встречающихся в аридных районах Южной и Юго-Западной Африки и в горных районах Капской области. Так же, как дримии, виды радамантуса имеют луковицы с плотными или рыхлыми чешуями, листья, появляющиеся после цветения, прицветники со шпорцем, сегменты околоцветника от свободных до сросшихся до середины, семена дисковидные, черные. Примечателен радамантус однобокий (R. secnndns), произрастающий на самом юге Юго-Западной Африки, в прибрежной горной пустыне Намиб. У него чешуи луковицы образуют рыхлую розетку, как у дримии хавортиевидной (рис. 44, 1). Характерной особенностью в роде является способ вскрывания пыльников неполной продольной щелью (рис. 44, 3).
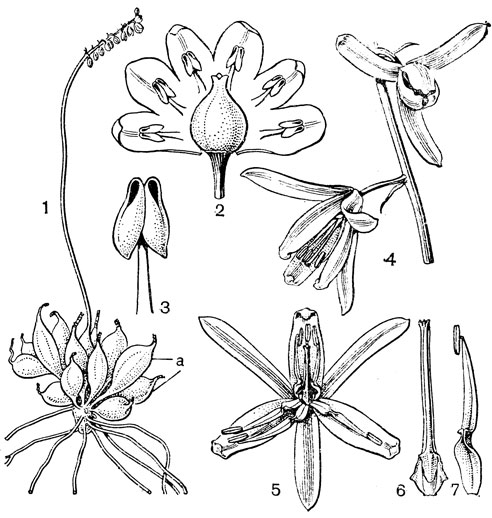
Рис. 44. Лилейные: триба пролесковых. Радамантус однобокий (Rhadamanthus secundus): 1 - общий вид (а чешуи луковицы); 2 - цветок в разрезе; 3 - тычинка. Альбука ангольская (Albuca angolensis): 4 - фрагмент соцветия; 5 - расправленный цветок, видны неправильно расположенные тычинки; 6 - гинецей; 7 - тычинка
Монотипный род уайтхедия (Whiteheadia) с одним видом уайтхедия двулистная (W. bifolia) встречается в сухих районах на западе Капской области и на самом юге Юго-Западной Африки. Это маленький наскальный геофит с розеткой из двух распластанных округлых листьев, как у видов массонии (Massonia), и невысоким многоцветковым цветоносом с зеленоватыми цветками. Сегменты околоцветника немного сросшиеся у основания; нити тычинок внизу расширенные и сросшиеся. Коробочки с 3 пленчатыми прозрачными крыльями. Семена черные, блестящие.
Род гиацинтоидес (Hyacinthoides) насчитывает примерно 7 видов, распространенных в Западной Европе и Западном Средиземноморье. Луковица гиацинтоидеса ежегодно возобновляется и состоит из одной замкнутой запасающей сочной чешуи, образовавшейся из нескольких (до 5) сросшихся влагалищ базальных листьев, а также пленчатой бесцветной влагалищной чешуи, которая охватывает основание зеленых листьев. Прицветников 2, крупных. Сегмента околоцветника свободные, колокольчатые или распростертые (гиацинтоидес итальянский - Н. italica), чаще голубые. Распространенный в Западной Европе до Северной Шотландии в лесах, в кустарниковых зарослях и вересковых пустошах гиацинтоидес неописанный (Н. nonscripta) имеет поникшие цветки и прямостоячие коробочки, которые рассеивают семена по способу баллистов.
Триба гиацинтовых (Hyacintheae) объединяет 18 родов (примерно 250 видов), часть из которых (8 родов) приурочены к Европе, Западной Азии и Северной Африке, преимущественно к Средиземноморской и Ирано-Туранской областям, другая часть родов ограничена в своем распространении Африкой, преимущественно Южной (8 родов); род родокодон (Rhodocodon) - эндемик Мадагаскара, дипкади (Dipcadi) - афро-индийский род. У всех представителей трибы сегменты околоцветника в большей или меньшей степени сросшиеся в трубку. Луковицы чаще многолетние, составлены основаниями базальных листьев и низовыми чешуями, чешуи чаще незамкнутые, реже луковицы ежегодно возобновляющиеся, у которых, кроме листовых и низовых чешуй, имеется еще влагалищная чешуя, чешуи замкнутые или незамкнутые. В большинстве случаев это эфемероиды.
Наше знакомство с родами первой группы начнем с рода гиацинт (Hyacinthus). В роде 3 вида (К. Персон и П. Венделбу, 1979), из которых гиацинт восточный (Н. orientalis) встречается в Юго-Западной Азии, а 2 других вида - гиацинт Литвинова (Н. litwinowii, табл. 10, 1) и гиацинт закаспийский (Н. transcaspicus) имеют ограниченное распространение в Северо-Восточном Иране и Южном Таджикистане (хребты Копетдаг, Восточный Эльбурс и Горган) до высоты 1700 м над уровнем моря. Луковицы у гиацинта многолетние, составлены незамкнутыми листовыми и низовыми чешуями. Сегменты околоцветника почти равны по длине трубке, серповидно изогнутые или отклоненные, голубые. Тычинки прикреплены примерно к середине трубки, однорядные. Плод - почти шаровидная мясистая коробочка. У гиацинта восточного семена шаровидные, черные, с крупным белым придатком - элайосомой, благодаря которой семена распространяются муравьями. Придаток семени гиацинта восточного, по-видимому, подобен по своему происхождению придатку семени пролески сибирской. Гиацинт восточный является родоначальником всех культурных сортов. Европейцы впервые увидели культивируемые гиацинты, так же как и тюльпаны, в XVI в. в садах турецких султанов в Константинополе. И, так же как в случае с тюльпанами, Голландия стала страной их массового выращивания. В природе у гиацинта восточного в кисти не более 10 цветков, они голубые и приятно пахнут. К настоящему времени получено множество сортов гиацинта, среди которых имеются многоцветковые (до 100 цветков), махровые сорта и сорта различной окраски. Название растения связано с древнегреческим мифом о прекрасном юноше Гиацинте: он был случайно убит Аполлоном при метании диска, и на месте его гибели вырос прекрасный цветок.

Таблица 10. Триба гиацинтовые: 1 - гиацинт Литвинова (Hyacinthus litwinowii), Западный Копетдаг, Туркмения; 2 - бельвалия сарматская (Bellevalia sarmatica), Ласпи, Крым; 3 - мускари незамеченный (Muscari neglectum), Крым, Никитский ботанический сад; 4 - хионодокса Люсили (Chionodoxa luciliae), парк Ботанического института АН СССР в Ленинграде; 5 - хионодокса сардыская (С. sardensis), там же; 6 - пушкиния пролесковидная (Puschkinia scilloides), там же; 7 - лашеналия золотистая (Lachenalia aurea), оранжереи Ботанической' института АН СССР в Ленинграде
Близкий гиацинту род белъвалия (Bellevalia) включает около 50 видов, распространенных от Западного Средиземноморья до Ирана и Средней Азии. Они встречаются на травянистых степных склонах, на полях и в посевах до нижнегорного пояса или у тающих снегов в субальпийском поясе. Листья у бельвалий сравнительно широкие, часто с белым хрящеватым краем и реснитчатые. Кисти цилиндрические (белъвалия Липского - В. lipskyi) или конические (белъвалия сарматская - В. sarmatica, табл. 10, 2). Верхние цветки бывают бесплодными. Сегменты околоцветника короче трубки, незначительно отклонены, часто на верхних сегментах имеются гребневидные вздутия, отчего околоцветник становится зигоморфным. Тычинки прикреплены у оснований сегментов, однорядные, нити их расширенные и сросшиеся у основания; пыльники находятся в зеве или на уровне верхушек сегментов. Окраска цветков меняется во время цветения от беловатой или синей (бутоны) до бурой, но есть и виды с синими цветками. Околоцветник при разбухании завязи целиком отделяется от основания и сбрасывается как колпачок, образующаяся коробочка с 3 ребрами - это признаки, присущие также и роду мускари. Семена более или менее шаровидные, гладкие, черные, с восковым налетом. Как и у мускари, стебель при плодах не полегает и распространение семян происходит по способу баллистов.
Около 20 видов, распространенных в Восточном Средиземноморье, Южной Европе до Южного Закавказья и Северного Ирана, имеет род гиацинтелла (Hyacinthella). Он отличается от бельвалии тем, что растения имеют 2 (редко 1 или 3) листа с выпуклыми жилкам. Цветки голубые, синие, не меняют своей окраски. Тычинки неясно двурядные, уровень пыльников варьирует у разных видов. Околоцветник разрывается продольно и остается при плодах в виде свободных сегментов. Семена сетчато-морщинистые. К роду гиацинтелла присоединен южнозакавказский вид гиацинтелла атропатенская (Н. atropatana), которую ранее относили к роду пролеска; она внешне идентична североиранскому виду гиацинтелла персидская (Н. persica), но имеет, в отличие от остальных видов рода, свободные сегменты, не сросшиеся в трубку (П. Венделбу, К. Персон, 1981).
Морфологически близок бельвалии и гиацинтелле ирано-туранский род алъравия (Alrawia), 2 вида которого встречаются в Северном Ираке и Западном Иране в разреженных дубовых или фисташковых лесах.
Гетерогенный род мускари, или мышиный гиацинт (Muscari), насчитывает около 30 видов (Д. Стюарт, 1970), распространенных в Европе, Северной Африке и Западной Азии до Средней Азии, но преимущественно в Средиземноморье, на травянистых склонах, в горах, в лесном поясе и у тающих снегов. Кисти плотные (мускари незамеченный - М. neglectum, табл. 10, 3) или рыхлые (мускари хохолковый - М. соmоsum). Верхние бесплодные цветки более или менее отличаются от плодущих. Околоцветник плодущих цветков актиноморфный или зигоморфный, от почти шаровидного до цилиндрического, стянутый у зева, с 6 отогнутыми зубчиками, обычно голубой или синий, с белыми зубчиками, реже коричневый, зеленоватый, желтый или белый. Тычинки в 1 или 2 кругах, прикреплены к середине трубки и заключены в ней. Семена, как у гиацинтеллы. Мускари опыляются пчелами или мелкими двукрылыми. Для привлечения насекомых у некоторых видов с невзрачными коричневато-зеленоватыми плодущими цветками имеются крупные стерильные верхние сине-фиолетовые цветки на длинных окрашенных цветоножках, образующие хохолок на вершину кисти. Плодущие цветки мускари гомогамные или слабопротогиничные. Многие виды мускари культивируют как декоративные растения и некоторые виды натурализовались в Европе, Северной Америке и Австралии. С XVI в. культивируется мускари мускусный (М. moschatum), имеющий сильный аромат.
Средиземноморский род бримёра (Brimeura) имеет 2 вида, у которых луковица возобновляется ежегодно и колокольчатые голубые или лиловые цветки имеют длинные прицветники.
По строению околоцветника (наличию трубки) и по серологическим данным (В. С. Чупов и Н. Г. Кутявина, 1981) близки кругу родства гиацинт - мускари, а также роду пролеска два рода - пушкиния (Puschkinia) и хионодокса (Chionodoxa). Род пушкиния с одним видом пушкиния пролесковидная (P. scilloides, табл. 10, 6) обитает в Западной Азии в среднегорном поясе до высоты 3000 м. Окраска цветков у нее варьирует от кремового до почти белого или голубого цвета с темной жилкой посередине сегментов. Тычинки прикреплены к разделенной на лопасти коронке или привенчику, образованному между тычинками и околоцветником (рис. 45, 1, 2). Семена, подобно семенам некоторых птицемлечников, не имеют отграниченных масляных придатков, у них масла, привлекающие муравьев, содержатся в наружной оболочке. У восточносредиземноморского рода хионодокса, насчитывающего примерно 6 видов, которые встречаются в горах у снежных пятен, нити тычинок шире, чем сами пыльники, и сложены в виде конуса вокруг столбика, как у птицемлечника Буше (рис. 45, 4-7). Белые нити и желтые пыльники образуют светлый глазок у ярко-синего колесовидного околоцветника хионодоксы сардыской (С. sardensis, табл. 10, 5). У этого вида и у хионодоксы Люсили (С. luciliae, табл. 10, 4) с голубым околоцветником известны формы с розовыми и белыми цветками. Хионодоксы широко культивируют как декоративные растения. Семена обоих видов имеют придатки одинаковой формы, но отличные по размеру (рис. 45, 8). Придаток хионодоксы похож по форме на придаток пролески двулистной, но представляет собой ариллус, образовавшийся в области семяножки (Г. А. Комар, 1974, 1978).
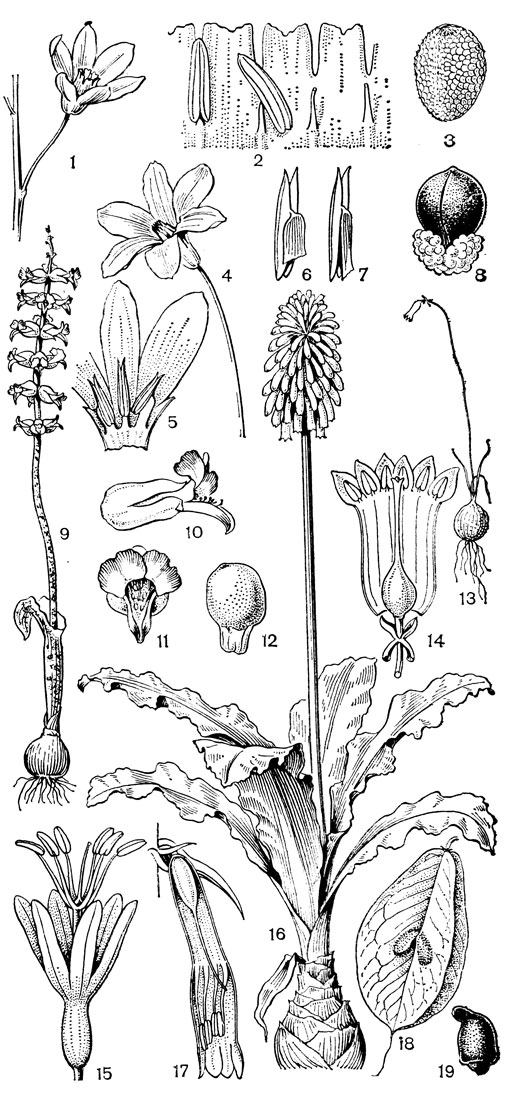
Рис. 45. Лилейные: триба гиацинтовых. Пушкиния про лесков и дная (Puschkinia scilloides): 1 - цветок; 2 - коронка с тычинками; 3 - семя. Xионодокса Люсили (Chionodoxa luciliae): 4 - цветок 5 - часть околоцветника с тычинками; 6, 7 -диморфные тычинки (видна тычиночная нить); 8 - семя. Лашеналия мутовчатая (Lachenalia verticillata): 9 - общий вид; 10 - цветок, вид сбоку; 11 - цветок, вид спереди; 12 - семя. Литантус крохотный (Litanthus pusillus): 13 - общий вид; 14 - цветок в разрезе. Андросифон капский (Androsiphon capensis): 15 - цветок. Вельтгеймия прицветниковая (Veltheimia bracteata): 16 - общий вид; 17 - цветок в разрезе; 18 - плод; 19 - семя
Перейдем теперь к группе африканских родов. Род лашеналия (Lachenalia) насчитывает около 90 видов (В. Баркер, 1978), произрастающих в Южной и Юго-Западной Африке, преимущественно в Капской области. Это некрупные растения с ежегодно возобновляющейся луковицей, имеющей влагалищную чешую и составленной замкнутыми чешуями у широколистных видов в незамкнутыми - у узколистных видов (М. В. Баранова, 1976). Листья от одного до многих, часто трубчатые в основании и пятнистые (рис. 45, 9). Околоцветник обычно зигоморфный, сегменты образуют трубку; они белые, желтые, красные, синие или зеленые. Внутренние сегменты равны наружным или длиннее их: у наружных сегментов на кончиках обычно темноокрашенные вздутия или пятнышки (табл. 10, 7). Некоторые виды опыляются дневными бабочками, а лашеналия трехцветная (L. tricolor) с поникшими длиннотрубчатыми цветками, у которых наружные сегменты красные, а внутренние длинные сегменты желтые, с зелеными кончиками, опыляется птицами. Семена лашеналий шаровидные, черные, с придатком, аналогичным придатку семян у пролески сибирской.
Эндемичный для Южной Африки монотипный род литантус (Litanthus) с видом литантус крохотный (L. pusillus) часто встречается в тенистых местах в лесах, в трещинах скал (рис. 45, 13, 14). Это мелкое, субтильное растение, у которого 1-3 узколинейных листочка появляются обычно после цветения. Во время цветения на тоненьком цветоносе распускается один колокольчатый белый или розоватый цветочек длиной 2-5 мм, который открывается после полудня и, по мнению С. Фогеля (1954), соответствует цветкам, опыляемым пчелиными.
Два монотипных рода андросифон (Androsiphon, рис. 45,15) и амфисифон (Amphisiphon) имеют чрезвычайно ограниченное распространение в Капской области - район Кальвинии, где, кроме них, встречаются еще такие же карликовые, почти бесстебельные розеточные лилейные, как поликсена (Polyxena) и массония (Massonia). Род поликсена (2 вида) широко распространен в кустарниковых, травянистых и полупустынных типах растительности Капской области. Этот род Дж. Джессоп (1976) присоединил к капским видам гиацинта, уменьшив общее число видов до двух. Поликсена отличается от азиатских видов гиацинта луковицей, которая ежегодно возобновляется и, кроме листовых и низовых чешуй, имеет еще влагалищную чешую, сегментами околоцветника - белыми или розовыми, тычинками, двурядными, прикрепленными ниже или в зеве трубки. Цветки поликсены ароматны и опыляются пчелиными.
В прибрежной зоне Капской области встречаются оба вида рода велътгеймия (Veltheimia). Велътгеймия капская (V. capensis) с сизыми листьями и луковицей, имеющей продолжительный период покоя, приурочена к западной части области, а велътгеймия прицветниковая (V. bracteata) с блестящими зелеными листьями и луковицей без периода покоя - к восточной части. Высота растений достигает 50-80 см. Их луковицы составлены незамкнутыми листовыми и низовыми чешуями, имеют многолетние ветвящиеся корни и в пазухах чешуй луковички-детки. Цветки длиной до 2,5 см на коротких изогнутых цветоножках в пазухе крупного прицветника с маленьким боковым прицветничком (рис. 45, 17). Околоцветник розоватый, образует слегка изогнутую цилиндрическую трубку пергаментной консистенции, с короткими отклоненными зубчиками (рис. 45, 17). Нити тычинок прикреплены примерно к середине трубки, низбегающие; столбик длинный и тонкий. Вельтгеймия, подобно орнитофильным лашеналиям, имеет признаки цветка, опыляемого птицами. Коробочки вздутые, очень легкие, с большими крыловидными выростами. Когда они падают на землю, ветер, ударяя в крылья, как в лопасти турбины, катит плод по земле (анемогеохория), и из него высыпаются семена. В каждом гнезде завязи по 2 семязачатка, но развивается в семя только один.
Ближайшим к вельтгеймии родом является монотипный род псевдогалътония (Pseudogaltonia) с видом псевдогалътония булавовидная (P. clavata), встречающимся от Анголы до Капской области. Она отличается крупной луковицей с волокнистыми остатками влагалищ листьев, сочными листьями, бледно-зелеными цветками длиной до 4 см на длинных повислых цветоножках.
В Драконовых горах в Южной Африке встречаются оба вида галътонии (Galtonia) с пахучими белыми или зеленоватыми трубчато-воронковидными повислыми цветками.
Род дипкади (Dipcadi) насчитывает примерно 30 видов, распространенных в Африке, на Мадагаскаре (3 эндемичных вида), на Сокотре и в Индии, а также по 1 виду в Южной Европе и Средней Азии. Цветки зеленые или коричневые, иногда с желтым, красным или белым на внутренней поверхности отогнутых кончиков сегментов, внизу сросшихся на 2/3 в трубку, а вверху сближенных. Наружные сегменты продолжаются в короткие до длинных хвостовые придатки, иногда красноватые и испускающий ночью запах, привлекающий ночных бабочек. Семена плоские. Некоторые виды дипкади съедобны, другие - ядовиты.
Род родокодон (Rhodocodon), имеющий 8 видов, эндемичен для Мадагаскара; 7 наземных или наскальных видов встречаются в центре или на западе острова, а родокодон ургинеевидный (R. urgineoides) - эпифит, произрастающий на востоке, во влажном тропическом лесу, в горах на высоте до 2000 м, где спорадически встречается на стволах деревьев, покрытых мхом. Это растение высотой 30-40 см с небольшой луковицей, 2-5 узкоэллиптическими листьями и цветоносом с 20-25 цветками. Прицветники внизу со шпорцем. Околоцветник колокольчатый, длиной 7-8 мм, белый, с зеленоватыми зубчиками. Семена, в отличие от остальных видов, плоские, с крыловидной каймой.
Триба массониевых (Massonieae) включает 2 рода - массония (Massonia) и добения (Daubenуа), виды которых распространены в Южной Африке, преимущественно в Капской области, в сухих районах, на открытых местах, на глинистой, каменистой или песчаной почве. Они зацветают в период зимних дождей и, как свойственно эфемероидам, вегетируют не более 2 месяцев. Это мелкие, розеточные, почти бесстебельные растения с 2 супротивными распластанными по земле листьями (рис. 46, 1), обычно широко-обратнояйцевидными, несколько мясистыми, голыми, волосистыми, с пупырышками или мелкоигольчатыми, иногда пятнистыми, охватывающими основание цветоноса. Цветонос короткий, подземный, с головчатым или щитковидным соцветием, расположенным на уровне земли. Цветоножки короткие, часто мясистые. Цветки многочисленные, прямостоячие, находятся в пазухах крупных зеленоватых прицветников (рис. 46, 2), причем наиболее крупные прицветники краевых (нижних) цветков образуют как бы обертку всего соцветия. Сегменты околоцветника сросшиеся в трубку. Тычинки прикрепляются к зеву трубки и обычно превышают сегменты околоцветника. Пыльники качающиеся, различной окраски. Столбик шиловидный с крошечным рыльцем. Семена шаровидные, черные.
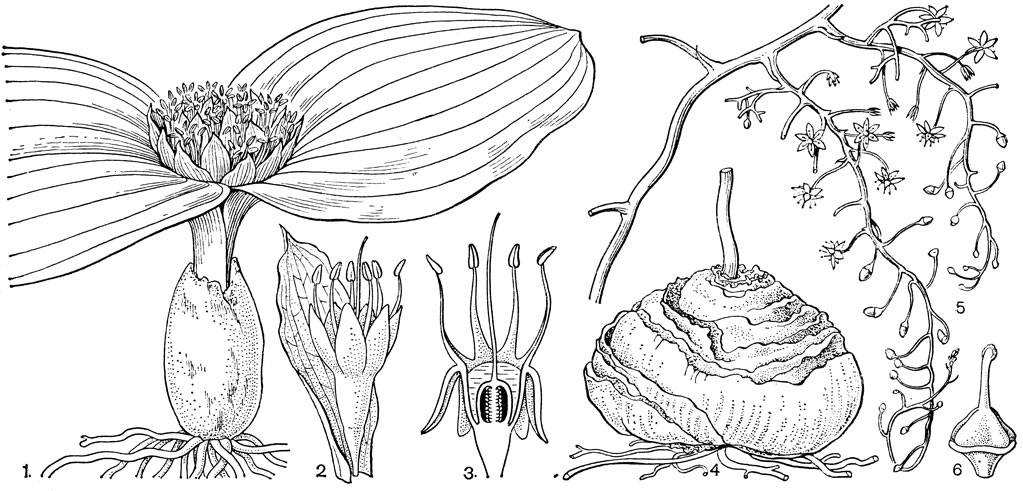
Рис. 46. Лилейные: трибы массониевые и бовиеевые. Массония прижатая (Massonia depressa): 1 - общий вид; 2 - цветок с прицветником; 3 - цветок в разрезе, видна трубка околоцветника, наполненная нектаром. Бовиея вьющаяся (Bowiea volubilis): 4 - луковица; 5 - часть соцветия; 6 - гинецей
Род массония вместе с присоединенным к нему родом необейкерия (Neobakeria) насчитывает 8 видов (Дж. Джессоп, 1976). Луковица массонии находится у поверхности почвы и не превышает в диаметре 2(3) см. Она ежегодно возобновляется и, подобно широколистным лашеналиям, имеет одну тонкую замкнутую влагалищную чешую, которая охватывает сочные запасающие чешуи, представленные основаниями базальных листьев и 1-3 низовыми чешуями. Вегетативное размножение у луковиц слабое (М. В. Баранова, 1976). Околоцветник обычно с длинной (до 20 мм) трубкой и продолговатыми прямостоячими или отклоненно отогнутыми сегментами, обычно розовыми, белыми или зелеными. Тычинки однорядные, иногда нити тычинок в основании спаяны, образуя вместе с трубкой околоцветника бокал, до краев наполненный нектаром, который выделяют септальные нектарники (рис. 46, 3). Цветки обычно имеют приятный запах. Широко распространенную в Капской области массонию колючую (М. echinata), у которой длина трубки околоцветника около 5 мм, опыляют пчелы. Массонию жасминоцветковую (М. jasminiflora), имеющую трубку длиной до 20 мм и диаметром около 1 мм, посещают дневные бабочки с длинным хоботком. У массонии прижатой (М. depressa, рис. 46, 1-3) желто-зеленые мясистые цветки имеют трубку околоцветника длиной около 20 мм и с диаметром зева около 12 мм. В широком зеве цветков блестит скопившийся нектар, привлекая птиц, которые, высасывая его, касаются клювом вверх стоящих тычинок и переносят пыльцу.
Второй род трибы - добения (Danbenya) - с одним видом добения золотистая (D. aurea) имеет ограниченное распространение: маленький район Юго-Западного Kappv в Капской области с годовым количеством осадков 250 мм. Она встречается на высоте 1350-1450 м над уровнем моря, где растет на тяжелых глинистых почвах. Попадаются популяции с красными или желтыми цветками, реже переходные формы и оранжевые, которые ранее принимались за самостоятельные виды. Добения очень похожа по облику на массонии, но отличается сильно зигоморфными краевыми цветками с косой трубкой околоцветника. Наружные 3 сегмента краевых цветков обратнояйцевидные, крупные, длиной до 4 см, лежат на двух супротивных листьях и на земле; 3 других сегмента длиной до 4 мм. Внутренние цветки более или менее актиноморфные, с редуцированными сегментами. Тычинки прикреплены у основания в зеве косой трубки и поэтому имеют разные уровни. Яркая окраска и форма цветков - приспособление к опылению их птицами.
Триба хлорогаловых (Chlorogaleae) включает 2 американских рода с немногочисленными представителями, у которых луковица ежегодно замещается и составлена основаниями базальных листьев и низовыми чешуями. Сегменты околоцветника свободные.
Род хлорогалум (Chlorogalum) насчитывает 5 видов, распространенных преимущественно в Калифорнии на сухих открытых равнинах, холмах или горных склонах, на глинистой или каменистой почве. Луковица хлорогалумов имеет оболочки, тонкие, пленчатые или толстые, с жесткими волокнами (хлорогалум послеполуденный - С. pomeridianum). Базальные листья многочисленные, линейные, распростертые. Стеблевые листья редуцированные, вверху преобразованные в пленчатые прицветники. Соцветие ветвистое, метельчатое. Цветки белые, розовые, голубые. У одних видов цветки открываются днем, у других - к вечеру. Так, у хлорогалума послеполуденного каждый цветок остается открытым только несколько часов. Цветок раскрывается внезапно, часам к 4 дня; до наступления ночи его интенсивно посещают пчелы, после чего он закрывается. Сегменты околоцветника скручиваются после цветения и остаются при плодах. Тычинки с качающимися пыльниками. Столбик тонкий, с 3-лопастным рыльцем. Семена почти шаровидные, обычно по 2 в каждом гнезде. Лишь хлорогалум послеполуденный размножается вегетативно путем деления луковицы. Некоторые виды используют для бордюрных посадок. Луковицы хлорогалума послеполуденного используют как заменитель мыла.
Род камассия (Camassia) объединяет 7 видов, из которых 6 распространены в Северной Америке и 1 вид - камассия двуцветковая (С. biflora) - встречается на западе Южной Америки в Андийской области. Камассии, в отличие от хлорогалумов, встречаются во влажных местах, на горных лугах, в прериях, на травянистых склонах, которые, если не постоянно, то, во всяком случае, весной, во время вегетации растений, бывают увлажнены. Базальные листья многочисленные, линейные. Цветонос высотой до 1 м. Кистевидное соцветие обычно многоцветковое, с довольно крупными прицветниками. Цветки крупные, актиноморфные или слегка зигоморфные, преимущественно голубые, синие, фиолетовые. Сегменты околоцветника остающиеся при плодах, чаще скрученные над завязью. Тычинки с качающимися пыльниками. Столбик нитевидный, с 3-лопастным рыльцем. Семена черные, обратнояйцевидные, по 2-5 и более в каждом гнезде. На западе Северной Америки, где камассии растут большими колониями, луковицы их служат пищей индейцам местных племен.
Триба бовиеевых (Bowieae), относимая ранее к подсемейству асфоделовых (Asphodeloideae), близка к пролесковым, о чем свидетельствуют биохимические данные (Р. Хегнауэр, 1963), строение семян (Г. Хубер, 1969) , жизненная форма (А. П. Хохряков, 1975) и строение луковицы (М. В. Баранова, 1976). Представители трибы, куда относятся 2 африканских рода - бовиея (Bowiea) и схизобазис (Schizobasis) - имеют базальные листья, которые рано (до начала цветения) засыхают и опадают; цветонос прямостоячий или лазающий, сильно ветвистый, безлистный, зеленый, функционирует как ассимилирующий орган и потому сохраняется после плодоношения до конца сезона. Прицветники со шпорцем, рано опадающие. Цветки представителей трибы бовиеевых многочисленные, мелкие, распускаются летом, запаха не имеют. Сегменты околоцветника распростертые, свободные. Семена немногочисленные, черные.
В роде бовиея 2 вида. Бовиея вьющаяся (В. volubilis, рис. 46, 4-6), цветонос которой достигает в высоту нескольких метров, широко распространена в Южной Африке, захватывает юг Юго-Западной Африки и доходит до Кении. Она встречается чаще среди кустарников и деревьев, особенно вдоль берегов рек, но также и в сухих районах, обычна на галечнике. В тропической Африке, в Кении встречается близкий ей вид бовиея килиманджарская (В. kilimandscliarica), обильно покрывающая соседние растения и скалы в горах до уровня 2300 м. Ее оттопыренные суккулентные конечные веточки и цветоножки опираются на вечнозеленые кустарники или камни, поддерживая свой основной стебель. Луковица у бовиеи часто находится на поверхности почвы и зеленеет на свету. Она крупная, достигает в диаметре 15 см и состоит из 7-10 полузамкнутых, очень сочных чешуй, принадлежащих 2-3 вегетационным циклам. В одном цикле формируются 2 листа и 1-2 низовые чешуи. Корни многолетние, ветвящиеся. В пазухах чешуй закладываются луковички-детки (М. В. Баранова, 1976). Луковицы ядовиты. Из луковиц бовиеи вьющейся получены гликозиды, подобные гликозидам дримии морской, обладающие высокой кардиотонической активностью. Цветки бовией зеленые. Сегменты околоцветника остаются при плодах.
Род схизобазис имеет 2 вида, распространенных в Южной и Юго-Западной Африке. У схизобазиса луковица составлена черепитчатыми чешуями, съедобная. Цветонос прямостоячий или лазающий. Цветки белые или бледно- розовые, ночные. Сегменты околоцветника опадают при плодах.
Семейство альстрёмериевые (Alstroemeriaceae) (М. В. Баранова)
Семейство альстрёмериевых объединяет 4 рода и свыше 200 видов, распространенных главным образом в Южной Америке. Лишь несколько видов рода бомарея (Bomarea) встречаются в центральных районах Мексики. Большинство альстрёмериевых - обитатели влажных низинных и горных тропических и субтропических лесов. Однако некоторые представители встречаются в высокогорьях, поднимаясь до высоты 4000 м над уровнем моря, и в пустынных районах. Наибольшее число видов принадлежит - родам бомарея (150 видов) и алъстрёмерия (Alstroemeria - 50 видов).
Семейство представляют корневищные травы с сочными веретеновидными и волокнистыми корнями с прямостоячими или лазающими стеблями или лианы. Листья на стеблях очередные, цельные, линейные, ланцетные или яйцевидные, голые или опушенные, сидячие или короткочерешковые. Особенностью большинства представителей семейства является перекручивание черешка листа на 180°, так, что морфологически нижняя его сторона располагается сверху, а верхняя - снизу. Интересно отметить, что черешок первого листа сеянца у альстрёмерий также уже перевернут. Эпидерма листа с аномоцитными устьицами. Сосуды обнаружены только в корнях, членики сосудов с простой перфорацией.
Цветки собраны в верхоцветные простые или сложные зонтиковидные или головчатые (леонтохир - Leontocfrir) соцветия, окруженные при основании листьями, или реже одиночные (шикенданция - Schickenclantzia). Цветки довольно крупные, диаметром до 5 см, актиноморфные или зигоморфные (алъстрёмерия, рис. 47, табл. 11, 12), желтые, красные, оранжевые, фиолетовые с темными штрихами или пятнами, обычно без запаха. Околоцветник из 6 свободных сегментов, расположенных в 2 круга. Сегменты околоцветника ланцетные или лопатчатые, с оттянутыми, узкими, в виде длинных ноготков основаниями, иногда сегменты наружного круга по длине и окраске отличаются от сегментов внутреннего круга (бомарея). В основании последних в виде узкой борозды находятся нектарники. Тычинок 6, расположены они в 2 круга. Их нити длинные, свободные, голые или в основании коротко опушены (альстрёмерия, бомарея), прикреплены к подпестичному диску. Пыльники удлиненные, они как бы насажены основанием сверху на ось тычиночной нити, их гнезда сросшиеся и вскрываются боковой продольной щелью, связник несколько возвышается над гнездами пыльников. Пыльцевые зерна крупные (длина большей оси зерна до 100 мк у альстрёмерии), с одной узкой и длинной бороздой, нередко переходящей на проксимальную сторону зерна (бомарея); экзина сетчатая или ямчатая. Гинецей синкарпный с центрально-угловой плацентацией (альстрёмерия, бомарея) или паракарпный с париетальной плацентацией (леонтохир, шикенданция), с многочисленными семязачатками. Завязь нижняя или полунижняя (бомарея); столбик нитевидный, трехгранный, долго не опадающий; рыльце цельное или трехлопастное.
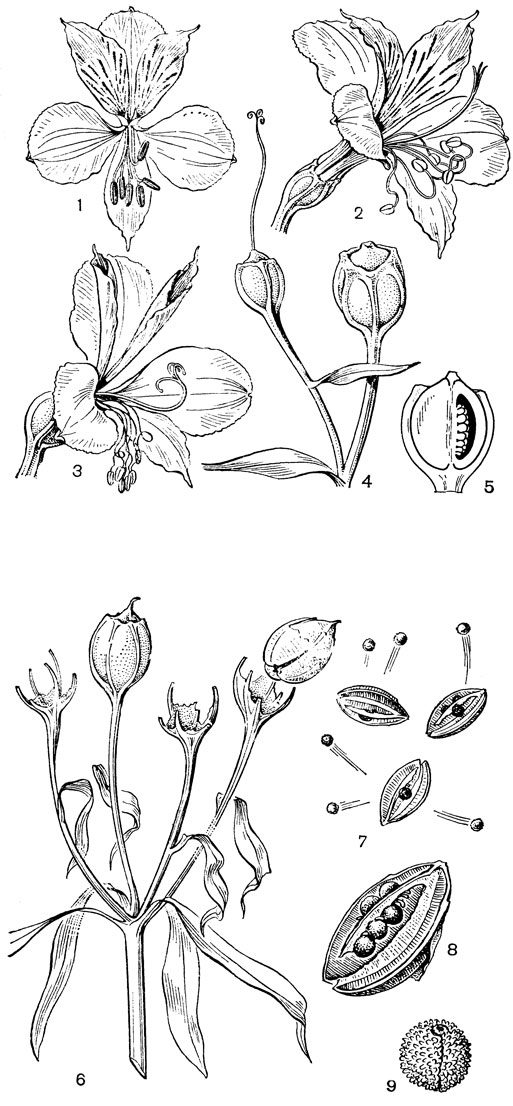
Рис. 47. Альстрёмерия золотистая (Alstroemeria aurantiaca): 1 - только что раскрывшийся цветок; 2 - он же в стадии вскрывания пыльников; 3 - он же после опыления; 4 - молодой плод; 5 - продольный разрез плода; 6 - созревшие плоды; 7 - распавшиеся гнезда плода с семенами; 8 - общий вид гнезда плода с семенами; 9 - семя

Таблица 11. Альстрёмериевые: 1 - альстрёмерия хорошенькая (Alstroemeria pulchella); 2 - бомарея Кальдаса (Bomarea caldasii)
Плод - локулицидная коробочка, более или менее усеченная, вскрывающаяся полностью от верхушки до основания (альстрёмерия) или только на верхушке (бомарея, леонтохир),) иногда с разбрасывающим семена механизмом. Семена многочисленные, шаровидные с мелкобугорчатой семенной кожурой и длинным узким семенным швом, с обильным эндоспермом и маленьким прямым зародышем.
Род альстрёмерия был описан К. Линнеем и назван по имени шведского ботаника К. Альстрёмера. Он впервые привез К. Линнею образцы семян алъстрёмерии лигту (A. ligtu) и алъстрёмерии пелегрина (A. pelegrina) из Испании, где выращивали эти растения. Альстрёмерии широко распространены по всей Южной Америке. Это корневищные травы с прямыми стеблями. Условия произрастания альстрёмерий очень разнообразны. Они растут в тропических лесах (альстрёмерия колокольчатоцветковая - A. campaniflora, альстрёмерия пиауисская - A. piauhyensis), в пустыне Атакама в Чили (альстрёмерия травянистая - A. graminea, альстрёмерия многолистная - A. polyphylla), в Аргентине, в Патагонии на гальке по берегам озер (альстрёмерия карликовая - А. папа), в высокогорьях Анд Боливии и Перу на высотах до 4000 м над уровнем моря (альстрёмерия снежная - A. nivalis). В нотофагусовых лесах и на полузаболоченных лугах растут виды альстрёмерии с длинными, сильно ветвящимися корневищами, залегающими у поверхности почвы на глубине 5-7 см (альстрёмерия золотистая - A. aurantiaca, табл. 12, 3). Виды открытых сухих районов отличаются плотными короткими корневищами, которые расположены в почве на значительной глубине - до 60 см. По внешнему виду альстрёмерии очень различны. Наряду с видами,; имеющими стебли высотой до 180 см и соцветия с 30 и более цветками (альстрёмерия бразильская - A. brasiliensis), встречаются виды карликовых размеров, едва заметные над землей, высотой 3-8 см, с одним цветком (альстрёмерия карликовая). Нельзя не отметить, что немало видов альстрёмерий (например, альстрёмерия хорошенькая - A. pulchella, табл. 11, 1 или альстрёмерия амазонская - A. amazoniса) формируют два типа побегов - репродуктивные и вегетативные. На вегетативных побегах листья крупные, многочисленные, обычно сближены у верхушки, образуют подобие мутовки, а на репродуктивных побегах листья мелкие, равномерно и редко расположены.
Таблица 12. Альстрёмериевые и филезиевые. Лапажерия розовая (Lapageria rosea): 1 - часть растения с цветком; 2 - листочки околоцветника наружного и внутреннего круга; 3 - альстрёмерия золотистая (Alstroemeria aurantiaca)
Оранжевые, красные, сиреневые цветки альстрёмерий очень декоративны и издавна выращивались на родине в садах под названием "перуанская лилия" или "лилия инков" (табл. 11 и 12).
Интересны приспособления к перекрестному опылению у зигоморфных альстрёмерий. Яркая окраска цветков, пыльца и нектар привлекают опылителей - пчел и колибри (X. Стинсон, 1942; В. Грант, 1966). Указателем места нектара, который скапливается в основаниях двух верхних сегментов околоцветника внутреннего круга, служат темные штрихи на них. Цветки альстрёмерий протандричны. При раскрывании цветков тычиночные нити с пыльниками располагаются вначале горизонтально (рис. 47, 1). При созревании пыльников (первыми созревают пыльники тычинок наружного круга) тычиночные нити загибаются вверх, а затем скручиваются спирально в верхней части, располагая пыльник так, что он оказывается на пути к двум верхним нектароносным сегментам околоцветника. Несколько позже то же происходит и с тычинками внутреннего круга. При сборе нектара на опылителя неизбежно попадает пыльца и переносится им на другой цветок. Любопытно, что после опыления нектароносные сегменты околоцветника блекнут, плотно продольно скручиваются и закрывают доступ к нектарникам. Во время созревания пыльцы столбик с плотно сомкнутыми лопастями рыльца отогнут вниз, в противоположную от тычинок сторону. Лишь после вскрытия и опустошения пыльников столбик отгибается вверх, длинные лопасти его рыльца расходятся в стороны и оно готово принять чужую пыльцу. При отсутствии опылителей виды альстрёмерий способны к ветроопылению. Подрбное явление наблюдал X. Стинсон (1942) у альстрёмерии золотистой при выращивании ее на Тихоокеанском побережье США. В том случае, когда по каким-либо причинам опыление не произошло, растения самоопыляются.
Не менее интересен механизм раскрывания плода и разбрасывания семян у альстрёмерий. Кожистые трехгнездные коробочки при созревании разрывают охватывающее их сросшееся пленчатое основание сегментов околоцветника с 6 проводящими пучками, лопаются на верхушке и распадаются на три створки (рис. 47, 6-8). При этом плаценты, расположенные в центре коробочки, отрываясь от ее основания, с силой разбрасывают в стороны шаровидные семена.
Практическое применение издавна находят сочные веретеновидные корни альстрёмерий. Они богаты крахмалом, состоящим из амилозы (30%) и амилопектина (70%). Зерна крахмала альстрёмерии лигту и альстрёмерии чилийской (A. chilensis) имеют очень сложную своеобразную форму. Крахмал корней используют в народной медицине и для изготовления высококачественного клея.
Особо следует отметить красоту цветков альстрёмерий, благодаря чему эти растения вызывают большой интерес у садоводов, которые в последние десятилетия проводят с ними активную селекционную работу. Получение полиплоидных крупноцветковых форм с различной окраской цветков и, что особенно ценно, со способностью сохранять декоративность в срезке до 30 и более дней (триплоидные сорта) явилось новым стимулом к дальнейшей работе с альстрёмериями. В настоящее время получено около 50 сортов.
Род бомарея (Bomarea) объединяет лазающие растения и лианы, которые произрастают на влажных лесных склонах, в кустарниковых зарослях, встречаются на сфагновых болотах и в горах, поднимаясь на высоту до 4200 м над уровнем моря. Это единственный род семейства, виды которого, кроме Южной Америки, встречаются в Центральной Америке, заходя на север до 20° с. ш. (бомарея коротковолосистая - В. hirtellus). Стебли бомарей длиной до 3-5 м оплетают кустарники и деревья (фейхоа, подокарпусы, дубы), или, не встречая опоры, их спутанные стебли лежат на земле. Некоторые виды засоряют посевы. Так, в Эквадоре и Колумбии злостным сорняком картофельных полей является бомарея Калъдаса (В. caldasii, табл. 11, 2). Листья на стеблях очередные, достигают в длину 20 см и нередко густо опушены иногда длинными вьющимися волосками, что придает растениям своеобразный вид (бомарея Калъбрейера - В. kalbreyeri). Яркие красные, 1 желтые, розовые цветки на цветоножках длиной до 25 см в свисающих зонтиковидных соцветиях привлекают колибри, которые, порхая в воздухе, собирают нектар из оснований внутренних пятнистых сегментов околоцветника и опыляют цветки. Обычно колибри привлекают красные цветки. Однако американскому исследователю В. Гранту (1966) в тропической Мексике не раз приходилось наблюдать, как колибри опыляют светло-розовые цветки бомарей остролистной (В. acutifolia).
Плоды бомарей - коробочки с толстым околоплодником - растрескиваются от верхушки почти до основания и висят, обнажая внутри яркие янтарно-оранжевые шаровидные семена, прикрепленные к плаценте. При ее ссыхании семена опадают. Есть также основания полагать, что яркая окраска семян бомарей привлекает птиц, которые, склевывая их, разносят семена на значительное расстояние. Своеобразную группу составляют карликовые одноцветковые виды бомарей, высота которых не превышает 5-8 см. Их плоды растрескиваются только на верхушке.
Бомарей известны главным образом как декоративные растения. Их крупные пониклые соцветия, насчитывающие нередко до 100 и более, трубчатых, длиной до 5 см ярких цветков, очень красивы. Среди них особенно декоративны бомарея патакохская (В. patacocensis) с ярко-малиновыми цветками, произрастающая в Эквадоре и Колумбии, бомарея Кальдаса с оранжевыми цветками из Эквадора (табл. 11, 2), бомарея сильнейшая (В. fortissima) с красными цветками из Перу и многие другие. Впервые бомареи в Европе цвели в 1846 г. в Англии в садоводстве Вича. В 1926 г. в Англии Г. Банкс получил первые межвидовые гибриды бомарей.
Сладкие корневые клубни бомареи яйцевидной (В. ovata) под местным названием "улубайя" используют в пищу в Перу и Боливии, а настойку из клубней бомареи салсиллы (В. salsilla) применяют в народной медицине как обезболивающее средство.
Единственный вид рода леонтохир - леонтохир Оваллъе (Leontochir ovallei) произрастает в горах Чили. Многочисленные прямые стебли растения высотой до 100 см густо покрыты широколанцетными с перевернутыми черешками листьями, которые заметно уменьшаются в размере вверх по стеблю. Красновато-оранжевые на коротких, длиной до 1 см, цветоножках, цветки собраны в плотные головчатые, почти шаровидные соцветия и расположены на верхушках стеблей. За некоторое сходство соцветия леонтохир называют на родине "лапой льва" или "когтями льва". Все сегменты околоцветника в цветке лопатчатые, с оттянутым узким основанием. Тычиночные нити длинные, пыльники прикреплены к ним основанием. Кеглевидная завязь одногнездная, с париетальной плацентацией. Столбик с трехлопастным рыльцем короче тычинок. Плод - ребристая коробочка, вскрывающаяся на верхушке. Мелкие шаровидные семена легко высыпаются из плода при ссыхании плаценты.
Род шикенданция (Schickendantzia) включает 2 вида, которые произрастают в горах Аргентины. Это прямостоячие травянистые одноцветковые растения, близкие роду альстрёмерия. Особенности опыления неизвестны.
Семейство луковые (Alliaceae) (Г. Л. Кудряшова)
Семейство луковых объединяет около 30 родов и 650 видов, распространенных на всех континентах, кроме Австралии. Большинство видов сосредоточено в умеренных областях северного полушария (Ирано-Туранская, Средиземноморская, Атлантическо-Северо-Американская, Мадреанская флористические области). В Циркумбореальной области число видов невелико, но они играют значительную роль в образовании растительного покрова. Один вид - лук скорода (Allium schoenoprasum) - заходит в Арктику до 75° с. ш. (Новая Земля). Это северная граница ареала семейства. В южном полушарии (тропические и умеренные области Южной Америки и Африки) представлены главным образом монотипные и олиготипные роды (около 14), объединяющие приблизительно 70 видов, и в меньшей степени виды родов, основной ареал которых находится в северном полушарии. Ареал семейства в южном полушарии доходит почти до 50° ю. ш. в Патагонии (тристагма патагонская - Tristagma patagonicum).
Виды луковых очень разнообразны по своей экологической приуроченности и встречаются почти повсюду от высокогорий до побережья морей. Более того, иногда два близких вида имеют разную экологию: один сухолюбивый (лук горный - Allium montanum), другой,, очень близкий к нему, влаголюбивый, растущий на влажных, богатых аллювиальной почвой лугах (лук угловатый - A. angulosum). Многие виды луков в Евразии - типичные луговые растения (лук сибирский - A. sibiricum, лук скорода, лук монгольский - A. mongolicum, лук двузубчатый - A. bidentatum, лук душистый - A. odorum и др.). Среди луков есть и сорняки сенокосных лугов, например черемша, или лук победный (A. victorialis). Многие виды луковых - обитатели лесов. Во влажных тенистых лесах на богатых гумусом почвах в громадных количествах встречается лук медвежий (A. ursinum). Характерный его запах распространяется далеко вокруг. Особенно часто растет он по оврагам в сообществе папоротников и осоки пальчатой (Carex digitata). В гирканских лесах Азербайджана и Ирана произрастает лук странный (A. paradoxum). Он имеет все характерные признаки рода лук, но крупные белые цветки делают его больше похожим на белоцветник летний (Leucojum aestivum) из семейства амариллисовых. В лесах средиземноморской области Европы (включая Крым) и Малой Азии растет нектароскордум Диоскорида (Nectaroscordum dioscoridis). На болотах в Америке растут некоторые виды рода бродиея (Brodiaea), в субтропическом поясе на сырых местах виды рода трителея (Triteleia), нотоскордума (Nothoscordum), милла (Milla). Среди луковых есть и обитатели засоленных почв (муилла приморская - Muilla maritima). Большинство же видов луковых, и притом наиболее своеобразные представители семейства, растут в степных и полупустынных областях низменностей и гор. Здесь наибольшее разнообразие форм луков - самого большого рода семейства. В высокогорьях Гималаев один из морфологически наиболее интересных представителей семейства - монотипный род милула (Milula), в полупустынях Калифорнии и Аризоны (США) - другой замечательный монотипный род семейства - гесперокаллис (Hesperocallis), в Андах - все роды своеобразной трибы гилисиевых (Gilliesiae).
Луковые - многолетние травы с луковицами, клубнелуковицами или иногда корневищами (триба агапантовых - Agapantheae). Корни обычно тонкие, нитевидные, но бывают и утолщенными. Толстые контрактильные корни многих луковых служат для втягивания луковицы на нужную глубину. Соцветие выносится на поверхность земли цветочной стрелкой, которая иногда выглядит как облиственный стебель, благодаря тому что листья снабжены влагалищами, охватывающими стрелку почти доверху. Луковицы морфологически очень разнообразны. Они бывают как одиночные, так и растущие на общем корневище. Первые, как правило, округлые и более мясистые, в то время как вторые узкие, удлиненные. Между этими крайними типами есть переходные формы, например у видов секции цепа (Сера), к которой относится культурный лук репчатый (А. сера). Луковицы у него крупные, округлые, но прикреплены по нескольку к общему корневищу, что можно наблюдать у дикорастущих видов, близких к луку репчатому. В культуре же у лука репчатого этот признак не выражен, так как он выращивается только в течение двух лет. Среди одиночных луковиц наиболее интересна сборная луковица, состоящая из маленьких узких луковичек, "зубков", например у хорошо всем знакомого чеснока (A. sativum). Луковицы характеризуются большим многообразием и по другим признакам. Наружные оболочки, которые защищают луковицы от неблагоприятных внешних воздействий, имеют различную консистенцию - толстые, кожистые или пленчатые, бумажистые, волокнистые, сетчатые. Последние образованы склеренхимными клетками с сильно утолщенными стенками. Листья луковых приземные (базальные), очередные, простые, трубчатые или чаще плоские, линейные, продолговатые, эллиптические, за редким исключением (лук победный, лук функиелистный - A. funkiifolium, рис. 48, 7-8 и др.), без черешков, с параллельным или дуговидным жилкованием, большей частью голые, но бывают и опушенные или по краю грубо-реснитчатые. Трубчатые листья имеют внутри полость, возникшую в результате разрушения бесхлорофилльной паренхимы. Цветки собраны в верхушечные зонтики, за исключением рода милула, у которого соцветие - колос, и гесперокаллиса, имеющего кистевидное соцветие. Зонтики луковых разнообразны как по количеству цветков в соцветии (от 1 до 500 и более), так и по длине цветоножек; у основания цветоножек часто имеются прицветнички. Зонтик до цветения окутан покрывалом из 1-2-5 сросшихся листочков, различающихся по форме и размерам. Цветки обычно маленькие, 3-10 мм, но бывают до 2,5 см (например, у видов рода бревуртия - Brevoortia), даже до 4 см (у гесперокаллиса, агапантусов - Agapanthus, тулъбагии - Tulbaghia). Они обоеполые, актиноморфные, редко зигоморфные. Околоцветник состоит из 6 свободных или при основании сросшихся лепестковидных сегментов, расположенных в двух кругах. Сегменты внутреннего круга иногда меньше наружных почти вдвое или отсутствуют, а при основании сегментов развит привенчик из 3-12 листовидных чешуек (триба гилизиевых). Сегменты околоцветника продолговатые, иногда с ноготком, с хорошо выраженными жилками в числе 1-7, в которых присутствует хлорофилл. Тычинок обычно 6, в двух кругах. Тычинки внутреннего круга часто отличаются от тычинок наружного круга расширенной уплощенной нитью и наличием зубцов; иногда фертильных тычинок 3, так как тычинки одного из кругов превращены в стаминодии (бродиея, трихлора - Trichlora, эринна - Erinna, солярия - Solaria и др.), иногда сильно видоизмененные (левкокорина - Leucocoryne, бревуртия) или редуцированы (анкрумия - Ancrumia). Пыльцевые зерна однобороздные, со слабо зернистой, морщинистой или сетчатой скульптурой экзины. Гинецей синкарпный, из 3 плодолистиков, с простым нитевидным столбиком, остающимся после цветения и созревания благодаря тому, что канал столбика доходит до дна завязи и там только открывается; завязь верхняя, 3-гнездная, с 1-2 или многими семязачатками в каждом гнезде. Плод - локулицидная коробочка, треугольная, наверху прижатая. Семена шаровидные, угловатые или сжатые, с толстой кожурой, черные, гладкие, диаметром около 3 мм. У некоторых видов (лук странный) семена с придатками (элайосомами) из паренхимных клеток, наполненных маслом. Зародыш маленький, с обильным эндоспермом. Характерный признак семейства луковых - присутствие в чешуях луковиц и зеленых листьях млечников, обычно членистых, заполненных млечным соком (латексом). Для представителей семейства луковых, за исключением рода нотоскордум и нескольких видов из родов лук, тульбагия, левкорина, тристагма, характерно также присутствие во всех тканях растения чесночного и близких к нему летучих масел. Эта масла содержат диалил дисульфид C6H10S2 или диалил трисульфид C6H10S3, определяющие специфический луковый или чесночный вкус и запах. Луковые - перекрестноопыляемые растения. Опылители цветков обычно насекомые (пчелы, мухи, жуки, бабочки), но есть виды с крупными яркими цветками (дихелостемма - Dichelostemma), опыляемые колибри. Насекомых привлекают окраска пветков, запах. В цветках луковых можно найти всю гамму цветов и их тончайших оттенков от чисто-белого до темно-бордового, почти черного. Часто цветки хорошо заметны для насекомых благодаря контрастной окраске сегментов околоцветника и пыльцы, окраске цветоножек, (лук Пачоского - Allium paczoskianum), листочков покрывала (например, виды бревуртии). Большинство цветков луковых обладает приятным для человека запахом. На протяжении всего времени цветения выделяется большое количество нектара. Нектарники у большинства луковых септальные, представляющие собой нектарные щели, которые начинаются при основании плодолистиков и поднимаются до середины их высоты или до верхней трети; нектарные щели выстланы клетками в виде сосочков, не имеющих кутикулы и выделяющих нектар путем диффузии. Нектар вытекает через отверстия, расположенные внизу, в середине или на верхушке завязи, на дно цветка и собирается там, как в блюдце, между завязью и основанием 3 внутренних тычинок. У видов с открытыми звездчатыми цветками нектар доступен многим насекомым, а у видов с цветками, трубчатыми при основании, только длиннохоботковым насекомым. У видов некоторых родов наряду с септальными развиты стаминальные нектарники, например у видов нектароскордума, у которого при основании тычинок развиты нектароносные бугорки. А у видов гилисии как нектарники функционируют язычковидные выросты при основании сегментов околоцветника. Перекрестному опылению способствует протандрия или, реже, протогиния (лук прямой - A. strictum, лук приземный - A. chamaemoly, рис. 49, 8-9). Раскрывание пыльников у протандричных видов (например, луков) начинается с тычинок внутреннего круга, нити которых быстро вытягиваются, выставляя пыльники наружу. Как только пыльники раскрылись двумя продольными щелями от верхушки вниз, тычинки склоняются наружу и интрорзные пыльники полностью сохраняют пыльцу для насекомого. После освобождения от пыльцы тычинки увядают и свешиваются наружу. С раскрыванием пыльников внутреннего круга начинается рост тычинок наружного круга. А после того как раскрылись последние, начинается быстрый рост столбика. И лишь после увядания всех тычинок рыльце бывает готово к восприятию пыльцы (на что указывает появление сосочков на его поверхности) и функционирует в течение нескольких дней. Интересно, что у самых нижних цветков соцветия развитие сильно задерживается; нити тычинок остаются прямыми, а пыльца остается в пыльниках, пока рыльце не достигнет зрелости. Это можно рассматривать как механизм, обеспечивающий самоопыление у какого-то количества цветков. Самоопыление у луковых происходит наряду с перекрестным довольно часто. Этому благоприятствует отсутствие самонесовместимости в цветках. Кроме того, механизмы предохраняющие от самоопыления, оказываются не очень действенными, так как у луков в соцветии обычно много цветков, находящихся к тому же в разных стадиях цветения, и самоопыление производят насекомые, прилетающие за нектаром и переползающие в соцветии с одного цветка на другой.

Рис. 48. Луковые. Лук алтайский (Allium altaicum): 1 - общий вид; 2 - сегменты околоцветника с тычинками. Лук многокорневой (A. polyrhizum): 3 - общий вид; 4 - сегменты околоцветника с тычинками. Лук щетинолистный (А. setifolium): 5 - общий вид; 6 - сегменты околоцветника с тычинками. Лук функиелистный (A. funkiifolium): 7 - общий вид; 8 - сегменты околоцветника с тычинками. Лук огородный (A. oleraceum): 9 - соцветие с луковичками при основании цветоножек и длинным листочком покрывала
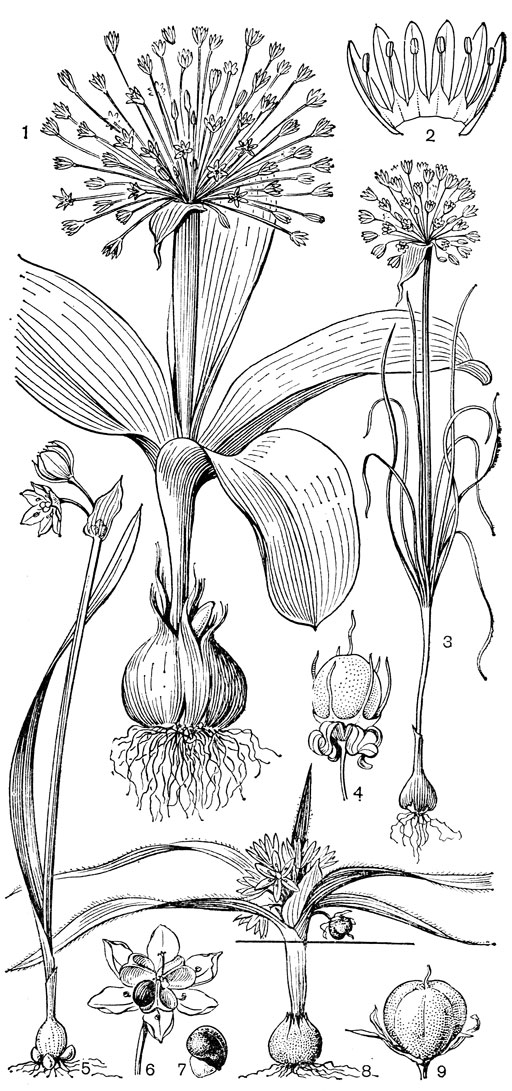
Рис. 49. Луковые. Лук Шуберта (Allium schubertii): 1 - общий вид; 2 - сегменты околоцветника с тычинками. Лук мутовчатый (A. verticil latum): 3 - общий, вид; 4 - коробочка с оставшимися столбиком, тычинками, сегментами околоцветника. Лук странный (A. paradoxum): 5 - общий вид; 6 - раскрывшаяся коробочка с оставшимися сегментами околоцветника; 7 - семя с элайосомой. Лук приземный (A. chamaemoly): 8 - общий вид; 9 - коробочка
В соцветии луков нередко образуются луковички при основании цветоножек. Луковички эти округлые, зеленые или коричневые, лишенные кожистых оболочек и тем значительно отличающиеся от подземных, которые также нормально образуются у таких растений. У видов, образующих луковички в соцветии, наблюдаются разные степени подавления полового размножения. Эмбриологическими и цитологическими исследованиями этих видов установлено, что даже в тех случаях, когда семенное размножение по внешним признакам протекает нормально, наряду с обычным ходом процессов семенообразования отмечается целый ряд аномалий (ранняя дегенерация материнских клеток пыльцы и др.). У нескольких видов, образующих большое количество луковичек в соцветии, в первую очередь назовем наиболее широко известный и хорошо изученный чеснок, наблюдаются столь глубокие нарушения процессов полового размножения, что семенообразование почти не имеет места. В целом отмечена корреляция между вегетативной и семенной репродукцией: чем больше фертильной пыльцы, тем меньше продукция луковичек в соцветии. Вегетативное размножение особенно развито у полиплоидов. Большая роль при вегетативном размножении у луковых принадлежит луковицам и корневищам. Независимо от образования семян на луковицах из вегетативных почек, расположенных на донце, развиваются новые дочерние луковицы. Помимо этого, многие виды, например широко распространенный у нас лук круглый (A. rotundum), лук виноградный (A. ampeloprasum), образуют на луковицах очень мелкие, величиной с горошину, так называемые детки-луковички, также служащие для вегетативного размножения. Иногда детки-луковички образуются не на луковицах, а на верхушках листьев (лук волшебный - А. magicum).
Большинство луковых образует большое количество семян, и молодые растения размножаются исключительно семенным способом. Семена в природе распространяются ветром, реже животными, в том числе муравьями. Семян бывает очень много за счет большого количества коробочек в зонтике. В каждой камере коробочки одно, несколько или много семян, которые часто остаются там в течение долгого времени после созревания, хотя коробочка открывается достаточно широко для их освобождения. Иногда коробочки поедаются овцами и другими животными, и семена распространяются с экскрементами, так как, проходя через пищеварительный тракт, они не повреждаются. Сухие растения предыдущих лет с жизнеспособными семенами, все еще заключенными в коробочки, часто можно встретить в природе. Постепенно они разносятся ветром по типу перекати-поле. Некоторые виды агапантусов и виды бродиеи имеют семена с пленчатой крыловидной окраиной, способствующей распространению их ветром. Иногда семена снабжены элайосомами (лук странный, лук медвежий), благодаря которым они растаскиваются муравьями. Луковички также распространяются на значительные расстояния ветром, дождем, при падении на них камней с обрывов, осыпях земли, разрушении скал и так далее. Распространению их ветром способствует наружная сухая очень легкая волокнистая или сетчатая оболочка. Луковички видов, растущих в прилиторальной зоне, например лука изменчивого, лука виноградного, переносятся морской водой. При этом луковичные оболочки обеспечивают плавучесть и защиту от намокания.
Согласно современным взглядам систематиков на семейство луковых, в нем выделяют 6 триб: агапантовые (Agapantheae), луковые (Аllieae), гесперокаллисовые (Hesperocallideae), гилисиевые (Gilliesieae), миллиевые (Milleae) и бродиеевые (Brodiaeae).
Триба агапантовых объединяет 2 африканских рода - агапантус (Agapanthus) и тулъбагию (Tulbaghia). Отличительные морфологические признаки этой трибы - воронковидный или цилиндрический околоцветник из сросшихся при основании сегментов, рыльце цельное, тычинки сросшиеся с трубкой околоцветника, в каждом из 3 гнезд завязи - многочисленные семязачатки, хорошо развитое короткое корневище, покрытое узкими основаниями листьев. Род агапантус (около 8 видов) целиком южно-африканский (Трансвааль, Наталь, Оранжевая провинция, Капская область). Это красивые растения с голубыми или белыми воронковидными, слегка зигоморфными цветками - по 20-30, иногда до 100 у агапантуса восточного (A. orientalis) в зонтичном соцветии с легко опадающим покрывалом из двух узких верховых листьев. В бутоне покрывало с открытым косым носиком, не полностью сросшимся, в отличие от других луковых. Отличительный признак рода - короткие пыльники на длинных нитях. При основании цветоноса длинные ремневидные листья, причем есть виды листопадные с ежегодно отмирающими листьями и вечнозеленые. Род тульбагия (около 30 видов) распространен в Южной и тропической Африке. Как и агапантусы, это крупные красивые растения, но отличаются кувшиновидным или цилиндрическим околоцветником с тремя мясистыми придатками у внутренних сегментов околоцветника, длинными пыльниками.
Триба луковых (Allieae) самая большая и разнообразная в семействе - 8 родов и около 550 видов, имеющих околоцветник свободный или сросшийся, открытый звездчатый или колокольчатый, очень редко с придатками у внутренних сегментов околоцветника, 6 тычинок, свободных или сросшихся между собой и с сегментами околоцветника, с покрывалом из 1-4 сросшихся долей, с хорошо выраженной луковицей. Центральное положение в трибе луковых занимает род лук (Allium), около 500 видов которого богато представлены почти по всему ареалу семейства: в Азии (несколько центров видового разнообразия), Европе (главный центр распространения - Средиземноморье), Северной Америке, Африке (несколько заходящих и близких к евразиатским видов). Род включает 6 подродов, 30 секций, большинство из которых состоит из нескольких групп родства. У луков актиноморфный околоцветник из 6 обычно свободных сегментов с одной жилкой, без придатков, 6 фертильных тычинок, также свободных или сросшихся между собой и с околоцветником; завязь верхняя; соцветие - большей частью шаровидный или полушаровидный зонтик, до цветения окутанный покрывалом, сросшимся из 2-4 долей; луковицы всегда развиты и очень разнообразны по характеру роста (одиночные или на корневищах), способности к делению и образованию луковичек-деток, по величине и форме, цвету, консистенции, жилкованию или сетчатости луковичной оболочки. Листья луков очень разнообразны: с черешками или безчерешковые, дудчатые, нитевидные, плоские (по форме линейные, продолговатые, овальные), одиночные или многочисленные. Общие родовые признаки в различных группах также выражены очень разнообразно, благодаря чему представители этих групп резко отличаются по внешнему виду. Наиболее близок к луку род нектароскордум (Nectaroscordum), насчитывающий 6 видов, похожих на крупные луки, но отличающихся дисковидными цветками с короткой трубкой и сегментами околоцветника с 3-7, а не с одной жилкой, покрывалом из одной доли. Ареал рода - Средиземноморье (включая Крым), Малая Азия, Иран, Закавказье. Другой близкий к лукам род - нотоскордум - распространен в Америке (1 вид в Северной, 15-16 видов в Южной) и Восточной Азии (2 вида). Сегменты околоцветников у нотоскордумов сросшиеся до середины, с одной жилкой; тычинок 6, приросших к основанию сегментов околоцветника; пыльники удлиненные; завязь 3-гнездная, с 6-12 семязачатками в каждом гнезде; коробочка кожистая, 3-лопастная. Американские и восточноазиатские виды нотоскордумов представляют собой разные группы родства: у первых покрывало из двух листочков, у вторых - из одного. Род нотоскордум в семействе луковых - один из немногих исключений, не имеющий лукового запаха.
В трибу луковых входит наиболее своеобразный род семейства - милула с одним видом - милула колосистая (Milula spicata, рис. 50, 5, б), обитающим в Восточных Гималаях. В отличие от остальных луковых у этого растения цилиндрическое колосовидное соцветие с покрывалом из одного острого листочка. Однако другие признаки милулы вполне соответствуют положению этого рода в трибе луковых: крупная луковица, линейные листья до одной трети цветоноса, до половины сросшийся околоцветник, 6 тычинок, 3 из них цельные, 3 с боковыми зубцами. Все растение имеет характерный луковый запах. Остальные монотипные и олиготипные роды этой трибы встречаются на ограниченных территориях в Северной и Южной Америке. Это калоскордум (Caloscordum), гаравентия (Garaventia), левкокорина, тристагма.
Триба гесперокаллисовых монотипная. Единственный представитель этой трибы - гесперокаллис волнистый (Hesperocallis undulata, рис. 50, 1) - распространен в Калифорнии и Аризоне, где он обитает на сухих песчаных и каменистых местах. Это единственный вид в семействе луковых с длинным (10-30 см) кистевидным соцветием. При основании соцветия - крупные листочки обертки. Цветки воронковидные, длиной до 3-4 см, с 6 тычинками, скрытыми в трубке околоцветника. Луковица яйцевидная, покрытая оболочкой. Растение имеет характерный луковый запах. 1
Триба гилисиевых наиболее изолированная в семействе луковых, иногда выделяемая в самостоятельное семейство гилисиевых (Gilliesiaceae). В нее входят 9 монотипных и олиготипных родов, встречающихся исключительно в Андах. Выделяются среди луковых зигоморфными цветками с 3-6 тычинками, с чешуйками при основании сегментов околоцветника, напоминающих коронку амариллисовых. Цветки бывают зигоморфными благодаря неравным тычинкам, сросшимся в кувшиновидное косо срезанное образование (гилисия, трихлора, солярия и др.) К тому же 3 тычинки с одной стороны (нижние) фертильные, с другой (верхней) стерильные. Наиболее орхидный вид имеют цветки гилисии злаковой (G. graminea, рис. 50, 7, 8). Зигоморфия цветков бывает также за счет неравных сегментов внутреннего круга. Особенно это выражено у солярии, у которой один из сегментов внутреннего круга почти не развит. Зонтиковидное соцветие гилисиевых обычно состоит из нескольких цветков на длинных цветоножках, верхние листочки покрывала очень неравные, в числе 2 или 1.
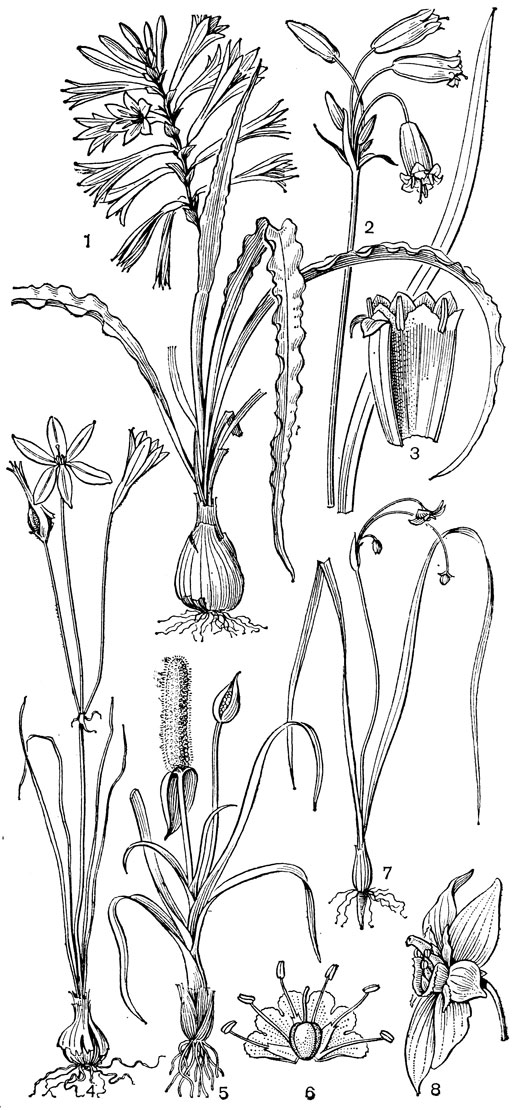
Рис. 50. Луковые. Гесперокаллис волнистый (Hesperocallis undulata): 1 - общий вид. Бродиея ярко-красная (Brodiaea coccinea): 2 - цветонос с соцветием; 3 - сегменты околоцветника с тычинками и чередующимися с ними стаминодиями. Милла двуцветковая (Milla biflora): 4 - общий вид. Милула колосистая (Milula spicata): 5 - общий вид; 6 - цветок в развернутом виде. Гилисия злаковая (Gilliesia graminea): 7 - общий вид; 8 - цветок с привенчиком при основании сегментов околоцветника

Таблица 13. Луковые: 1 - лук победный (Allium victorialis), парк Ботанического института АН СССР в Ленинграде; 2 - лук каратавский (A. karataviense), там же
Триба миллиевых включает 4-6 монотипных и олиготипных родов, сосредоточенных на юге Северной Америки (Калифорния, Мексика, Гватемала). Наиболее крупный род - милла (6 видов). Это небольшие растения с маленькими луковицами, малоцветковым зонтиком и покрывалом из 2-3 узеньких листочков. Цветки у видов этой трибы с актиноморфным цилиндрическим околоцветником, сегменты околоцветника при основании сросшиеся, в результате чего наблюдается тенденция к образованию нижней завязи.
В трибу бродиеевых входят 6-7 родов с актиноморфным околоцветником, сегменты которого до половины срослись в трубку, цветоножками, имеющими, как правило, в верхней части сочленение. В большинстве своем цветки ярко окрашенные, ширококолокольчатые. Ярко окрашенными бывают и листочки покрывала. Все роды этой, трибы распространены в Северной и Южной Америке.
Среди видов семейства луковых много полезных растений - пищевых, лекарственных, декоративных. Это прежде всего виды рода лук, культурные и дикие. Наиболее широко известен и распространен лук репчатый (Allium сера). Предположительная родина его - Средняя Азия, где произрастают ближайшие к нему дикие виды: лук Ошанина (A. oschaninii), лук Вавилова (A. vavilovii), лук смешанный (А. praemixtum), лук пскемский (A. pskemense). Культурной разновидностью лука репчатого считают аскалонский лук, или лук шалот (А. ascalonicum). Вторым по значению после лука репчатого является чеснок (A. sativum), происходящий также из Средней Азии. Довольно широко распространен в культуре лук-порей (A. porrum). Родина его - Средиземноморье, где он больше всего и культивируется. Лук батун, или татарка (A. fistulosum), лук многоярусный (A. fistulosum var. viviparum), лук душистый (A. odorum) наиболее популярны как культурные растения в Восточной Азии, особенно в Китае, где они произрастают в большом количестве и в диком состоянии. Шнитт-лук (A. schoenoprasum) выращивают главным образом в Европе. В диком виде он широко распространен в Евразии и Северной Америке. Наиболее близки к культурным формам шнитт-лука популяции дикого вида, встречающиеся в Альпах. Кроме того, многие дикорастущие виды лука используются населением в пищу: черемша, лук косой (A. obliquum), лук алтайский (A. altaicum, рис. 48,1-2), лук пскемский, лук Вавилова, лук млечноцветный (A. galanthum). Несколько видов лука имеют кормовое значение. Это центральноазиатские виды - лук монгольский, лук многокорневой (A. polyrhizum, рис. 48, 3-4), лук двузубчатый, лук душистый, составляющие основу так называемых луковых степей - прекрасных пастбищ. Из зеленой массы монгольского и многокорневого луков местные скотоводы делают очень питательные подсоленные брикеты для зимней подкормки скота.
Культурные луки не близки систематически и относятся к разным подродам. Лук репчатый, татарка, шнитт-лук, лук душистый относятся к подроду корневищных луков (Rhizirideum); чеснок и лук-порей из подрода настоящих луков (Allium). В культуре же их обычно классифицируют на однолетние (чеснок), двулетние (лук репчатый, порей) и многолетние (татарка, лук многоярусный, шнитт-лук, лук душистый). Первые выращивают 1-2 года главным образом ради луковиц, хотя молодая листва также употребляется в пищу; вторые возделывают преимущественно ради зеленых листьев, которые начинают отрастать очень рано и потом растут в течение всего вегетационного периода.
Луки как культурные растения сопутствуют человеку с глубокой древности. Несомненно, что уже первобытный человек, разыскивая различные растения с острым вкусом, обращал особое внимание на дикие луки, а затем сознательно разводил их вокруг своих стоянок. Первые изображения растений репчатого лука датируются 3200-2700 гг. до н. э. Начало культуры лука репчатого относят к 4000 г. до н. э. Упоминания о луках находят в клинописи древних шумеров, египетских папирусах. Более поздние многочисленные литературные документы и произведения древнего искусства изобилуют сведениями о популярности лука в Древнем Египте, Греции, Римской империи, в странах Ближнего Востока, где он почитался не только как пищевое и лекарственное растение, но и как совершенно необходимое при религиозных ритуалах, мумификации, захоронениях, для магических действий. Среди надписей в храмах, на статуях, на крышках гробов сохранились формулы с упоминанием лука и чеснока для совершения обрядов поклонения, захоронения, произнесения клятв.
Древние египтяне считали лук и чеснок божественными растениями, за что позднее были высмеяны Ювеналом в его "Сатирах". Лук и чеснок приносили па алтари богам вместе с хлебом, мясом, другими овощами и фруктами. На Великой пирамиде в Гизе среди кое-где еще сохранившихся надписей, большей частью религиозного содержания, есть названия лука и чеснока как овощей, необходимых для питания царей в загробном мире. Сохранились изображения лука репчатого в виде крупной очищенной луковицы и нежной зелени на столах пиршеств царей, где он соседствует с мясом, дичью, вином, хлебом, фигами, виноградом. В письменных документах 260-80 гг. до н. э., например в архиве Зенона, управляющего поместьем одного из придворных царя Птолемея II, содержатся уже рекомендации по выращиванию лука, сведения о различных его сортах.
Столь же почитаемы были лук и чеснок у древних греков. Лук, особенно крупные луковицы, приносили в дар пифиям во время пиршеств в честь богов в Дельфах в храме Аполлона. Лук дарили новобрачным (по преданию, у богини Латоны перед рождением близнецов Аполлона и Артемиды улучшилось состояние здоровья благодаря луку). Известно, что афинский полководец Ификрат получил в качестве одного из свадебных подарков бочку лука. Римский полководец Ксенофонт ввел лук в ежедневный рацион своих солдат, так как луку приписывали способность восстанавливать силу и энергию у воинов. Римляне особенно ценили лук как средство, якобы прогоняющее души умерших - демонов и лемуров, которых они особенно боялись. Наряду с этим известны области, например Пелузиум, где к луку было прямо противоположное отношение. По свидетельству Плутарха, там считали лук даже опасным для человека, как растение, чей неприятный запах привлекает злых духов. В Индии, где лук выращивают с глубокой древности и где было хорошо известно его благотворное действие на организм, в пищу его совсем не употребляли из-за плохого запаха, а пользовались только как лекарством.
Популярность лука как ценного пищевого и лекарственного растения постоянно возрастала. В средние века культура лука распространилась в Европе сначала во Франции, Испании, Португалии, позднее в России, Германии, Англии. И очень скоро у народов этих стран лук стал незаменимой приправой к самым различным блюдам, а у бедных слоев населения составлял повседневную пищу.
Наибольшего развития культура лука достигла в X-XII вв. в Испании. Здесь были созданы знаменитые сорта испанского лука, превосходящего и сейчас все известные сорта по сладости и величине. Сорта лука репчатого создавались также и в других странах и вначале были известны по названиям географических мест, где они возникли: в России - Бессоновский (село Бессоновка Пензенской области), Вишенский (село Вишенки Горьковской области), Мячковский (село Мячково Московской области); в Западной Европе - Эрфуртский, Нюрнбергский, Страссбурский и др. Сейчас лук репчатый самый распространенный из луков в культуре. Его возделывают практически везде, где есть земледельческая культура, даже за полярным кругом. Только в СССР известно более 80 его сортов.
Начало культуры чеснока относят примерно к 2000 г. до н. э. Именно чесноку род обязан своим названием Allium - так называли чеснок древние римляне. Чеснок почти так же широко используют, как и лук репчатый. В культуре известно около 30 его сортов.
Первоначально луки использовались как пряные растения, возбуждающие аппетит. Значение луков для современного человека еще шире и многообразнее. Лук и чеснок используют в вареном, жареном (острые и полуострые сорта) и сыром (сладкие или салатные сорта) виде. Чеснок, кроме того, незаменим при солении и мариновании овощей, в колбасном производстве. Средняя норма потребления лука на одного человека в год в различных областях значительно колеблется, но обычно не бывает меньше 6 кг (северные районы), в Средней Азии и Закавказье она составляет 14-17 кг. Значение луков как витаминоносных растений также было очень давно осознано человеком (как противоцинготное). Сейчас хорошо известно, что все съедобные луки, особенно их зеленая часть, отличаются высоким содержанием витамина С: в луковицах - от 12 до 30 мг на 100 г сырой массы, в листьях - от 25 до 90 мг (суточная потребность человека в витамине С составляет около 60 мг). В зеленом луке содержатся также каротины, витамины В1, В2 и РР, но в небольшом количестве. В чешуях лука содержится витамин Р. Луки широко известны с древних времен как лекарственные растения. Свидетельства этого есть в сочинениях Диоскорида, Авиценны. Наиболее известны их бактерицидные свойства, которые давно используют в народной медицине. Современная медицина использует препараты (их около 10) из различных видов луков. Наиболее распространенные из них - аллилчеп и аллилглицер. Главное их назначение - лечение инфекционных заболеваний, кроме того, они усиливают двигательную и секреторную деятельность. Маннит - продукт, предназначенный для питания диабетиков,- выделен из лука в 1957 г.
Других представителей семейства луковых также используют в пищу, в народной медицине (тульбагии, агапантусы, нотоскордумы и др.), но они имеют только местное значение.
Среди луковых очень много красивоцветущих растений, но использование их в декоративном садоводстве ограничивает луковый или чесночный запах. Тем не менее целый ряд видов нашел широкое применение в садах, парках, в оранжереях и комнатном цветоводстве. Очень популярны как садовые растения агапантусы, тульбагии, особенно у себя на родине в Африке, где выращивают почти все виды, прежде всего агапантус восточный. В Европе наиболее широко распространен агапантус зонтичный (Agapanthus umbellatus). Очень красивы тульбагии. Среди них наиболее широко распространенная тулъбагия душистая (Т. fragrans) - единственный вид среди тульбагий без чесночного запаха. Используются как декоративные многие виды луков, нотоскордумов, бродией и других родов луковых.
Семейство гемерокаллисовые (Hemerocallidaceae) (Г. Л. Кудряшова)
Семейство гемерокаллисовых включает род гемерокаллис (Hemerocallis), чаще называемый красоднев или лилейник (около 15 видов), и, вероятно, также род левкокринум (Leucocrinum). Семейство впервые было выделено в качестве самостоятельного знаменитым английским ботаником Робертом Брауном в 1810 г.
Виды рода гемерокаллис - обитатели умеренно теплых районов Восточной Азии. Они распространены в Китае, на полуострове Корея и в Японии, а также на советском Дальнем Востоке и в Сибири. Дальше всех на северо-запад (бассейн верховьев реки Оби в Западной Сибири) проникает гемерокаллис желтый (Hemerocallis lilio-asphodelus, или H. flava). Европейские ботаники (А. Энглер, Г. Хеги и др.) приводили два вида гемерокаллисов-гемерокаллис желтый и гемерокаллис рыжий (H. fulva) - также и для флоры Европы. Другие ботаники, особенно американские и китайские, считали, что эти виды с незапамятных времен натурализовались в Европе, а затем и на других континентах и приобрели там вторую родину. Согласно последним данным ирландского ботаника Д. А. Уэбба (1981), в Европе в диком состоянии встречается только один вид - гемерокаллис желтый (предгорья Юго-Восточных Альп и соседние равнины).
Типичные местообитания большинства гемерокаллисов - негустые леса, кустарниковые заросли, лесные и пойменные луга. Они хорошо выносят затенения лесов и высокотравий, прекрасно растут и на открытых местах. Гемерокаллис желтый, например, в изобилии растущий на Дальнем Востоке СССР, предпочитает здесь хорошо увлажненные, богатые гумусом почвы, где его заросли образуют желтые ковры. В Западной Сибири гемерокаллис желтый встречается на более сухих местах - степных и каменистых склонах гор. Левкокринум горный (Leucocrinum montanum) распространен на западе Северной Америки (от Орегона и Северной Калифорнии до Колорадо и Нью-Мексико), в Скалистых горах.
Гемерокаллисовые - многолетние травянистые растения высотой от 10-20 (левкокринум, рис. 51, 7) до 50-100 см (гемерокаллис, рис. 51, 2), с подземным корневищем, от которого у многих видов отрастают ползучие побеги (рис.51, 2). Корни мочковатые, шнуровидные, часто с мясисто утолщенными шишковидными образованиями. Листья приземные, расположены в два круга, линейные, на конце острые (гемерокаллис) или тупые (левкокринум), цельнокрайные, с выдающимися жилками, прямые или дуговидно изогнутые. В умеренных районах листва большинства гемерокаллисов увядает осенью или погибает от морозов ранней зимой и растение доживает до весны в состоянии покоя в то время как у некоторых видов, например у гемерокаллиса золотистого (Hemerocallis aurantiaca), листья остаются зелеными до следующей весны, особенно в районах с мягкой зимой. Из середины двурядного круга, образуемого красивыми листьями гемерокаллисов, поднимается цветонос, равный по высоте листьям или превышающий их, редко более низкий, чем листья; у левкокринума горного одиночные цветки на цветоножках разной длины, выходящих из пазух листьев и скученных при их основании. Соцветие у гемерокаллисов метельчатое, обычно из 10-12 цветков, с небольшими листовидными или пленчатыми прицветниками при основании цветоножек, или цветки почти сидячие, скученные на конце цветоноса в головковидное соцветие; иногда цветков лишь несколько или даже один, однако есть виды, у которых количество цветков в соцветии доходит до 100 (гемерокаллис многоцветковый - Н. multiflora). Цветки актиноморфные (левкокринум) или слабо зигоморфные (гемерокаллис, рис. 51, 3), белые (левкокринум), различных оттенков желтого, оранжевого, иногда красноватого тона (гемерокаллис), часто с приятным запахом, очень похожи на цветки лилий, воронковидные, длиной от 2-2,5 см у левкокринума до 10 см и более у гемерокаллисов, довольно широко раскрытые, при основании с хорошо выраженной узкой трубкой. Сегменты околоцветника расположены в два круга; внутренние сегменты отличаются от наружных большей шириной, более нежной консистенцией и волнистым краем (гемерокаллис). Тычинок 6, они короче околоцветника или даже скрыты в его трубке, прикреплены к последней, с крупными пыльниками, желтыми, оранжевыми, темно-бурыми, почти черными. Столбик нитевидный, изогнутый, намного превосходящий по длине тычинки, с головчатым рыльцем; завязь верхняя, 3-гнездная. Плод - сухая, кожистая, растрескивающаяся, трехкамерная коробочка, иногда поперечно морщинистая (у гемерокаллиса желтого, например, рис. 51, 5). Семян по 9-12 в каждом гнезде; они яйцевидные, слегка угловатые, черные, блестящие, длиной до 4 мм (рис. 51, 6).
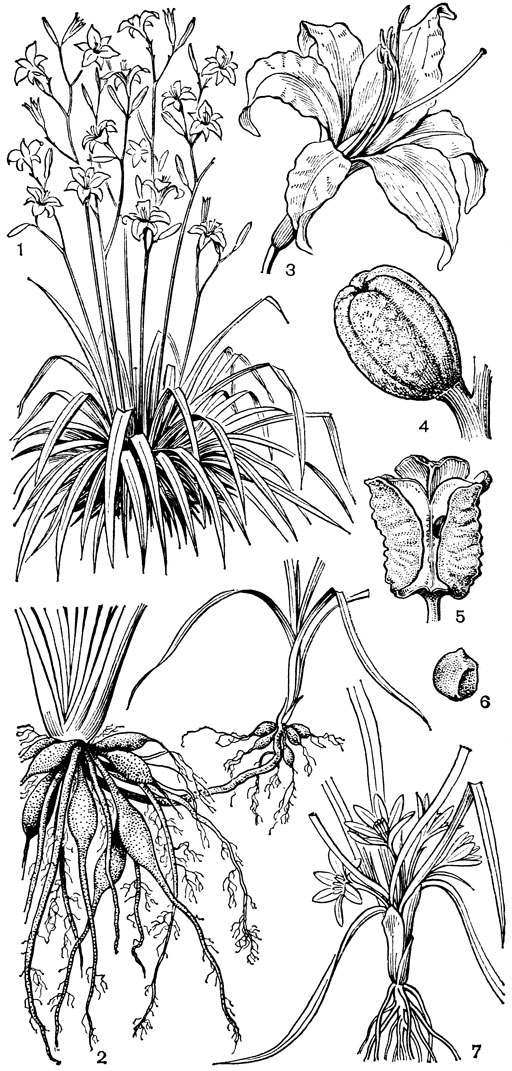
Рис. 51. Гемерокаллисовые. Гемерокаллис рыжии (Hemerocallis fulva): 1 - общий вид; 2 - корневище с клубневидно утолщенными корнями и ползучими побегами, с помощью которых растение вегетативно размножается; 3 - цветок; 4 - коробочка. Гемерокаллис желтый (Н. lilio-asphodelus): 5 - раскрывшаяся коробочка; 6 - семя. Левкокринум горный (Leucocrinum montanum): 7 - общий вид
Название "гемерокаллис" отражает кратковременность цветения этих растений (лат. hemera - день, callos - красота). Цветок увядает на следующий день после распускания, иногда даже еще раньше (гемерокаллис рыжий), но иногда цветки живут и дольше, почти 6 суток (гемерокаллис желтый), особенно в пасмурную погоду при низкой температуре. Несмотря на эфемерность отдельного цветка гемерокаллисов, во время цветения на растении всегда есть раскрытые цветки, так как распускание их происходит в строгой последовательности на каждой оси соцветия, и в целом период цветения растения достигает 2-5 недель (в зависимости от количества цветков в соцветии). Цветут гемерокаллисы с мая по сентябрь (октябрь), при этом большинство видов летом - с июня по август. Среди гемерокаллисов есть виды, цветущие днем, например широко распространенный в культуре гемерокаллис рыжий, гемерокалис золотистый; есть виды, цветки которых распускаются только ночью (гемерокаллис лимонно-желтый - Н. citrina). Цветки у первых раскрываются после восхода солнца и остаются открытыми в течение всего дня, при этом доли околоцветника постепенно отгибаются назад; перед заходом солнца, до наступления темноты, цветки быстро закрываются и вскоре опадают. Цветки гемерокаллисов ночного типа цветения раскрываются незадолго до захода солнца, а закрываются на следующее утро после рассвета. В прохладную и облачную погоду цветки остаются раскрытыми в течение некоторого времени на следующий день. В Ленинграде в период белых ночей цветки гемерокаллиса лимонно-желтого не успевают полностью раскрыться.
Яркие нарядные душистые цветки гемерокаллисов опыляются насекомыми. Септальные нектарники расположены в стенках нижней части завязи и открываются наружу между плодолистиками. Нектар скапливается на дне трубки цветка и доступен только длиннохоботковым насекомым, которых очень много в Восточной Азии, но совершенно нет в Европе. Посещают цветки гемерокаллисов также насекомые, питающиеся пыльцой. В Америке опылителями многих видов гемерокаллисов стали колибри. Большая разница в длине тычинок и столбика, который значительно возвышается над тычинками, препятствует самоопылению. Обычно гемерокаллисы образуют много семян и прекрасно ими размножаются. Распространяются семена ветром и животными. Многие виды размножаются вегетативно ползучими побегами от корневища. Для гемерокаллиса рыжего это вообще единственный способ размножения, так как он абсолютно стерилен в Европе и Америке, где он широко распространился в культуре и одичал. Причиной этого является, по-видимому, то, что все растения его представляют собой клон от одного экземпляра, завезенного в незапамятные времена из Китая. Завезенное растение, размножившееся вегетативно и распространившееся по всему свету за много столетий, было триплоидным, что и является причиной его неспособности давать семена.
Некоторые виды гемерокаллисов уже давно вошли в культуру. Описания гемерокаллисов приводятся в "Естественной истории" Плиния, а Диоскорид сообщает и о медицинских свойствах их листьев и цветков. В I в. н. э. гемерокаллис желтый уже хорошо известное садовое растение. Медицинские свойства гемерокаллисов были давно известны в Китае, о чем упоминается во многих древних китайских травниках. В наши дни, однако, гемерокаллисы используют почти исключительно как декоративные и пищевые растения. Сухие цветочные почки гемерокаллиса желтого употребляют в пищу как овощи и особенно охотно как приправу к мясу, которая считается утонченным деликатесом в Китае. Для этих целей цветочные почки гемерокаллиса заготавливают там в огромных количествах, и их можно найти в любой деревенской лавке. Эти сморщенные, темно-коричневые, прозрачные, сухие почки при приготовлении издают чудесный аромат. Употребляют их также с пшеном, соей. В других странах гемерокаллисы используют только как декоративные растения. Их выращивают везде, за исключением некоторых тропических областей. Эти красивые растения так широко распространились благодаря тому, что они легко приспосабливаются к любым климатическим условиям, обладают высоким иммунитетом, легко культивируются и размножаются. В 90-х годах прошлого столетия известные в культуре виды гемерокаллисов, главным образом гемерокаллис желтый, гемерокаллис рыжий, гемерокаллис Миддендорфа (Н. middendorfii), гемерокаллис малый (Н. minor), гемерокаллис Дюмортъе (Н. dumortierii), стали объектами опытно-исследовательских селекционных работ. К началу XX в. число известных садовых сортов дошло до 100, в настоящее время их свыше 10 000.
Семейство амариллисовые (Amaryllidaceae) (З. Т. Артюшенко)
Амариллисовые включают около 70 родов и более 1000 видов, распространенных на всех континентах, кроме Антарктиды. Большинство из них произрастает в тропической и субтропической зонах и лишь некоторые виды заходят в зону умеренного климата, например подснежник белоснежный (Galanthus nivalis), который встречается в самой северной точке ареала семейства, в окрестностях Киева. Наибольшее разнообразие амариллисовых сконцентрировано в трех местах: в Центральной и Южной Америке, тропической и Южной Африке и Средиземноморье. Особенно интересна в отношении видового разнообразия Южная Африка.
Амариллисовые произрастают в самых разнообразных местообитаниях, но большинство приурочено к гористым местам, от подножий гор и до высоты 4000-5000 м. В ряде мест амариллисовые играют заметную роль в растительном покрове.
Амариллисовые - многолетние травянистые растения. Высота их варьирует от нескольких сантиметров до 2 м. Многолетней частью всех амариллисовых является луковица или очень редко клубнелуковица (только у рода иксиолирион - Ixiolirion). Находятся они или под землей, как у представителей умеренных широт, или на поверхности почвы, как у ряда обитателей тропической зоны. Луковицы весьма разнообразны. Они различаются по форме, размеру, окраске чешуй, а также по внутреннему строению. Форма луковиц варьирует от яйцевидной, продолговато-яйцевидной до почти цилиндрической, а размер - от нескольких до десятков сантиметров. Очень важен характер чешуй, формирующих луковицу. Это или основания ассимилирующих листьев, разрастающиеся ко времени отмирания листовых пластинок, или низовые листья, никогда не несущие листовых пластинок. Кроме того, чешуя может быть замкнутой или туникатной, полутуникатной и черепитчатой. Второй характерной чертой луковицы является количество слагающих ее чешуи. Каждый новый прирост луковицы несет более или менее определенное число листьев, за которыми следует цветонос с цветками, если луковица взрослая. Следовательно, каждый новый прирост несет серию листьев и цветонос. У разных родов и даже видов листовая серия состоит только из ассимилирующих листьев, например у гиппеаструма (Hippeastrum), или, помимо зеленых листьев, образуются низовые, например у подснежника (Galanthus). Количество тех и других в каждой серии для каждого рода и даже вида более или менее постоянно и является хорошим отличительным признаком. Амариллисовым свойственны два типа ветвления побега: моноподиальный и симподиальный. Первый тип встречается чаще у растений умеренных широт, а второй - у субтропических и тропических. На схематическом рисунке (рис. 52) изображены луковицы белоцветника весеннего (Leucojum vernum) и гиппеаструма дворцового (Hippeastrum aulicum), причем для большей наглядности луковицы представлены с вытянутыми междоузлиями. Если у луковиц умеренных широт в течение вегетационного периода закладывается одна листовая серия с цветоносом, которая реализуется во время будущей вегетации растений, то у субтропических и тропических растений формирование листовых серий идет непрерывно, и во взрослой луковице можно насчитать до 6-8 листовых серий с цветоносами, из которых ежегодно реализуются только 2-3, а остальные продолжают медленно развиваться, например у кринума (Crinum).
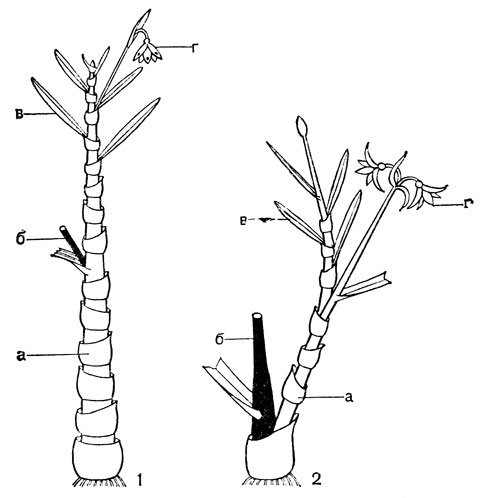
Рис. 52. Типы ветвления у амариллисовых: 1 - белоцветник весенний (Leucojum vernum) с моноподиальным ветвлением; 2 - гиппеаструм дворцовый (Hippeastrum aulicum) с симподиальным ветвлением (а - мясистые чешуи, б - отмерший цветонос, в - вегетирующие листья, г - цветонос с цветком)
Листья амариллисовых собраны в приземной розетке, за исключением иксиолириона, у которого 2-3 листа приземные, а остальные стеблевые. Листья, как правило, сидячие, но у некоторых растений они с хорошо выраженным черешком, который у основания переходит в расширенное влагалище, формирующее чешую луковицы, как у эвхариса (Eucharis), гемантуса (Haemanthus) и др. Листорасположение очередное, причем у большинства двурядно-очередное. У многих листья плоские, линейные или нитевидные, но встречаются и вальковатые, образовавшиеся в результате срастания краев листа, завернутых кверху. Срастание происходит на ранних этапах развития листа. Свидетельством этого служит его анатомическое строение, показывающее, что проводящие пучки в листе расположены не в один ряд, а кольцом, с ксилемой, обращенной внутрь кольца, а флоэмой - наружу. Листья обычно плотные, кожистые, с хорошо развитой кутикулой и устьицами, расположенными на верхней и нижней поверхностях листа. Часто листья покрыты восковым налетом, придающим им сизоватый оттенок. Жилкование дуговидное, центральная жилка часто образует хорошо выраженный киль на нижней поверхности листа. Размер листьев колеблется от нескольких сантиметров до метра и более. В листьях многих амариллисовых, в полостях, расположенных между проводящими пучками, образовавшихся в результате облитерации клеток, содержится большое количество слизей, свободно вытекающих из листа при его повреждении. В слизи содержится много рафид и алкалоидов. Помимо листьев, алкалоиды содержатся и в других частях растения, в цветоносах и чешуях.
У большинства амариллисовых надземный стебель представлен безлистным цветоносом или облиственным стеблем (иксиолирион). Он полый внутри или выполнен паренхимной тканью. По форме цветонос бывает округлым или сплюснутым, заостренным к обоим концам. На верхушке цветоноса расположены два прицветника, охватывающих основание цветоножки. Прицветники могут быть свободными, как у кринума и гиппеаструма, или со сросшимися краями, преимущественно у одноцветковых амариллисовых, таких, как подснежник, нарцисс (Narcissus, табл. 14, 5), парамонгайя Вебербауэра (Paramongaia weberbaueri). В последнем случае края прицветников пленчатые и перед цветением разрываются с одной стороны бутоном. У многоцветковых амариллисовых прицветники первого цветка соцветия хорошо развиты, тесно соприкасаются друг с другом краями, закрывая все соцветие. Прицветники у основания часто срастаются в трубку, охватывающую завязь и основание цветоножки. Верхняя их часть остается свободной и называется покрывалом или крылом, так как полностью закрывает молодой цветок (бутон) или все соцветие (рис. 53). К началу цветения цветок или соцветие разрывает покрывало, которое остается на цветоносе до полного созревания плодов и отмирает вместе с цветоносом. Цветоножка, расположенная в пазухе каждой пары прицветников, бывает разного размера - тонкой или толстой, короткой или длинной.
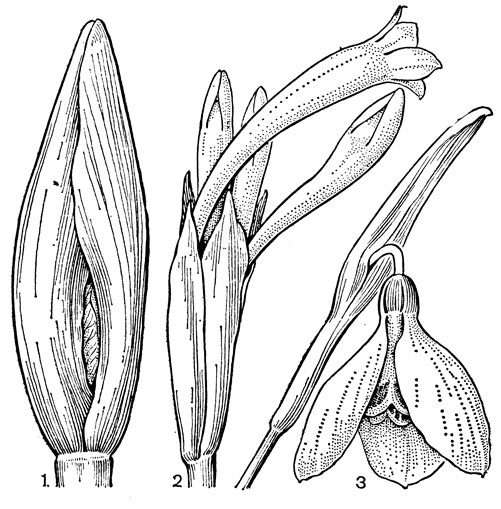
Рис. 53. Характер покрывала у амариллисовых: 1 - гиппеаструм дворцовый (Hippeastrum aulicum) с покрывалом из двух свободных прицветников; 2 - циртантус желто-белый (Cyrtanthus ochroleucum) с покрывалом из нескольких свободных прицветников; 3 - подснежник Воронова (Galanthus woronowii) с покрывалом из двух сросшихся прицветников

Таблица 14. Амариллисовые: 1 - ликорис золотистый (Lycoris aurea); 2 - гиппеаструм пришлый (Hippeastrum advenum); 3 - нарцисс крупный (Narcissus major); 4 - нарцисс узколистный (N. angustifolius); 5 - шпрекелия прекраснейшая (Sprekelia formosissima); 6 - урцеолина повислая (Urceolina pendula); 7 - кринум капский (Crinum capense); 8 - белоцветник летний (Leucojum aestivum)
Цветки амариллисовых обычно собраны в более или менее ясно выраженные зонтики, иногда завитки, как у белоцветника летнего (Lencojimi aestivum, табл. 14, 8), снабженные покрывалом из 2 или больше (редко 1) обычно пленчатых прицветников. Соцветие-кисть ветречается редко (род иксиолирион). В семействе ясно выражена тенденция к редукции соцветия от зонтика к одиночным цветкам (табл. 14-17). В культуре у некоторых амариллисовых, например гиппеаструма, при хорошем уходе вместо 4 цветков в зонтике образуется до 8, а при плохих условиях - только 2, а иногда 1 цветок.
Цветки прямостоячие, изогнутые или поникающие, обоеполые, лишь в редких случаях вследствие недоразвития гинецея или андроцея однополые (например, у кливии благородной - Clivia nobilis, табл. 16, 3). У единственного представителя амариллисовых, произрастающего в Мексике,- шпрекелии прекраснейшей (Sprekelia formosissima, табл. 14, 5) - зигоморфность цветка выражена очень сильно и ее околоцветник типично двугубый. Сегменты околоцветника, расположенные в 2 круга, свободные или сросшиеся в трубку. Трубка околоцветника длиной от нескольких миллиметров до 10 см, узкая или расширяющаяся к зеву, цилиндрическая или воронковидная. Тычинки расположены в 2 круга, но в зигоморфных цветках они собраны в пучок и часто образуют 3 ряда по 2 тычинки в каждом ряду. Нити тычинок равны между собой или тычинки наружного круга короче; в зигоморфных цветках, например у габрантуса (Habranthus), тычиночные
нити расположены тремя парами, причем наружные нити длиннее внутренних. Длина тычиночных нитей варьирует от нескольких миллиметров у подснежника и белоцветника до 10 см у гемантуса и кринума. Нити тычинок могут быть прямыми или изогнутыми, нитевидными или утолщенными, расширенными и даже крылатыми, как у иеронимиеллы (Hieronymiellа). Пыльники большей частью продолговатые, но бывают и копьевидные, как у подснежника и лапиедры (Lapiedra), или шаровидные, как у гессеи (Hessea) и карполизы (Carpolyza). Прикрепляются пыльники к тычиночной нити по-разному: основанием (тогда пыльник прямой или изогнутый и неподвижный), вблизи основания (тогда он подвижный), серединой спинки (тогда он качающийся и от легкого движения воздуха сильно раскачивается, высыпая пыльцу). Раскрываются пыльники или продольными щелями от верхушки до основания, или порообразными щелями, как у белоцветника. Щели обращены к гинецею, открываются внутрь цветка. Пыльцевые зерна одно- или двубороздные, иногда с трехлучевой бороздой; экзина сетчатая или шиповатая.
Для многих амариллисовых характерно наличие в цветке особых образований, возвышающихся в виде трубки или небольшой оборочки над зевом околоцветника. Это так называемые корона и привенчик, которые имеют двоякое происхождение. Корона образуется из разросшихся оснований тычиночных нитей, причем они или срастаются между собой, как у гименокаллиса корзинкового (Hymenocallis calathina), элизены длиннолепестной (Elisene longipetala), парамонгайи Вебербауэра, или остаются несросшимися в виде зубцов, как у панкратиума иллирийского (Pancratium illiricum), калифрурии Хартвега (Caliphruria hartwegiana) и др. (рис. 54). Привенчик образуется из выростов сегментов околоцветника, которые также могут срастаться между собой, образуя трубку разной формы и размера, как у нарцисса крупного (Narcissus major, табл. 14, 3) и нарцисса поэтического (N. poёticus), или выросты в виде чешуевидных зубчиков, как у шпрекелии прекраснейшей, ликориса золотистого (Lycoris aurea) и др. (табл. 14, 2, 5). Корона и привенчик могут быть трубчатыми или блюдцевидными, с более или менее ровным, а чаще с гофрированным или разделенным на лопасти или чешуйки краем. Корона и привенчик придают цветку особую привлекательность и являются, кроме того, хорошим отличительным признаком (табл. 14-15).

Рис. 54. Амариллисовые: 1 - элизена длиннолепестная (Elisene longipetala) с узкими длинными сегментами и цилиндрической короной; 2 - панкратиум иллирийский (Pancratium illiricum) с широкими сегментами и несросшимися зубцами короны; 3-8 - парамонгайя Вебербауэра (Paramongaia weberbaueri) с широкими сегментами, воронковидной короной и частями цветка (4 - гинецей; 5 - продольный и 6 - поперечный разрезы завязи; 7 - лист; 8 - поперечный разрез листа); 9 - гименокаллис корзинковый (Hymenocallis calathina) с узкими сегментами и бокаловидной короной с отогнутым краем; 10 - семя; 11, 12 - калифрурия Хартвега (Caliphruria hartwegiana) (11 - общий вид цветка; 12 - разрез цветка с несросшимися зубцами короны в зеве)
Гинецей из 3 сросшихся плодолистиков; завязь нижняя, 3-гнездная, в каждом гнезде от одного (как у гемантуса) до многих семязачатков. Столбик нитевидный, вздутый, у некоторых видов треугольный у основания, угловатый или крылатый с 3-лопастным, 3-раздельным или головчатым рыльцем. Плод у амариллисовых - локулицидная коробочка, иногда сочная, ягодообразная. Семена с мясистым эндоспермом, окружающим маленький зародыш, плоские, угловатые или крылатые, легко разносятся ветром. У некоторых представителей семена имеют мясистый, сочный придаток - элайосому, который поедают муравьи, растаскивающие семена.
Цветки у многих амариллисовых протандричны, но некоторые протогиничны. Например, у гиппеаструма полосатого (Hippeastrum vittatum, табл. 17, 1) рыльце созревает на 24 ч позже раскрывания пыльников. У некоторых растений наблюдается гетеростилия. Так, у нарцисса и подснежника встречаются 3 формы цветка - длинностолбчатая, короткостолбчатая и с тремя фертильными тычинками. Амариллисовые, как правило, перекрестноопыляемые растения, но иногда у них происходит и самоопыление. Это явление чаще всего можно обнаружить у пониклых цветков типа подснежника, у которого тычинки перед пониканием цветка возвышаются над гинецеем и из конусовидных пыльников пыльца попадает прямо на рыльце. Иногда при посещении цветка опылитель стряхивает пыльцу из пыльников на рыльце, и тогда также происходит самоопыление. В ряде случаев вследствие неравномерного роста тычиночных нитей пыльники многократно соприкасаются с рыльцем и опыляют его. Некоторые насекомые и птицы берут нектар из цветков, порхая над ними. Взмахи крыльев создают поток воздуха, под действием которого пыльца осыпается на рыльце и осуществляется самоопыление. Опыляются амариллисовые бабочками, пчелами, шмелями, мухами и птицами. Для привлечения опылителей у амариллисовых в цветке имеется ряд приспособлений: необычайно яркая окраска, пятна и полоски на сегментах околоцветника, сильный запах и наличие нектарников или нектароносной ткани. Корона и привенчик цветка служат прекрасным опознавательным знаком для опылителей; зеленое пятно на внутренних и наружных сегментах околоцветника белоцветника и подснежника обозначает вход для насекомых, оно отличается более сильным запахом. Более сильным запахом обладает также красная каемка привенчика ряда нарциссов, например нарцисса поэтического.
Для цветков амариллисовых характерно обильное выделение нектара. У гемантуса, нарцисса, кливии и других цветочная трубка до половины и более заполнена нектаром, а у таких растений, как панкратиум (Pancratium), гименокаллис (Hymenocallis), шпрекелия (Sprekelia), нектар заполняет цветочную трубку доверху и часто выливается из нее, стекая по тычиночным нитям (рис. 55). Нектарники и нектароносная ткань - эффективное приспособление для привлечения опылителей, не только насекомых, но и птиц. Наиболее обычны септальные нектарники, приуроченные к перегородкам завязи и нижней части столбика; у многих нектарники находятся у основания сегментов околоцветника. Нектароносная ткань обычно располагается на дне цветочной трубки на верхушке завязи. Она состоит из железистой ткани, сверху покрытой толстой кутикулой. При выделении нектара кутикула разрывается и нектар вытекает через щели. Небольшое количество нектара выделяется в бороздках на внутренней стороне зеленого пятна сегментов околоцветника, как у подснежника и белоцветника (Leucojum), но основная функция пятна - указывать вход для опылителей.
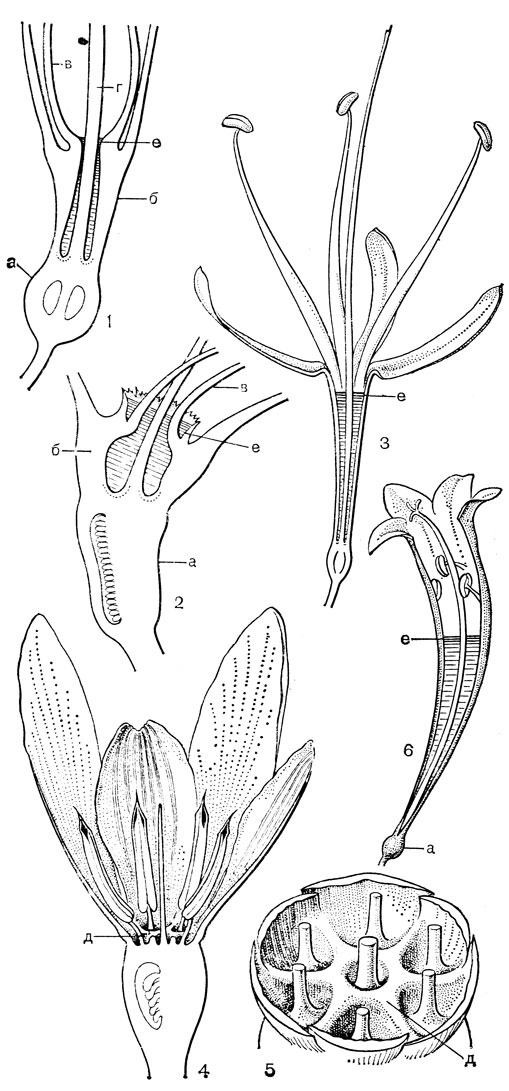
Рис. 55. Нектарники амариллисовых: 1 - кливия благородная (Clivia nobilis); 2 - гиппеаструм дворцовый (Hippeastrum aulicum); 3 - гемантус Екатерины (Наеmanthus katharinae); 4 - подснежник киликийский (Galanthus cilicicus); 5 - он же, часть нектарного диска (тычинки и столбик обрезаны); 6 - циртантус Макована (Cyrtanthus makowanii) (а - завязь, б - трубка околоцветника, в - тычинки, г - столбик, д - нектароносная ткань, е - увеличено, ж - уровень нектара в трубке околоцветника)
У некоторых амариллисовых, например у нарцисса, опыление производят опылители, посещающие цветки не только из-за нектара, но и ради пыльцы, которую они поедают. Особенно охотно посещаются цветки, у которых пыльца склеивается в комочки вязким веществом, как у подснежника. При легком прикосновении насекомого к цветку, направленному вниз, комочки пыльцы высыпаются на его спинку и легко транспортируются.
Большое значение при опылении имеют корона и привенчик. Если привенчик короткий блюдцевидной формы, как у нарцисса поэтического, насекомые садятся на него и просовывают в трубку голову; если привенчик длинный, с отогнутым гофрированным краем, то насекомые садятся на край, который играет роль посадочной площадки. В зависимости от длины короны или привенчика цветки опыляются коротко- хоботковыми или длиннохоботковыми насекомыми. В других случаях насекомые или птицы садятся на цветонос и высасывают нектар снизу из свешивающихся цветков; если соцветие с многочисленными прямостоячими цветками и расположено на крепком цветоносе, опылители садятся на соцветие или порхают над ним, высасывая нектар, как у гемантусов.
Запах цветка также играет большую роль для привлечения опылителей. Душистые цветки посещаются преимущественно в утренние и вечерние часы, когда их запах особенно интенсивен. В некоторых случаях запах усиливается ночью. Так, длиннотрубчатые цветки кринума и панкратиума особенно любят посещать в вечерние и ночные часы длиннохоботковые бабочки, привлекаемые сильным запахом цветков.
Многие амариллисовые опыляются птицами, привлекаемыми обильно выделяющимся нектаром. Основным представителем орнитофильных амариллисовых является шпрекелия прекраснейшая, у которой все части цветка выделяют нектар. Его в цветке так много, что он заполняет его трубку и нижняя часть тычинок и пестика погружена в нектар. Птицы опыляют также виды гемантуса, кливии, кринума, гименокаллиса, эвхариса, эвкрозии (Eucrosia, табл. 15, 3), циртантуса (Cyrtanthus, табл. 17, 7) и др.
Таблица 15. Амариллисовые: 1 - калостемма желтая (Calostemma lutea); 2 - кринум погруженный (Crinum submersum); 3 - эвкрозия двуцветная (Eucrosia bicolor); 4 - иксиолирион горный (Ixiolirion montanum)
Строение и способы распространения плодов и семян амариллисовых также разнообразны. У многих плод - сухая коробочка, из которой легкие уплощенные семена рассеиваются при малейшем дуновении ветра. Примером могут служить унгерния (Ungernia), гиппеаструм, валлота (Vallota) и др. У ряда растений коробочка мясистая или даже ягодообразная. В одних случаях с созреванием плода цветонос, теряя тургор, полегает на почву вместе с коробочкой; последняя раскрывается по гнездам, ткани околоплодника высыхают и скручиваются, а семена тут же рассыпаются. Такое вскрытие коробочек наблюдается у подснежников, белоцветников. В других случаях мясистую коробочку разрывают прорастающие в ней семена. С образованием корешка и первого листа из разорванной коробочки проросшие семена падают на почву, укореняются и продолжают развиваться. Это наблюдается у кливии матово-красной (Clivia miniata), панкратиума карибского (Pancratium caribaeum) и др. У кринумов и амариллиса (Amaryllis) околоплодник коробочки бумажистый. Он также часто разрывается увеличивающимися в объеме семенами, но последние прорастают не в коробочке, а лишь после освобождения из нее, на почве.
Семена амариллисовых разнообразны. Они округлые, угловатые, с небольшим пленчатым крылом, окаймляющим семя и способствующим перенесению его на значительные расстояния. Но особенно интересны семена с элайосомами, как, например, у подснежника и белоцветника. Этот придаток семени охотно поедают муравьи, которые растаскивают семена по подземным ходам, способствуя тем самым их распространению. Не менее примечательны семена кринума и амариллиса. У них сильно разрастается эндосперм, который ко времени созревания становится сочным и слегка зеленеет снаружи. Сквозь тонкий пленчатый околоплодник хорошо просвечивают зеленоватые крупные семена с очень маленьким зародышем.
Ягодовидные мясистые коробочки боофоны (Boopliane) и гемантуса вместе с семенами опадают с зонтиковидного соплодия на - почву и охотно поедаются животными, способствующими их распространению.
Семейство амариллисовых подразделяется на два очень неравных по объему подсемейства - амариллисовые (Amaryllidoideae) и иксиолирионовые (Ixiolirioideae). Второе подсемейство состоит из одного только рода иксиолирион, который так заметно отличается от остальных амариллисовых, что японский ботаник Т. Накаи (1945) даже выделил его в самостоятельное семейство иксиолирионовых (Ixioliriaceae). Несмотря на то что иксиолирион (табл. 15, 4), в отличие от остальных амариллисовых, для которых характерны луковица и безлистный цветонос, имеет клубнелуковицу и облиственный стебель, несущий на верхушке соцветие,- тип гинецея, строение цветка, кариотип позволяют включить его в семейство амариллисовых.
Подсемейство амариллисовых делится на ряд триб, объем которых до настоящего времени твердо не установлен. Наиболее известны системы, предложенные Г. Траубом (1957, 1963), Д. Хатчинсоном.(1960) и Г. Мельхиором (1964). Последний в основу разделения подсемейства на трибы положил систему Трауба, опубликованную в 1957 г., значительно ее переработав. Новые данные по исследованию вегетативных и репродуктивных органов амариллисовых показали, что ни одна из предложенных систем не может удовлетворить исследователей этого семейства.
Характерной чертой цветка амариллисовых является наличие или отсутствие короны и привенчика, возникших в процессе приспособления к опылителям. В соответствии с этим признаком трибы делятся на две группы, а внутри групп распределяются в соответствии с признаками, свидетельствующими об их примитивности или продвинутости.
Более примитивными следует признать трибы, представителям которых свойственны цветки без короны и привенчика, многочешуйчатые луковицы, состоящие из многих листовых циклов, реализующихся в течение нескольких лет, и цветоносы с зонтиковидными соцветиями, окруженные свободными прицветниками; многие из них отличаются непрерывной вегетацией. Среди этой группы триб самой примитивной является триба криновых (Crineae). Эта самая крупная по объему триба семейства, насчитывающая 10 родов и около 200 видов, распространенных в тропических и субтропических областях обоих полушарий. Для представителей этой трибы характерен многоцветковый зонтик, окруженный свободными прицветниками; цветки большей частью крупные, слегка зигоморфные, длиннотрубчатые, с прямой или изогнутой трубкой; семязачатков много в каждом гнезде. К этой трибе относится самый крупный род семейства - кринум (Crinum, табл. 14, 7; 15, 2). Большинство видов кринума (около 40) произрастают в тропической Африке, более 20 видов - в тропической Азии, свыше 10 - в тропической Америке, около 10 видов - в Австралии и Полинезии и до 10 видов - в Капской области. Условия их обитания очень разнообразны: болотистые берега рек и ручьев, заболоченные луга, песчаные берега морских побережий, ручьи, озера, опресненные морские лагуны, травянистые горные склоны, до высоты более 1000 м над уровнем моря. Цветки кринума собраны в зонтике по 1-2, как у кринума одноцветкового (С. uniflorum), произрастающего на открытых песчаных местах Северной Австралии, или до 50 в соцветии, как у кринума азиатского, широко распространенного в тропической части Азии. Крупные белые или бледно-розовые цветки кринумов, располагающиеся на довольно высоком и крепком цветоносе, обладают тонким ароматом. Виды кринума широко используют в цветоводстве. В зарубежных странах с теплым климатом и на юге Советского Союза некоторые его виды культивируют в открытом и полузакрытом грунте; в районах с холодным климатом выращивают в оранжереях, используя для выгонки, для оформления закрытых помещений и как комнатное растение.
Род валлота (Vallota), относящийся к этой трибе, представлен одним видом - валлотой пурпурной (V. purpurea), эндемиком Капской области. У этого растения луковица крупная, яйцевидная, несет ремневидные листья длиной до 60 см, появляющиеся одновременно с цветоносом. Цветонос полый, длиной до 90 см, несет до 10 багряно-красных прямостоячих цветков. Цветки с длинной трубкой и 6 ланцетными сегментами длиной до 5 см и шириной 2,5 см; тычинки достигают половины длины сегментов, столбик одной длины с сегментами. Плод - слегка угловатая коробочка; семена уплощенные, с крылом у основания. Имеются формы с белыми цветками, а также межродовые гибриды. Культивируется повсеместно: в странах с теплым и жарким климатом - в открытом грунте, а с холодным и умеренным - в оранжереях и комнатных условиях.
Род циртантус (Cyrtanthus, табл. 17, 7) включает до 50 видов, произрастающих в тропической и Южной Африке. Приурочены они к горным склонам и долинам. Цветки среднего размера, длиной 3-4 см, прямостоячие или поникающие, со слегка искривленной трубкой; в зонтике до 12 цветков; у некоторых видов зонтики малоцветковые, с 1-3 цветками. Окраска околоцветника преимущественно красных тонов, реже белая и желтая. Тычинки с тонкими нитями и качающимися пыльниками. Столбик длинный, нитевидный, с 3-лопастным рыльцем. Плод - коробочка, с плоскими темными семенами, имеющими у основания крыловидный вырост. В Европе и Америке издавна культивируют несколько наиболее декоративных видов циртантуса. Имеются сорта и гибриды с валлотой.
Род унгерния (Ungernia) представлен 6 видами, распространенными в Центральной Азии и приуроченными к горным массивам Гималаев и Тянь-Шаня до высоты 1300-2300 м над уровнем моря. Они растут на каменистых, щебнистых, глинистых, реже гипсовых склонах в степном поясе гор, на закрепленных песках и выходах коренных пород. Это растения с крупной луковицей, залегающей на глубине более полуметра. Весной появляется пучок ремневидных листьев, а в начале лета, после их отмирания, появляются цветоносы, несущие зонтик цветков. Околоцветник воронковидный, почти правильный, со слегка изогнутой трубкой, с сегментами длиной около 3 см, окрашенными в розовые, желтоватые, коричневатые тона, с полоской другого цвета на внутренней стороне. Тычинки расположены в 2 круга, наружные короче внутренних; продолговатые, качающиеся пыльники; столбик длиной около 3 см, увенчанный головчатым рыльцем. Плод - локулицидная коробочка, несущая черные, угловатые, слегка сплюснутые семена (рис. 56). Виды унгернии не обладают особыми декоративными качествами, в отличие от других амариллисовых, но они имеют большое практическое значение в медицине. Из унгернии выделены алкалоиды, применяющиеся для лечения различных заболеваний.
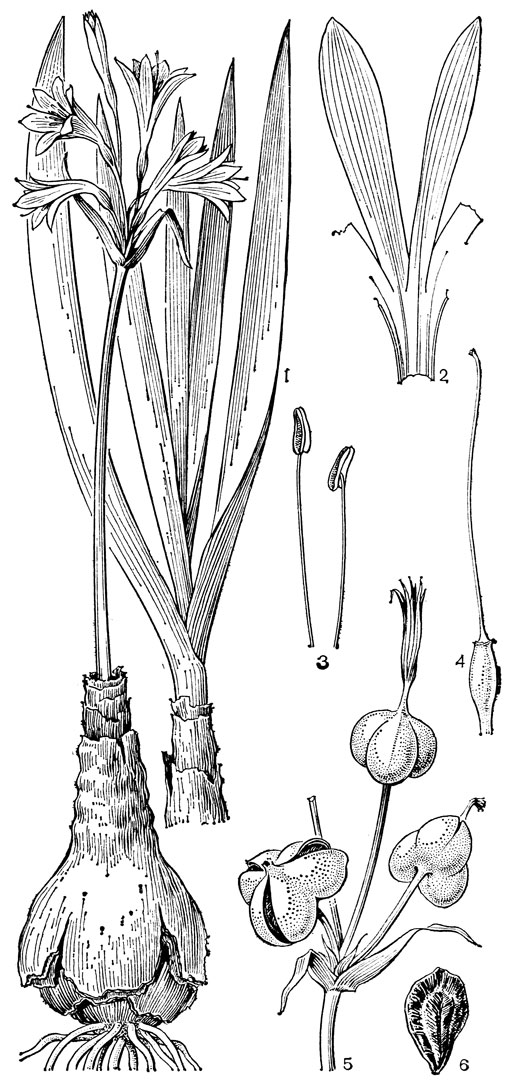
Рис. 56. Амариллисовые. Унгерния трехсферная (Ungernia trisphaera): 1 - луковица, цветонос с цветками и листья. Унгерния Северцова (U. sewertzowii): 2-4 - части цветка; 5 - плоды; 6 - семя
Триба гемантусовых (Haemantheae) включает 8 родов и около 90 видов, приуроченных к Южной Африке, за исключением рода грифиния (Griffinia), произрастающего в Бразилии. Представители этой трибы характеризуются более или менее мелкими цветками, собранными, как правило, в многоцветковый плотный зонтик, окруженный свободными, иногда ярко окрашенными прицветниками. Семязачатки немногочисленные.
Самый крупный род трибы - гемантус (Наеmanthus, табл. 16,2; 17,5). В него входит свыше 50 видов, распространенных в тропической и Южной Африке, а один вид - гемантус крупнолистный (Н. grandifolius) растет в горах острова Сокотра. Произрастают гемантусы в различных экологических условиях: по берегам рек, на каменистых склонах гор, в горах и в лесах. Наибольшее число видов сосредоточено в Капской области. Цветки гемантуса белые или красные, не очень крупные, длиной до 5 см, трубчатые, собраны в густой зонтик, окруженный прицветниками. Плод - ягодовидная разрывающаяся коробочка; семена округлые. В районах Африки с жарким климатом культивируют многие виды, в других странах выращивают в основном три вида: гемантус белоцветковый (Н. albiflos), гемантус ярко-красный (Н. coccineus, табл. 17, 3) и гемантус Екатерины (Н. katharinae, табл. 16, 1). В условиях умеренного и холодного климата все эти виды культивируют только в закрытом грунте.
Одними из излюбленных растений, выращиваемых в помещениях при любых климатических условиях, являются виды рода кливия (Clivia, табл. 16, 5), относящегося к этой же трибе. Этот небольшой род содержит всего 3 или 4 вида, произрастающих в горных районах Южной Африки. Цветки длиной до 3 см собраны в густой, многоцветковый зонтик (до 60 цветков). Околоцветник от желтого до темно-оранжевого цвета, с короткой трубкой, с приросшими в зеве тычинками и с длинным нитевидным столбиком. Плод - разрывающаяся ягодовидная коробочка с единичными семенами, которые часто прорастают в коробочке, разрывая ее при этом. Наиболее часто культивируют кливию матово-красную (С. miniata) и ее многочисленные сорта, создающие пастельную гамму расцветок от лимонно-желтой до красновато-оранжевой.
Триба амариллисовых (Amaryllideae) состоит из 3 родов, распространенных в Южной Африке. Луковицы большие; цветонос с довольно крупными, яркими, слегка зигоморфными цветками. Семязачатков в гнезде много, они погружены в плаценту или на ножках.
Род амариллис (Amaryllis) представлен единственным видом - амариллис красавица (А. bella-donna, табл. 17, 7), произрастающим на юге Африки и являющимся эндемиком Капской области. Это крупное растение с цветками длиной до 10 см, собранными по 4-12 в зонтик, расположенный на высоком крепком цветоносе. Околоцветник от бледной до темно-розовой окраски. Плод - разрывающаяся коробочка, с крупными семенами, снабженными мясистой оболочкой. В зарубежных странах амариллис издавна культивируют; имеется много садовых форм, а также многочисленные межродовые гибриды, созданные в последнее время. От гиппеаструма, который также часто называют амариллисом, амариллис очень хорошо отличается многоцветковым зонтиком, сочной коробочкой, крупными, как у кринума, семенами и рядом других менее броских черт.
Не менее интересен в декоративном отношении и род нерина (Nerine), содержащий около 30 видов, произрастающих в Южной Африке от подножий гор до высоты 1500 м над уровнем моря. Цветки бледно- или темно-красные, длиной до 4 см, с ланцетными сегментами, слегка волнистыми по краю. Ярко-красные тычинки придают цветку особую прелесть. Цветки собраны по 4-6 в зонтике. Коробочка округлая, глубоко-3-лопастная, перепончатая, с небольшим количеством семян в гнездах. Культивируют более 5 видов. В странах с жарким и теплым климатом их выращивают в открытом грунте, с умеренным и холодным - в оранжереях и комнатных условиях.
Триба зефирантесовых (Zephyrantheae) включает 9 родов, из которых 7 приурочены к Южной Америке, 2 - к Южной Африке. Растения этой трибы с мелкими луковицами, небольшими листьями и цветками, расположенными по 2-1 на невысоком цветоносе и окруженными двумя сросшимися прицветниками. Самый крупный род этой трибы - зефирантес (Zephyranthes), включающий до 40 видов, распространенных в субтропической и тропической областях Америки. Это небольшие растения с относительно крупными белыми, розовыми или желтыми цветками, растущие в разных условиях: на заболоченных местах долины реки Ла-Плата, в горах Мексики, на песчаных плоскогорьях Чили. Листья у них линейные, цветонос несет один цветок, окруженный трубкой из двух сросшихся прицветников. Цветок актиноморфный, прямой или слегка наклоненный, с воронковидным околоцветником; сегменты ланцетные, наружные немного шире внутренних, белые, розовые, желтые. Плод - коробочка, с уплощенными черными семенами. Имеются межвидовые и межродовые гибриды, которые культивируются на Западе. Зефирантес крупноцветковый (Z. grandiflora) и зефирантес розовый (Z. rosea) - излюбленные комнатные растения северных широт. Цветонос с цветком у них развивается очень быстро: после появления цветоноса над почвой через 1-2 дня растение уже цветет. За эту особенность любители-цветоводы называют его "выскочкой". В странах с теплым и умеренным климатом зефирантесы прекрасно растут в открытом грунте, занимая в короткое время обширные пространства, и легко дичают.
Близкие к зефирантесу роды - габрантус (Habranthus) и аргиропсис (Argyropsis) сходны по биологии, экологии и декоративным качествам. Чаще всего культивируют габрантус мощный (Habranthus robustus) и аргиропсис белоснежный (Argyropsis Candida). В условиях теплого и умеренного климата их выращивают в открытом грунте, а в условиях холодного - в оранжереях и комнатных условиях.
В трибе подснежниковых (Galantheae) 4 рода и около 40 видов, распространенных в Средней Европе, Средиземноморье, на Кавказе, в Иране, Афганистане и Центральной Азии. Цветки у них одиночные, по 2 или несколько в зонтике; 2 прицветника срастаются основаниями в трубку. Семязачатков много в каждом гнезде; семена у многих видов с элайосомой.
Самый крупный род трибы - подснежник (Galanthus, табл. 17, 2), в состав которого входит 17 видов, распространенных в Средней и Южной Европе, в Малой Азии и на Кавказе. Произрастают подснежники в разных экологических условиях: по опушкам леса, среди кустарников, в предгорьях и горах, поднимаясь на высоту до 2500 м над уровнем моря. В природе некоторые виды занимают значительные площади. Подснежники - небольшие растения, достигающие в высоту 30 см, с единственным поникающим цветком молочно-белого цвета длиной до 3,5 см. Сегменты околоцветника расположены в 2 круга: наружные ланцетные, вогнутые; внутренние почти наполовину короче, клиновидные, на верхушке с вырезкой, окаймленной зеленым подковообразным пятном. У некоторых видов имеется второе зеленое пятно, расположенное у основания внутренних сегментов, например у подснежника греческого (G. graecus) и подснежника византийского (G. byzanthynus). Плод - мясистая локулицидная коробочка, с семенами, снабженными сочным придатком - элайосомой. Из некоторых подснежников был выделен алкалоид галантамин, показавший положительные результаты при лечении ряда заболеваний.
В Европе подснежники культивируют издавна, особенно в Англии. Эти растения легко натурализуются. Они хорошо приживаются в открытом грунте даже в северных районах, например в Ленинграде, Кировске, где их успешно выращивают в Полярно-Альпийском ботаническом саду Кольского филиала АН СССР.
Близкий к подснежнику род белоцветник (Leucojum) содержит около 10 видов, ареал которых простирается от Ирландии через Среднюю и Южную Европу, заходя в Крым и на Кавказ и захватывая северное побережье Африки. Растут белоцветники в разных условиях: на болотистых местах, вдоль ручьев из талого снега, по опушкам леса и в разреженных листопадных лесах, на сухих местах побережья Северной Африки. Это небольшие растения, высотой до 30 см. Цветки также невелики, длиной до 3 см; они расположены на цветоносе по 1-2 или собраны в малоцветковый завиток. Околоцветник чисто-белый, с сегментами одинаковой формы и размера, отмеченными снаружи у верхушки зеленым пятном. Тычинки с белыми нитями и ярко-желтыми конусовидными пыльниками; столбик нитевидный или веретеновидный, с маленьким головчатым рыльцем. Плод - локулицидная мясистая коробочка, несущая семена, у некоторых видов снабженные элайосомой. Белоцветник весенний издавна выращивают в открытом грунте. У белоцветников также обнаружен галантамин, который используют в медицинской практике в Болгарии.
В трибу подснежниковых включен и род штернбергия (Sternbergia, рис. 57, 8-14, табл. 16, 4) с 5 видами, распространенными в Средиземноморье, Малой Азии, на Кавказе, в Иране и в Средней Азии. Виды штернбергии произрастают в предгорных равнинных районах, в нижнем и среднем горных поясах, на каменистых местах и сухих степных склонах. Это небольшие растения, высотой 20-40 см, с розеткой линейных листьев и с одним-двумя (реже тремя) цветоносами, каждый из которых несет по одному цветку, заключенному в покрывало. Цветки актиноморфные, из 6 эллипсоидальных сегментов, причем внутренние немного уже наружных. Околоцветник воронковидный, с небольшой трубкой, окрашенный в сернисто- или ярко-желтый цвет. Тычинки разной длины, расположены в 2 круга; внутренние нити длиннее наружных; столбик с 3- лопастным рыльцем; завязь сидячая или на короткой ножке, 3-гнездная, с многочисленными семязачатками. Плод - мясистая коробочка, раскрывается слабо. Семена темные, округлые, с элайосомой. Виды штернбергии очень декоративны, их можно использовать для весеннего цветения на открытых местах в парках и альпинариях.
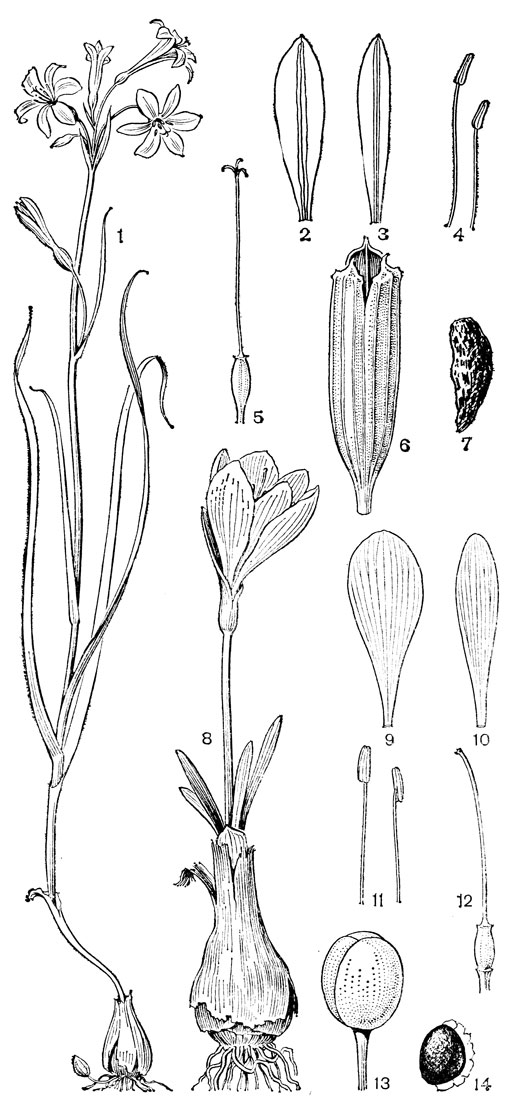
Рис. 57. Амариллисовые. Иксиолирион ферганский (Ixiolirion ferganicum): 1 - общий вид; 2-5 - части цветка; 6 - плод; 7 - семя. Штернбергия желтая (Sternbergia lutea): 8 - общий вид; 9-12 - части цветка; 13 - плод; 14 - семя

Таблица 16. Амариллисовые: 1 - гемантус Екатерины (Haemanthus katharinae), оранжереи Ботанического института АН СССР в Ленинграде; 2 - гименокаллис прибрежный (Hymenocallis littoralis), там же; 3 - кливия благородная (Clivia nobilis), там же; 4 - штернбергия желтая (Sternbergia lutea), Крым, Никитский ботанический сад
В следующую группу амариллисовых входят трибы, представителям которых свойственно наличие в цветке короны или привенчика. Среди них более примитивными следует признать трибы, у представителей которых корона формируется из расширенных оснований тычиночных нитей.
Триба эвстефиевых (Eustephieae) включает 8 родов и свыше 30 видов, приуроченных к Центральной и Южной Америке. Цветки актиноморфные, редко слегка зигоморфные, с длинной или короткой трубкой околоцветника и короной, образованной крыловидными выростами тычиночных нитей. Род эвстефия (Eustephia) состоит из 6 видов, распространенных в Перу и Аргентине. Произрастают преимущественно в горах. Листья линейные; цветонос несет зонтиковидный завиток из 3-12 и более трубчатых красных, розовых или пурпурных цветков; завязь 3-гнездная, с многими семязачатками в каждом гнезде; тычинки у основания с боковыми выростами, заканчивающимися зубцами, которые соединяются в корону до середины тычинки; столбик короткий с головчатым или слегка 3-лопастным рыльцем. Из трибы эвстефиевых культивируют только один вид из рода эвстефия - эвстефию ярко-красную (Е. coccinea) и один вид из рода урцеолина (Urceolina) - урцеолину матово-красную (U. miniata).
Триба панкратиевых (Pancratieae) включает 6 родов и более 20 видов, распространенных в восточном полушарии: на Канарских островах, в Средиземноморье, Малой Азии и тропической Азии, Австралии и Юго-Западной Африке. Произрастают панкратиевые в самых разнообразных местообитаниях: по берегам водных бассейнов и в долинах рек, в пустынных районах, на песках, на склонах холмов и гор. Цветки у них крупные, душистые, собраны в зонтик, редко одиночные; околоцветник с длинной трубкой; тычинки с придатками у основания, которые часто срастаются в корону.
Самый крупный род трибы - панкратиум (Pancratium). В его состав входят 15 видов, распространенных на Канарских островах, в Средиземноморье, Малой Азии, тропической Азии и Южной Африке. Панкратиум растет в различных условиях: на сухих местах на островах Корсика, Сардиния, Мальта, в Испании и на юге Италии, в пустынных районах на границе Египта и Сирии, на склонах гор влажных и сухих тропиков Индии, на песчаных берегах Средиземного и Черного морей. Листья у панкратиума сидячие, ремневидные. Цветки крупные, белые, душистые, собраны в зонтик по 2-10, иногда одиночные. Околоцветник с длинной трубкой и ланцетными или линейными сегментами. Расширенные и сросшиеся между собой основания тычиночных нитей образуют корону, возвышающуюся над зевом околоцветника. Над короной тычиночные нити свободные, короткие; столбик длинный, нитевидный, с головчатым рыльцем. Плод - коробочка, с угловатыми сдавленными черными семенами. Культивируют несколько видов, в том числе панкратиум морской (P. maritimum) и панкратиум иллирийский (P. illiricum). В странах с жарким и теплым климатом их выращивают в открытом грунте, в условиях умеренного и холодного климата - в оранжереях.
Триба эвхарисовых (Eucharideae) включает 9-10 родов и около 100 видов, распространенных на Американском континенте. Произрастают в разнообразных местообитаниях: в долинах рек, на склонах гор, на высокогорных лугах. Листья сидячие или на черешках; цветки актиноморфные или слегка зигоморфные, с длинной трубкой и развитой короной.
Род эвхарис (Eucharis, табл. 17, 4) включает 10 видов, распространенных в тропической и субтропической областях Центральной и Южной Америки, преимущественно в горных районах, а также по берегам рек в Колумбии и в бассейне реки Амазонки. Листья ланцетные, на длинных черешках. Цветки собраны в рыхлый малоцветковый зонтик, душистые, белые. Околоцветник с длинной трубкой, правильным или слегка зигоморфным отгибом. Разросшиеся основания тычиночных нитей образуют крупную корону, придающую цветку особую привлекательность. Свободные нити тычинок над короной короткие, с качающимися пыльниками. Плод - 3-лопастная коробочка, содержит крупные семена с мясистой кожурой. Эвхарис - одно из распространенных среди любителей-цветоводов тропических луковичных растений. В странах с теплым и жарким климатом его выращивают в открытом грунте, с умеренным и холодным - в оранжереях или в комнатных условиях. В промышленном цветоводстве зарубежных стран его широко используют для выгонки и срезки. Наиболее известны в культуре эвхарис крупноцветковый (Е. grandiflora, табл. 17,4), эвхарис белоснежный (Е. Candida) и эвхарис Фостера (Е. fosteri).

Таблица 17. Амариллисовые: 1 - гиппеаструм полосатый (Hippeastrum vittatum), оранжереи Ботанического института АН СССР в Ленинграде; 2 - подснежник закавказский (Galanthus transcaucasicus), Армения, Кафан, Ваанаван; 3 - гемантус ярко-красный (Haemanthus coccineus), оранжереи Ботанического института АН СССР в Ленинграде; 4 - эвхарис крупноцветковый (Eucharis grandiflora), там же; 5 - гиппеаструм дворцовый (Hippeastrum aulicum), там же; 6 - амариллис красавица (Amaryilis bella-donna), там же; 7 - циртантус Макована (Cyrtanthus makowanii), там же
Самый крупный род трибы эвхарисовых - гименокаллис (Hymenocallis, табл. 16, 2), насчитывающий 56 видов, распространенных в тропической и субтропической Америке: на юго-западе США, в Мексике, Вест-Индии, Центральной и Южной Америке. Растут в различных условиях: в предгорьях и горах, поднимаясь на высоту до 2500 м, в долинах и по берегам рек. Листья сидячие или на коротких черешках. Цветки в многоцветковом зонтике, белые, иногда желтоватые, душистые. Околоцветник с длинной трубкой и узкими линейными или ланцетными сегментами отгиба, актиноморфный или слегка зигоморфный. Тычиночная корона широковоронковидная, крупная, длиной от 2 до 7 см, с короткими тычиночными нитями над верхним краем, несущим несколько зубцов между нитями; пыльники прикреплены основаниями или посередине. Рыльце точечное или головчатое, на нитевидном столбике. Плод - коробочка, с крупными округлыми или угловатыми семенами, по 1-2 или несколько в каждом гнезде. Семена часто прорастают в еще не созревшей коробочке, разрывая ее. Гименокаллисы издавна выращивают в оранжереях и комнатных условиях во всех странах, а в районах с теплым климатом - в открытом грунте. Наиболее часто культивируют гименокаллис красивый (H. speciosa), гименокаллис прибрежный (Н. littoralis, табл. 16, 2), гименокаллис корзинковидный (H. calathina) и др.
Из этой же трибы следует упомянуть род парамонгайя (Paramongaia), описанный сравнительно недавно (в 1949 г.). Род представлен одним видом - это парамонгайя Вебербауэра (P. weberbaueri), произрастающая на склонах гор в Перу. Крупные, правильные, иногда слегка зигоморфные цветки ярко-лимонной окраски, с крупной тычиночной короной очень эффектны и напоминают нарциссы с крупным привенчиком; завязь 3-гнездная, с многими семязачатками в каждом гнезде; локулицидная коробочка несет сочные, довольно крупные, уплощенные светло-коричневые семена, снабженные крылом. Культивируется только в Перу местными жителями.
Для двух последних триб - гиппеастровых и нарциссовых характерно наличие в цветке привенчика, образованного выростами околоцветника.
Триба гиппеастровых (Hippeastreae) включает 8 родов, из них 6 распространены в тропической и субтропической Америке, 1 - в Восточной и Центральной Азии и 1 - в Сирии и Западной Африке. Привенчик у гиппеастровых представлен небольшими выростами в виде чешуек, расположенных в зеве околоцветника. Цветки довольно крупные, слегка или сильно зигоморфные, собраны в зонтик, реже одиночные.
Самый крупный род трибы - гиппеаструм (Hippeastrum, табл. 14, 2, табл. 17, 2, 5) включает около 75 видов, распространенных преимущественно в тропической части Южной Америки, но некоторые виды приурочены к субтропикам (Чили, Аргентина, Уругвай). Произрастают в гористых местах: на склонах гор, плоскогорьях, высокогорных лугах и песках. Листья линейные или продолговато-ланцетные, плотные, с выраженным килем на нижней стороне. Цветки крупные, ярко окрашенные, с нежным запахом, собраны в зонтик по 2-4 (редко 5 и более). Околоцветник с короткой трубкой и широко воронковидным отгибом, в зеве которого у ряда видов имеется привенчик в виде чешуевидных выростов. Тычинки изогнутые, собраны в пучок, над которым возвышается столбик, увенчанный 3-раздельным рыльцем. Гиппеаструмы очень декоративны. Крупные (длиной до 15 см) цветки от белой до ярко-красной окраски украшены полосками, точками и пятнами другого оттенка; многие обладают нежным ароматом. В странах с теплым и жарким климатом их выращивают в открытом грунте, с умеренным и холодным - в оранжереях. Гиппеаструм - одно из излюбленных комнатных растений. Наиболее широко культивируют гиппеаструм полосатый (H. vittatum, табл. 17, 1), гиппеаструм Эванс (Н. evansieae), гиппеаструм дворцовый (H. aulicum, табл. 17, 5), гиппеаструм сетчатый (H. reticulatum) и др. Гиппеаструмы легко гибридизируют, и имеется множество как межвидовых, так и межродовых гибридов.
Род ликорис (Lycoris, табл. 14, 1) представлен 10 видами, распространенными в Азии от Восточных Гималаев до Японии (Центральный Китай, остров Тайвань, Корея и Япония). Произрастает в гористых местах, на щебнистых и глинистых склонах гор и высокогорных лугах. Листья узкие, линейные, сизоватые. Цветки слегка зигоморфные, от нескольких до 12 собраны в зонтик. Околоцветник с короткой трубкой и ланцетными, волнистыми по краю сегментами красной и желтой окраски, образующими в зеве чешуевидные выросты. Тычинки собраны в пучок и окружают нитевидный столбик с маленьким головчатым рыльцем. Плод - локулицидная перепончатая коробочка, содержащая немногочисленные округлые морщинистые черные семена с плотной семенной оболочкой. Почти все виды ликориса культивируются. Имеется много сортов, которые в странах с теплым климатом выращивают в открытом грунте, а с холодным - в оранжереях.
Род шпрекелия (Sprekelia) представлен одним видом. Шпрекелия прекраснейшая (S. formosissima, табл. 14, 5) растет в гористых районах Мексики и Гватемалы. Листья у нее линейные, появляются весной одновременно с цветоносом, несущим 1, редко 2 цветка. Цветки зигоморфные, крупные, длиной 8-10 см, почти двугубые, ярких красных тонов. Трубка околоцветника очень короткая; 3 сегмента прямостоячие, с отогнутой назад верхушкой и чешуевидными выростами в зеве, 3 сегмента отогнуты книзу. Тычинки, собранные пучком, окружают пестик. Плод - коробочка, несет многочисленные уплощенные семена, окруженные узким крылом. На родине шпрекелия широко распространена в садах, в Европе ее культивируют с XVI в. в оранжереях для срезки.
Триба нарциссовых (Narcisseae) включает 3 рода и около 65 видов, распространенных главным образом в Средиземноморской флористической области и в Африке. Привенчик цветка хорошо развит, имеет вид трубки или небольшой зубчатой чаши; цветки одиночные или собраны в зонтик.
Основной род трибы - нарцисс (Narcissus, табл. 14, 3, 4). В него входит около 60 видов, распространенных от Центральной Европы до Северной Африки и от Западной Европы до Западной Азии. В этих пределах нарциссы растут преимущественно во влажных условиях: высокогорные и долинные луга, орошаемые талыми водами, травянистые склоны гор. Пример разнообразия условий обитания дает нарцисс узколистный (N. angustifolius, табл. 14,4) - единственный представитель этого рода, произрастающий в Советском Союзе на восточной границе его ареала, в Закарпатской области. Как правило, он встречается небольшими группами на альпийских лугах, склонах гор и низменностях. Исключение составляет крупный массив чистого нарциссового луга на влажной низменности (200 м над уровнем моря) в районе города Хуст, простирающийся на 15 га и занимающий значительную площадь на хребте Свидовец. Листья у нарцисса линейные. Цветки по 1 или нескольку собраны в зонтик. Околоцветник актиноморфный, с длинной трубкой и ланцетными или эллиптическими сегментами белой и желтой окраски; в зеве околоцветника расположен хорошо развитый привенчик различной формы: от длинно-трубчатой, как у нарцисса ложнонарциссового (N. pseudonarcissus), до очень короткой, как у нарцисса поэтического (N. poeticus). Тычинки и столбик с небольшим рыльцем, не выступают из трубки околоцветника. Плод - коробочка, несет много угловатых черных семян. Нарцисс с его привлекательными, очень душистыми цветками - одно из излюбленных растений цветоводов. Его выращивают в открытом грунте в районах с умеренным и даже холодным климатом (например, в Ленинграде, на Кольском полуострове), где он успешно переносит даже суровые зимы без всякой защиты от морозов. Созданы многочисленные сорта нарциссов. За последние годы получен ряд сортов с розовым крупным привенчиком, придающим цветку особую прелесть.
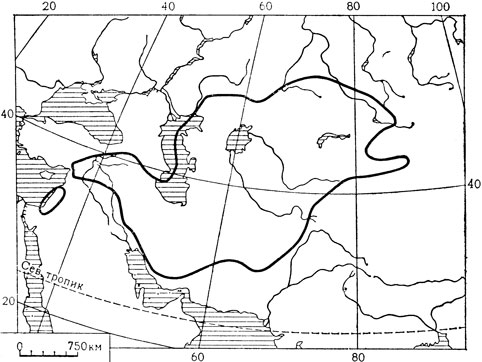
Карта 2. Ареал рода иксиолирион
Подсемейство иксиолирионовых (Ixiolirioideae) представлено единственным родом иксиолирион (Ixiolirion, рис. 57,1-7, табл. 15, 4), в состав которого входят 5 видов, распространенных в Западной, Средней и Центральной Азии. Произрастают преимущественно в предгорьях и горах, на степных склонах и пустынно-степных шлейфах гор на высоте от 800 до 1500 м над уровнем моря, часто спускается ниже; охотно расселяется на пахотных землях, засоряя посевы. Небольшая клубнелуковица иксиолириона к концу вегетации отмирает и заменяется новой. Стебель облиственный, длиной от 15 до 50 см, несет от 3 до 7 желобчатых, у верхушки заостренных листьев, из которых большинство прикреплены к нижней части стебля. Цветки собраны в щитковидном или кистевидном соцветии; околоцветник воронковидный или колокольчатый, сине-фиолетовый, почти до основания шестираздельный; тычинки расположены в 2 круга, наружные короче внутренних; столбик нитевидный, с 3-лопастным рыльцем. Плод - коробочка, раскрывающаяся на верхушке, с многими черными, угловатыми семенами (рис. 57, 7). Культивируется только в открытом грунте, к выгонке не пригоден.
Большинство амариллисовых - декоративные растения. Разнообразие формы и окраски цветков, их размер и аромат придают им особую ценность (табл. 14-17). Все амариллисовые легко гибридизируют. В настоящее время имеются сотни сортов и садовых форм и число их с каждым годом возрастает. Но наиболее интересны межродовые гибриды, которые дают возможность получить растения с совершенно новыми декоративными признаками, несвойственными существующим родам амариллисовых. Межродовая гибридизация открывает широкие перспективы по увеличению ассортимента декоративных амариллисовых.
Семейство формиевые (Phormiaceae) (Р. А. Удалова)
Семейство объединяет 5 родов и около 36 видов, распространенных в тропических и отчасти субтропических областях всех континентов,) за исключением Европы. Самый крупный род - дианелла (Dianella) - насчитывает около 25 видов и широко представлен в странах Юго-Восточной Азии, Австралии и на островах Тихого океана. Два вида дианеллы встречаются только в Новом Свете: дианелла боливийская (D. boliviana) растет в Андах на высоте около 3500 м над уровнем моря в парамосах,, отличающихся достаточно суровым климатом; дианелла сомнительная (D. dubia), так же как и монотипный род экскремис (экскремис сжатый - Excremis coarctata), встречается в высокогорных областях Перу, Колумбии и Венесуэлы. Богаты формиевыми Восточная и Юго-Восточная Австралия и остров Тасмания. Здесь представлено около 9 видов дианеллы и эндемичные роды стипандра (Stypandra) с 3 видами и небольшой род блендфордия (Blandfordia, рис. 58, 6-9). Одно из знаменитых текстильных растений формиум прочный, или новозеландский лен (Phormium tenax), широко распространено в Новой Зеландии и на островах Чатем, Норфолк, Стьюарт и Окленд.
Экологические условия обитания формиевых чрезвычайно разнообразны. Особенно показательна в этом плане дианелла. Дианелла промежуточная (Dianella intermedia), например в Новой Зеландии растет на пемзовых почвах с небольшим слоем гумуса среди кустарников из лептоспермума и папоротника орляка. На островах Новые Гебриды она встречается в подлеске араукариевых лесов, а на Новый Каледонии - в зарослях казуарины. Ее можно встретить также на железнодорожных откосах, на каменистых берегах рек, на песчаных побережьях. Но, вероятно, самым пластичным видом, дающим много форм, является восточно-азиатская дианелла мечевидная (D. ensata). От морских побережий она поднимается в горы до 2700 м. Дианелла мечевидная растет в глубине тропических дождевых лесов и в светлых зарослях кустарников, у потухших кратеров и по берегам рек, на рисовых полях и на влажных скалах. Она является единственным представителем этого семейства, встречающимся в Восточной Африке и на острове Мадагаскар. Виды блендфордии растут большей частью в горных вечнозеленых лесах, иногда на торфянистых болотах.
Характерную, ни с чем не сравнимую черту ландшафта Новой Зеландии создает формиум прочный (рис. 58,2). Особенно пышно новозеландский лен развивается по берегам рек и медленно текущих ручьев, по краям болот и озер. Он растет на прибрежных холмах и дюнах, а поднимаясь в горы, встречается в зарослях вечнозеленых кустарников из лептоспермума, персонии и папоротника орляка. Крупные заросли этого растения встречаются на Северном острове в долине реки Манавату. Отдельными группами новозеландский лен растет и в злаковниках - формациях, образованных различными злаками. Второй вид этого рода - формиум Кука (Phormium cookianum), или горный лен, не так широко распространен, как формиум прочный, хотя встречается на обоих островах Новой Зеландии. Он является характерным компонентом субальпийских буковых лесов из нотофагуса (Nothofagus cliffortioides). Растет он и на побережьях, но, в отличие от формиума прочного, предпочитает сухие скалистые места. Нередко после лесных пожаров формиум Кука становится пионером в возобновлении растительного покрова.
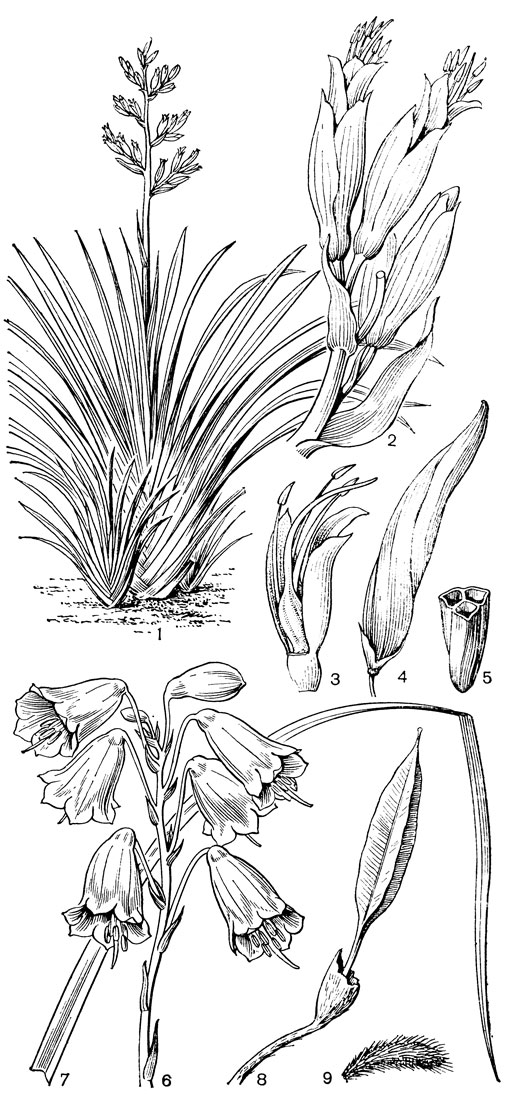
Рис. 58. Формиевые. Формиум прочный, или новозеландский лен (Phormium tenax): 1 - общий вид растения с соцветием; 2 - ветвь соцветия; 3 - цветок (удалены 3 сегмента околоцветника и 3 тычинки). Формиум Кука (P. cookianum): 4-5 - плод. Блендфордия крупноцветковая (Blandfordia grandiflora): 6 - соцветие; 7 - лист; 8 - плод; 9 - семя
Формиевые - многолетние травянистые растения с коротким толстым корневищем. От корневища вверх развиваются побеги, которые образуют достаточно плотные группы - куртины. Стебли формиевых большей частью не превышают 30 см. У дианеллы яванской (Dianella javanica), австралийского вида стипандры крупноцветковой (Stypandra grandiflora) они достигают в высоту 1-2 м. Первоначально их стебли равномерно покрыты листьями, но с возрастом нижние листья опадают и растения приобретают вид драцен, с характерной кроной. Дианелла голубая (Dianella caerulea, рис. 59, 1) по характеру роста напоминает бамбук. Ее стебли быстро растут в высоту, сохраняя почти одинаковый диаметр. Они покрыты длинными коричневыми чешуевидными листьями, и только верхушка несет веер зеленых листьев. После опадения чешуй отчетливо видны утолщенные узлы и междоузлия, равномерное чередование которых придает стеблю вид бамбука. У новогвинейской дианеллы мелкопилъчатой (D. serrulata) стебель настолько мал, что практически становится невидимым.

Рис. 59. Формиевые. Дианелла голубая (Dianella caerulea): 1 - верхняя часть растения; 2 - цветок; 3-4 - тычинка; 5 - плод. Дианелла дубравная (D. nemorosa): 6 - общий вид цветущего растения
Листья формиевых линейные, линейно-ланцетные, большей частью длиной 15-30 см и шириной 0,5-3,5 см. У дианеллы промежуточной и дианеллы тасманской (D. tasmanica) они длиной почти 1,5 м. Темно-зеленые с сизым налетом листья новозеландского льна достигают в длину 2-3 м при ширине 5-12 см. Они, как меч, устремляются кверху. Листья формиевых возникают на очень коротких междоузлиях и располагаются в два ряда в одной плоскости, как веер. Основание листа имеет влагалище, которое подобно черепице плотно налегает одно на другое. Иногда, как, например, у новогвинейского вида дианеллы однолистной (D. mоnophylla), число листьев редуцируется до одного.
Листья новозеландского льна по праву считают одними из самых прочных. Механические волокна имеют толстую оболочку и выдерживают максимум натяжения. Немецкий ученый С. Швенденер вычислил, что сила сопротивления таких клеток выше, чем у каленого железа и кованой стали. Опыт, проведенный в Ботаническом институте АН СССР, показал, что лист новозеландского льна выдерживает массу в 410 кг. Второй вид этого рода - формиум Кука - имеет значительно менее прочные листья, которые не превышают в длину 2,5 м.
Цветение формиевых на родине приурочено к ноябрю - январю, что соответствует в южном полушарии лету. Обоеполые, правильной формы, 3-членные цветки собраны в верхушечное метельчатое, кистевидное (у блендфордии) соцветие. Мощную метелку высотой до 5-6 м ярко-красных цветков развивает новозеландский лен. У прочих формиевых соцветие, как правило, невысокое, хотя у некоторых видов дианеллы оно достигает почти метровой высоты. Число цветков соответственно величине соцветия очень различно: от 4-6 у дианеллы мелкопильчатой до 500 и более у новозеландского льна. Доли околоцветника свободные, почти одинаковые по форме и длине. У блендфордии они срастаются и напоминают цветки колокольчика. Она обладает самыми крупными цветками длиной 4-5 см; самые мелкие (около 5 мм) у дианеллы черной (D. nigra). Преобладающая окраска у дианеллы небесно-голубая; у блендфордии - красная, переходящая на верху околоцветника в оранжевый цвет; у формиума Кука наружные доли околоцветника желтые или красные, а внутренние зеленые. Однако окраска цветков формиумов очень изменчива. У формиума Кука есть разновидности с красными цветками, а у новозеландского льна - с желтыми. Цветки дианеллы остаются полностью открытыми короткое время. Затем у большинства видов сегменты околоцветника смыкаются и плотно склеиваются с завязью и тычинками, так что отделить их друг от друга невозможно. Вскоре они засыхают и с развитием завязи не выдерживают натяжения, разрываются и опадают. Остается лишь небольшая часть околоцветника у основания завязи. У новозеландского льна при выращивании его на Кавказе в Аджарии цветонос развивается около месяца и затем еще в течение двух месяцев (июль - август) постепенно снизу вверх идет раскрытие цветков. Они держатся открытыми в течение 3-5 дней, а потом увядают. Цветки являются протандричными. Обилие пыльцы и нектара привлекает к ним птиц-опылителей. Как сообщает П. Кнут (1904), это колибри, попугаи кака, медососы (семейство Meliphagidae) - птицы из отряда воробьиных, которые с помощью особой щетки на кончике языка высасывают нектар.
Тычинки расположены в два круга и прикреплены к основанию сегментов околоцветника. У блендфордии они прикрепляются в середине околоцветника или несколько выше. Тычиночные нити тонкие, нитевидные, у стипандры в основании или сразу под пыльниками с шерстистым или войлочным опушением. У экскремиса они утолщены в середине, а внизу срастаются в валик. У дианеллы непосредственно под пыльниками тычиночные нити сильно вздуты. Это образование в виде зоба, желтого или оранжевого цвета, особенно развито у дианеллы голубой и дианеллы гладкой (D. laevis). Некоторые ботаники описывали их как желёзки. В целом их строение напоминает желёзки, однако проверенных данных о функции этого образования нет. У дианеллы увенчанной (D. coronata) вместо зобовидного образования имеются сосочковидные выросты. Пыльники линейные или линейно-ланцетные, раскрываются верхушечными щелями. У стипандры, блендфордии и дианеллы яванской они после цветения скручиваются в виде улитки. Пыльцевые зерна у блендфордии однобороздные, а у формиума и дианеллы с трехлучевой бороздой.
Гинецей синкарпный и состоит из 3 плодолистиков; завязь верхняя, 3-гнездная, с многими семязачатками; столбик тонкий, нитевидный, с небольшим головчатым рыльцем; у экскремиса столбик довольно толстый, с маленькими долями рыльца. Плоды формиевых удлиненные локулицидные коробочки с многими семенами. У формиума прочного коробочка достигает в длину 20 см. У формиума Кука три ее грани перекручены. У дианеллы плоды шаровидные или удлиненно-шаровидные голубые ягоды. Семена формиевых черные, блестящие, а у блендфордии покрыты волосовидными сосочками.
Из двух видов формиума мировую известность завоевал новозеландский лен. С давних времен коренные жители Новой Зеландии - маори использовали его в качестве текстильного растения. Отсюда происходит родовое название, означающее "циновка", "корзиночка". Для того чтобы получить волокно, маори соскребали острым краем раковины наиболее жесткую часть листьев, затем листья вымачивали в проточной воде в течение 4-5 дней, чтобы удалить слизь, и, наконец, мяли, отбеливали на солнце и высушивали. В народной медицине использовали также сок корней и основания листьев. Молодые листья новозеландского льна служили для изготовления традиционных украшений, которые юноши дарили своим избранницам. Маори активно не разводили новозеландский лен, а использовали его природные запасы.
Экспорт волокна начался в 1828 г., однако в Англии это растение стало известно еще раньше от французского мореплавателя и исследователя Л. К. Фрейсинета. Его выращивали как горшечное растение при дворе короля Георга II. Благодаря способности новозеландского льна выдерживать понижение температуры до -12...-15 °С, его стали разводить во многих странах мира: первоначально в Англии, затем в Мексике, Бразилии, Чили, Аргентине. В Японии до второй мировой войны ежегодно производилось 10-12 тыс. т волокна. В СССР новозеландский лен культивируют с конца прошлого столетия на Черноморском побережье Кавказа, главным образом в ботанических садах.
Современный процесс обработки листьев новозеландского льна напоминает древний способ маори, с той лишь разницей, что в настоящее время почти все операции выполняют машины. Из новозеландского льна ткут дорожки, маты, обивочный материал, причем его часто комбинируют с сизалем (агава сизалевая). Из новозеландского льна делают традиционные юбки "пиу-пиу". Из свежего зеленого листа вырезают узкие полоски, которые затем подвергают специальной обработке: скоблят, вымачивают, красят. В результате из каждой полоски листа получается тонкая длинная трубочка белого цвета с черными поперечными "поясками". На одну юбку необходимо приблизительно 240 таких трубочек.
Среди формиевых немало декоративных растений. Из австралийских видов наиболее популярны дианелла голубая - невысокий кустарник с нежными голубыми цветками и плодами, блендфордии - с крупными красными или желто-красными цветками. Во многих ботанических садах и оранжереях выращивают культурные формы новозеландского льна с красно-пурпурными или пестрыми листьями.
Семейство агавовые (Agavaceae) (Р. А. Удалова)
Семейство объединяет 10 родов и около 450 видов, распространенных в тропических и субтропических засушливых областях Северной и Центральной Америки, на севере Южной Америки, на островах бассейна Карибского моря, а также в Восточной Азии. Северная граница ареала агавовых доходит до 40-41° с. ш., где в штате Колорадо в Скалистых горах на высоте 2000-2500 м над уровнем моря растет стелющаяся среди камней юкка сизая (Yucca glauca). Южная граница ареала проходит через Венесуэлу и Колумбию. В Венесуэле несколько видов агав распространены главным образом в прибрежной зоне и на близлежащих островах (Кюрасао, Аруба). В Колумбии в горах бассейна реки Каука встречается агава Валлиза (Agave wallisii) и широко известная в культуре фуркрея Селло (Furcraea selloa, табл. 18, 5). Очень разнообразны агавы на Багамских островах и Ямайке, однако основное видовое богатство сосредоточено в Мексике и на юге США. Здесь наряду с широко известными растениями, как юкка (Yucca) и агава (Agave, рис. 60, табл. 18), представлены редкие, малоизвестные роды псевдобравоя (Pseudobravoa), прохниантес (Prochnyanthes) и геспералоэ (Неsperaloe). Только один род хоста (Hosta, табл. 18, 4) приурочен к умеренно теплым областям Восточной Азии (Япония, Китай, полуостров Корея, СССР - Приморский край и остров Сахалин).
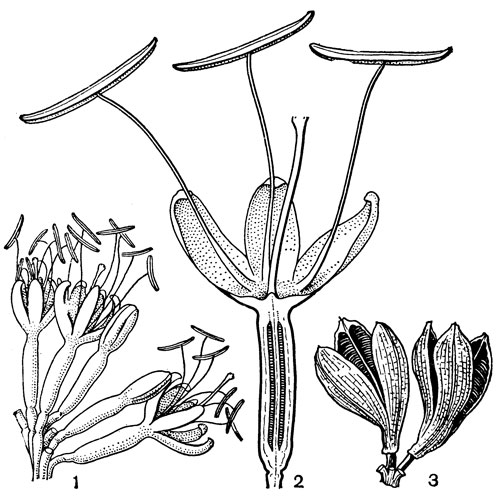
Рис. 60. Агава восковая, подвид почти восковая (Agave cerulata subsp. subcerulata): 1 - пучок цветков боковой ветви соцветия; 2 - продольный разрез цветка; 3 - плоды

Таблица 18. Агавовые и дориантовые: 1 - юкка коротколистная (Yucca brevifolia), пустыня Мохаве, Калифорния, США; 2 - юкка Трекуля (Y. treculeana), Сухумский ботанический сад; на заднем плане - юкка алоэлистная (Y. aloifolia); 3 - агава американская (Agave ameriсаnа) в цвету, Алжир; 4 - хоста подорожниковая (Hosta plantaginea), Крым, Никитский ботанический сад; 5 - фуркрея Селло (Furcraea selloa), оранжерея Ботанического института в Ленинграде; 6 - юкка повислая, форма цельная (Yucca flaccida f. integra), Батумский ботанический сад; 7 - соцветие юкки повислой, форма цельная; 8 - дориантес высокий (Doryanthes excelsa), Австралия
Агавовые - в основном корневищные древовидные растения, для которых характерно вторичное утолщение стебля однодольного типа. Так, у юкки коротколистной (Yucca brevifolia, табл. 18,2) ствол бывает высотой до 12 ми диаметром 60 см. У агавы он сильно укорочен и обычно не превышает 30-50 см, но отдельные виды, как, например, агава Карвинского (Agave karwinskii), образуют ствол высотой до 4 м. Роды хоста, полиантес (Polianthes), псевдобравоя и прохниантес представлены многолетними травянистыми формами (рис. 62, 7-11). Сосуды находятся только в корнях (с простыми перфорациями у агавы и лестничными у хосты). Листья агавовых суккулентные или кожистые, большей частью крупные и собраны в прикорневой розетке или расположены пучком на вершине ствола. Цветки большей частью белые, кремовые или зеленоватые. Они двудомные, актиноморфные или слабозигоморфные (у полиантеса). У большинства представителей они собраны в крупное верхушечное соцветие - кисть, колос или метелку. У агавы и фуркреи (Furcraea) соцветие достигает в высоту 8-10 м. Сегментов околоцветника 6, расположенных в два круга, одинаковых по форме и почти не отличающихся по окраске. Обычно они срастаются (кроме юкки и фуркреи) в более или менее длинную трубку. Шесть длинных тычинок прикрепляются к основанию лепестков или трубки венчика. Пыльники интрорзные, открываются продольными щелями. Оболочка пыльцевых зерен однобороздная или реже (полиантес) с двумя дистальными бороздами,, с толстой сэкзиной. Столбик нитевидный, а у фуркреи он трехгранно утолщен; рыльце 3-лопастное. Завязь 3-гнездная, у большинства родов нижняя, но у юкки, геспералоэ и хосты она верхняя. Семязачатки анатропные, многочисленные, расположенные в два ряда друг над другом. Плод - локулицидная коробочка, а у юкки ягодовидной (Yucca baccata), юкки алоэлистной (Y. aloifolia, табл. 18, 2) и некоторых других ягодообразный. Семена плоские, черные.
Семейство агавовых делят на 3 трибы: хостовые (Hosteae), юкковые (Yucceae) и агавовые (Agaveae).
Триба хостовых включает один род хоста, насчитывающий около 40 видов. По внешнему виду хоста очень заметно отличается от всех других членов семейства. Это - многолетние травянистые растения с поверхностным компактным или слабо разветвленным корневищем. Листья на черешках от узколанцетных до широкояйцевидных расположены пучками в основании укороченного стебля. Белые или нежно-лиловые цветки собраны в невысокую кисть. Они растут по скалистым и каменистым берегам рек, нередко у самой воды, по лесным опушкам, у ручьев, а иногда на заболоченных участках.
Хоста вздутая (Hosta ventricosa) и хоста ланцетовидная (Н. lancifolia) встречаются в СССР на Курильских островах и Сахалине. В современной ботанической литературе хосту относят к семейству лилейных. Однако с агавовыми ее сближает одинаковый набор хромосом (тип Yucca-Agave), целый ряд эмбриологических признаков и данные серологии (исследования В. С. Чупова и Н. Г. Кутявиной, 1978). Хосту издавна выращивают в Японии как декоративное растение, откуда в конце XVIII в. она попадает в европейские сады. Наиболее известны в культуре хоста курчавая (Н. crispnla), хоста подорожниковая (Н. plantaginea, табл. 18, 4), хоста Фортъюна (Н. fortunei) и ее культурные разновидности с полосатыми или пятнистыми листьями. В китайской медицине используются также корни хосты вздутой как средство от зубной боли.
В трибе юкковых (Yucceae) 2 рода - геспералоэ и юкка. Последний рассматривается здесь в широком объеме и включает в себя роды геспероюкка (Hesperoyncca), клистоюкка (Сlistoyucca), самуэла (Samnela) и саркоюкка (Sarсоуnсса). У представителей этой трибы, в отличие от большинства других агавовых, верхняя завязь, а сегменты околоцветника свободные, не срастающиеся в трубку. Геспералоэ долгое время оставалось загадкой для ботаников. Эти почти бесстебельные растения с длинными линейными кожистыми листьями, подобно злакам, образуют плотные дерновины. Их листья с отделяющимися по краю волокнами напоминают листья юкки, а колокольчатовидные цветки - алоэ (отсюда и название рода). Вот почему во второй половине XIX в. одни ботаники относили эти растения к роду юкка, а другие - к алоэ. Спор разрешил американский ботаник Г. Энгельманн, который в 1871 г. установил для них новый род геспералоэ, указав, что его листья, пыльца и семена такие же, как у юкки, а околоцветник и гинецей как у алоэ, в то же время тычинки, как у агавы.
Цветки геспералоэ эфемерны, опыляют их птицы. В течение дня цветки меняют вертикальное положение на горизонтальное, а затем цветок повисает. К концу дня он теряет свою красоту. Обильный нектар, выделяемый тремя крупными желёзками, склеивает в единую массу столбик, лепестки и тычинки.
В роде геспералоэ 3 вида, из них геспералоэ мелкоцветковое (Hesperaloe parviflora, рис. 62) имеет редкую в семействе агавовых розово-красную окраску цветка.
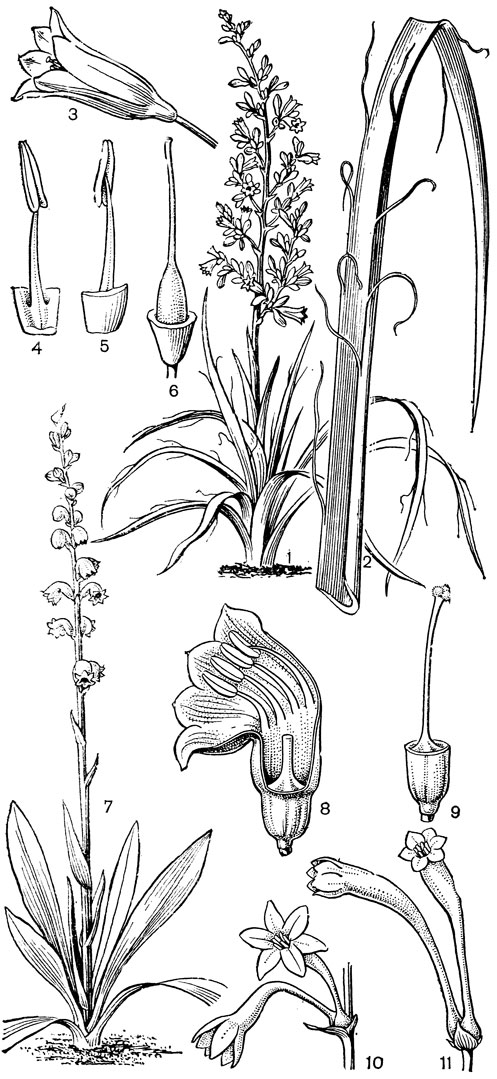
Рис. 62. Агавовые. Геспералоэ мелкоцветковое (Hesperaloe parviflora): 1 - общий вид; 2 - лист; 3 - цветок; 4, 5 - тычинка; 6 - гинецей. Прохниантес Балла (Prochnyanthes bulliana): 7 - общий вид; 8 - цветок с удаленным столбиком и частью околоцветника; 9 - гинецей. Полиантес дурангийский (Polianthes durangensis): 10 - цветки. Полиантес болотный (P. palustris): 11 - цветки
Юкка, в отличие от геспералоэ, большей частью образует древовидный ствол высотой нередко до 6 м, даже 12 м. Такие великаны, как юкка карнерозанская (Yucca carnerosana), ранее относившаяся к роду самуэла, юкка сильная (Y. valida), и некоторые другие, возвышаются в пустынях среди невысоких колючих кустарников. Их стволы укутаны старыми повисшими вниз засохшими листьями, а вершину растения венчает мощное соцветие из белых цветков. Некоторые виды, например юкка нитчатая (Y. filamentosa), юкка повислая (Y. flaccida, табл. 18, 6, 7), юкка равнинная (Y. саmpestris), не имеют ствола. Они обильно ветвятся, образуя плотные дерновины.
Известно около 50 видов юкки. Ареал рода протянулся от Атлантического побережья Флориды и островов Карибского моря, через южные штаты США и Мексику до Калифорнии. Юкки растут в засушливых местах, предпочитая сухие открытые пространства с песчаными, каменистыми или известковыми почвами. Многие виды встречаются в креозотовых пустынях из креозотового куста (Larrea tridentata) в ассоциации с дазилирионом (Dasylirion sp.), окотилло (Fouquieria splendens), опунцией (Opuntia sp.) и другими колючими растениями. В сосново-можжевеловых лесах на высоте 2000-2450 м растут юкка преузкая (Yucca angustissima), юкка ягодовидная (Y. baccata), занимающая очень широкий ареал на юго-западе США, юкка Стэндли (Y. standley) - одно из самых маленьких растений этого рода, стебель которой в высоту не превышает 20 см. Один из красивейших видов - юкка Трекуля (Y. treculeana, табл. 18, 2) на большей части своего ареала растет в зарослях колючих жестколистных кустарников (чаппараль). Многие виды юкки являются также непременным элементом кактусово-акациевых саванн. По берегам пересыхающих ручьев, у подножия гор или по краям крутых каменистых каньонов - везде можно встретить одиноко растущие юкки, которые, как правило, не образуют плотных насаждений. Исключением в этом плане является юкка коротколистная, известная под названием "дерево Джошуа" ("Yoshuatree"). Поражает необычный внешний вид этого растения. Огромное дерево, высотой до 9 м, с мощным, обильно и причудливо ветвящимся стволом, как древний исполин, возвышается в пустыне среди бесплодных песков и невысоких колючих кустарников. Эта юкка одна из немногих, развивающих огромную крону. Самые старые экземпляры, по некоторым данным, достигали возраста 800 лет. Ареал юкки коротколистной проходит через пустыню Мохаве в штатах Аризона и Калифорния. В этих местах она иногда образует своеобразные разреженные леса, которые тянутся на несколько километров. Для сохранения этого удивительного растения был создан в окрестностях Риверсайда (штат Калифорния) заповедник "Национальный памятник дерева Джошуа" ("Yoshua Tree National Monument").
У видов юкки наблюдается очень своеобразный и интересный механизм перекрестного опыления. Цветки открываются ночью и опыляются маленькой белой бабочкой из рода пронуба (Pronuba yuccasella). С наступлением тем-ноты, когда белые цветки выделяют приятный аромат, самка пронубы приступает к сбору пыльцы. Она клейкая и напоминает замазку. Пронуба скатывает пыльцу в комочек и благодаря зубоподобному цепкому и колючему придатку переносит ее к другому цветку. Здесь бабочка вдавливает пыльцу в рыльце, а затем откладывает в завязь цветка свои личинки. В результате семязачатки оплодотворяются, а личинки питаются развивающимися семенами. Их так много в плоде, что хватает и для личинок бабочки, и для воспроизведения растения. С наступлением дня бабочки обоих полов (их скапливается достаточно большое количество) остаются в цветке. Они сидят на тычиночных нитях и, вероятно, в это время питаются пыльцой.
Впервые об опылении юкки стало известно в 1872 г. из короткого сообщения известного американского ботаника Г. Энгельманна. Несколькими годами позже, в 1878 г., американский энтомолог Ч. Рили подробно изучил и описал удивительное поведение этой бабочки. Другой вид пронубы - P. synthetica - вступает в контакт только с юккой коротколистной. Единственным самоопыляющимся видом является юкка алоэлистная.
Юкка - прекрасное декоративное растение. Ее нередко выращивают в садах, парках и оранжереях многих стран мира. Некоторые виды юкки стали известны науке благодаря коллекциям ботанических садов. Так, в 1859 г. русским ботаником Э. А. Регелем была описана юкка слоновая (Y. elephantipes), выращенная в оранжереях Санкт-Петербургского ботанического сада. На территории нашей страны юкку культивируют на Черноморском побережье Кавказа и Крыма с 1816 г. Особенно известны юкка нитчатая, юкка славная (Y. gloriosa), юкка алоэлистная. Два последних вида морозоустойчивые и выдерживают температуру до -15 °С. В культуре юкки, за исключением юкки алоэлистной, из-за отсутствия опылителей не плодоносят. Из жестких листьев юкки получают прочное техническое волокно. Из цветков юкки слоновой местные жители приготавливают салат. Многие виды юкки используют в качестве живых изгородей.
Триба агавовые (Agaveae) - самая крупная в семействе. В нее входят 7 родов и около 360 видов, из них более 300 относятся к роду агава (Agave). В переводе с греческого "агава" означает "статная, видная". Первые упоминания европейцев о ней относятся к середине XVI в. В Европу агавы попали во второй половине XVI в., первоначально в Испанию, а затем в Италию и Францию. Итальянский ботаник А. Чезальпино видел цветущую агаву в городе Пизе в 1583 г., а И. Камерариус - во Флоренции в 1586 г. Агава быстро завоевала признание как интересное декоративное растение. Благодаря сравнительной легкости вегетативного размножения ее разводят во многих странах Средиземноморья.
Как уже говорилось вначале, агава, за несколькими исключениями, не образует ствола. Ее листья крепкие, большей частью сочные, собраны в плотную розетку. У большинства видов ее диаметр достигает 3 м, а у агавы Франдзозини (A. franzosini) - 4,5 м. Как исключение можно назвать агаву клобучковую (A. cucullata) с листьями длиной до 10 см и агаву карликовую (A. pumila), листовая розетка которой диаметром не более 3-4 см. В розетке обычно от 20 до 50 листьев, а у агавы парноцветковой (A. geminiflora) их до 200. Они такие тонкие и узкие, что ее называют "тростниковолистной". По краям листа агавы имеются крепкие шипы, особенно внушительные у агавы неустрашимой (A. ferox), а у агавы нитеносной (A. filifera) и некоторых других отделяются тонкие волокна. Конец листа заканчивается колючкой. Паренхимная ткань листа узкоспециализирована как водозапасающая, а эпидерма покрыта голубоватым восковым налетом, препятствующим чрезмерному испарению влаги. В течение периода роста агавы развивают от 3 до 7 листьев, каждый из которых живет несколько лет.
Агавы, за небольшим исключением,- монокарпики, цветущие один раз в жизни. Одни виды цветут на родине в возрасте 5-15 лет, другие - в 50 и даже 100 лет. На Черноморском побережье Кавказа и Крыма агава американская (A. americana, табл. 18, 3) цветет на 10-12-й год. В оранжереях ботанического сада Ботанического института АН СССР в Ленинграде наблюдалось цветение агавы многоколючковой (A. polyacantha) в возрасте 25 лет, а агавы Вера-Крус (A. vera-cruz) - в 35 лет.
Цветение агав - зрелище очень впечатляющее, запоминающееся надолго. У большинства видов цветонос поднимается на высоту до 3 м. У агавы американской он достигает 8 м, а у агавы Салъма (A. salmiana) и агавы неустрашимой - даже 10 м! Цветонос развивается 2-3 месяца, и затем еще в течение нескольких месяцев постепенно раскрываются цветки. Они окрашены в зеленоватый или желтоватый цвет и располагаются пучками по 2-4 - 6-8 на боковых ветвях соцветия. У агавы узколистной (A. angustifolia) в соцветии насчитывают более 2000 цветков. Длинные тычинки далеко выступают из околоцветника. Их пыльники крупные, подвижные, созревают раньше завязи (протандрия). По наблюдениям Е. Кокрума и Б. Хейворда (1962), цветки агавы Шотта (A. schottii) и агавы узколистной опыляются летучими мышами (хироптерофилия) из рода лептониктерис (Leptonycteris nivalis). Раскрытие цветков в ночное время, обилие сладкого нектара и пыльцы со специфическим запахом привлекают ночных опылителей. Наличие высокого прочного цветоноса позволяет летучим мышам "приземлиться" и, медленно спускаясь вниз по соцветию, слизывать пыльцу и нектар. В дневное время цветки посещают пчелы, не исключена так-же возможность и ветроопыления. В Эквадоре наблюдали опыление агав гигантскими колибри из рода натагония (Patagonia gigas). На Ямайке, по данным Л. Галдинга, агава отпрысконосная (A. sobolifera) опыляется райскими колибри (Topaza pella). По данным А. Бергера (1915), в Африке одичавшие агавы активно посещаются птицами-нектарницами (Nectarinia famosa). Случайными опылителями бывают также мыши и крысы. Плоды агав долгое время остаются на засохшем цветоносе. С каждым порывом ветра из растрескавшихся коробочек часть семян высыпается и разносится на большие расстояния. Однако не у всех видов развивающиеся плоды полноценны. Нередко семена недоразвиты и в этом случае органом размножения являются бульбиллы - небольшие дочерние растеньица, развивающиеся на цветоносе из адвентивных (придаточных) почек. Бульбиллы имеют корни и, отрываясь ветром от соцветия, быстро укореняются. Их жизнеспособность сохраняется в течение нескольких лет.
Агавы - характерные представители флоры юга США, Центральной Америки и Вест-Индии. В Южной Америке они обитают в прибрежных областях Венесуэлы и близлежащих островов, а в Колумбии - в бассейнах рек Магдалены и Кауки. Агавы встречаются и в Перу, однако, по всей вероятности, в данном случае речь идет об их разведении, как и в странах Средиземноморья и Индии. Для некоторых видов, особенно издавна культивируемых, таких как агава сизалевая (A. sisalana) и агава фуркреевидная (A. fourcroides), установить родину невозможно. Агавы растут в различных аридных растительных формациях. В Мексике - главном центре их развития - они встречаются в зарослях колючих кустарников из ипомеи древовидной (Ipomoea arborescens), бурзеры (Burserasp.), представителей семейства бобовых. В горных дубово-сосновых и сосновых лесах они часто поднимаются на высоту 2000-3600 м над уровнем моря. На побережье Калифорнийского залива многие виды агавы доминируют в саванновых формациях из фукьерий колончатой (Fouquieria columnaris), фукьерии блестящей (F. splendens), пахицереуса (Pachycereus sp.), юкки и видов рода опунция (Opuntia). Большие заросли агавы восковой (A. cerulata) тянутся на несколько километров в центральной части Калифорнийского полуострова.
Близок к агаве род манфреда (Manfreda, рис. 61). Около 18 видов этого рода представлены многолетними травянистыми растениями с сильно укороченным и клубневидно утолщенным стеблем. Листья у некоторых видов пестрые и, в отличие от агав, не имеют верхушечной колючки. У манфреды вирджинской (М. virginiса) - самого северного вида, встречающегося в штате Мэриленд, листья ежегодно отмирают. Цветки манфреды располагаются в пазухе прицветника по одному, а не пучками, как у агавы.
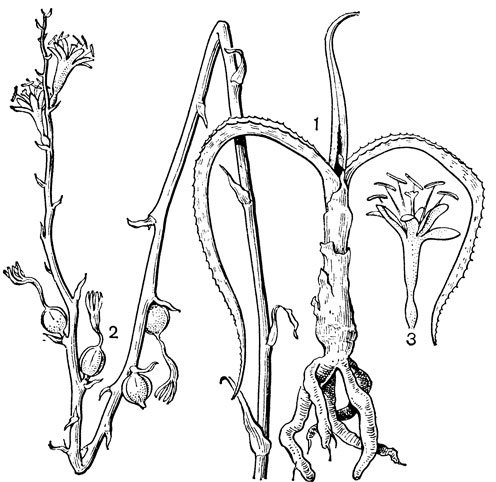
Рис. 61. Манфреда многопятнистая (Manfreda maculosa): 1 - общий вид; 2 - соцветие; 3 - цветок
Резко отличаются по внешнему виду от остальных представителей агавовых полиантес (Polianthes), близкий к нему род прохниантес (Prochnyanthes) с 2-3 видами из пустынь Мексики и монотипный род псевдобравоя (Pseudobravoa). Все эти растения - небольшие многолетние травы, стебель которых сильно укорочен и луковицеобразно утолщен. Листья собраны в розетку, большей частью они узкие и тонкие. Наибольший интерес представляет полиантес (в переводе с греческого - беловатый цветок) и особенно полиантес клубневый (Р. tuberosa), известный в садоводстве под названием тубероза. Белые душистые цветки туберозы содержат эфирные масла, используемые в парфюмерной промышленности. Тубероза известна и как прекрасное декоративное растение, цветущее поздним летом и осенью.
Агавовые имеют большое практическое значение, прежде всего как волокнистые растения. Мировую известность завоевала агава сизалевая, с давних времен культивируемая во многих странах. Ее прочное волокно, называемое "сизаль" или "пита", идет на изготовление веревок, канатов, рыболовных сетей и лассо. Агаву сизалевую разводят главным образом на Багамских островах, в Вест-Индии, Бразилии и Танзании (1/3 мирового производства). Плантации сизаля используют в течение 12-15 лет, ежегодно обрезая по 7-9 листьев с каждого растения. Второй важной промышленной культурой является агава кантала (A. cantala), которую разводят преимущественно на Яве, Филиппинских островах и в восточной части Индии. Агава фуркреевидная дает волокно "хенекен" ("энекен") или "юкатанский сизаль". На плантациях энекена каждое растение используется в течение 18 лет. Главный центр ее производства - Мексика, Куба, полуостров Юкатан. Белое волокно энекена идет на изготовление бумаги, веревок, тарной ткани. Известны также агава складчатая (A. falcata), из которой получают волокно "эспадин", агава жесткая (A. rigida), дающая волокно "кенжут". На острове Св. Елены и острове Маврикия широко культивируется фуркрея вонючая (F. foetida), или "маврикийская конопля", а на Кубе - фуркрея шестилепестная (F. hexapetala), или "кубинская конопля". Трудно переоценить значение агав в жизни индейцев. Они используют в пищу печеную сердцевину ствола и основания листьев, из них же изготовляют муку. Индейцы едят цветки агав в вареном виде или высушивают их и добавляют в маисовые лепешки. Некоторые агавы и манфреды содержат в листьях большое количество сапогенинов, которые применяют для мыловарения. Из агавы темно-зеленой (A. atrovirens) и агавы Сальма приготовляют алкогольный напиток пульке. Используют агавы и в качестве живых изгородей и в народной медицине как средство от укусов насекомых и змей. Агава, фуркрея, бешорнерия (Beschorneria) снискали себе славу как прекрасные декоративные растения. Их широко разводят в садах, парках и оранжереях во многих странах мира. Особенно известны агава неустрашимая, агава ярко-красная (A. coccinea), агава американская и ее культурные разновидности.
Семейство дориантовые (Doryanthaceae) (Т. В. Вельгорская)
Монотипное семейство дориантовые включает род дориантес (Doryanthes), который состоит из 2-3 видов, распространенных в прибрежных районах штатов Новый Южный Уэльс и Квинсленд на востоке Австралии.
Дориантовые - крупные многолетние травы с утолщенным подземным стеблем, коротким вертикальным корневищем и толстыми контрактильными (втягивающими) корнями, которые развивают еще и боковые корни (рис. 63,4). В течение сухого сезона корни дориантеса сокращаются и слегка втягивают растение в грунт; у старых особей стебли обычно сидят глубоко в почве. Листья большие, достигающие в длину 1,5-2 м, собраны в большую приземную розетку (иногда содержащую до 100 листьев), интенсивно-зеленые, с ребристой поверхностью, гладкими краями и суживающиеся примерно с середины длины листа к концу. Все листья, включая и кроющие листья соцветия, начинают свое развитие с появления белого, с овальной губчатой верхушкой твердого придатка молодого свернутого в трубку листа. Этот придаток, лишенный хлорофилла, имеет многочисленные устьица. По мере того как лист постепенно вытягивается, развертывается и в нем появляется хлорофилл, придаток высыхает (рис. 63, 3) и становится коричневым, слегка засыхает также и верхушка листа. Затем засохшие части иногда опадают, оставляя неровный зубчатый край. Австралийский ученый Айвар Ньюман (1929) предполагает, что этот белый придаток, или "наконечник", осуществляет респираторную функцию еще свернутого листа, но экспериментальные доказательства этого пока отсутствуют. Устьица у листьев дориантеса парацитные.
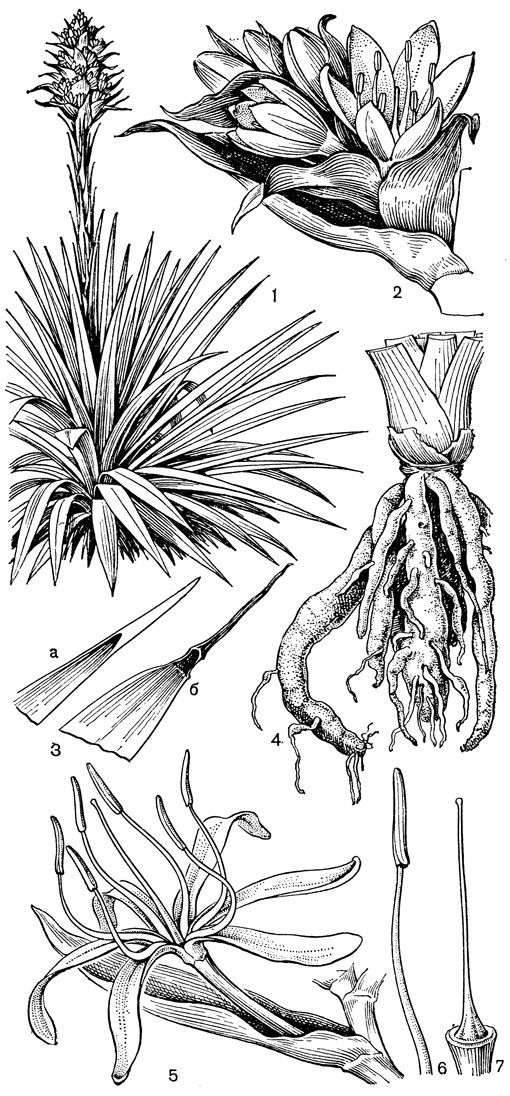
Рис. 63. Дориантовые. Дориантес Пальмера (Doryanthes palmeri): 1 - общий вид надземной части; 2 - кисть соцветия; 3 - придаток листа (а - ранняя стадия, б - взрослый лист). Дориантес высокий (D. excelsa): 4 - подземные органы; 5 - цветок; 6 - тычинка; 7 - столбик
Цветки обоеполые, актиноморфные, собранные в большие головчатые (у дориантеса высокого - D. excelsa) или тирсоидные (у дориантеса Палъмера - D. palmeri, рис. 63, 1) соцветия, состоящие из пазушных кистей с укороченной главной осью. Впечатляют размеры как соцветий, так и цветоносов. Например, у наиболее крупных экземпляров дориантеса высокого соцветия, достигающие в диаметре 60 см, подняты мощным цветоносом на высоту до 5,5 м (табл. 18,8). Молодое соцветие полностью окружено большими кроющими листьями, которые по мере его развития краснеют и расходятся в стороны. Каждая кисть соцветия расположена в пазухе кроющего листа, а каждый цветок в кисти, в свою очередь, имеет свой прицветник. В соцветиях бывает до 150 цветков, и диапазон возрастов их весьма различен: когда нижние цветки уже завяли и начинают образовываться плоды, верхние только раскрываются. Цветки темно-красные, пурпурные снаружи и розовые или розовато-белые внутри, очень крупные. Околоцветник венчиковидный. Сегментов околоцветника 6, расположенных в 2 круга, сросшихся почти до половины своей длины в трубку и у дориантеса высокого слегка отгибающихся. Тычинок 6, расположены в 2 круга. У дориантеса высокого длинные тонкие тычиночные нити до половины своей длины как бы опираются на сегменты околоцветника, а затем, изогнувшись почти под прямым углом, становятся вертикально, вынося вверх свои очень длинные желтовато-зеленые пыльники (рис. 63, 5,6). У дориантеса Пальмера пыльники менее крупные; сначала они желтые, затем приобретают фиолетовую окраску. Пыльники интрорзные, прикрепленные к нитям основаниями и раскрывающиеся продольной щелью. Пыльцевые зерна однобороздные. Гинецей синкарпный, из 3 плодолистиков; завязь нижняя, с многочисленными (от 120 до 150) анатропными семязачатками; длинный суживающийся к верхушке столбик заканчивается маленьким треугольным рыльцем. В месте срастания обоих кругов сегментов околоцветника основание их расширяется, слегка возвышаясь над завязью, и образует нектарную чашу. Плод дориантеса - крупная деревянистая локулицидная коробочка. Семена светло-коричневые, довольно крупные, эллипсовидной формы, снабженные тонким широким крылом. Семя с крупным зародышем, окруженным эндоспермом.
Интересна история систематического положения рода дориантес. Обычно его помещали в трибу агавовых семейства амариллисовых. В системе Дж. Хатчинсона этот род включен в отдельное семейство агавовых. В 1969 г. немецкий ботаник Г. Хубер выделил дориантес, основываясь, в частности, на анатомии семени, в самостоятельное семейство дориантовые, которое впоследствии было принято Р. Дальгреном (1975, 1980) и А. Л. Тахтаджяном (1980). Действительно, от всех агавовых дориантес отличается очень своеобразной морфологией листьев, парацитными устьицами, пыльниками, прикрепленными к нитям основаниями, симультанным типом образования микроспор и некоторыми другими эмбриологическими особенностями, а также кариотипом. На основании сравнительных серологических исследований В. С. Чупов и Н. Г. Кутявина (1981) пришли к выводу, что род дориантес очень обособлен в пределах порядка лилейных.
Дориантес - обычное растение светлых эвкалиптовых лесов Восточной Австралии. Он растет на песчаных, хорошо дренированных почвах по склонам оврагов, чаще всего обращенных к юго-востоку, иногда встречается и на северных склонах, но, как правило, только в тех случаях, если почва там глинистая. Дориантес - светолюбивое растение, и выросшие в тени особи заметно уступают по размерам розеток и листьев своим более удачливым соседям - обитателям открытых мест. Растут дориантесы обычно большими группами, где границы каждой популяции часто очень отчетливы, так как наряду с половым для дориантеса свойственно и вегетативное размножение.
Цветет дориантес австралийской весной - в октябре - ноябре, и цветение его крупных соцветий продолжается от 2 до 3 месяцев. Наличие в цветке нектара и обильной пыльцы, яркие тона околоцветника указывают на то, что дориантес - биотически опыляемое растение. Есть основания полагать, что у себя на родине он опыляется птицами - медососами и нектарницами (в пользу этого предположения говорит и еще один факт, о котором читатель узнает чуть позже). Не исключено также, что опылителями этого растения могут быть и насекомые. К сожалению, сведений об экологии этого эндемичного рода сравнительно мало.
Плоды дориантеса созревают в январе - феврале. Высвободившиеся из коробочки крылатые семена распространяются анемохорно. Несмотря на то что семена готовы к прорастанию, дориантесы прорастают медленно, и проходит много месяцев, прежде чем растение разовьется, а от начала прорастания семени до цветения более 10 лет. После цветения и созревания плодов растение погибает, другими словами, дориантес - типичный монокарпик.
Дориантес был привезен в Европу в самом начале прошлого века и своим необычайным видом, великолепием красок и форм снискал восхищение и у ботаников, и у садоводов. Его называли одним из чудес растительного царства еще и потому, что впервые привезенный из Австралии стебель дориантеса, не имея подземных органов, вдруг зацвел. Начиная с 1870 г. и по настоящее время дориантес, особенно дориантес Пальмера, широко культивируется на юге Соединенных Штатов Америки и в Мексике. Растения очень декоративны и их используют в пейзажных и регулярных парках; хорошо смотрится дориантес и в маленьких внутренних двориках - патио. В Америке дориантес привлек внимание не только садоводов, но и маленьких колибри, которые в обмен на обильный нектар взяли на себя роль опылителей этого замечательного растения.
Семейство асфоделовые (Asphodelaceae) (Т. В. Егорова)
Семейство асфоделовых, состоящее из 42 родов и почти 1500 видов, распространено преимущественно в Старом Свете, главным образом в Южной и тропической Африке и Австралии, а также Макаронезии, на Мадагаскаре и Маскаренских островах, в Средиземноморье, Европе (до юга Швеции и Ирландии), Западной, Юго-Западной и Средней Азии, на северо-западе Центральной Азии, Алтае, в Гималаях, Восточной (кроме СССР), Южной и Юго-Восточной Азии, Новой Гвинее, на юге Северной Америки, в Центральной и Южной Америке.
Асфоделовые - преимущественно многолетние травы, реже древовидные и кустарниковидные растения, кустарнички и полукустарники либо очень редко лианы и однолетние травы. Немногим менее половины видов этого семейства являются листовыми суккулентами, из которых наиболее известен род алоэ (Aloe, табл. 19, 1). Травянистые представители асфоделовых имеют горизонтальное, вертикальное или косовосходящее корневище, от которого отходят мясистые придаточные корни, нередко утолщенные (веретеновидные, цилиндрические, клубневидные), служащие, как и корневище, вместилищами запасных питательных веществ. Очень немногие представители асфоделовых (некоторые виды алоэ) имеют луковицу, образованную подземными расширенными мясистыми основаниями листьев. У некоторых южноафриканских видов алоэ эти расширенные основания листьев имеют сочленение с узколинейными ассимилирующими пластинками; в месте сочленения пластинки отделяются и опадают. У одного из видов рода артроподиум (Artliropodium) в основании однолетних побегов развиваются небольшие конические клубни.
Таблица 19. Асфоделовые. Алоэ мыльное (Aloe saponaria): 1 - общий вид растения; 2 - бутон; 3 - цветок; 4 - отцветающий цветок. Асфоделина тонкая (Asphodeline tenuior): 5 - часть растения с соцветием и верхними листьями. Гастерия двурядная (Gastena disticha): 6 - общий вид растения; 7 - бутон; 8 - цветок; 9 - отцветшим цветок
Листья у большинства травянистых асфоделовых образуют прикорневую розетку или пучок, из середины которых или, чаще, из пазух листьев выходят цветоносы. Облиственные стебли имеют сравнительно немногие представители семейства (например, асфоделина - Aspliodeline, табл. 19, 5, табл. 20, 6, виды булъбины - Bulbine, несколько видов алоэ). У древесных видов листовые розетки расположены на верхушках ветвей или стволов. Листья очередные, многорядные, редко двурядные. Они очень разнообразны по форме, консистенции и продолжительности жизни. Нередко листья дифференцированы на пластинку и влагалище, а у листьев австралийского рода совербеа (Sowerbaea) имеется и язычок, как у злаков (рис. 75, 9).
Цветки асфоделовых обычно небольшие по размеру (самые крупные длиной не более 5 см), собраны в простые или сложные кисти, метелки, колосовидные, зонтиковидные и головчатые соцветия, расположенные на верхушке стебля и его ветвей либо на пазушных или центральных цветоносах, достигающих у некоторых растений высоты 1,5-3 м. Очень редко цветки одиночные. На оси соцветия они расположены по одному в пазухе прицветника либо в пазушных пучках (обычно 2-, 6-цветковых); на цветоножках часто имеется сочленение. Цветки чаще всего обоеполые, актиноморфные, реже более или менее зигоморфные. Околоцветник простой, венчиковидный, из 6 расположенных в 2 кругах свободных или в различной степени сросшихся между собой сегментов. Внутренние и наружные сегменты более или менее одинаковые или резко различающиеся, как у родов тизанотус (Thysanotus) и боттионея (Bottionea), внутренние сегменты которых значительно шире наружных и снабжены по краям бахромкой (рис. 74, 2). Околоцветник при завядании цветка обычно сохраняется довольно продолжительное время, полностью окружая развивающийся плод или располагаясь на его верхушке в виде шапочки (в родах гастерия - Gasteria, бульбина и др., рис. 67, 6). У растений австралийского рода ходжсониола (Hodgsoniola) и близких к нему родов околоцветник при отцветании спирально скручивается. Тычинок обычно 6, свободных или приросших к основаниям сегментов околоцветника, а иногда (австралийский род бория - Воrуа) к его зеву. В цветках некоторых родов имеется только 3 тычинки, а 3 другие полностью редуцированы или превращены в стаминодии. Нити тычинок обычно длинные и тонкие, но иногда внизу расширенные и сросшиеся между собой и околоцветником (род джонсония - Johnsonia). У рода асфоделина расширенные ниже середины нити образуют вокруг завязи колпачок, препятствующий проникновению к нектару мелких ползающих насекомых, неспособных произвести опыление (рис. 67, 8). У некоторых родов (например, у бульбины) тычиночные нити снабжены бородкой из густых волосков, которые, вероятно, задерживают высыпающуюся из пыльников пыльцу, препятствуя ее попаданию на рыльце собственного цветка. Пыльники обычно интрорзные" прикрепляются к нити спинкой (и в этом случае они обычно качающиеся) или основанием. Характерной особенностью асфоделовых является наличие у многих представителей в основании или посередине связника пыльника особой ямки или канавки, в которую входит тычиночная нить (рис. 64, 6). В роде эхеандия (Echeandia) пыльники срослись между собой в коническую трубку, окружающую столбик (рис. 72, 6). У рода ходжсониола трубку образуют удлиненные сросшиеся связники пыльников. Гинецей синкарпный, состоящий из 3 плодолистиков. Столбик обычно нитевидный, с очень маленьким головчатым, дисковидным" редко 3-лопастным рыльцем. Завязь верхняя,; 3-гнездная, иногда глубоко лопастная.
Рис. 64. Асфоделовые. Парадизея лилиевидная (Paradisea liliastrum): 1 - цветонос с соцветием и лист; 2 - продольный разрез цветка. Эремурус Ольги (Eremurus olgae): 3 - верхняя часть соцветия; 4 - лист; 5 - цветок; 6 - тычинка (на связнике видна ямка, в которую входит тычиночная нить); 7 - тычинка с раскрывшимся пыльником; 8 - семя с крыловидным ариллусом; 9 - нижняя часть растения с утолщенными мясистыми корнями. Эремурус бурый (Е. fuscus): 10 - цветок
В перегородках между гнездами завязи у некоторых родов были обнаружены септальные нектарники. Согласно Э. Дауманну (1970), они принадлежат к 3 типам. Нектарники первого типа имеют роды алоэ, гастерия, хавортия (Наworthia), книпхофия (Kniphofia), антерикум (Anthericum), хлорофитум (Chlorophytum) и парадизея (Paradisea). Эти нектарники крупные (нектарная щель каждого из них занимает почти всю перегородку между гнездами); через тонкий выводной каналец, выходящий у основания столбика, нектар по каплям выделяется на верхнюю поверхность завязи и стекает на дно околоцветника (см. рис. 73, 2). Видам рода эремурус (Eremurus) свойствен второй тип нектарников, представляющий собой редуцированный вариант нектарников алоэ. Они значительно короче последних, а у некоторых видов в одной или двух перегородках часто совсем не развиваются. Третий тип нектарников встречается у родов асфоделус (Asphodelus) и асфоделина. Они менее крупные, чем у алоэ, а выводной каналец их выходит не на верхушку завязи, а на ее боковую стенку. Это самый специализированный тип септального нектарника. В каждом гнезде завязи имеется от 2 или нескольких до многочисленных (12-30) ортотропных, гемитропных или анатропных семязачатков.
Плод у большинства асфоделовых - локулицидная коробочка. У австралийского рода коринотека (Corynotheca) плод часто орешковидный, невскрывающийся. Из 1-3 односемянных сухих или сочных и также невскрывающихся долей состоит плод растений из рода трикорина (Tricoryne). Мадагаскарский род ломатофиллум (Lomatophyllum) характеризуют сочные ягодовидные, но вскрывающиеся плоды. Семена имеют довольно крупный зародыш и мясистый или твердый эндосперм.
Семейство асфоделовых объединяет 2 подсемейства: собственно асфоделовые (Asphodeloideae) и антериковые, или венечниковые (Аnthericoideae). Эти два подсемейства различаются между собой по целому ряду признаков, в особенности эмбриологически. Так, подсемейство асфоделовых характеризуется симультанным (одновременным) типом образования микроспор, ортотропными и гемитропными семязачатками, гелобиальным эндоспермом, отсутствием гаусторий у зародышевого мешка и наличием ариллуса у семян (последний признак - одна из характерных особенностей подсемейства асфоделовых). Как указывает Г. А. Комар (1978), из всего семейства лилейных в широком его понимании настоящий ариллус имеется только у представителей подсемейства асфоделовых. Он крыловидный, беловатый, полупрозрачный, не срастающийся с наружным интегументом. У родов трибы асфоделовых (Asphodeleae) ариллус окружает семя полностью, а у трибы алоевых (Aloeae) - не полностью, обнаруживая некоторые черты редукции. В противоположность подсемейству асфоделовых представителям антериковых свойственны сукцессивный (последовательный) тип образования микроспор, анатропные и очень редко ортотропные (у родов совербея и лаксманния - Laxmannia) семязачатки, наличие гаусторий у зародышевого мешка, отсутствие ариллуса (у немногих родов имеется ариллоид - элайосома). Оба подсемейства обнаруживают довольно большие различия также в анатомическом строении листьев. У подсемейства асфоделовых листья унифациальные (т. е. имеющие только одну поверхность), с 2 рядами проводящих пучков. Хотя у большинства их представителей листья имеют как бы две поверхности, верхнюю и нижнюю, но, как показывают анатомические исследования, фактически они являются ложнодорсивентральными или ложнобифациальными. Листья видов под-семейства асфоделовых характеризуются еще и тем, что у них всегда есть водоносная паренхима. Она очень сильно развита у суккулентных родов и в редуцированной форме присутствует у слабо суккулентных и несуккулентных (например, у книпхофии, бульбины, асфоделуса и др.). У представителей подсемейства, антериковых листья обычно дорсовентральные с одним рядом проводящих пучков. Исключение составляют только роды трибы джонсониевых (Johnsonieae), имеющие унифациальные листья. Водоносная паренхима в листьях антериковых полностью отсутствует. Подсемейства асфоделовых и антериковых различаются также и по характеру подземных органов. Собственно асфоделовые имеют очень короткое, у некоторых родов почти дисковидное (донце) или бочонковидное корневище, от которого отходят большей частью утолщенные мясистые цилиндрические или веретеновидные корни. Для представителей подсемейства антериковых более характерно удлиненное горизонтальное или косовосходящее корневище с клубневидно утолщенными у концов корнями. Некоторые ботаники, как X. Хубер (1969) и Р. Дальгрен (1980), возводят эти подсемейства в ранг самостоятельных семейств. Однако в строении цветка оба подсемейства не обнаруживают существенных различий.
Подсемейство асфоделовых состоит из 11 родов (около 850 видов); 10 из них встречаются в восточном полушарии, а один монотипный род глифосперма (Glyphosperma) - в Мексике. Подсемейство объединяет три трибы: асфоделовые, алоевые и книпхофиевые (Kniphofieae).
В трибу асфоделовых входят 6 родов, включающих обычно слабо суккулентные или несуккулентные и большей частью не вечнозеленые многолетние или (редко) однолетние травы и полукустарники. Водоносная паренхима присутствует в листьях только в редуцированной форме, и вместилищами воды и запасных питательных веществ являются разнообразно утолщенные мясистые корни. Цветонос у растений этой трибы обычно центральный; околоцветник не мясистый, обычно звездчатый, колесовидный, редко чашевидный или колокольчатый, со свободными или немного сросшимися при основании сегментами. В каждом гнезде завязи находится от многочисленных до 2 семязачатков. Плод - коробочка.
Род эремурус из трибы асфоделовых - характерный представитель ирано-туранской флоры. В роде свыше 60 видов, распространенных на юго-востоке Европы (Крым, юго-восток Украины) и в Азии от Ливана до запада Центральной Азии (Джунгария), Алтая и северо-западных Гималаев. Изолированно от основного ареала рода произрастает эремурус китайский (Eremurus chinensis), немногие местонахождения которого лежат в Юго-Западном Китае. Наиболее богаты видами эремуруса Средняя Азия (45 видов), в особенности Памиро-Алай, и Афганистан (15-20 видов). Большинство видов встречается в горах - от предгорий до субальпийского пояса, но преимущественно в степном поясе и нижней части лесного. Растут они обычно по открытым солнечным склонам. Некоторые виды эремурусов, встречаясь в большом количестве, играют заметную роль в сложении растительного покрова сухих горных склонов.
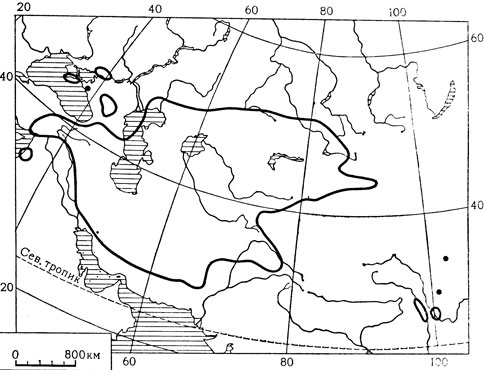
Карта 3. Ареал рода эремурус
Эремурусы - многолетние растения с густой приземной розеткой линейных листьев длиной 30-100 см и высоким (до 2 м у эремуруса мощного - Е. robustus, табл. 20, 7) центральным простым безлистным цветоносом с длинными, нередко густыми султановидными кистями из белых, розовых, желтых, зеленовато-желтых или оранжевых очень многочисленных цветков. В соцветии, например у эремуруса Ольги (Е. olgae, рис. 64, 5), свыше 500 цветков. Околоцветник бывает колесовидный, колокольчатый или чашевидный, со свободными или немного сросшимися у основания сегментами. Корневище эремуруса вертикальное, короткое, имеет форму утолщенного диска, бочонка или усеченного конуса. От него радиально отходят веерообразно расположенные мясистые, часто утолщенные веретеновидные, иногда цилиндрические, редко шнуровидные корни. Эремурусы цветут с апреля до середины лета. Раскрывание цветков в соцветии происходит снизу вверх. Зона цветения постепенно передвигается к верхушке, а в нижней части соцветия уже зреют плоды. Биология цветения эремурусов весьма своеобразна. Еще Ф. Хильдебранд (1881) отметил необычное явление при цветении эремуруса замечательного (Е. spectabilis). Вскрывание пыльников и опыление рыльца у этого вида происходило только в том случае, если первоначально широко раскрытые цветки закрывались. Эту особенность ученый назвал сенсационным исключением из правил. Известные русские ботаники Б. А. Федченко (1899) и его мать, монограф рода эремурус О. А. Федченко (1909), описавшие движения сегментов околоцветника, тычинок и столбика в процессе цветения эремуруса замечательного, эремуруса Ольги (названного в честь О. А. Федченко) и эремуруса мощного, обратили внимание на тот факт, что у первого вида насекомые посещают только цветки с завернутыми внутрь лепестками, в то время как у двух других они садятся и на раскрытые цветки.

Таблица 20. Асфоделовые: 1 - эремурус Ольги (Eremurus olgae), Крым, Никитский ботанический сад; 2 - бульбина широколистная (Bulbine latifolia), оранжереи Ботанического института АН СССР в Ленинграде; 3 - книпхофия ягодная (Kniphofia uvaria), Батумский ботанический сад; 4 - артроподиум усиконосный (Arthropodium cirrhatum), оранжереи Ботанического института АН СССР в Ленинграде; 5 - хавортия батесианская (Haworthia batesiana), там же; 6 - асфоделина желтая (Asphodeline lutea), Крым, Никитский ботанический сад; 7 - эремурус мощный (белый) (Eremurus robustus) и эремурус согдийский (желтый) (Е. sogdianus), там же
Разное поведение насекомых связано с тем, что у первого вида пыльники вскрываются только после закрывания цветков, а у остальных - вскоре после их раскрывания. Как показали исследователи У. Даммер (1888), П. Кнут (1904), А. П. Хохряков и М. Т. Мазуренко (1969) и др., цветки эремуруса замечательного и других видов этого рода являются протандричными.
Э. Дауманн (1968) очень детально исследовал процесс цветения у эремуруса замечательного и эремуруса гималайского (Е. himalaicus). Весь процесс цветения этих растений (от раскрывания цветков до их увядания) длится око-ло 60 ч. В течение этого периода у обоих видов наблюдаются 4 основные фазы цветения, продолжительность которых и характер протекания различны (рис. 65). Первую фазу, во время которой в раскрывшихся цветках не функционируют ни тычинки, ни рыльце и не происходит выделения нектара, называют бесполой. У эремуруса замечательного она длится 25 ч. Примерно за 5 ч до наступления следующей, мужской фазы сегменты околоцветника, прежде почти горизонтально отклоненные, поднимаются, их верхушки буреют и заворачиваются внутрь цветка, который таким образом оказывается почти закрытым. Из него высовываются тычинки и столбик. В эту вторую фазу, протекающую 9 ч, тычинки расположены косо вверх, нити удлинены, пыльники вскрыты, а столбик, тоже удлинившийся, отогнут вниз; начинается слабое выделение нектара. Третья фаза цветения - обоеполая. В течение 16 ч функционируют и пыльники, и достигшее зрелости рыльце; усиливается секреция нектара. Но самоопыления цветков, по-видимому, не происходит, так как выпрямившийся и удлинившийся столбик заметно превышает тычинки. Наконец, в четвертую, женскую фазу в течение 11 ч функционирует только рыльце, столбик остается в прежнем положении, тычинки засыхают и опускаются вниз; выделение нектара в начале этой фазы достигает максимума. У эремуруса гималайского первая фаза продолжается только 4 ч, а следующая (вскрывание пыльников) наступает вскоре после раскрывания цветка и длится 20 ч. В течение первой и второй фаз столбик отогнут вниз, а околоцветник раскрыт. В третьей, обоеполой фазе цветения (24 ч) сегменты околоцветника поднимаются, образуя широкий колокол, столбик выпрямляется и превышает тычинки. Женская фаза протекает аналогично с таковой у эремуруса замечательного. Эремурусы опыляют медоносная пчела, шмели и мухи-журчалки. Насекомые собирают с них главным образом пыльцу и в значительно меньшей степени нектар, которого у многих эремурусов образуется мало, поскольку их септальные нектарники находятся в рудиментарном состоянии.
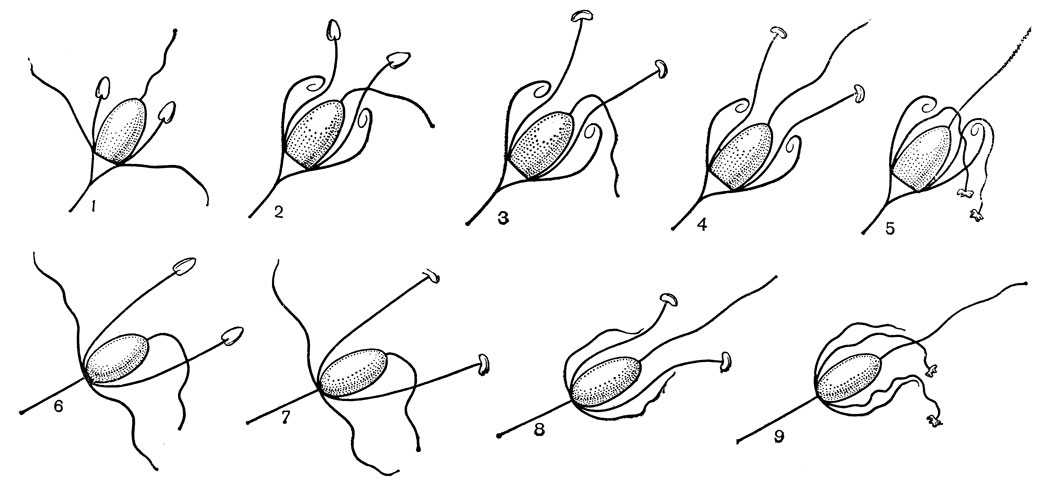
Рис. 65. Схематическое изображение фаз цветения эремурусов. Эремурус замечательный (Eremurus spectabilis) и эремурус гималайский (Е. himalaicus): 1, 6 - бесполая фаза; 2, 3, 7 - мужская фаза; 4, 8 - обоеполая фаза; 5, 9 - женская фаза
Эремурусы, как и другие представители асфоделовых, являются растениями-баллистами. Их коробочки всегда ориентированы прямо вверх, а плодоножки направлены косо вверх или горизонтально. При раскачивании цветоноса ветром, животными или человеком крылатые семена высыпаются из коробочек и падают на землю на небольшом расстоянии от материнского растения. Размножаются эремурусы только семенами. Многие виды - необычайно красивые растения. Их разводят как декоративные в районах с умеренно-теплым климатом. Но, как показал опыт культуры эремурусов в Главном ботаническом саду АН СССР в Москве, их можно выращивать и в умеренной зоне. Корни различных эремурусов иногда используют для получения клея. Клеящее вещество представляет собой особый полисахарид эремуран, считающийся ценным заменителем гуммиарабика. Молодые листья и богатые крахмалом корни иногда употребляют в пищу. Многие эремурусы - прекрасные медоносы и перганосы.
Другой род трибы асфоделовых - асфоделус (Asphodelus) - объединяет 12 видов, произрастающих на Канарских островах и острове Мадейра, в Средиземноморье, Западной Азии (исключая Кавказ) до Ирана; 5 видов встречаются на юге Европы, от Азорских островов до Балканского полуострова. Асфоделусы обитают на сухих каменистых и песчаных местах, встречаясь от уровня моря до субальпийского пояса. Это многолетние, иногда однолетние или двулетние (асфоделус дудчатый - Asphodelus fistulosus, рис. 66, 4) травы с прикорневой розеткой линейных или полуцилиндрических листьев, с простым или разветвленным центральным цветоносом высотой от нескольких сантиметров, как у североафриканского асфоделуса бесстебелъного (A. acaulis), до 1-2 м. Корни асфоделусов обычно толстые, веретеновидные или редьковидные. Цветки длиной 5-40 мм, воронковидные или почти колесовидные, белые или бледно-розовые, со свободными или сросшимися только в основании сегментами. Септальные нектарники хорошо развиты и выделяют обильный нектар. Коробочка обычно морщинистая, с 6 семенами. Встречающийся по всему Средиземноморью асфоделус летний (A. aestivus) - крупное растение с разветвленным цветоносом высотой до 2 м и рыхловатыми кистями белых цветков. Он растет вблизи морских побережий по пескам, сухим травянистым и каменистым склонам и осыпям, образуя местами большие заросли. Асфоделус летний применяют в народной медицине со времен Гиппократа, Теофраста и Диоскорида. Толстые корни иногда используют для получения клея и окрашивания шерсти в желтый и желто-коричневые цвета. Асфоделус белый (A. albus), имеющий простой с густой кистью цветонос высотой 30-100 (150) см, очень похож внешне на некоторые виды эремуруса (рис. 66, 1). Он растет в Южной, отчасти Средней и Атлантической Европе на горных лугах, в разреженных лесах и верещатниках. Асфоделус белый разводят как декоративное растение. По всему ареалу рода распространен асфоделус дудчатый - небольшое растение, имеющее узкие полуцилиндрическйе листья и полый, почти от основания разветвленный цветонос высотой 15-70 см с рыхлыми кистями мелких цветков (рис. 66, 4). Он растет по сухим песчаным и каменистым местам, краям дорог, в посевах. Листья его иногда употребляют в пищу.
К асфоделусу относительно близок род асфоделина, в котором 15-16 видов, распространенных в Восточном Средиземноморье, Крыму и Западной Азии (Кавказ, Малая Азия, Иран). Асфоделины растут в горах и на равнине по су-хим каменистым и щебнистым местам, степным склонам, светлым сухим лесам и кустарникам, скалам. Асфоделина крымская (Asphodeline taurica), встречающаяся на Балканском полуострове, в Крыму и на Северном Кавказе, иногда образует асфоделиновые степи. У асфоделины, в отличие от большинства представителей семейства, высоко (до половины или до самого соцветия) и густо облиственный стебель, иногда ветвистый (как у однолетней кавказской асфоделины древовидной - A. dendroides). Листья имеют узколинейные или линейно-шиловидные пластинки, верхушки которых располагаются на одном уровне, и пленчатые влагалища. Соцветие этого растения кистевидное, от очень густого до рыхлого, обычно образованное многочисленными пучками из 3-5 цветков. Своеобразно выглядят молодые соцветия асфоделины крымской. Они крупные, густые и толстые, как початки, и сплошь серебристо-белые от крупных, пленчатых, полупрозрачных, значительно превышающих бутоны прицветников. Цветки асфоделины белые или желтые, почти колесовидные или воронковидные. Тычинки неравные (внутренние длиннее наружных). Основания тычиночных нитей расширены и плотным колпачком окружают завязь, благодаря чему обильно выделяющийся нектар оказывается недоступным для мелких ползающих насекомых, которые не могут участвовать в опылении. Зацветание асфоделины начинается в нижней части соцветия, причем в каждом его пучке распускается сначала только один цветок. Зона цветения постепенно передвигается вверх, и, когда она доходит примерно до середины, в нижней части соцветия появляется вторая зона цветения, что связано с раскрыванием следующих цветков в пучках.

Рис. 66. Асфоделовые. Асфоделус белый (Asphodelus albus): 1 - общий вид; 2 - коробочка; 3 - семя с крыловидным ариллусом. Асфоделус дудчатый (A. fistulosus): 4 - общий вид; 5 - коробочка. Асфоделина крымская (Asphodeline taurica): 6 - общий вид; 7 - цветок
У полностью раскрывшихся цветков асфоделины крымской и асфоделины желтой (A. lutea, табл. 20, 6) сегменты околоцветника расположены зигоморфно: один, наружный сегмент обращен вниз, а пять остальных, сближенных между собой сегментов обращены вверх. Вскрывание пыльников происходит вскоре после распускания цветка. Опылителями асфоделины крымской являются пчелы и шмели. У кавказского вида асфоделины тонкой (A. tenuior, табл. 19, 5) раскрывание цветков происходит к 5-6 ч вечера, а утром следующего дня все раскрывшиеся накануне цветки находятся в увядшем состоянии. Вечернее распускание цветков, несомненно, связано с опылением их ночными или сумеречными насекомыми. Асфоделина либурнская (A. liburnica), распространенная в Средиземноморье от Италии до Турции, опыляется сумеречными бабочками-бражниками. Асфоделины - хорошие медоносы. Некоторые виды, например асфоделину желтую, разводят как декоративное растение.
Родом бульбина мы завершим обзор трибы асфоделовых. К этому роду принадлежат около 55 видов, распространенных в Южной Африке и отчасти тропической Африке (4 вида) и Австралии (3 вида). Это многолетние травы с облиственным стеблем или чаще с прикорневой розеткой листьев и простым цветоносом (табл. 20, 2) или, реже, сильно ветвистые полукустарники. Листья этих растений более или менее суккулентные, полуцилиндрические, линейные или ланцетные, нередко с влагалищами. Цветки мелкие, обычно ярко-желтые, редко белые, собранные в цилиндрическую или щитковидную многоцветковую кисть. Сегменты околоцветника свободные, расположены звездчато или отогнуты вниз. Тычиночные нити снабжены длинными густыми и пушистыми, расположенными немного ниже пыльников волосками, придающими цветкам декоративный вид (рис. 67, 2-5). Полагают, что эти волоски служат приспособлением против самоопыления, так как они задерживают высыпающуюся пыльцу из пыльников. Тычинки у бульбин обычно одинаковые, но у австралийской булъбины полубородчатой (Bulbine semibarbata) тычинки внутреннего круга длиннее тычинок наружного круга, которые к тому же лишены волосков (рис. 67). Некоторые виды бульбин разводят как декоративные.
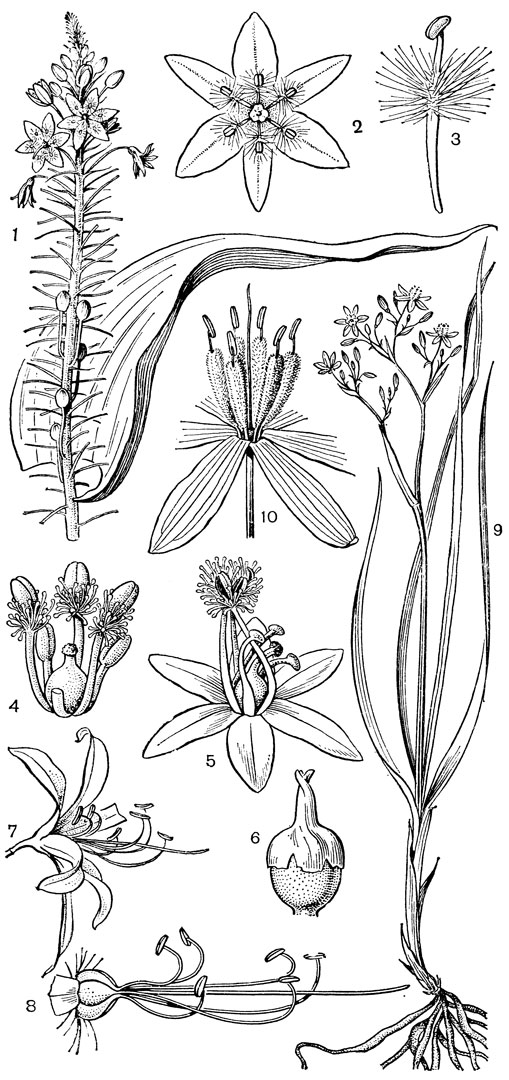
Рис. 67. Асфоделовые. Бульбина широколистная (Bulbine latifolia): 1 - верхняя часть соцветия и лист; 2 - цветок; 3 - тычинка с волосками. Бульбина полубородчатая (В. semibarbata): 4 - положение тычинок и столбика в бутоне (околоцветник и одна тычинка удалены); 5 - положение тычинок и столбика в раскрывшемся цветке; 6 - плод с остатком ('шапочкой') околоцветника. Асфодели на тонкая (Asphodeline tenuior): 7 - цветок; 8 - цветок (околоцветник удален), видны расширенные основания тычиночных нитей, плотным колпачком окружающие завязь. Симетис плосколистный (Simethis planifolia): 9 - общий вид; 10 - цветок
В трибу алоевых входят 4 рода и более 600 видов, подавляющее большинство которых является суккулентными ксерофитами, свойственными преимущественно аридным областям Южной и тропической Африки и Мадагаскара. Эти растения имеют толстые, мясистые, сочные, обычно вечнозеленые листья, почти целиком состоящие из бесцветной водоносной паренхимы, расположенной за узкой полосой хлоренхимной ассимилирующей ткани. Водоносная паренхима несет не только водозапасающую функцию, но является и вместилищем запасных питательных веществ. Листья часто имеют толстую кутикулу и глубоко погруженные тетрацитные устьица. Триба включает многолетние травы, а также большое число древовидных и кустарниковидных форм, стволы и ветви которых имеют вторичный рост однодольного типа. Околоцветник у алоевых сочный, мясистый, обычно трубчатый, редко колокольчатый, иногда со звездчатым либо завернутым наружу отгибом; сегменты его свободные или, чаще, более или менее сросшиеся. Завязь с многочисленными ортотропными или гемитропными семязачатками в каждом гнезде. Плод - локулицидная коробочка и только у мадагаскарского рода ломатофиллум (Lomatophyllum) он сочный и ягодовидный, но все же раскрывающийся. Семена обычно плоские и крылатые.
Самым крупным и широко известным родом трибы алоевых является род алоэ (Aloe, табл. 19, 1). В нем около 350 видов, произрастающих в Южной и тропической Африке, на острове Мадагаскар, Аравийском полуострове, а также на острове Сокотра и в Макаронезии. Наиболее богаты видами алоэ Капская область и Трансвааль, а в тропической Африке - Эфиопия и Сомали. Виды алоэ произрастают преимущественно в областях с жарким и крайне сухим климатом, где встречаются от приморской полосы до высоты 2500 м над уровнем моря. В некоторых местах, например на севере плоскогорья Намакваленд в Юго-Западной Африке и в пустыне Намиб, своеобразные суккулентные пустыни и полупустыни образованы алоэ. Многие виды алоэ встречаются в саваннах по песчаным и каменистым местам, нередко среди крупных камней. Очень часто алоэ являются ландшафтными растениями.
Алоэ - очень разнообразные по своему внешнему облику растения. Преобладают среди них многолетние травы, хотя нередко встречаются также древовидные и кустарниковидные формы, иногда и лианы. Листья этих растений обычно суккулентные, толстые, мясистые, очень сочные, редко слабо суккулентные или кожистые и жесткие. Они образуют прикорневые или верхушечные (у древесных форм) розетки, в которых располагаются по спирали или иногда двурядно. Отчетливую спираль образуют короткие дельтовидные листья южноафриканкого алоэ многолистного (A. polyphylla), широко известного под названием спирального алоэ. У сравнительно немногих видов листья располагаются как на верхушке, так и по всей длине побега. По форме листья бывают ланцетными, линейно-ланцетными, мечевидными, дельтовидными, реже линейными. Края листьев обычно снабжены крепкими шиповидными зубцами. У некоторых видов обе поверхности листьев усажены твердыми колючками. В пазухах листьев образуются простые или разветвленные, 3 иногда очень высокие (до 2-3 м) цветоносы. Очень редко цветоносы бывают центральные, развивающиеся из верхушечной почки. Цветки у алоэ обычно крупные (длиной 3,5-5 см и диаметром около 1 см), красные, оранжевые, желтые или редко белые. Они собраны в кисти, метельчатые или колосовидные соцветия. Кисти варьируют от удлиненных (до 1 м) до сильно укороченных - щитковидных и почти головчатых. У некоторых видов кисти односторонние. Их оси расположены почти горизонтально, а цветки обращены в одну сторону - прямо или косо вверх (рис. 68, 69). Околоцветник сочный, мясистый, обычно трубчатый, иногда колокольчатый, цилиндрический или слегка трехгранный, прямой или несколько изогнутый, внизу нередко почти шаровидно расширенный, обычно актиноморфный, очень редко двугубый. Сегменты околоцветника свободные или (чаще) в разной степени сросшиеся; свободные их части прямые или более или менее сильно изогнуты наружу и имеют более бледную окраску, чем трубка околоцветника, обычно зеленоватую. Тычинок 6, равных или неравных по длине, о линейными или продолговатыми пыльниками, прикрепляющимися к нити основанием. В раскрытых цветках тычинки сближены все вместе, примыкая, как и столбик, к стороне околоцветника, обращенной к оси соцветия. Плоды у алоэ большей частью почти цилиндрические, в кожистые или деревянистые, с многочисленными неправильно 3-гранными или уплощенными семенами, окруженными беловатым полупрозрачным крыловидным ариллусом.
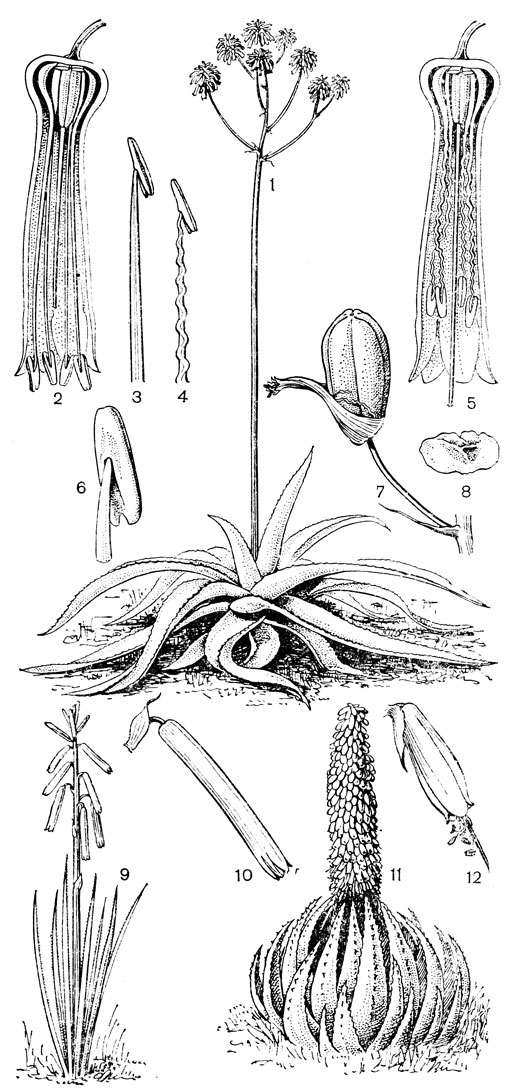
Рис. 68. Виды алоэ. Алоэ Дукера (Aloe duckeri): 1 - общий вид; 2 - продольный разрез цветка в мужской фазе цветения; 3 - тычинка с прямой тычиночной нитью; 4 - тычинка с извитой тычиночной нитью; 5 - продольный разрез цветка в женской фазе цветения (тычинки втянуты в околоцветник); 6 - тычинка (на связнике видна канавка, в которую входит тычиночная нить); 7 - плод с остатком околоцветника; 8 - семя с крыловидным ариллусом. Алоэ книпхофиевидное (A. kniphofioides): 9 - общий вид; 10 - цветок. Алоэ Пеглер(А. peglerae): 11 - общий вид; 12 - цветок
Цветки алоэ очень богаты нектаром, вытекающим у основания столбика из септальных нектарников и заполняющим нижнюю часть околоцветника. Яркая окраска цветков и изобилие нектара привлекают разных опылителей. По данным С. Фогеля (1954), большинство видов алоэ являются орнитофильными. Алоэ неустрашимое (A. ferox), например, опыляют небольшие птицы-нектарницы. Они садятся на его соцветие ниже цветка, из которого пьют нектар,, погружая клюв в околоцветник, или на ближайший к соцветию лист (рис. 69, 7-8). Нектариицы опыляют и другие виды алоэ, в частности, алоэ многолистное, алоэ дихотомическое (A. dichotoma). Последний вид посещают и пчелы, которые опыляют также алоэ бело-цветковое (A. albiflora) и, по-видимому, многие другие виды. Алоэ наименьшее (A. minima) опыляют дневные бабочки, а алоэ Сюзанны (A. suzannae) - ночные. Цветки алоэ протандричны. Вначале из распустившегося цветка показываются тычинки с уже вскрывшимися пыльниками, несущими обильную комковатую (не сыпучую) пыльцу. Вскоре выставляется и столбик с маленьким головчатым рыльцем, которое располагается на одном уровне с пыльниками или незначительно выше или ниже их. При таком расположении тычинок и столбика не исключено попадание пыльцы на рыльце того же цветка. Но, по-видимому, вначале своей экспозиции рыльце еще не зрелое и не способно воспринять пыльцу. Позже, вероятно, происходит совпадение мужской и женской фаз цветения, и в это время возможно самоопыление, которое происходит, по-видимому, в ограниченных пределах из-за наличия комковатой пыльцы. Спустя некоторое время после одновременной экспозиции тычинок и столбика можно наблюдать, что из цветка высовывается лишь один, несколько удлинившийся столбик. Если цветок в этой стадии цветения вскрыть, то можно увидеть, что тычинки находятся внутри околоцветника. Произошло же это потому, что их первоначально прямые нити зигзагообразно извились, в результате чего длина их сократилась и тычинки оказались втянутыми в околоцветник (рис. 68, 5). Оставшись один, столбик может теперь опылиться только пыльцой с другого растения или с другого цветка своего растения.

Рис. 69. Виды алоэ. Алоэ многолистное (Aloe polyphylla): 1 - общий вид. Алоэ реснитчатое (A. ciliaris): 2 - ветвь с соцветием; 3 - часть стебля с двумя влагалищами листьев (нижнее - в продольном разрезе) и основаниями листовых пластинок (по краю верхнего влагалища видны реснички). Алоэ хохолковое (A. comosa): 4 - общий вид; 5 - часть соцветия, раскрывшиеся цветки и молодые бутоны прижаты к его оси, а цветки, находящиеся между ними, отклонены от оси, образуя как бы хохолок. Алоэ Пиланса (A. pillansii): 6 - общий вид. Алоэ неустрашимое (A. ferox): 7 - общий вид (ствол скрыт за увядшими листьями); 8 - часть соцветия с сидящей на нем нектарницей, пьющей из цветков нектар
Виды алоэ, подобно большинству представителей семейства асфоделовых, являются растениями-баллистами. При раскачивании цветоноса ветром или животными семена из раскрывшихся плодов высыпаются па землю неподалеку от материнского растения. Крыловидный ариллус придает семенам некоторую летучесть и способствует их лучшему рассеиванию. Семенное возобновление является единственным способом размножения у большинства алоэ. Но некоторые виды, например алоэ настоящее (A. vera) и алоэ пестрое (A. variegata), размножаются также и вегетативно посредством подземных столонов, которые, выходя на поверхность земли, развивают молодые розетки. Мадагаскарское алоэ луковичконосное (A. bulbillifera) является живородящим. В его соцветии, в пазухах нижних прицветников вместо цветков развиваются луковички, которые, падая на землю, при благоприятных условиях могут дать начало новым растениям.
Познакомимся теперь с некоторыми наиболее интересными видами алоэ. Начнем с древовидных его представителей. Одни из них, например алоэ дихотомическое, имеют разветвленный ствол и достигают в высоту 9-18 м. Эти алоэ отличаются от настоящих деревьев тем, что вторичное утолщение у них (как и у всех древовидных однодольных) происходит за счет деятельности клеток меристематической зоны, расположенной по периферии ствола, а не за счет камбия, как у двудольных. Самым крупным и высоким из всех древовидных алоэ является алоэ Байнеса (A. bainesii), растущее в Капской области и в Юго-Восточной Африке в густых кустарниковых зарослях и невысоких лесах по склонам гор и холмов. Это довольно стройное развесистое древовидное растение высотой 10-18 м, с гладким стволом диаметром 1-2 (3) м несет на верхушках ветвей розетки длинных (60-90 см) темно-зеленых изогнутых листьев и плотные кисти розовых цветков. Алоэ Байнеса очень декоративно, и его часто выращивают в садах и парках Африки. В пустынных районах Капской области и Юго-Западной Африки, лишенных какой-либо иной древесной растительности, по каменистым склонам и вершинам холмов среди крупных камней растет алоэ дихотомическое - растение очень своеобразного, причудливого облика. Оно высотой 6-9 м (ствол диаметром около 1 м), сильно дихотомически разветвленное, с почти вертикальными довольно длинными беловато-серыми голыми ветвями, увенчанными розетками очень толстых листьев и метелками из нескольких густых лимонно-желтых кистей. Цветки алоэ дихотомического опыляют птицы-нектарницы и несметное количество пчел. Цветущее алоэ привлекает также и павианов, которые разрывают кисти цветков, высасывая из них нектар. Местные жители (бушмены и готтентоты) использовали в прошлом выдолбленные ветви алоэ дихотомического, как колчаны для стрел. Поэтому его называют еще колчанным деревом.
На алоэ дихотомическое очень похоже внешне алоэ Пиланса (A. pillansii), но оно более высокое (до 10 м) и стройное, обычно менее разветвленное (рис. 69, 6). Алоэ Пиланса растет также в Южной Африке, пустыне Намиб и в аридных районах плоскогорья Намакваленд - на плоских каменистых вершинах невысоких холмов. Уникальная среди алоэ особенность этого вида состоит в том, что его цветоносы, несущие до 50 кистей лимонно-желтых цветков, выходят из пазух самых нижних (а не верхних) листьев розетки. Сам цветонос при этом свешивается вниз, а его боковые ветви с кистями изогнуты кверху. Ветвистые древовидные алоэ встречаются также в тропической Африке (Сомали) и на юге Аравийского полуострова. В Южной Африке по каменистым склонам гор встречается алоэ складчатое (A. plicatilis) - невысокое, слабо дихотомически разветвленное деревце или кустарник, с гладкими ветвями и двурядно расположенными необычными для алоэ листьями. Они узкоэллиптические, плоские, утолщенные, кожистые, с совершенно гладкими краями и округлой верхушкой. Увядшие его листья быстро опадают, оставляя слабый рубец. Одним из самых широко распространенных видов алоэ является алоэ древовидное (A. arborescens), хорошо известное читателю по комнатной культуре. В комнатных условиях (в умеренном климате) это алоэ цветет крайне редко и именно с этой его особенностью связано его народное название - столетник (как якобы цветущее один раз в сто лет). Но на своей родине, в засушливых районах Южной и тропической Африки, алоэ древовидное цветет ежегодно, образуя довольно длинные (до 80 см) цветоносы с густыми коническими кистями алых крупных цветков (длиной 4 см). Алоэ древовидное - низко и сильно ветвящееся деревце высотой 2-3(5) м, со стволом толщиной 30 см или кустарник. Его листья образуют крупные густые розетки диаметром до 80 см (рис. 70, 7). Растет оно на каменистых местах, нередко среди кустарниковой растительности, встречаясь от уровня моря до 1800 м. Алоэ древовидное исключительно декоративно, легко размножается черенками, и поэтому его выращивают чаще других видов этого рода. Как декоративное растение алоэ разводят на его родине, а также в Юго-Западной Европе и Северной Африке, где оно нередко натурализуется. В СССР это растение выращивают как лекарственное на Черноморском побережье Кавказа.

Рис. 70. Асфоделовые. Книпхофия ягодная (Kniphofia uvaria): 1 - общий вид (слева - розетка нового побега); 2 - основание старого побега с почками возобновления и горизонтальным побегом-столоном; 3 - цветок. Алоэ Мендеса (Aloe mendesii): 4 - общий вид растения, свисающего со скалы. Алоэ гребенчатое (A. ortolopha): 5 - общий вид растения с односторонними кистями. Алоэ Ричардса (A. richardsii): 6 - луковица с отходящими от ее основания утолщенными корнями. Алоэ древовидное (A. arborescent): 7 - общий вид
Большую группу древовидных алоэ составляют растения с простым, неветвистым, относительно невысоким (обычно 2-3 м, но иногда до 8 м) стволом, увенчанным крупной розеткой листьев. Такие алоэ похожи на пальмы. Одним из них является алоэ неустрашимое (рис. 69, 7), широко распространенное во многих районах Южной Африки по холмам и равнинам, на каменистых почвах с низким травяным покровом. В Капской области этот вид нередко является ландшафтным растением. Края его листьев, а также обе их поверхности обычно обильно усажены твердыми шиповидными зубцами, за что это алоэ и получило свое видовое название.
Немногие виды алоэ являются кустарниковидными лианами. Из них наиболее известно распространенное в Южной Африке алоэ реснитчатое (A. ciliaris) с почти плоскими линейно-ланцетными пластинками листьев, влагалища которых по верхнему краю усажены мягкими белыми ресничками (рис. 69, 2, 3). Ветвистый тонкий ствол этого растения высотой до 6 м цепляется (опирается) за растущие рядом деревья и кустарники.
Самую многочисленную группу видов алоэ составляют многолетние травы с прикорневой розеткой листьев, отличающиеся большим разнообразием внешнего облика. Среди них есть совсем небольшие растения, высотой 20-40(50) см, с узколинейными, как у многих злаков, но более или менее суккулентными листьями. Их называют злаковидными алоэ. Таковы, например, алоэ колючелистное (A. myriacantha) - один из самых распространенных видов рода, ареал которого охватывает тропическую и Южную Африку, и алоэ кпипхофиевидное (A. kniphofioides), встречающееся в Южной Африке, имеющее самые крупные (до 5 см) в роде алоэ ярко-красные цветки (рис. 68, 10). Оба вида растут на каменистых местах, занятых другой низкотравной растительностью, и в нецветущем состоянии их почти невозможно найти. В необычных для алоэ местообитаниях встречается другой злаковидный вид - алоэ облачное (A. nubigena). Это растение образует густые дерновины на влажных замшелых выступах скал, в тенистых, богатых перегноем местах, иногда в брызгах водопадов.
Несколько видов алоэ имеют подземную луковицу. Она есть, например, у алоэ Бутнера (A. buettneri), имеющего самый большой из всех алоэ ареал. Этот вид встречается почти по всей тропической Африке, в изобилии произрастая по сухим травянистым склонам, в открытых и кустарниковых саваннах и даже в местах, которые в сезон дождей бывают затопленными и растения оказываются наполовину в воде. Луковицы у алоэ Бутнера довольно крупные (диаметром 8-10 см) и служат настоящими запасающими органами; листья не вечнозеленые, как у большинства алоэ; зимой они высыхают и исчезают.
К травянистым многолетникам принадлежит алоэ настоящее, более известное под названием алоэ барбадосское (A. barbadensis). Это растение образует довольно компактные розетки широколанцетных, сизовато-зеленых, почти прямостоячих листьев и цветонос высотой 60-90 см. Относительно родины алоэ настоящего нет единого мнения. Одни авторы полагают, что этот вид произрастает на островах Канарских и Зеленого Мыса; другие считают областью его естественного произрастания Северо-Восточную Африку, а возможно также и Аравийский полуостров. С давних времен алоэ настоящее культивировали как лекарственное растение по всему Средиземноморью, и во многих районах оно натурализовалось. Испанцами это растение было завезено в Новый Свет, где оно также натурализовалось, особенно широко в Вест-Индии - на острове Барбадос. С этого острова оно в конце XVI в. попало в ботанические сады Англии. В культуре алоэ настоящее распространилось в Индию, Южный Китай и некоторые районы Юго-Восточной Азии. Существует мнение, что алоэ настоящее упоминается в аккадских текстах (2000 лет до н. э.) древней Ассиро-Вавилонии как растение для украшения дверей дома. Древнейший обычай некоторых народов Ближнего Востока - вешать алоэ над входом в дом, особенно новый,- был связан с тем, что растению приписывали магические свойства. Считали, что оно способствует долгой жизни и процветанию обитателей дома и самого дома. Этот обычай сохранялся в Египте до середины XIX в., кое-где он существует и в настоящее время. Висящие на домах растения алоэ могли жить в течение нескольких лет без воды и даже цвести! На аккадском языке (древнейшем из всех известных семитских языков) алоэ настоящее обозначалось si-bu-ru. От него произошло арабское sabr, или saber, что означает "терпение", "выносливость". Этим же словом называют также получаемый из листьев разных видов алоэ (их около 15) сухой сгущенный сок сабур, который с древних времен широко использовали как лекарственное средство. Первые сведения о лечебных достоинствах сухого сока алоэ настоящего имеются еще у Диоскорида (около 78 лет н. э.). В его работе было и цветное изображение этого растения в бутонах. Но еще раньше грекам было, по-видимому, известно о лекарственном значении алоэ с острова Сокотра. В сочинениях известного арабского путешественника и географа Идриси (середина XII в.) говорится о том, что остров Сокотра был завоеван Александром Македонским из-за того, что на острове производили целебный сухой сок алоэ. О разнообразном медицинском использовании сока алоэ сообщалось в трактатах классика античной медицины римского врача и естествоиспытателя Галена. Медицинское значение имеет и мякоть (пульпа) свежих листьев алоэ - бесцветная сочная водоносная паренхима, образующая основную массу листа. Свежие листья алоэ имеют широкое применение в народной медицине некоторых тропических и субтропических стран. В СССР медицинское использование имеет сок алоэ древовидного, культивируемого на Черноморском побережье, в Аджарии. Сок извлекают прессованием измельченных свежесобранных листьев.
Многочисленные виды алоэ являются излюбленными декоративными растениями, широко разводимыми в африканских садах. К ним принадлежит и алоэ древовидное. Этот вид, а также алоэ неустрашимое и некоторые другие виды часто используют в Африке для живых изгородей. Несколько видов алоэ культивируются как декоративные в Европе, на побережье Средиземного моря, некоторые из них там натурализовались.
Род гастерия (Gasteria), также входящий в трибу алоевых, эндемичен для Южной Африки. Он включает около 70 видов суккулентных многолетних трав, произрастающих главным образом в ее центральных районах, по каменистым и мелкоземистым местам, нередко среди травяного покрова. Это небольшие растения с плотной розеткой многорядных или двурядных, толстых, мясистых, удлиненно-эллиптических листьев с сильно развитой кутикулой и обычно с мелкими белыми пятнами. Пазушные, несколько изогнутые наверху цветоносы гастерии оканчиваются рыхлой, обычно односторонней (из-за свешивающихся вниз цветков) кистью (табл. 19, 6-9). Цветки розовые, трубчатые, со сросшимися сегментами околоцветника; трубка изогнутая, в верхней половине цилиндрическая, а в нижней - вздутая, расширенная. Так же как у алоэ, тычинки в результате изгибания нитей располагаются по одну сторону от столбика, прилегая к выпуклой стороне околоцветника. Они тесно соприкасаются друг с другом пыльниками, которые полукругом окружают верхушку столбика с рыльцем. И хотя конец столбика немного изогнут в противоположном от тычинок направлении, контакт рыльца со вскрывшимися пыльниками неизбежен. По данным С. Фогеля, у гастерии имеет место и орнитофилия. Предполагают, что опылителями их могут быть и пчелы. Коробочки у гастерий 3-гранные, кожистые, как и у других растений-баллистов, обращены они прямо вверх (рис. 71, 10) вследствие изгибания цветоножек, становящихся при плодах твердыми, проволоковидными. Семена выпадают из коробочек при раскачивании цветоноса ветром. Виды гастерии разводят в суккулентных садах, но чаще их культивируют как горшечные растения.
Эндемичным для Южной Африки является большой род хавортия (Haworthia) (куда в настоящее время включается также хортолирион - Chortolirion), насчитывающий около 200 видов, из которых 1 достигает тропической Африки (Ангола и Зимбабве). Наиболее богата видами Капская область. Представители этого рода - маленькие суккулентные растения с исключительно плотной 3-многорядной, иногда довольно высокой (рис. 71) розеткой листьев и пазушными цветоносами с одной или несколькими рыхлыми кистями. У немногих видов имеется облиственный стебель. Листья хавортий обычно очень толстые и мясистые, нередко твердые и короткие. Поверхность листьев у многих видов шероховатая от многочисленных зеленоватых или беловатых твердых бугорков. По краям листьев иногда имеются зубцы или жесткие реснички. У некоторых видов из пазух нижних листьев развиваются столоны, служащие для вегетативного размножения (рис. 71, 6). Цветки обращены косо вверх. Они мелкие, невзрачные, беловатые, с прямой цилиндрической трубкой, образованной сросшимися до половины сегментами и двугубым отгибом из свободных и отогнутых наружу частей. Верхняя губа образована двумя наружными и одним внутренним сегментами, а нижняя - 2 внутренними и одним наружным. Тычинки (3 короткие и 3 длинные) и расположенный ниже их уровня столбик скрыты в трубке околоцветника. У цветков хавортий очень много нектара, и в Африке их опыляют пчелы. Хавортии часто выращивают в оранжереях, иногда как комнатную культуру.
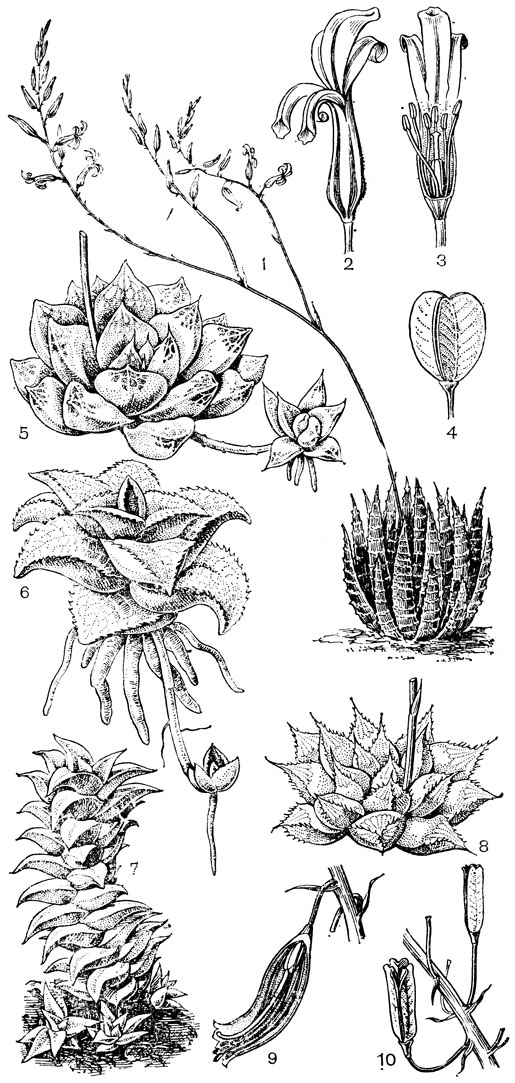
Рис. 71. Асфоделовые. Хавортия полосатая (Haworthia fasciata): 1 - общий вид; 2 - цветок; 3 - цветок в разрезе (3 сегмента околоцветника удалены); 4 - плод. Хавортия ладьевидная (Н. cymbiformis): 5 - розетка взрослого растения со столоном, несущим дочернюю розетку. Хавортия шахматная (Н. tesselata): 6 - розетка с корнями и столоном с дочерней розеткой. Хавортия извилистая (Н. tortuosa): 7 - взрослая и молодые розетки. Хавортия волосконосная (Н. pilifera): 8 - общий вид. Гастерия двурядная (Gasteria disticha): 9 - цветок в разрезе (ориентирован косо вниз); 10 - коробочки (ориентированы вверх)
Следующая триба подсемейства асфоделовых представлена одним родом - книпхофия (Kniphofia). Он объединяет около 60 видов, распространенных в Южной и тропической Африке, на острове Мадагаскар (2 вида) и на Аравийском полуострове (1 вид). Книпхофия растет в горах на высоте 1800-4000 м в избыточно увлажненных местообитаниях: по сырым и болотистым местам среди травянистой растительности, по берегам горных потоков, в зарослях приречных кустарников, иногда в местах, периодически затопляемых в сезон дождей.
Книпхофии - многолетние травы с густой розеткой листьев и центральным цветоносом, который у некоторых видов (например, у тропической книпхофии Томсона - К. thomsonii) достигает в высоту 3 м. Только у книпхофии облиственной (К. foliosa) из Эфиопии имеется облиственный стебель. У некоторых видов (например у книпхофии ягодной - L. uvaria, рис. 70, 2, табл. 20, 3 и книпхофии Томсона) в пазухах самых нижних листьев закладываются почки, из которых развиваются столоны длиной до 15 см, дающие начало новым особям. Листья у книпхофии линейные или линейно-ланцетные, снизу килеватые, сверху плоские или желобчатые, обычно длинные (до 2 м). По строению соцветий и цветков книпхофия похожа на алоэ, но цветонос у нее всегда простой, неразветвленный, с короткой, часто густой кистью. Раскрывание цветков в соцветии обычно происходит снизу вверх, и только у книпхофии карликовой (К. pumila) и книпхофии изоетолистной (К. isoetifolia) оно происходит в обратном направлении. Околоцветник трубчатый или трубчато-воронковидный, иногда колокольчатый, со сросшимися почти доверху или редко до половины своей длины сегментами. Прицветники часто пленчатые, отогнутые вниз и прижатые к цветоносу.
Процесс опыления у книпхофии, по-видимому, сходен с таковым у алоэ. Цветки у них протандричные. Тычинки, скрытые вначале, как и столбик, в трубке околоцветника, при цветении выставляются из него на разную длину (иногда они втрое длиннее околоцветника) и находятся примерно на одном уровне с высовывающимся немного позднее столбиком. В конце мужской фазы нити тычинок у большинства видов книпхофии зигзагообразно извиваются, втягивая пыльники внутрь околоцветника. Цветки книпхофии опыляются птицами. Цветение различных видов происходит в разное время года, но некоторые, как, например, книпхофия Томсона, цветут почти круглый год. Многие виды книпхофии - очень эффектные долго цветущие декоративные растения, нередко разводимые в садах и парках с теплоумеренным и субтропическим климатом. С начала XVIII в. в Западной Европе культивируют интродуцированную из Юго-Восточной Африки книпхофию ягодную с густыми яйцевидными или почти шаровидными кистями огненно-красных бутонов и оранжево-желтых цветков. Этот вид выращивают в СССР на Южном берегу Крыма.
В подсемействе антериковых (Anthericoideae) известны 31 род и около 630 видов; из них 27 родов распространено в Старом Свете, 1 (антерикум) встречается также и в Америке, 4 рода произрастают только на Американском континенте - от юга США (штаты Юта и Аризона) до Перу и Чили. В этом подсемействе различают 5 триб. Самой большой из них является триба антериковых (Anthericeae), состоящая из 14 родов, представленных многолетними травами и очень редко небольшими кустарниковидными и полукустарниковидными растениями. Цветки у них звездчатые, колесовидные, редко воронковидные или колокольчатые, со свободными или сросшимися при основании, редко до половины сегментами. Тычинок иногда 3 (анемаррена - Anemarrhena). Гнезда завязи обычно с многочисленными (до 30), реже несколькими или 2 (роды анемаррена и алекторурус - Alectorurus) семязачатками. Плод - коробочка.
Наиболее известным родом трибы антериковых является хлорофитум (Chlorophytum). Он включает около 220 видов, большинство которых распространено в тропической Африке, 20 видов встречаются в Южной Африке, около 15 - на Мадагаскаре, по 1 - на юге Аравийского полуострова и на острове Сокотра, около 15 видов - в Индии, несколько видов - в Таиланде, Индокитае, на полуострове Малакка, в Южном Китае, Австралии и Тасмании. Виды хлорофитума преимущественно мезофильные растения, обитающие во влажных, обычно тенистых местах. На востоке Южной Африки в субтропических вечнозеленых муссонных лесах обычен хлорофитум хохолковый (С. comosum), являющийся важным компонентом их травяного покрова. Это растение имеет довольно необычный облик, благодаря тому что в его соцветии на верхушке цветоноса и боковых ветвей, а нередко и на других частях развиваются новые растения с розетками листьев и короткими корнями. При полегании цветоноса розетки укореняются, и таким путем у этого вида происходит быстрое вегетативное размножение. Хлорофитум хохолковый хорошо знаком читателям, так как он широко распространен в комнатной культуре. Некоторые виды хлорофитума произрастают в аридных районах Южной Африки. В качестве примера можно привести хлорофитум липкий (С. viscosum) - очень жесткое, опушенное железистыми волосками растение, обитающее по пескам в пустыне Намиб и на полупустынном плоскогорье Намакваленд, а также хлорофитум аридный (С. aridum), встречающийся в кустарниковых саваннах Трансвааля.
Виды хлорофитума - многолетние травы с ползучим корневищем, обычно тонкими, но у концов часто клубневидно утолщенными корнями, с розетками листьев и с центральным простым или разветвленным цветоносом. Самый высокий цветонос (до 2 м), а также наиболее длинные (до 140 см) слабосуккулентные листья имеет похожий на алоэ хлорофитум Крука (С. krookianum), встречающийся по сырым местам в восточных районах Южной Африки (рис. 72). Листья у хлорофитумов от линейных до широколанцетных, по краю обычно бахромчатые, а при основании нередко суженные в черешок. Мелкие, белые, реже зеленоватые или розоватые цветки собраны в 1-6-цветковые пучки, расположенные в пазухе прицветников и образующие рыхлые или более или менее густые сложные кисти. Каждый цветок такого пучка сидит непосредственно на оси соцветия в пазухе прицветника. Пучки представляют собой редуцированные боковые ветви соцветия, оси которых подверглись полной редукции. У некоторых видов (например, у хлорофитума капского - С. capense) иногда развиваются очень короткие оси и цветки на них расположены по спирали. Околоцветник у хлорофитумов со свободными сегментами, обычно расположенными звездчато или отклоненными вниз. Тычинки с крупными пыльниками, прикрепляющимися к нити основанием, при увядании изгибаются назад. Гинецей состоит из 3 плодолистиков, но у хлорофитума клубневого (С. tuberosum), распространенного в тропической Африке и Индии, он образован только 2 плодолистиками. В каждом гнезде завязи имеется от 6 до 30 семязачатков. Коробочка 3-гранная, часто глубоколопастная. Семена крупные, диаметром 2-4 мм, плоские, округлые, черные и блестящие.
Рис. 72. Асфоделовые. Хлорофитум Крука (Chlorophytum krookianum): 1 - надземная часть растения; 2 - часть соцветия. Хлорофитум клубневой (С. tuberosum): 3 - общий вид растения; 4 - цветок; 5 - плод. Эхеандия крупноплодная (Echeandia macrocarpa): 6 - цветок со шмелем, собирающим пыльцу и производящим перекрестное опыление. Алекторурус йедоенский (Alectorurus yedoensis): 7 - семя с пучком волосков при основании. Анемаррена асфоделовидная (Anemarrhena asphodeloides): 8 - цветок
Цветет хлорофитум в течение всего года, но разные его виды - в различное время. Цветки в пучках раскрываются последовательно, и первым распускается нижний бутон. При неблагоприятных для цветения условиях рост бутонов может задержаться на продолжительное время, и если они вновь начнут расти и распускаться, то в одних и тех же пучках можно видеть и зрелые плоды, и раскрытые и увядшие цветки, и бутоны. Цветоножки при цветении направлены косо вверх и не изменяют своего положения при плодах. Цветки опыляются пчелами, цветочными мухами и другими насекомыми, привлекаемыми пыльцой и нектаром. Несколько видов хлорофитума разводят как декоративные в комнатах и оранжереях. Из них наиболее распространен хлорофитум хохолковый, имеющий ряд культурных разновидностей, в том числе пестролистную форму с продольными белыми полосами, а также хлорофитум капский.
К роду хлорофитум очень близок род антерикум (Anthericum), отличающийся от него главным образом мелкими, шаровидными в очертании, угловатыми, черными семенами. В род антерикум входит около 250 видов, распространенных преимущественно в тропической Африке. Из них 17 видов произрастают в Южной Африке, 2 - в Северной Африке, 3 - в Европе, несколько - на юге Северной Америки, в Центральной и Южной Америке. Два европейских вида - антерикум ветвистый (А. rаmosum) и антерикум обыкновенный (A. liliago, рис. 73) - достигают в своем распространении юга Швеции и, следовательно, дальше других представителей семейства асфоделовых проникают в северные широты. Антерикум ветвистый встречается и в южных, и отчасти центральных районах европейской части СССР. Оба вида обитают по сухим солнечным местам, травянистым и каменистым склонам гор и холмов, сухим разреженным лесам и кустарникам. Европейские виды имеют простые кисти, в то время как у большинства остальных видов они сложные, образованные не одиночными цветками, а пучками цветков. Африканские виды антерикума обитают в условиях, в основном сходных с таковыми у видов хлорофитума. Многие'тропические виды растут в горах, по открытым травянистым и лесным склонам, скалам, берегам горных рек. В Южной Африке представители этого рода чаще всего встречаются в горных и равнинных кустарниковых песчаных саваннах. У антерикума обыкновенного пыльники вскрываются одновременно с раскрыванием цветков или сразу после этого. Самоопыление устраняется благодаря разной длине тычинок и столбика, который превышает тычинки примерно на 5 мм и, кроме того, немного отогнут вниз.
На верхней поверхности завязи из септальных нектарников крупными каплями выделяется нектар (рис. 73, 2). Антерикум разводят как декоративное растение в открытом грунте.
Рис. 73. Асфоделовые. Антерикум обыкновенный (Anthericum liliago): 1 - общий вид; 2 - гинецей (на верхушке завязи видны капельки нектара, выделившиеся из септальных нектарников). Артроподиум усиконосный (Arthropodium cirrhatum): 3 - соцветие (цветоножки в верхней половине белые, похожие на трубку околоцветника); 4 - лист; 5 - тычинка (от пыльника отходят два волосистых придатка). Диурантера большая (Diuranthera major): 6 - верхняя часть соцветия с бутонами, цветками и плодами; 7 - лист; 8 - тычинка (на пыльнике два придатка)
Следует упомянуть об эндемичном для Африки роде трахиандра (Trachyandra), насчитывающем 50 видов, из которых 45 встречаются в Южной Африке, преимущественно в Капской области, а остальные - в тропической Африке. Среди видов этого рода имеется несколько кустарниковидных форм, очень редких в подсемействе антериковых. В качестве примера назовем встречающуюся на юго-западе Капской области трахиандру Адамсона (Т. adamsonii) - кустарник высотой до 180 см, с ветвями, покрытыми твердыми основаниями опавших листьев и несущими на верхушках пучки узких, сизых, слабосуккулентных листьев. Оригинально выглядят некоторые травянистые представители рода, например трахиандра рыхлая (Т. lаха), обитающая в пустыне Калахари и других засушливых районах Южной Африки. После образования плодов веточки его сильно ветвистого раскидистого соцветия, а также цветоножки дуговидно загибаются книзу, и тогда все соцветие приобретает шаровидную форму. Отрываясь от растения, эти сухие шаровидные соцветия (перекати-поле) перекатываются ветром по земле, разбрасывая семена.
Три монотипных рода из трибы антериковых - анемаррена, тераухия (Terauchia) и алекторурус - представляют семейство асфоделовых в Восточной Азии. Анемаррена асфоделовидная (Anemarrhena asphodeloides) растет по сухим склонам гор в лесостепных и степных районах Северо-Восточного и Северного Китая и на юге полуострова Корея, образуя иногда своеобразные анемарреновые степи. Она имеет горизонтальное ветвящееся корневище, на конце которого развивается густой пучок злаковидных желобчатых листьев и цветонос длиной до 1 м с негустой кистью мелких невзрачных цветков со слабым сладковатым запахом. Сегменты околоцветника, сросшиеся до половины своей длины, образуют тускло-пурпуровую трубку и желтый или фиолетовый отгиб. Цветки имеют только 3 тычинки, почти полностью сросшиеся нитями с внутренними сегментами околоцветника (рис. 72, 8). Тычинки скрыты в трубке околоцветника и расположены выше столбика. Анемаррена цветет в середине лета. Цветки ее раскрываются после 7 ч вечера и опыляются мелкими ночными бабочками из группы совок (Acronicta psi) и огневок (Pyrausta gracilis).
К анемаррене очень близка тераухия анемарренолистная (Terauchia anemarrhenaef olia), встречающаяся на севере полуострова Корея. У этого вида известны только мужские цветки с недоразвитой завязью. Единственный вид алекторуруса - алекторурус йедоенский (Аlеctorurus yedoensis) изредка встречается в Японии, на островах Сикоку и Кюсю, в горах на влажных скалах. Это небольшое двудомное растение с розеткой двурядных вечнозеленых широколинейных листьев и цветоносом высотой 15-40 см, с рыхлым метельчатым соцветием из мелких белых колокольчатых цветков. Семена его снабжены при основании длинными белыми волосками (рис. 72, 7). Эта особенность свойственна только данному роду. Алекторурус занимает в трибе антериковых обособленное положение.
В горах Средней и Южной Европы распространен род парадизея (Paradisea), названный по имени итальянского ботаника конца XVIII и начала XIX в. Д. Парадизи. В род входят 2 вида, из которых более известна парадизея лилиевидная (P. liliastrum), растущая на альпийских и субальпийских лугах в Пиренеях, Юре, Альпах и Апеннинах (рис. 64,1). Этот вид имеет самые крупные в подсемействе цветки (длиной 3-5 см). Они белые, воронковидные, со свободными сегментами, собранные в 2-20-цветковую более или менее одностороннюю кисть. Их 3 длинные и 3 короткие тычинки с качающимися пыльниками дуговидно изогнуты наверх так же, как и немного превышающий их столбик. Парадизею лилиевидную разводят как декоративное растение, в особенности на альпийских горках.
В Новом Свете обитают 3 эндемичных рода из трибы антериковых. Из них 2 - эремокринум (Eremocrinum), распространенный на юге США (штаты Аризона и Юта), и чилийский род боттионея (Bottionea) - являются монотипными. Третий, исключительно тропический род эхеандия (Echeandia), объединяет 13 видов, распространенных в Центральной Америке (от Мексики до Панамы) и на севере Южной Америки (Венесуэла и Эквадор). Виды эхеандии - многолетние травы с утолщенными от основания корнями, длинными линейными листьями и собранными в пучки цветками, образующими на верхушке цветоноса и его боковых ветвей рыхлые кистевидные соцветия. Пыльники у эхеандии срослись в довольно длинную коническую трубку, окружающую столбик, что является приспособлением к перекрестному опылению. Срастание пыльников - характерная особенность этого рода. Австралийским ботаником П. Бернхардтом и американским Э. Монталво (1977, 1979) детально изучена биология цветения эхеандии крупноплодной (Е. macrocarpa), произрастающей в горах Центральной Америки в дубово-можжевеловых лесах на высоте до 1800 м над уровнем моря. Как отмечают названные авторы, эхеандии в строении андроцея обнаруживают замечательное сходство с двудольными растениями умеренного пояса - додекатеоном обыкновенным (Dodecatheon meadia) и пасленом сладко-горьким (Solanum dulcamara), у которых пыльники также образуют конус вокруг столбика, хотя и не срастаются при этом между собой. Указанное сходство обусловливает и одинаковый механизм опыления у этих растений. Цветки эхеандии крупноплодной желтые, с колесовидным околоцветником и во время цветения обращены косо вниз (рис. 72, 6). Они строго протогиничны. В женскую фазу, которая продолжается один день, из пыльникового конуса высовывается столбик. И только после того, как рыльце потеряет способность воспринимать пыльцу и столбик втянется в пыльниковый конус, происходит вскрывание пыльников. Поэтому самоопыление у эхеандии совершенно исключено. Мужская фаза продолжается от 2 до 7 суток в зависимости от погодных условий. Следовательно, каждый отдельный цветок большую часть жизни функционирует как донор пыльцы, а не как ее получатель. Цветки эхеандии не имеют нектара, и насекомые посещают их ради пыльцы. Главными опылителями, в большом количестве встречающиеся на цветках эхеандии, являются шмели - рабочие особи и царицы. Подлетая к цветку, рабочий шмель обхватывает пыльниковый конус передними парами ног и мандибулами. При вибрации грудных крыльев пыльниковый конус сотрясается и пыльца высыпается из его внутренних продольных щелей на голову, грудь и брюшко насекомого, которое стряхивает ее затем второй парой ног в пыльцевые мешки, находящиеся на задних ногах. Шмели-царицы ведут себя аналогично рабочим особям с той лишь разницей, что они иногда прекращают вибрацию и слизывают выпавшую из конуса пыльцу длинным язычком. Каждую минуту насекомые посещают в среднем 14 цветков, причем 24 с насекомое тратит на полет, а 36 с - на получение пыльцы путем вибрации. Несмотря на устойчивые взаимоотношения с насекомыми-опылителями, в соцветиях эхеандии крупноплодной образуется мало плодов (в среднем 8 плодов на соцветие, в котором до 66 бутонов). Такая малая продуктивность может быть объяснена двумя причинами. Во-первых, только 15-20% цветков в популяции находятся одновременно в женской фазе. Во-вторых, из 4-5 цветков пазушного пучка только один (нижний) способен после опыления образовать плод, остальные же цветки пучка (их называют вспомогательными) являются функционально пыльниковыми. Они хотя и имеют внешне нормально развитый гинецей, но оказываются не в состоянии завязать плоды. Наличие вспомогательных цветков и большую продолжительность мужской фазы цветка по сравнению с женской Бернхардт и Монталво считают приспособлением, выработанным растениями, чтобы обеспечить насекомых обильной пищей - пыльцой - и тем самым постоянно привлекать их к себе и опылять цветки, от которых они ничего не получают (в женской фазе пыльники закрыты). Описанная "стратегия" опыления, как полагают, является выгодной для растения, так как, с одной стороны, она надежно обеспечивает перекрестное опыление, а с другой - благодаря образованию небольшого количества плодов позволяет растению меньше истощать энергетические резервы и полнее сохранить их для следующей вегетации. Эхеандия крупноплодная цветет с начала июля, в сезон дождей, при температуре 14-15 °С. Вскрывание плодов у нее приурочено к периоду сильных ветров (конец октября - начало ноября), что способствует более успешному разбрасыванию семян из обращенных вверх коробочек этого растения баллиста.
Триба тизанотовых (Thysanoteae) состоит из одного рода тизанотус. Он включает 37 видов, распространенных в Австралии, из них 2 вида встречаются в Новой Гвинее, а 1 - тизанотус китайский (Thysanotus chinensis) - в Южном Китае, Таиланде, Вьетнаме, на Филиппинах и в Индонезии. Большинство представителей рода растет в умеренном и субтропическом поясах, часто встречаясь на песчаных равнинах, покрытых верещатниками, на приморских песках, по сухим холмам. Большую экологическую амплитуду имеет распространенный почти по всему ареалу рода тизанотус клубневой (Т. tuberosus), встречающийся от пустынных районов Центральной Австралии, где растет в сообществах ксерофитных злаков, до тропических районов Квинсленда (рис. 74, 1). Все тизанотусы - многолетние травы с коротким корневищем, от которого отходит пучок многочисленных тонких или (чаще) клубневидно утолщенных корней. Листья прикорневые, линейные, злаковидные, иногда полуцилиндрические или несколько мясистые. У немногих представителей (тизанотуса китайского и тизанотуса многоцветкового - Т. multiflorus) они вечнозеленые; у некоторых других видов (например, у тизанотуса колюченосного - Т. spiniger) листья отмирают еще до начала цветения и ассимиляционная функция переходит к цветоносу. У тизанотуса колючего большинство веточек соцветия заканчивается колючим острием, и только немногие из них 1-, 2-цветковым частным соцветием (рис. 74, 4). У единственной в роде лианы - тизанотуса Патерсона (Т. patersonii) - цветонос извилистый, вьющийся, длиной до 1 м, с короткими несущими пучки цветков боковыми ветвями (рис. 74, 3). Своеобразное строение имеют цветки тизанотусов: внутренние сегменты распростертого околоцветника резко отличаются от наружных - они значительно шире благодаря перепончатым, часто фиолетовым и обычно бахромчато-реснитчатым краям (рис. 74, 2). Тычинки, как правило, неравные: 3 внутренние имеют более длинные нити и особенно пыльники; у некоторых видов внутренние тычинки совсем не развиваются. Пыльники линейные, длиной до 1 см, прикрепляются к нити основанием, реже спинкой и вскрываются верхушечной порой. При увядании они обычно спирально скручиваются. В каждом гнезде завязи 2 семязачатка, но иногда их 10-17. При основании семян имеется маленький придаток - элайосома. Род тизанотус благодаря особенностям строения цветков, а также наличию в хлоренхиме листьев и стеблей рафидных каналов с оксалатом кальция, которых нет у других представителей порядка лилейных, занимает в подсемействе антериковых довольно обособленное положение.
В трибе ходжсониоловых (Hodgsonioleae) различают 7 родов, их представители обитают в южном полушарии. Монотипный род паситея (Pasithea) встречается в Южной Америке (юг Перу, Чили), 5 моно- и олиготипных родов - в Австралии, Тасмании и на востоке Новой Гвинеи и 1 род цесия (Caesia, 12 видов) - на юге Африки, Мадагаскаре и в Австралии. Все представители этой трибы - многолетние травы с прикорневыми, иногда двурядными (виды цесии) листьями и безлистными цветоносами или редко (роды трикорина и коринотека) с облиственными ветвистыми стеблями. Листья обычно линейные, злаковидные, иногда шиловидные, при основании нередко с влагалищами. Цветки мелкие, обычно голубые, редко белые или желтые, собранные в метелку, простую или сложную кисть или в малоцветковые зонтиковидные верхушечные соцветия (например, у трикорины). Околоцветник звездчатый, со свободными, редко сросшимися внизу в короткую трубку сегментами. При отцветании околоцветник спирально скручивается (рис. 74, 8), что является характерным признаком этой трибы, который свойствен еще только одному австралийскому роду - арнокринуму (Arnocrinum) из трибы джонсониевых. Скрученный околоцвет-ник более или менее долго сохраняется при развивающемся плоде, но затем опадает. Тычинок обычно 6, пыльники прикрепляются спинкой и вскрываются интрорзно, а у рода агростокринум (Agrostocrinum) - верхушечной порой. У рода ходжсониола (Hodgsoniola) имеются 3 фертильные и 3 стерильные тычинки. У первых связники удлинены и срослись в цилиндрическую трубку, из которой выступает столбик. У рода трикорина нити тычинок очень густо опушены обращенными вверх воло-сками (рис. 74, 7). Число семязачатков в каждом гнезде завязи варьирует от многочисленных (у родов паситея и хамесцилла - Chamaescillа) до 2 у остальных ее представителей. Роды трикорина и коринотека имеют необычные для семейства асфоделовых плоды. У трикорины в каждом гнезде ее глубоко трехлопастной завязи из 2 семязачатков развивается 1 и, кроме того, 1 или 2 гнезда завязи часто дегенерируют.
Рис. 74. Асфоделовые. Тизанотус клубневой (Thysanotus tuberosus): 1 - общий вид; 2 - цветок с цветоножкой. Тизанотус Патерсона (Т. patersonii): 3 - общий вид. Тизанотус колюченосный (Т. spiniger): 4 - часть соцветия. Tрикорина высокая (Tricoryne elatior): 5 - общий вид; 6 - цветок; 7 - тычинка; 8 - скрученный (после отцветания) околоцветник; 9 - плод
Образующийся плод состоит из 3-1 односемянных невскрывающихся сухих или сочных долей (рис. 74, 9). Односемянным невскрывающимся часто бывает плод у растений из рода коринотека, когда в их завязи развивается только одно гнездо с 1 семязачатком. Но если развивается больше гнезд и семян, то тогда образуется довольно поздно вскрывающийся плод-коробочка. Таким образом на примере родов трикорина и коринотека можно наглядно видеть возможные пути трансформации коробочки в односемянный невскрывающийся плод. Семена цесии и коринотеки снабжены при основании элайосомами; у цесии элайосомы являются строфиолями, у коринотеки - карункулами.
С представителями рассмотренной трибы обнаруживает родство по эмбриологическим признакам род симетис (Simethis), выделенный X. Хубером (1969) в трибу симетисовых (Simethideae). Единственный вид рода - симетис плосколистный (S. planifolia) произрастает в Западном Средиземноморье (Алжир, Марокко Тунис, юг Европы до Италии), на западе Франции и юго-западе Ирландии, встречаясь по верещатникам, каменистым местам, разреженным сосновым лесам. Это многолетнее растение с мясистыми шнуровидными корнями, пучком длинных прикорневых влагалищных листьев и цветоносом длиной до 40 см (рис. 67, 9, 10). Соцветие у симетиса рыхлое, метельчатое, с немногими ветвями, несущими пучки из 3-7 цветков. Околоцветник звездчатый, со свободными сегментами, сверху белыми, снизу пурпурными. Тычинки с длинными пыльниками и густо шерстисто-волосистыми нитями, прикрепляющиеся к ямке связника. В каждом гнезде завязи образуются по 2 семязачатка. Плод - шаровидная коробочка. Семена при основании с белым придатком - строфиолой.
Очень своеобразна эндемичная для Австралии триба джонсониевых, иногда выделяемая в отдельное семейство. В нее входит 8 родов, включающих многолетние травы, реже кустарничковидные формы. Многие представители трибы являются ксерофильными растениями. Характерным признаком джонсониевых является наличие головчатого, реже зонтиковидного соцветия, окруженного оберткой кроющих листьев; иногда (у рода джонсония - Johnsonia) соцветие-простой колос. Другая особенность джонсониевых - унифациальные (билатеральные или 3-гранные) листовые пластинки. Листья джонсониевых расположены в прикорневых или верхушечных пучках или по всему стеблю. Цветки сидят в пазухе прицветников, причем у некоторых родов каждый цветок имеет еще и прицветничек, расположенный на его вентральной (брюшной) стороне и в разной степени охватывающий цветок (рис. 75). Сегменты
околоцветника свободные или сросшиеся в основании или наполовину своей длины, образуя в последнем случае трубку и отгиб (например, у рода бория - Воrуа). В цветке 6 или 3 тычинки; в каждом гнезде завязи от многих до нескольких семязачатков.
В роде джонсония 3 вида, обитающих в Юго-Западной Австралии по сухим приморским пескам. Наиболее известна джонсония обыкновенная (J. lupulina) -плотнодерновинный многолетник с жесткими двурядно расположенными билатеральными узколинейными листьями и уплощенным стеблем (рис. 75, 1). Джонсонию можно принять за растение из семейства злаков. Злаковидный облик имеют не только дерновины этого вида, но и его соцветие - густой продолговато-эллиптический колос с крупными скрывающими цветки, тонкокожистыми, как у злаков, чешуями. При основании колоса имеется кроющий лист, направленный вертикально вверх, благодаря чему создается впечатление, что он является продолжением стебля, а соцветие занимает не верхушечное, а боковое положение. Такое расположение нижнего прицветного листа свойственно также некоторым представителям семейств ситниковых и осоковых. Цветки у джонсонии мелкие, со сросшимися внизу и образующими короткую трубку сегментами околоцветника. Тычинок 3, расширенные основания их нитей срослись между собой и с околоцветником (рис. 75, 3). Столбик расположен выше уровня пыльников. Семена джонсо-нии снабжены элайосомами.
Другой род трибы джонсониевых - совербея (Sowerbaea) состоит из 5 близкородственных видов, дизъюнктивно распространенных по всему побережью Австралии и Тасмании. Растут они преимущественно на сырых и болотистых приморских песках и в субприбрежных верещатниках. Это небольшие, образующие дерновины многолетние травы с прикорневыми листьями, имеющими узкую 3-гранную пластинку и влагалище, а также длинный язычок, расположенный, как и у злаков, на границе между ними (рис. 75, 8-9). По внешнему виду совербея походит на лук. Ее кувшинчатые фиолетовые, розоватые или синие цветки со свободными сегментами околоцветника собраны в довольно густое многоцветковое зонтиковидно-головчатое (как у луков) соцветие, окруженное общей оберткой из нескольких чешуевидных листочков. Каждый цветок располагается в пазухе сильно бахромчато рассеченного прицветника. Для рода характерно наличие 3 тычинок, супротивных внутренним сегментам околоцветника, и 3 чередующихся с ними стаминодиев. Тычинки с очень короткими нитями и удлиненными раздвоенными пыльниками. Столбик расположен значительно выше тычинок, и поэтому самоопыления у видов этого рода, по-видимому, не происходит. В каждом гнезде завязи 2-6 семязачатков.
Род бория (Вогуа, рис. 75, 5-7) включает низкие, сильно ксерофилизованные кустарничковидные растения, с очень жесткими колю-чими 3-граннымй листовыми пластинками и широкими короткими влагалищами. Плотные головчатые соцветия окружены черепитчатой оберткой из жестких кожистых чешуй. Супротивно кожистым прицветникам, в пазухах которых сидят цветки, на вентральной стороне последних располагаются чешуевидные прицветнички, охватывающие своими завернутыми краями нижнюю часть цветка (рис. 75, 6). Цветки мелкие, желтоватые, с околоцветником, дифференцированным на длинную трубку из сросшихся частей сегментов и колесовидный отгиб. Тычинок 6, прикрепленных тонкими нитями к зеву околоцветника, что является необычным для этого семейства. Столбик расположен на одном уровне с пыльниками. Семязачатки в каждом гнезде завязи многочисленные. В род бория входят 3 вида, встречающихся в Западной Австралии и Квинсленде. Бория блестящая (В. nitida) замечательна тем, что может сохранять жизнеспособность даже после сильного пересыхания. Она обитает в аридных районах юго-запада Австралии. Это низкий (высотой 3-15 см) кустарничек с очень жесткими трехгранными игловидными листьями, скученными на верхушке побегов. Пластинки листьев со временем опадают, а их короткие расширенные черепитчато налегающие друг на друга основания остаются на ветвях, придавая им своеобразный чешуйчатый вид. Листья бории блестящей имеют ярко выраженное ксероморфное строение: массивное кольцо склеренхимы под эпидермой, очень глубокое расположение устьиц. При наступлении летней засухи этому виду приходится существовать в течение многих недель в отсутствии влаги. Что же происходит в это время с растением? Его листовые пластинки, прежде расположенные более или менее горизонтально, поднимаются вверх, образуя плотный жесткий пучок, в котором молодые листья и верхушечная почка оказываются под прикрытием старых листьев. Продольные бороздки листа с находящимися в них устьицами закрываются благодаря сцеплению зубцевидных выростов на эпидермальных клетках. Эти приспособления несколько замедляют, но не приостанавливают процесс потери воды растением. Продолжая все больше и больше обезвоживаться, листья становятся вначале ярко- оранжевыми, а затем соломенно-желтыми. Такие растения производят впечатление совершенно засохших, безжизненных. Но при наступлении периода дождей они вновь становятся зелеными. Специальные наблюдения в природе показали, что если засохшие растения, находившиеся без воды в течение нескольких недель, полить, то через 24 ч после полива листья возвращаются в прежнее, почти горизонтальное положение и начинают зеленеть от основания. Через 16 суток после полива листья становятся зелеными больше чем наполовину своей длины, и на этом процесс их позеленения обычно останавливается. Экспериментально установлено, что бория блестящая может обходиться без воды в течение четырех лет!
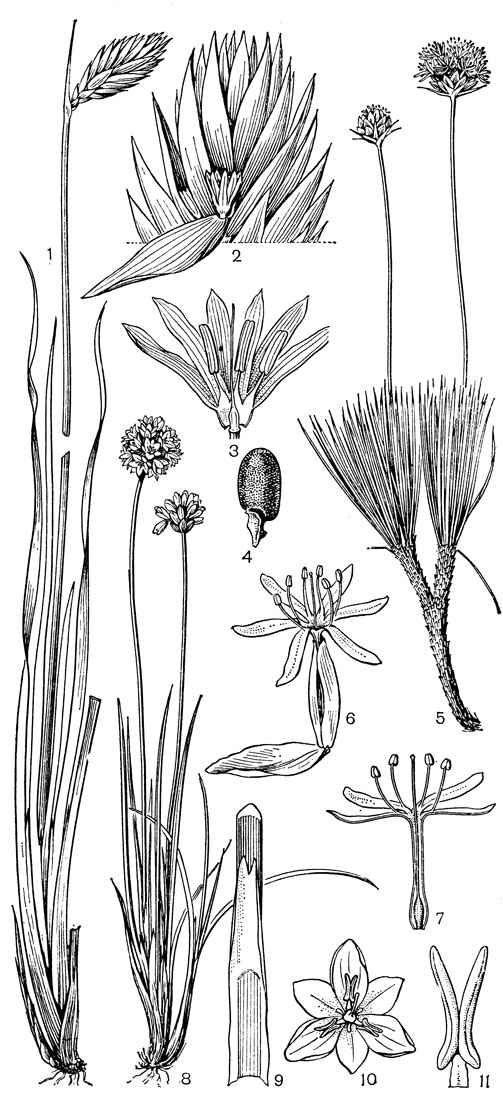
Рис. 75. Асфоделовые. Джонсония обыкновенная (Johnsonia lupulina): 1 - общий вид; 2 - верхняя часть соцветия с отогнутым прицветником (виден сидящий в его пазухе цветок с прицветничком); 3 - цветок в разрезе; 4 - семя с элайосомой. Бория северная (Borya septentrionalis): 5 - общий вид; 6 - цветок с отогнутым прицветником и окружающим трубку околоцветника прицветничком; 7 - продольный разрез цветка. Совербея ситниковая (Sowerbaea juncea): 8 - общий вид; 9 - часть листа (на границе влагалища и пластинки расположен длинный язычок); 10 - цветок; 11 - тычинка
Семейство ксанторреевые (Xanthorrhoeaceae)
К наиболее замечательным по внешнему облику растениям Австралийского флористического царства принадлежат многие представители небольшого семейства ксанторреевых. Из 9 родов и 65-70 видов этого семейства лишь 2 вида самого богатого видами (их около 42) рода ломандра (Lomandra) заходят из Восточной Австралии на юг Новой Гвинеи и в Новую Каледонию, а недавно установленный род ромналъда (Romnalda) с единственным видом ромналъда папуасская (R. papuana) эндемичен для Новой Гвинеи. Все остальные роды и виды ксанторреевых встречаются только в Австралии и Тасмании. Особенно большое ландшафтное значение имеют древовидные представители ксанторреевых, называемые "травяными" или "злаковыми" деревьями ("Grass-Trees") из-за большого сходства их длинных и узких листьев с листьями злаков. Это кингия австралийская (Kingia australis, рис. 76, 9) и многие виды рода ксанторрея (Xanthorrhoea, рис. 77), содержащего около 15 видов. Хотя семейство ксанторреевые было установлено известным бельгийским ботаником Б. Ш. Ж. Дюмортье еще в 1829 г., в течение долгого времени принадлежащие к нему роды относили или к семейству ситниковых (Дж. Вентам и Дж. Д. Хукер, 1862- 1883), или к семейству лилейных в очень широком его объеме (А. Энглер, 1887 и многие другие авторы). Хотя по строению цветков (они ветроопыляемые, с сухим, обычно перепончатым или кожисто-перепончатым околоцветником) ксанторреевые действительно на-поминают ситниковые, они все же значительно более тесно сближаются с семейством асфоделовых, а через него и с лилейными в современном более узком понимании этого семейства.
Роды ксанторреевых очень разнообразны как в отношении жизненной формы, так и по строению цветков. Не случайно многие из них выделяются некоторыми авторами в самостоятельные монотипные или олиготипные семейства кингиевых (Kingiaceae), бакстериевых (Вахteriaceae), ломандровых (Lomandraceae), дазипогоновых (Dasypogonaceae) и калектазиевых (Calectasiaceae). Кроме уже упомянутых древовидных форм, в семействе ксанторреевых представлены как многолетние дерновинные или длиннокорневищные травы, так и сильно разветвленные полукустарники, в том числе не менее оригинальная эрикоидная (верескоподобная) калектазия синецветковая (Calectasia суаnеа) с очень мелкими, хвоеподобными листьями (рис. 78). Общим для всех ксанторреевых является ксероморфное строение их вегетативных органов и особенно листьев, всегда очень узких (редко достигающих в ширину 10-12 мм) и жестких, с мощными тяжами склеренхимы, а также с относительно толстостенными клетками многих других тканей, в том числе даже клеток палисадной паренхимы. В отличие от ситниковых проводящая система стеблей и листьев большинства ксанторреевых состоит из трахеид и только в корнях имеются довольно примитивные по строению сосуды с простой, реже (у кингии и бакстерии - Baxteria) лестничной перфорацией. Лишь у некоторых видов ксанторреи и акантокарпуса (Acanthocarpus) сосуды с лестничной перфорацией наблюдались и в листьях.
Из древовидных ксанторреевых наибольшей высоты (до 9 м) достигает кингия австралийская - единственный вид эндемичного для Юго-Западной Австралии рода. Разрастание ее стебля в толщину - результат деятельности камбиеподобной меристемы. Толстый неразветвленный ствол взрослый особей кингии заканчивается розеткой многочисленных густорасположенных узколинейных листьев длиной до 1 м и обычно несколькими шаровидными соцветиями на довольно длинных и толстых цветоносах, густо покрытых чешуевидными верховыми листьями (рис. 76, 9, 10). В вегетативном состоянии с кингией сходны и древовидные виды ксанторреи, также имеющие толстый ствол-каудекс, подобно стволам саговников покрытый остатками оснований листьев. Однако ствол ксанторреи обычно заканчивается одним густым цилиндрическим соцветием, напоминающим початок рогозов и расположенным на длинной, лишенной листьев ножке. У наиболее крупного вида этого рода - ксанторреи австралийской (Xanthorrhoea australis) - ствол может достигать в высоту 6-7 м и канделябровидно ветвиться в верхней части, образуя несколько листовых розеток с соцветиями или без них. У других древовидных ксанторрей (ксанторреи четырехгранной - X. quadrangulata рис. 77, ксанторреи Прейса - X. preissii и ксанторреи древовидной - X. arborea) стволы-каудексы не превышают в высоту 2-3 м и обычно не ветвятся. Некоторые другие виды, примером которых может служить наиболее известный вид рода - ксанторрея смолистая (X. resinosa, рис. 77, 5), имеют лишь немного возвышающийся над поверхностью земли каудекс (очень редко он достигает в высоту 1-1,5 м). У широко распространенной в Южной Австралии и на Тасмании ксанторреи малой (X. minor, рис. 77, 8) каудекс уже полностью скрыт в земле, а ее побеги образуют более или менее густые дерновины с соцветиями высотой до 80 см. Еще мельче ксанторрея карликовая (X. pumilio) из Квинсленда, соцветия которой возвышаются над поверхностью земли всего на 20-40 см.

Рис. 77. Ксанторреевые. Ксанторрея четырехгранная (Xanthorrhoea quadrangulata): 1 - общий вид; 2 - лист на поперечном сечении; 3 - гинецей; 4 - раскрывшийся плод. Ксанторрея смолистая (X. resinosa): 5 - общий вид; 6 - гинецей; 7 - он же на поперечном сечении. Ксанторрея малая (X. minor): 8 - общий вид; 9 - верхняя часть соцветия; 10 - лист на поперечном сечении; 11 - цветок с прицветничками
Все виды ксанторреи имеют интересную особенность, вероятно являющуюся средством защиты от перегрева или механических повреждений: их каудексы, основания листьев, а иногда и соцветия выделяют большое количество клейкой смолы, слой которой на стволах древовидных видов может достигать в толщину 2-4 см. У большинства видов она имеет коричневую или темно-красную окраску, но у ксанторреи смолистой, ставшей известной европейцам раньше других видов рода, смола желтая, откуда и происходит название рода Xanthorrhoea, которое может быть переведено на русский язык как "желтосмолка".
Виды самого крупного рода семейства - ломандры (рис. 76) - сходны по жизненной форме с травянистыми видами ксанторреи, но нередко имеют ползучие корневища. У видов секции схеноломандра (Schoenolomandra) как удлиненные вегетативные, так и более короткие репродуктивные побеги несут лишь видоизмененные в чешуи листья. Жизненную форму розеткообразующего многолетнего растения имеют также немногие виды хамексероса (Chamaexeros), ромнальда папуасская и бакстерия австралийская (Baxteria australis). Последний вид, в отличие от всех других представителей семейства, образует довольно крупные одиночные цветки на коротких ножках, несущих чешуевидные листья. Более расставленные листья на побегах свойственны видам дазипогона (Dasypogon, рис. 78, 2), а акантокарпус Прейса (Асаnthocarpus preissii, рис. 76) из Юго-Западной Австралии и калектазия синецветковая имеют жизненную форму полукустарника с сильно разветвленными и густо облиственными стеблями.
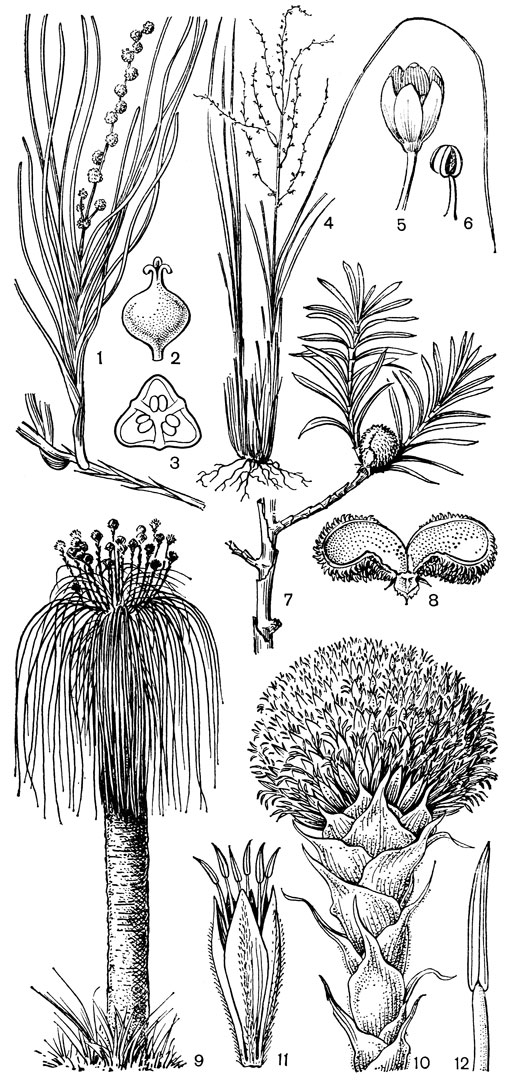
Рис. 76. Ксанторреевые. Ломандра сизая (Lomandra glauca): l - общий вид; 2 - гинецей; 3 - он же на поперечном сечении. Ломандра мелкоцветковая (L. micrantha): 4 - общий вид; 5 - мужской цветок; 6 - тычинка. Акантокарпус Прейсa (Acanthocarpus preissii): 7 - ветвь с плодами; 8 - вскрывшийся плод. Кингия австралийская (Kingia australis): 9 - общий вид; 10 - соцветие; 11 - цветок; 12 - тычинка
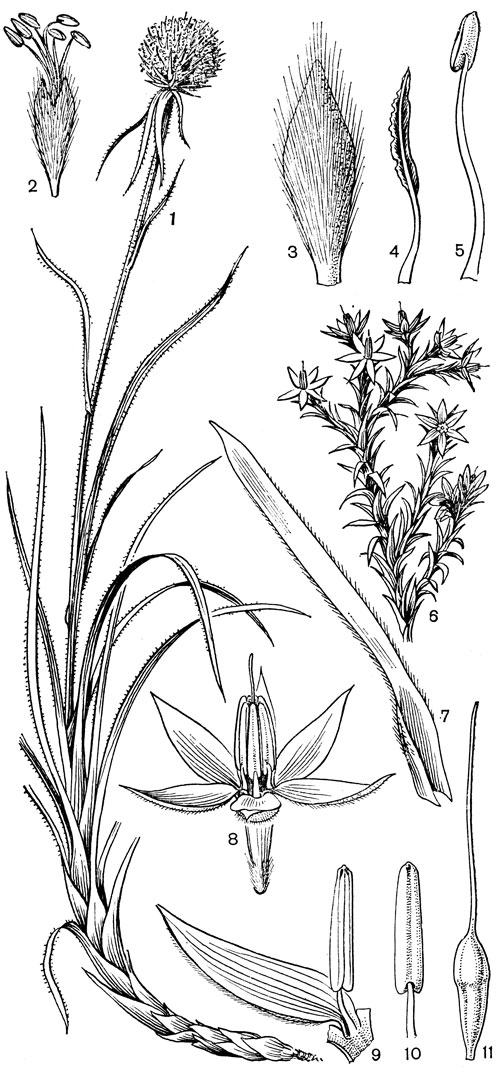
Рис. 78. Ксанторреевые. Дазипогон бромелиелистный (Dasypogon bromeliifolius): 1 - общий вид; 2 - цветок; 3 - сегмент внешнего круга околоцветника; 4 - сегмент внутреннего круга околоцветника; 5 - тычинка. Калектазии синецветковая (Calectasia cyanea): 6 - цветущая ветвь; 7 - лист; 8 - цветок; 9 - часть цветка с сегментом околоцветника и тычинкой; 10 - тычинка; 11 - гинецей
Листья ксанторреевых почти всегда узколинейные, с более или менее развитыми влагалищами при их основании. Как у видов ксанторрей, листья которых достигают в длину 1 м и более, так и у мелколистных акантокарпуса и калектазии старые листья обычно обламываются близ их основания, а влагалища некоторое время еще сохраняются на каудексе или ветвях стебля. Нередко по краю листьев имеются шипиковидные или сосочковидные выросты, а также волоски, особенно обильные близ основания листьев кингии. Существенное систематическое значение имеет форма листьев на поперечном сечении, которая в пределах рода ксанторрея может быть веретеновидной, трехгранной или четырехгранной. У ломандры листья варьируют от плоских и довольно широких до щетиновидно вдоль свернутых, а у ломандры цилиндрической (L. cylindrica) и близких к ней видов они цилиндрические. Хвоеподобные листья калектазии, имеющие длину всего 3-15 мм, заканчиваются на верхушке острым шипиком. Парацитные или анемоцитные устьица листьев ксанторреевых часто погружены в борозды, что, как и сильно развитые склеренхимные тяжи, является приспособлением к засушливым условиям обитания.
Всегда актиноморфные 3-членные обоеполые (у большинства родов) или однополые и тогда двудомные (у видов ломандры) цветки ксанторреевых обычно собраны в соцветия самого различного строения: очень густые цилиндрические колосья у ксанторреи, шаровидные головки у кингии и дазипогона, метелки, кисти или мелкие головки у ломандры и других родов. Хотя цветки в колосьях или головках кажутся сидячими, эти соцветия в действительности являются сложными и образованы спирально расположенными на оси общего соцветия частными цимозными соцветиями в виде пучков сидячих или почти сидячих цветков. Этим объясняется обилие в таких соцветиях прицветникообразных листочков, часть которых может быть верховыми листьями, часть - прицветниками и часть - прицветничками. Крупные одиночные цветки на коротких ножках, выходящих из пазух прикорневых листьев, имеет бакстерия, а у калектазии более мелкие, но также одиночные сидячие цветки расположены на верхушках ветвей и в их развилках. Початкообразные соцветия ксанторрей могут достигать очень большой длины (у ксанторреи смолистой - до 4 и даже 5 м). Мужские и женские соцветия у видов ломандры часто заметно отличаются друг от друга: мужские соцветия обычно крупнее женских, но мужские цветки мельче женских. У видов секции тифопсис (Typhopsis) этого рода, например у ломандры белоголовчатой (L. leucocephala), прицветникообразные листочки в соцветии рассечены на многочисленные волоски, окутывающие цветки, подобно цветкам в початкообразных соцветиях рогоза (Typha).
Как уже упоминалось, околоцветник ксан торреевых производит впечатление сухого обычно он перепончатый или кожисто-перепончатый, иногда при плодах еще более отвердевающий. Его 6 сегментов, расположенные двумя кругами, у видов ксанторреи вполне свободные, но у других родов семейства могут различным образом срастаться между собой. У калектазии синецветковой 6-лопастный отгиб околоцветника окрашен в синий цвет, а нижние части сегментов срослись в довольно длинную и узкую трубку, заостренную к основанию. У дазипогона 3 внешних сегмента срастаются в нижней части в трубку, а более узкие внутренние сегменты остаются свободными. Некоторые виды ломандры, напротив, имеют внешние сегменты свободные, а внутренние почти на половину своей длины сросшиеся. У ксанторреи и некоторых других родов наружные сегменты околоцветника перепончатые и слабо окрашенные, похожие на прицветники, а сегменты внутреннего круга заметно более крупные и окрашены в белый, желтоватый или красноватый цвет, напоминая этим лепестки.
Тычинок 6, расположенных 2 кругами. Пыльники прикреплены к нитям или спинкой (у ксанторреи и некоторых других родов), или более или менее двулопастным основанием (у кингии и калектазии), что является очень существенным систематическим признаком. Тычинки калектазии имеют довольно короткие нити, прикрепленные у основания отгиба околоцветника. У других ксанторреевых нити тычинок обычно более длинные, после распускания цветков отгибающиеся кнаружи и могут быть или до основания свободными, или (у трех внутренних тычинок) срастающимися с основанием внутренних сегментов околоцветника. Пыльники вскрываются интрорзно продольными щелями, но у калектазии пыльца в основном высыпается не через них, а через поры на верхушках обоих гнезд пыльника. Пыльцевые зерна у большинства родов имеют однобороздную оболочку и гладкую поверхность, хотя у некоторых видов ломандры на ней отмечаются мелкие шипики. У акантокарпуса кольцевая бороздка делит зерно пополам. Очень оригинально строение оболочки пыльцевых зерен у бакстерии, где ее поверхность разделена на несколько геометрически правильных пятиугольных или шестиугольных участков.
Гинецей ксанторреевых имеет много общего с другими родственными лилейным семействами. Обычно он образован 3 плодолистиками и имеет 3-гнездную завязь со столбиком, заканчивающимся цельным или трехлопастным рыльцем. У ксанторреи в каждом гнезде завязи имеется от 2 до 8 гемитропных семязачатков, из которых в семя обычно развиваются 1-3. Большинство других родов содержит по одному семязачатку в каждом гнезде, но у калектазии и дазипогона в одногнездной завязи имеются 3 семязачатка, из которых лишь один развивается в семя. Плод большинства ксанторреевых - локулицидная коробочка. Нераскрывающиеся односемянные орешкообразные плоды, одетые сохраняющимся и способствующим распространению диаспор околоцветником, характерны для видов кингии, дазипогона и калектазии. У всех родов семейства остаток столбика сохраняется при зрелом плоде в виде носика, часто немного смещенного в сторону от верхушки плода. У акантокарпуса вскрывающиеся плоды покрыты шипикообразными выростами, вероятно, имеющими защитное значение. Семена ксанторреи имеют блестящую черную оболочку, почти горизонтально расположенный зародыш и небольшой беловатый рубчик на одном из концов. Если они образуются по нескольку в одном гнезде, то они имеют различную величину и форму. У других родов в каждом гнезде развивается только одно семя, обычно имеющее коричневую оболочку, твердый маслянистый эндосперм и прямой или слабо согнутый зародыш.
Выше уже отмечалась существенная роль древовидных видов ксанторреи и кингии в ландшафтах многих районов Австралии. Обычно они встречаются группами на открытых участках австралийской саванны, иногда напоминающих вересковые пустоши северного полушария, или среди зарослей ксерофильных кустарников, так называемого скрэба, реже на обнажениях песчаника и других каменных пород по склонам холмов и гряд. Менее крупные виды - ксанторреи смолистая и малая - растут в более влажных местообитаниях, часто на песчаных и торфянистых пустошах, а иногда даже на болотах. Можно отметить, что, кроме широко распространенного в Австралии названия древовидных ксанторрей - "злаковые деревья", местное население использует для них еще одно название - "черный мальчик" ("black boy"), основанием для которого послужило некоторое сходство особей этих растений с темнокожими людьми, несущими на голове пучок травы. Другие роды ксанторреевых обычно также встречаются в саваннах и саванноподобных группировках растительности, лишь редко заходя в разреженные леса. Замечательный своим обликом, а также блестящими синими цветками с ярко-желтыми тычинками, полукустарник калектазия синецветковая обычно растет близ морского побережья на хорошо дренированных почвах песчаных пустошей вместе с другими эрикоидными кустарниками и полукустарниками или видами банксии из семейства протейных. Сильно разветвленные стебли этого вида обычно достигают в высоту лишь 50-60 см.
Биологические особенности ксанторреевых еще недостаточно изучены. По-видимому, все они являются поликарпиками, но могут цвести не каждый год. По способу опыления они принадлежат к числу ветроопыляемых растений, хотя не исключено, что некоторую роль в переносе пыльцы могут играть и короткохоботковые насекомые, поедающие пыльцу или посещающие соцветия в поисках убежища. Неприятный запах, свойственный цветкам ксанторреи Прейса, возможно, привлекает мух. Если у видов ломандры самоопылению препятствует двудомность, то у ксанторреевых с обоеполыми цветками, вероятно, имеет место дихогамия. Так, в цветках калектазии пыльца высыпается из верхушечных пор уже в самом начале цветения или еще в бутоне. Развитие же столбика всегда запаздывает, вследствие чего рыльце оказывается на одном уровне с порами пыльников только тогда, когда пыльцы в них уже нет (протандрия). Однако часть пыльцы может сохраняться на лопастях околоцветника, так что самоопыление у этого вида все же не исключается полностью. В роде ксанторрея отмечается довольно широкое распространение межвидовой гибридизации, что отчасти определяется присутствием у всех изученных видов этого рода одного и того же числа хромосом (2n = 22).
У родов ксанторреевых с плодами-коробочками, например у ксанторреи, семена просто выпадают из коробочек во время раскачивания плодоносящих соцветий ветром. В дальнейшем они могут распространяться с помощью временных водных потоков или с комочками почвы на ногах животных. У родов с не вскрывающимися односемянными плодами - кингии, дазипогона и калектазии - диаспорами служат плоды, одетые сухим околоцветником, а иногда и прицветниками. Присутствие околоцветника придает диаспорам значительную парусность, и они могут разноситься на большие расстояния. Специализация к анемохории особенно хорошо выражена у калектазии. Ее мелкие односемянные плоды располагаются в нижней части узкой трубки околоцветника на короткой плодоножке. Основание трубки заострено и покрыто жесткими, вверх направленными волосками, что позволяет диаспорам калектазии, подобно диаспорам ковылей, не только закрепляться среди травяной подстилки, но и как бы ввинчиваться в нее или в песок. Однако жизнеспособность семян у этого вида, вероятно древнего реликта, очень низкая.
Несмотря на небольшое количество родов (9), система ксанторреевых все еще недостаточно разработана. Не без оснований некоторые авторы предлагают разделить его на несколько самостоятельных семейств. В последнее время A. Л. Тахтаджян (1980), основываясь на анатомических, палинологических и других данных, делит это семейство на 4 подсемейства: собственно ксанторреевые (Xanthorrhoeoideae) с родом ксанторрея; ломандровые (Lomandroideae) с родами ломандра, ромнальда, акантокарпус, хамексерос; кингиевые (Kingioideae) - кингия и бакстерия и дазипогоновые (Dasypogonoideae) с родами дазипогон и калектазии. Следует отметить, что пары родов, составляющие два последних подсемейства, настолько отличаются друг от друга по внешнему облику и другим признакам, что каждое из этих подсемейств может быть разделено на две монотипные трибы.
Хозяйственное значение ксанторреевых невелико. Многие из них, несмотря на жесткость листьев, могут использоваться в качестве кормовых растений для скота, однако некоторые виды ксанторреи и ломандры токсичны и вызывают расстройства органов движения. Древесина древовидных видов этого семейства служит строительным материалом и идет на различные поделки. Выделяемую ксанторреями смолу местное население использует для изготовления лака, но не очень высокого качества. Особенно известна в этом отношении желтая смола ксанторреи смолистой. Она имеет острый вкус и горит с приятным бензойным запахом. В меньшем количестве используют коричневую или красную смолу других ксанторрей. Смолу их применяют также в виде клея. У аборигенов Австралии ножки соцветий ксанторрей шли на изготовление копий, наконечники к которым приклеивали этой смолой. Древовидные виды ксанторреевых очень декоративны и, несомненно, заслуживают широкого введения их в культуру в садово-парковых хозяйствах наряду с юкками, драценами, агавами и дориантесом. Не менее ценное декоративное растение - калектазия синецветковая, которую часто считают одним из красивейших растений Австралии. К сожалению, биологические особенности всех этих видов изучены слабо, а потому возможности введения их в культуру очень ограничены.
Семейство афиллантовые (Aphyllanthaceae) (Т. В. Вельгорская)
К семейству афиллантовых относится один род афиллантес (Aphyllanthes), единственный представитель которого - афиллантес монпельенский (A. monspeliensis, рис. 79) - обитает в странах Западного Средиземноморья: на Пиренейском полуострове (кроме северо-западных районов Кантабрийского нагорья), на Балеарских островах, в Южной Франции, в Северной Италии (Пьемонт, Лигурия) и на острове Сардиния. В 1943 г. итальянские ботаники В. Джакомини и Н. Ариетти обнаружили афиллантес в Итальянских Альпах в окрестности города Брешиа (Ломбардия). На Африканском континенте афиллантес встречается в Марокко и Алжире.
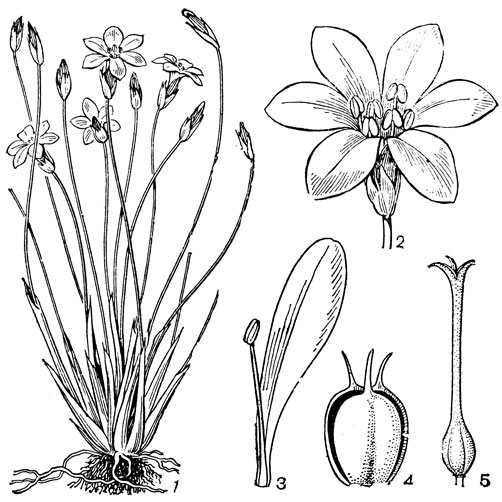
Рис. 79. Афиллантес монпельенский (Aphyllanthes monspeliensis): 1 - общий вид; 2 -цветок (гинецей удален); 3 - тычинка с сегментом околоцветника; 4 - плод; 5 - гинецей
Растет афиллантес монпельенский на каменистых и скалистых сухих почвах и, наряду со злаками, осоками, некоторыми представителями семейств губоцветных (видами розмарина, тимьяна, лаванды) и ладанниковых, является одним из главных компонентов растительности средиземноморской гарриги. В европейском Средиземноморье афиллантес широко распространен на высотах от 500 до 1200 м над уровнем моря, но никогда не встречается в прибрежных районах. В Северной Африке, в Атласских горах Марокко, афиллантес растет в кустарниках и на открытых местах горного леса вплоть до высоты 2700 м над уровнем моря.
Афиллантовые - многолетние травы с коротким симподиальным корневищем, характеризующимся вторичным ростом. Многочисленные жесткие, высотой 20-50 см, тонкие, ребристые, безлистные стебли являются единственным фотосинтезирующим органом растения. Сильно редуцированные двурядные пленчатые листья длиной 3-8 см собраны у основания стеблей и превращены в красновато-коричневые нефотосинтезирующие защитные органы. Сосуды только в корнях, членики их с простой перфорацией. Каждый стебель несет на своей верхушке редуцированное головчатое соцветие с 1, реже 2-3 цветками. Соцветие окружено 5 красновато- или желтовато-коричневыми перепончатыми прицветниками, расположенными черепитчато в 2 ряда. Цветки обоеполые, актиноморфные, диаметром 2,5см, ярко-синие или фиолетово- голубые, с темными прожилками, редко белые. Околоцветник из двух 3-членных кругов, венчиковидный; сегменты околоцветника одинаковые, у основания сросшиеся в трубку. Тычинок 6, расположенных в 2 круга (3 короткие и 3 более длинные) и прикрепленных тонкими нитями к нижней части сегмента околоцветника. Пыльники маленькие, интрорзные; оболочка пыльцевых зерен более или менее спирапертурная, очень сходна с оболочкой пыльцевых зерен ломандры Эндлихера (Lomandra endlicheri) из семейства ксанторреевых. Гинецей синкарпный, с нитевидным столбиком, заканчивающимся 3-лопастным рыльцем; завязь верхняя, 3-гнездная, с одним базальным анатропным семязачатком в каждом гнезде. Плод - перепончатая локулицидная коробочка с 3 овальными семенами, покрытыми черной мелкоморщинистой кожурой.
Цветет афиллантес в апреле - июле; опыляется насекомыми, главным образом пчелами и дневными бабочками, которых привлекает яркая окраска цветков. В открытом цветке афиллантеса пыльники трех более длинных тычинок находятся чуть выше лопастей рыльца, а три короткие тычинки более или менее ниже длинного столбика, и в начале цветения пыльники не касаются рыльца. Если по какой- либо причине не произошло перекрестного опыления, то не исключена контактная автогамия, так как, отцветая, сегменты околоцветника смыкаются и тычинки склоняются к рыльцу.
Снабженный тремя шиповидными выростами плод афиллантеса, скорее всего, распространяется зоохорно.
Практического значения афиллантес монпельенский почти не имеет.
Семейство гангуановые (Hanguanaceae) (Е. А. Земскова)
Семейство включает один род гангуана (Hanguana), представленный одним видом - гангуана малайская (Н. malayana, рис. 80), распространенным на Шри-Ланке, Малайском архипелаге, острове Палау и на севере Австралии.
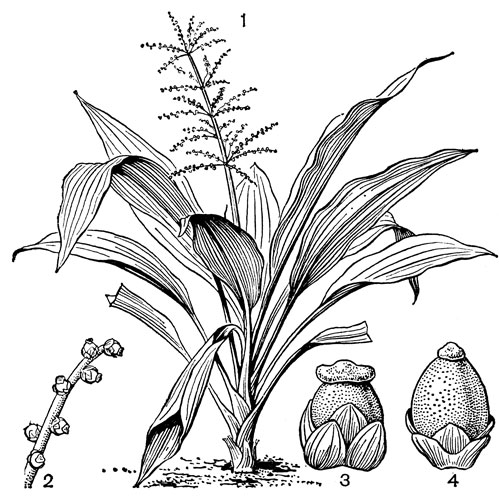
Рис. 80. Гангуана малайская (Hanguana malayana): 1 - общий вид; 2 - часть соцветия; 3 - женский цветок; 4 - плод
Гангуана - многолетнее травянистое растение с высоким прямостоячим жестким стеблем и длинным ползучим или плавающим корневищем. Среди представителей рода встречаются водные формы, растущие на болотах или в медленно текущей воде, и наземные формы, обитающие во влажных лесах. Листья большей частью прикорневые, на длинных черешках. Цветки мелкие, зеленые или желтые, собранные в метелки, с крупными брактеями, актиноморфные, двудомные. В женских и мужских цветках околоцветник чашечковидный, остающийся, из 6 свободных, расположенных в два круга сегментов. В мужских цветках 6 тычинок, сросшихся у основания с околоцветником. Пыльники маленькие, прикрепленные основанием, вскрывающиеся интрорзно продольной щелью. В женских цветках 6 стаминодиев (3 более крупных и 3 мелких, лишенных пыльников). Гинецей синкарпный, из 3 плодолистиков, с 3 широкими сидячими рыльцами, в каждом гнезде завязи по одному семязачатку. В мужских цветках завязь рудиментарная. Плод ягодообразный, мясистый, с толстыми стенками, с 1-3 семенами. Семена имеют толстую кожуру, обильный эндосперм и маленький зародыш.
Гангуановые - энтомофильные растения. Верхняя часть рыльца четко выделяется коричневой окраской, которая привлекает насекомых. В мужских цветках эту роль выполняют нектарники, атрофированные в женских цветках. Водные и наземные формы гангуаны различаются главным образом по размеру. Водные формы, кроме прикорневых листьев, имеют также почти сидячие стеблевые листья. Водные растения более мощные, длина стебля иногда достигает 2 м.
От основания стебля отходят длинные побеги, которые на верхушке развиваются в новое растение, в свою очередь также дающее побеги, часто образуя большую плотную непроницаемую массу. Части этой массы могут отделяться течением воды от материнского растения и образовывать "плавающие острова". Способность плавать обусловлена наличием многочисленных воздухоносных полостей у растения. Свежее корневище длиной 50 см, диаметром 10 см имеет массу 1,5 кг. Наземные формы, как правило менее мощные, с немногочисленными короткими и слабыми побегами, не образуют плотные массы, так как после образования цветков только один столон развивается в новое растение, на некотором расстоянии от материнского. На гангуановых болотах гангуана часто подавляет другие виды растений за исключением некоторых орхидных.
Жители Новой Гвинеи употребляют в пищу стебли и побеги водных форм гангуаны.
Семейство спаржевые (Asparagaceae) (Л. И. Иванина)
В семействе 25 родов и около 550 видов, большинство из которых распространено в северном полушарии, в тропической и Южной Африке и на Мадагаскаре; лишь немногие представители спаржевых встречаются в Центральной Америке, на островах Малайского архипелага, а также в Австралии и Тасмании.
Спаржевые преимущественно многолетние, нередко вечнозеленые травы или кустарнички, редко кустарники, иногда лианы. Все представители семейства имеют корневища, обычно густо покрытые придаточными корнями. Сосуды большей частью только в корнях и обычно с лестничной перфорацией, но у данаи (Danae) сосуды с лестничной перфорацией имеются и в стеблях, а у семелы (Semele) и спаржи (Asparagus) сосуды в корнях с простой перфорацией, в то время как в стеблях перфорация лестничная.
Листья приземные (базальные) или расположены по всему стеблю (большей частью очередные, реже супротивные и мутовчатые), с параллельным или дугонервным жилкованием, часто с влагалищным основанием, сидячие или редко с черешками. Листья у представителей подсемейства иглицевых (Ruscoideae) и собственно спаржевых (Asparagoideae) недоразвитые, в виде мелких пленчатых чешуй, из пазух которых развиваются особые ассимиляционные листовидные органы (видоизмененные побеги) - филлокладии или кладодии.
Цветки у спаржевых некрупные или мелкие с прицветниками, пазушные или верхушечные, одиночные или чаще образуют кистевидные, колосовидные, а иногда зонтиковидные соцветия. У большинства видов цветки обоеполые, реже однополые, обычно функционально мужские или функционально женские, двудомные или однодомные. Они обычно 3-членные, реже 2- или 4-членные. Сегменты околоцветника более или менее сросшиеся, реже почти свободные. Тычинок обычно 6, реже 3 или 4 и очень редко 8 или 12. Тычиночные нити обычно срастаются с сегментами околоцветника на разных уровнях, редко тычинки срастаются по всей длине в колонку (у иглицевых). Пыльники прикреплены к нити основанием или спинкой, большей частью интрорзные, реже полуэкстрорзные или экстрорзные. Пыльцевые зерна однобороздные. У некоторых представителей семейства имеются септальные нектарники, реже петальные нектарники (у видов диспорума - Disporum). Гинецей большей частью из 3 плодолистиков, реже из 4 или 2, синкарпный (только у данаи - с неполными перегородками). Завязь преимущественно верхняя (за исключением родов пелиосаптеса - Peliosanthes и офиопогона - Ophiopogon, у которых она полунижняя или почти нижняя). Семязачатков в каждом гнезде 2, редко 1 или несколько; они ортотропныег гемитропные или анатропные (спаржа).
Плод обычно ягода, и только у офиопогона, лириопы (Liriope) и пелиосантеса плод переходный между ягодой и коробочкой, обычно с 1-2 или несколькими шаровидными семенами. Семена с эндоспермом и недифференцированным зародышем, у некоторых видов купены (Polygonatum) и майника (Maianthemum) семена имеют ариллусовидные образования.
Семейство спаржевые в принятом здесь более широком его понимании включает 3 подсемейства: ландышевые, иглицевые и собственно спаржевые (А. Л. Тахтаджян, 1980). Некоторые современные ботаники, как X. Хубер (1969) и М. Дальгрен (1980), считают их самостоятельными семействами.
Представители подсемейства ландышевых (Convallarioideae) - некрупные многолетние травы, встречающиеся почти по всему ареалу семейства, за исключением Африки. Листья у них сравнительно крупные, приземные или стеблевые. Цветки почти всегда обоеполые, средних размеров, обычно собраны в кистевидные или колосовидные соцветия, редко одиночные. Околоцветник колокольчатый, трубчатый, почти полушаровидный, реже другой формы. Тычинок 4, 6, 8 или 12; тычиночные нити свободные. Завязь верхняя, редко полунижняя или почти нижняя, обычно 3-гнездная, редко 2- или 4-гнездная. Ландышевые - самое большое подсемейство среди спаржевых. В него входят 5 триб: аспидистровые (Aspidistreae), пелиосантовые (Peliosantheae), офиопогоновые (Ophiopogoneae), купеновые (Polygonateae) и собственно ландышевые (Convallarieae).
Самыми своеобразными по строению цветка (нередко также и по строению соцветий) и относительно наиболее примитивными являются представители трибы аспидистровых, которых японский ботаник Т. Накаи (1936) выделял в самостоятельное семейство аспидистровые (Aspidistraceae). Они распространены в Гималаях и в Восточной и Юго-Восточной Азии. Это вечнозеленые многолетние травы, обычно с толстым корневищем и приземными листьями. Цветки в колосовидных или кистевидных соцветиях, иногда одиночные. Околоцветник мясистый, из 6-8 (редко 12) сросшихся сегментов. Число тычинок равно числу сегментов околоцветника; пыльники почти сидячие. Столбик короткий, рыльце большей частью щитовидное или широко-трехлопастное. В трибе аспидистровых 6 родов. Наиболее известный род этой трибы - аспидистра (Aspidistra, около 11 видов, рис. 81), произрастающая в Восточных Гималаях, Юго-Западном и Южном Китае, на островах Хайнань и Тайвань, а также в Индии (Верхний Ассам) и в Северном Вьетнаме. Виды аспидистры обитают по склонам ущелий, на тенистых скалах, в лесах, в горах, главным образом в нижнем и среднем поясах на высоте от 300 до 1800 м над уровнем моря. Некоторые виды (особенно аспидистра высокая - A. elatior) широко распространены в культуре, их выращивают как комнатные растения, применяют для озеленения помещений, а в условиях теплого климата сажают в садах и парках. Как и у остальных видов, корневище аспидистры высокой мясистое, толщиной 1-1,5 см и жесткое. Оно состоит из коротких междоузлий, несущих бледные чешуевидные листья. В пазухах каждого из них образуется по одному цветку, сидящему на короткой цветоножке. Цветки распускаются весной. Опыление аспидистры высокой было изучено еще известным итальянским ботаником Ф. Дельпино (1875). Некрупные, преимущественно зеленовато-красные, мясистые, неприятно пахнущие цветки почти лежат на земле. Широкое зонтиковидное складчатое рыльце закрывает вход в полушаровидный околоцветник. Мелкие короткохоботковые мухи, садясь на цветок, сначала попадают на большое рыльце, затем сползают по его складкам на стенки околоцветника и попадают на дно цветка. Некоторое время они остаются в цветке, возможно, находят там приют в прохладную погоду (Э. Лёв, 1895), а может быть, просто не могут сразу вылезти, так как выход из цветка закрыт рыльцем. Выползая по стенкам между расположенными в нижней части околоцветника пыльниками, они вымазываются пыльцой и затем, перелетая на другой цветок, совершают перекрестное опыление. Э. Лёв полагал, что в плохую погоду, когда мало насекомых, их заменяют улитки. Однако, по мнению других авторов, улитки являются скорее лишь случайными опылителями.
К аспидистре близок монотипный и своеобразный род эврардиелла (Evrardiella), произрастающий во Вьетнаме, в горах Чыонгшон (Аннам) (рис. 81).
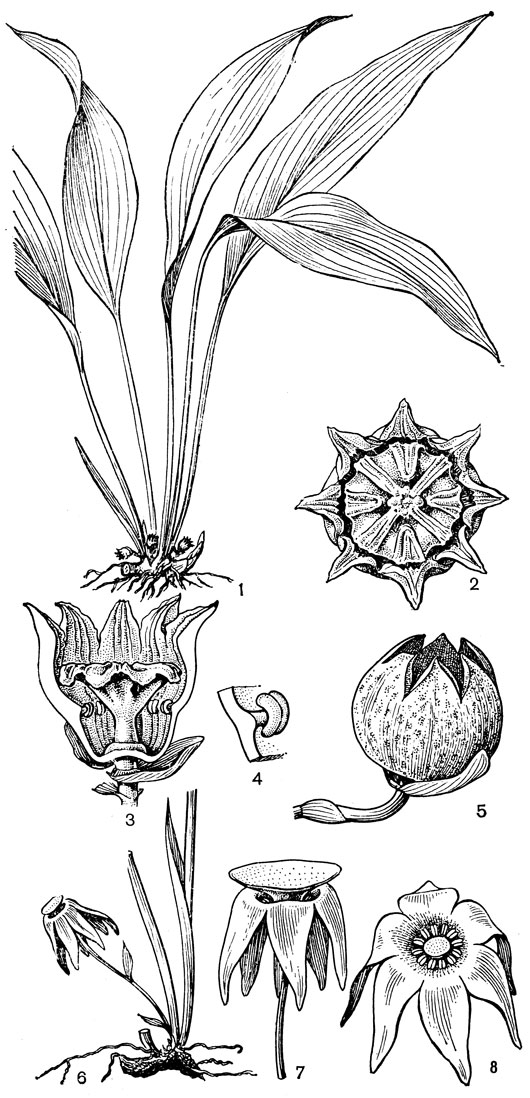
Рис. 81. Спаржевые. Аспидистра высокая (Aspidistra elatior): 1 - общий вид; 2 - цветок (вид сверху); 3 - продольный разрез цветка; 4 - тычинка. Аспидистра типичная (A. typica): 5 - бутон. Эврардиелла двенадцатитычиночная (Evrardiella dodecandra): 6 - нижняя часть растения; 7 - цветок (вид сбоку); 8 - цветок
На островах Японии, на полуострове Корея и во многих провинциях континентального Китая встречается относящийся к аспидистровым монотипный род родея (Rhodea). Родея японская (R. japonica) отличается красивыми темно-зелеными приземными листьями. Весной, в мае - июне, появляется невысокий цветонос с колосовидным соцветием и с бледно-желтыми цветками. Столбик у родеи короткий, с неясно 3-раздельными округлыми лопастями. Ягоды красные, шаровидные, односемянные. Родея - декоративное растение, особенно распространенное с древних времен в Японии и Китае. В листьях родеи обнаружены гликозиды карденолидной природы и другие вещества, "обладающие высокой кардиотонической актив-ностью. В лесах Восточных Гималаев, Юго-Западного Китая и Индокитая произрастают 9 видов еще одного рода аспидистровых - кампиландры (Campylandra), у которой цветки собраны в короткий колос с хорошо заметными длинными, острыми, линейно-ланцетными прицветниками, по размерам в несколько раз превосходящими цветки. К кампилаидре и родее весьма близок род тупистра (Tupistra), насчитывающий около 15 видов, обитающих в Гималаях, Бирме, Индии (штаты Ассам, Манипур), Юго-Западном и Южном Китае, на полуострове Малакка и острове Суматра. Виды тупистры произрастают в тенистых лесах и в зарослях кустарников (на высоте 600-1500 м над уровнем моря). Цветки короткоколокольчатые, их рыльца с расширенными лопастями. Тупистры - декоративные растения.
В трибу пелиосантовых входит один род - пелиосантес (Peliosanthes), отличающийся формой завязи и строением плода. В роде пелиосантес около 10 близких видов, объединяемых Дж. Джэссопом (1976) в один вид. Они распространены в Восточных Гималаях, в Южном и Юго-Восточном Китае, на островах Хайнань и Тайвань, в Индии, Бангладеш, Бирме, Индокитае, на полуострове Малакка, островах Малайского архипелага. Произрастают преимущественно в дождевых лесах, от равнины до 300-1600 м над уровнем моря, иногда поселяются на скалах и часто близ воды. Пелиосантесы (рис. 82) - многолетние вечнозеленые растения с более или менее коротким корневищем. Приземные листья от линейных до продолговато-яйцевидных, постепенно переходящие в длинный черешок. Мелкие цветки на цветоножках собраны по одному или нескольку в пазухах прицветников в более или менее длинное, сравнительно тонкое, кистевидное соцветие с редкими чешуевидными листьями при основании. Околоцветник мясистый, в нижней части с конусовидной трубкой, часто приросшей к завязи, разнообразный по окраске - серо-голубой, иногда белый, фиолетовый, пурпурный. Андроцей из 6 тычинок с очень короткими нитями, прикрепленными к трубке околоцветника. Завязь полунижняя или почти нижняя. Рыльце сидячее. Плод ягодообразный, близкий к коробочке, рано растрескивающийся и освобождающий семена, которые затем развиваются вне плода. Семена обычно голубые, твердые, с мясистой оболочкой.
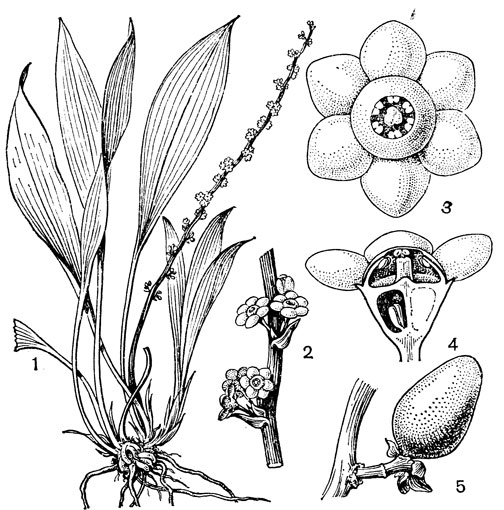
Рис. 82. Пелиосантес тэта (Peliosanthes teta): 1 - общий вид; 2 - часть соцветия; 3 - цветок (вид сверху); 4 - разрез цветка; 5 - плод
К трибе офиопогоновых относятся 2 рода - лириопа и офиопогон, распространенные в Гималаях и Восточной Азии. Это небольшие многолетние травы с линейными и ланцетными длинными приземными листьями. Корневища с несколькими клубневидными корнями, иногда со столонами. Цветки расположены на цветоносе в пучках по 3-8, реже одиночные и образуют обычно кистевидное соцветие. Сегменты околоцветника широко открытые; у офиопогона околоцветник внизу образует короткую трубку, сросшуюся с завязью; завязь полунижняя или нижняя; 6 свободных тычинок; столбик с маленьким 3-раздельным рыльцем. Плод офиопогоновых отличается характером вскрытия перикарпия. Околоплодник скоро разрушается и открывает мясистые красные семена, в распространении которых большую роль играют птицы.
В роде лириопа (Liriope) 8 видов, произрастающих в Японии, на полуострове Корея, в континентальном Китае, на острове Тайвань, во Вьетнаме и на Филиппинах. Виды лириопы обитают на равнинах и у подножий холмов, преимущественно по тенистым склонам и берегам рек, в горах встречаются до высоты 1400-2300 м. У лириопы голубоватые, фиолетовые или бледно-сиреневые цветки с колесовидным околоцветником, собранные в кистевидное соцветие. В бутоне (рис. 83, 2) нити тычинок прямые, почти равные по длине столбику, и незрелые пыльники и рыльце находятся почти на одном уровне. По мере распускания цветка нити поворачиваются в одну сторону и как бы слегка полегают. В таком положении открытые пыльники не соприкасаются с рыльцем. Опыление может происходить только при участии насекомых опылителей. Под конец цветения тычиночные нити слегка выпрямляются и пыльники располагаются почти на одном уровне с рыльцем. В это время может произойти самоопыление. Лириопа встречается в культуре во многих странах, особенно в Японии и Китае. Как декоративные преобладают 3 вида - лириопа колосистая (L.spicata, табл. 21,1), близкий к ней вид лириопа злаколистная (L. graminifolia) и лириопа мускари (L. muscari). В нашей стране лириопу используют для бордюров в садах и парках Черноморского побережья Кавказа. Ее можно выращивать и как горшечную культуру на севере страны. Сладкие ароматные клубни лириопы колосистой и лириопы злаколистной употребляют в медицине в Японии, Китае и Индокитае.
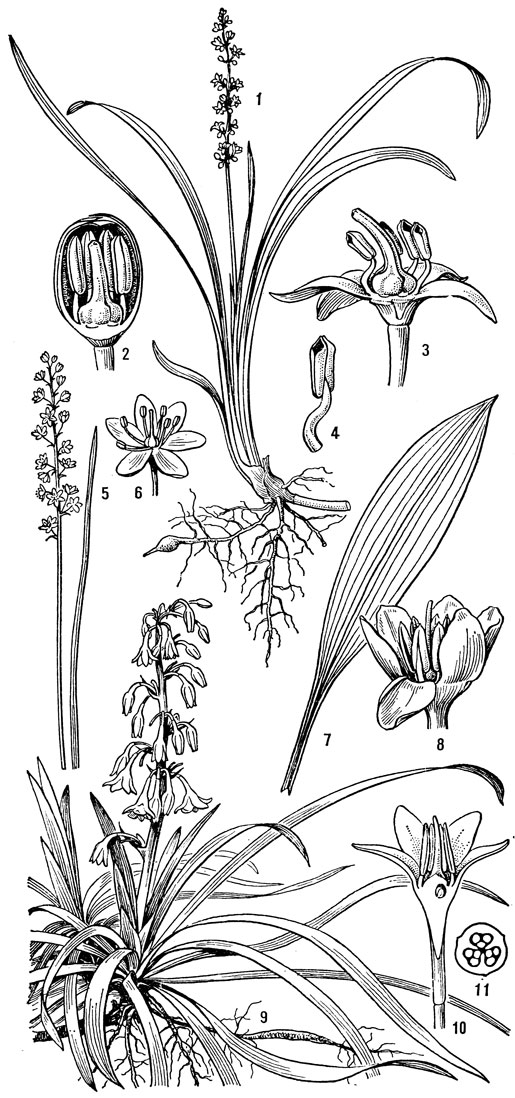
Рис. 83. Спаржевые. Лириопа колосистая (Liriope spicata): 1 - общий вид; 2 - бутон; 3 - цветок (с частью удаленных сегментов); 4 - тычинка. Лириопа ганьсуйская (L. kansuensis): 5 - соцветие и лист; 6 - цветок. Офиопогон тонкинский (Ophiopogon tonkinensis): 7 - лист; 8 - цветок. Офиопогон ябуран (О. jaburan): 9 - общий вид; 10 - цветок в разрезе; 11 - поперечный разрез завязи

Таблица 21. Спаржевые: 1 - лириопа колосовидная (Liriope spicata), оранжереи Ботанического института АН СССР в Ленинграде; 2 - иглица понтийская (Ruscus ponticus), Крым, государственный заповедник 'Мыс Мартьян'; 3 - иглица понтийская, Крым, Никитский ботанический сад; 4 - иглица подъязычная (R. hypoglossum), Крым
В близком к лириопе роде офиопогон, или змеебородник (Ophiopogon), насчитывается 50 видов, из которых 33 встречаются в Китае. Возможно, что число видов несколько преувеличено, так как для офиопогонов характерна большая изменчивость признаков, и некоторые формы или разновидности были описаны в качестве видов. Представители рода офиопогон произрастают в Тибете, Гималаях, Индии (Ассам, Манипур), в континентальном Китае (за исключением Северного) и на острове Хайнань, в Северном Вьетнаме, Северном Таиланде, Японии и Малезии. Офиопогоны широко используются в субтропических районах как бордюрные и почвопокровные растения. Наиболее часто встречаются офиопогон ябуран (О. jaburan, рис. 83, 9) и офиопогон японский (О. japonicus). Особую роль в качестве декоративных растений играют их пестролистные формы. В Японии и Китае офиопогон японский применяют в качестве лекарственного растения.
Триба купеновых содержит 8 родов. Представители большинства из них имеют облиственный стебель. Цветки у них пазушные, одиночные или в пучках, реже в полузонтиках или метелках. Сегменты околоцветника почти свободные или сросшиеся. Тычинок 6 или 4, семязачатков обычно много. Плод - ягода. Ареал родов не ограничен Восточной Азией, как у предыдущих триб, а более обширен: он заходит в умеренную и частично в тропическую зону обоих полушарий. Среди купеновых наименее продвинуты 3 рода - диспорум, стрептопус (Streptopus) и клинтония (Clintonia).
В роде диспорум (Disporum) известно 20 видов, распространенных в Гималаях, Восточной и Юго-Восточной Азии и в Северной Америке (в Кордильерах и в восточной части). Они растут во влажных горных лесах, по холмам и склонам гор до 3000 м над уровнем моря. Диспорумы характеризуются тонкими ползучими корневищами, маловетвистыми стеблями и продолговатыми или яйцевидными, почти сидячими листьями. Кремово-белые или зеленоватые, реже розовые цветки диспорума с околоцветником из свободных сегментов расположены в пазухах листьев на концах побегов по одному или по 2-5 на длинных цветоножках. Преобладают две формы околоцветника: воронковидная и чашевидная, им соответствуют разные способы опыления (Ф. Утех и Ш. Кавано, 1976). Воронковидная форма цветков рассматривается авторами на примере диспорума сидячего (D. sessile). У него цветки относительно большие, длиной 2-3 см, основание сегментов слегка мешковидной формы (рис. 84, 5, 4). В нижней части сегментов находятся базальные петальные нектарники. Постоянный опылитель цветков этого вида - крупная одиночная пчела. Подлетая к цветку снизу, она сначала задевает выступающее из цветка-рыльце, а затем легко раздвигает свободные сегменты околоцветника и при этом прижимает 3-раздельное рыльце и садится "верхом". Касаясь пыльников мохнатой головой и грудью, пчела сметает с них пыльцу. Длинным хоботком она достигает нектарников, расположенных сравнительно глубоко в мешковидном основании сегментов околоцветника. Покидая цветок, пчела "пятится" между сегментами и пыльниками и опять обильно вымазывается пыльцой, теперь уже лапками и нижней частью брюшка. Перелетая на другой цветок, она опускается на рыльце и производит перекрестное опыление. Движения пчелы быстрые: на один цветок она тратит примерно 6 секунд. Диспорум смилациновый (D. smilacinum) - пример цветков с чашевидным околоцветником. Благодаря относительно открытым нектарникам при основании сегментов околоцветника и открытому венчику его цветок посещают более многочисленные неспециализированные посетители, часть из которых является и опылителями. Кроме того, открытый венчик способствует ветроопылению и, возможно, самоопылению. Несколько видов диспорума изредка используются в качестве декоративных растений и выращиваются в ботанических садах и парках.
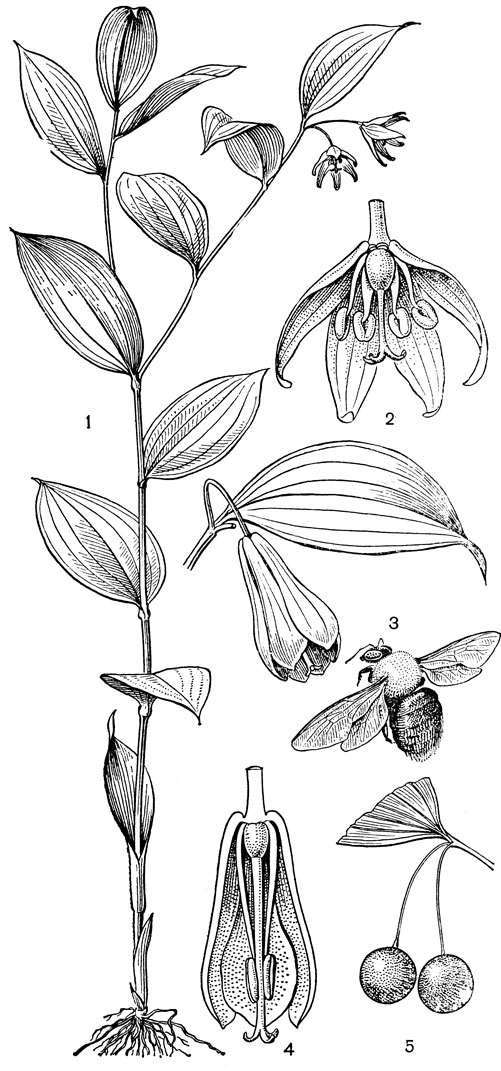
Рис. 84. Диспорум (Disporum). Диспорум зеленоватый (D. viridescens): 1 - общий вид; 2 - цветок. Диспорум сидячий (D. sessile) 3 - цветок с опылителем - пчелой из рода ксилокопа (Xylo сора); 4 - продольный разрез цветка; 5 - плоды
Представители рода стрептопус (Streptopus), объединяющего 9 видов, обитают, как и многие купеновые, в Гималаях, Восточной Азии и Северной Америке, а также в умеренной зоне Средней и Южной Европы. Стрептопусы - многолетние травы высотой от 10 до 120 см с мало разветвленным облиственным стеблем. Листья многочисленные, сидячие или стебле-объемлющие, от яйцевидных до продолговато-ланцетных, с острой верхушкой и густой сетью жилок. Цветки расположены по 1-2 в пазухах стеблевых листьев на тонких, у середины более или менее скрученных цветоножках; иногда они прижаты к расположенному выше междоузлию, так что кажутся сидячими или приросшими к следующему выше расположенному очередному листу. Красный или розовый, реже зеленовато-желтый околоцветник состоит из 6 продолговатых или ланцетных сегментов, свободных или сросшихся. Тычинки в 2-3 раза короче околоцветника, их нити расширены, иногда с шиловидным концом. Столбик нитевидный, с раздельным или почти цельным тупым рыльцем. Завязь 3-гнездная, с многочисленными семязачатками. Плод - многосемянная продолговатая или почти шаровидная красная ягода. Стрептопусы растут в густых сырых лесах и среди кустарников, у ручьев и ключей. Самый большой ареал имеет стрептопус стеблеобъемлющий (S. amplexifolius), произрастающий в Средней и Южной Европе (в Альпах на высоте 750-2300 м), в Восточной Сибири, на Дальнем Востоке СССР, в Японии и в Северной Америке (от Аляски и полуострова Лабрадор до Калифорнии). Корневище у него короткое, покрытое толстыми корнями. Околоцветник воронковидный. Молодые растения съедобны и имеют запах огурцов. В Советском Союзе в Восточной Сибири и на Дальнем Востоке обитает стрептопус стрептопусовидный, или аянский (S. streptopoides), обладающий нитевидным ползучим корневищем, которое ежегодно дает одногодичный стебель. Околоцветник у него звездчатый. В горных лесах Северной Америки широко распространен стрептопус розовый (S. roseus), корневище которого иногда бывает со столонами. Стрептопусы - декоративные растения, они пригодны для озеленения тенистых влажных мест в парках.
Род клинтония (Clintonia), как и два предыдущих рода, характеризуется разорванным ареалом, но в отличие от них имеет центр распространения видов в Северной Америке: здесь встречается 4 вида, а в Гималаях и Восточной Азии - всего 2 очень близких вида, часто объединяемых в один - клинтонию удскую (Clintonia udensis). Клинтонии мезофиты. Это обычные растения темнохвойных, реже смешанных лесов таежных и широколиственных зон; в Гималаях они встречаются на высоте 2500-3800 м. Растения имеют тонкие ползучие корневища, от которых отходят 2-6 приземных широких обратнояйцевидных листьев. Цветки одиночные или собраны в зонтиковидное соцветие или кисть. Воронковидный околоцветник разделен до основания на 6 ланцетных сегментов. Клинтонии - декоративные растения. Белые, как фарфоровые, цветки клинтонии удской и своеобразная форма листьев делают ее очень привлекательной.
В подсемействе ландышевых самый большой род - купена (Polygonatum). В него входит около 50 видов, распространенных в умеренных районах Евразии и Северной Америки, в горных районах субтропиков, реже тропиков Китая и Индокитая. Наибольшее видовое разнообразие купен в Восточной Азии (в Китае 31 вид). В СССР встречаются около 17 видов купен. Для купены характерны симподиальные узловатые корневища с похожими на печати круглыми вдавленными рубцами на местах отмерших годичных побегов (рис. 85, 6) - отсюда второе, народное название рода: "соломонова печать". Стебель у купен очень разной высоты, от 2,5-5 см у гималайско-китайской купены Хукера (P. hookeri, 85, 9) до 2 м и более. Некоторые виды купен с лазающими стеблями. Листья очередные, супротивные или мутовчатые, от линейных до широкоэллиптических, иногда с усиками. Расположенные в пазухах листьев, одиночные или собранные в малоцветковые кисти, цветки белые, желтые, розовые- или фиолетовые, со сросшимися при основании цветоножками. Сегменты околоцветника сросшиеся; околоцветник трубчатый или колокольчатый, лопасти отгиба короче, чем трубка. Тычиночные нити приросшие к околоцветнику до половины своей длины и больше. Пыльники стреловидные, качающиеся, интрорзные. Плод - шаровидная ягода с 1-2 семенами. Цветки купен опыляются главным образом шмелями, пчелами, иногда бабочками. Насекомые посещают их из-за обильного нектара, выделяемого септальными нектарниками.
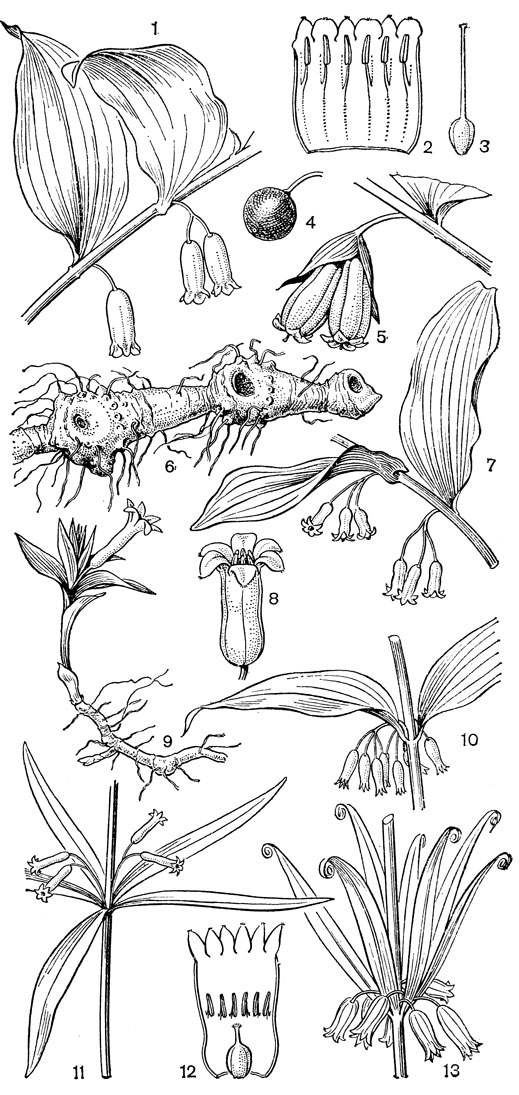
Рис. 85. Купена (Polygonatum). Купена душистая (P. odoratum): 1 - часть соцветия; 2 - развернутый венчик (без гинецея); 3 - гинецей; 4 - плод. Купена обертковая (P. involucratum): 5 - часть соцветия. Купена многоцветковая (P. multiflorum): 6 - корневище. Купена обильноцветковая (P. polyanthemum): 7 - часть соцветия; 8 - цветок. Купена Xукера (P. hookeri): 9 - общий вид. Купена супротивнолистная (P. oppositifolium): 10 - часть соцветия. Купена розовая (P. roseum): 11 - часть соцветия; 12 - развернутый венчик. Купена Кинга (P. kingianum): 13 - часть соцветия
Среди купен с очередными листьями, белыми цветками и темно-синими ягодами наиболее известны широко распространенные евразиатские виды купена душистая, или лекарственная (P. odoratum), и купена многоцветковая (P. multiflorum). Цветки купены душистой, обладающие запахом горького миндаля, очень богаты нектаром, но из-за сравнительно длиннотрубчатого околоцветника он доступен только длинкохоботковым шмелям. Замечено, что у растений, произрастающих в Альпах, трубка околоцветника внизу часто бывает прокушена шмелями, которые похищают нектар, а в проделан-ные отверстия за нектаром проникают и другие насекомые. Иногда в соцветиях купены встречаются цветки с неразвитым гинецеем, со столбиком вдвое короче, чем у нормальных цветков (функционально мужские цветки), а иногда и совсем без гинецея (мужские цветки). У купены многоцветковой цветки собраны в пазухах листьев по 2-5, у них менее длинная трубка, чем у купены душистой. Они опыляются главным образом шмелями, но, возможно, и пчелами, а в отдельных случаях и маленькими бабочками. Трубка околоцветника с перетяжкой, с зеленоватыми отогнутыми на концах опушенными зубцами. Цветок гомогамный (одновременно созревают пыльники и рыльце). Вход в трубку цветка закрывают 3-лопастное рыльце и пыльники. Нити тычинок опушены узловатыми волосками. Рыльце расположено немного ниже пыльников, а цветки на поникших цветоножках направлены вниз, поэтому пыльца сначала не попадает на рыльце. Шмели, просовывая головку в расширенную часть трубки, касаются рыльца и пыльников, вскрывающихся интрорзно, противоположными сторонами своего хоботка и собирают нектар, который находится на дне трубки венчика. Перелетая затем на другой цветок, они производят перекрестное опыление. Вполне возможно и самоопыление, если только рыльце придвинется к близко расположенным пыльникам.
Купены с мутовчатыми листьями имеют белые или розовые цветки с трубчатым околоцветником. У купены мутовчатой (P. verticillatum), распространенной от Европы до Китая и обитающей в лесах и на лугах от равнины до альпийского пояса, околоцветник сравнительно короткотрубчатый, в основании богатый нектаром. Цветки белые, с отогнутыми зеленоватыми лопастями отгиба, опыляются как шмелями, так и короткохоботковыми пчелами и отдельными маленькими бабочками. Легко может происходить и самоопыление. К мутовчатолистным относится и купена розовая (P. rosenm, рис. 85, 11, 12) с небольшим трубчатым розовым околоцветником и красными ягодами. Она встречается в горах Юго-Западной Сибири, на Памиро-Алае, Тянь-Шане и в Северо-Западном Китае. У маленькой высокогорной купены Хукера (P. hookeri) со скученными супротивными или очередными листьями (рис. 85, 9) всего один фиолетовый цветок. Ягода у нее красная.
Купены используют как декоративные растения благодаря изящной форме их стеблей, большим зеленым листьям, приятным белым или розовым цветкам, а позднее - крупным темно-синим или красным ягодам. В цветоводстве известны махровые формы купен, купена многоцветковая с розовыми цветками, купены с полосатыми листьями. Большинство видов цветет в конце весны - начале лета. Хорошо растут в культуре все европейские, кавказские, дальневосточные, американские и многие среднеазиатские купены; они обычно нормально переносят довольно суровые зимы Ленинграда. Корневища купены душистой применяют в народной медицине, в них найдены алкалоид гликонин, сердечные гликозиды, сапонины.
Род смилацина (Smilacina), к которому относится около 25 видов, занимает довольно обширный ареал, от Гималаев и штата Ассам до Восточной Сибири, советского Дальнего Востока, Японии, континентального Китая, Тайваня, Вьетнама и Малезии и от Канады до гор Мексики и Центральной Америки (Гватемала). Большинство смилацин-гигрофиты. Они растут в сырых или заболоченных хвойных и лиственных лесах и на их опушках, на заболоченных лугах и моховых болотах, главным образом в среднегорном и субальпийском поясах. В Гималаях они распространены до высоты 4000 м над уровнем моря. Смилацины обычно невысокие стройные растения со слегка извилистым в верхней части стеблем, с, продолговато-ланцетными или яйцевидными, обычно сидячими листьями с многочисленными жилками. Белые или пурпурные цветки смилацин обладают легким приятным ароматом. Они обычно обоеполые, реже однополые и двудомные, собраны в верхушечные кисти или метелки (рис. 86). Околоцветники отличаются сранительно узкими звездчато растопыренными сегментами. Только у китайско-тибетского высокогорного вида смилацины Генри (S. henryi) околоцветник трубчатый. Тычинки у всех видов прикреплены при самом основании сегментов; пыльники качающиеся, интрорзные. Завязь почти шаровидная, 2-гнездная, с 2 семязачатками в каждом гнезде, с коротким и тупым 2-, 3-лопастным рыльцем. Ягода шаровидная, с одним или несколькими семенами. Смилацины - декоративные растения. В культуре наиболее распространена одна из самых красивых смилацин - смилацина звездчатая (S. stellata).
К смилацинам очень близок род майник (Maianthemum, рис. 86,10,11), к которому относится всего 3 вида, встречающихся в умеренных областях Евразии и Северной Америки в хвойных и смешанных лесах, среди кустарников и на лугах. У майников тонкое ползучее корневище; в нецветущем состоянии на стебле имеется всего один лист, а цветущее растение развивает 2-3 листа с широкой, глубоко сердцевидно-яйцевидной пластинкой на коротком черешке. Мелкие, белые, приятно пахнущие цветки майника собраны в густую верхушечную кисть. Околоцветник звездчатый, из 4 опадающих сегментов. Тычинок 4. Цветки у евразиатского майника двулистного (М. bifolium) слабо протогиничны. В тот период, когда бутоны только открываются, рыльце уже появилось и восприимчиво, а пыльники еще не вскрылись. Затем вскрываются пыльники, а рыльце еще продолжает функционировать (обоеполая стадия), и наконец рыльца теряют восприимчивость (мужская стадия). В тот момент, когда пыльники уже вскрыты и высыпается пыльца, а рыльце еще не завяло, возможна контактная автогамия под действием маленьких насекомых (или других факторов). Она облегчается почти вертикальным положением цветков и расположением пыльников выше рыльца. Ягоды у майников мелкие, светло-красные, с 1-2 семенами. У майника двулистного ягоды вишнево-красные, их поедают рябчики и дрозды. Проходя через желудок, семена сохраняют всхожесть на 80-85% (П. Мюллер, 1934). В качестве декоративных растений майники пригодны для посадки на влажных тенистых участках и на срез для миниатюрных букетов.
Представители трибы ландышевых, состоящей из 4 монотипных родов, хотя и имеют много общего с купеновыми, хорошо отличаются от них безлистным цветоносом с цветками, собранными в верхушечную кисть. Все листья у них приземные. Наиболее известный род этой трибы - ландыш (Convallaria) с одним очень полиморфным видом - ландышем майским (С. majalis). Это растение широко распространено в умеренных и холодных областях северного полушария от Атлантической Европы и Западного Средиземноморья до Японии, полуострова Корея и Северного Китая, а также на юго-востоке Северной Америки. На огромном пространстве своего ареала этот вид дифференцировался на несколько географических рас, которые разными авторами, в зависимости от их взглядов на структуру и объем вида, рассматриваются как разновидности, подвиды и виды. Ландыш произрастает на влажных почвах в хвойных и лиственных лесах и кустарниках, по лесным опушкам и на влажных лугах.
У ландыша довольно сложная система корневищ (рис. 87, 1). Прежде всего мы замечаем длинное, горизонтальное, ползучее, ветвистое корневище с удлиненными междоузлиями. В узлах корневища находятся чешуевидные листья. Из их пазушных почек развиваются новые горизонтальные корневища. Затем заметен небольшой, длиной 1-2 см, почти вертикальный участок корневища с очень укороченными междоузлиями, ежегодно дающий надземный годичный побег. Этот побег состоит из укороченной оси с погруженной в почву верхушечной почкой, 3-5 заметных низовых чешуевидных стеблеобъемлющих листьев с замкнутыми трубчатыми влагалищами (темно-фиолетовой, коричневой или светло-зеленой окраски), 1 ланцетного чешуевидного листа и 2 (реже 3) хорошо всем знакомых блестящих зеленых листьев. В пазухе последнего чешуевидного листа развивается боковой безлистный цветонос. В. неблагоприятные годы ландыш образует побеги с одним зеленым листом. Влагалища всех упомянутых чешуевидных и зеленых листьев, в основании охватывающих друг друга, вместе с осью побега составляют надземный ложный стебель. Образование горизонтальных корневищ происходит с интервалом от 1 года до нескольких лет, в зависимости от различных условий. Вертикальные корневища образуют листья ежегодно, а цветоносы лишь периодически - с перерывами в 2-3 года. Этим в основном объясняется то обстоятельство, что в зарослях ландыша иногда наблюдается много листьев, но мало или совсем отсутствуют цветоносы. Система горизонтальных корневищ продолжает нарастать симподиально за счет пазушных почек корневищ горизонтального или вертикального материнского корневища.
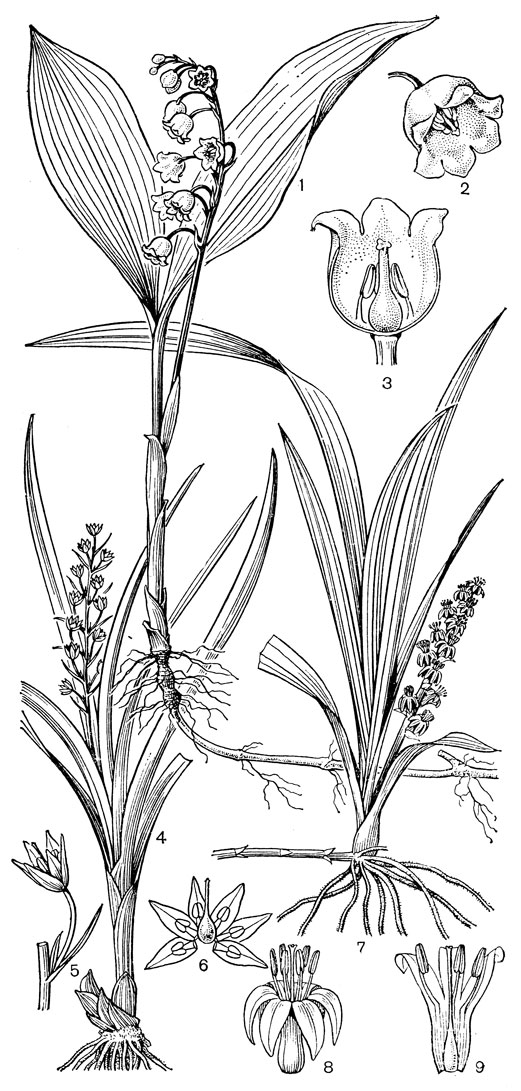
Рис. 87. Спаржевые. Ландыш майский (Convallaria majalis): 1 - общий вид; 2 - цветок; 3 - продольный разрез цветка. Теропогон бледный (Theropogon pallidus): 4 - общий вид; 5 - цветок с прицветником; 6 - раскрытый цветок. Рейнекея мясо-красная (Reineckea carnea): 7 - общий вид; 8 - щеток; 9 - продольный разрез цветка
Ландыш - геофит. На зиму остаются только корневища в почве. С годами горизонтальные корневища перегнивают и их система распадается на отдельные особи, образуя клон. Изучение зарослей ландыша в Закарпатье показало, что средний собственный возраст репродуктивных особей около 9 лет, к 10-12 годам они утрачивают способность к образованию цветков, максимальный возраст - 21 год. Некоторые старые корневища, собранные в Ленинградской области, имели возраст 38-42 года.
Первое цветение ландыша в природе наступает не ранее 7-го года жизни, а в культуре гораздо раньше. Обычно растения зацветают в средней полосе европейской части СССР в конце мая - начале июня. Цветение продолжается 15-20 дней. Трехгранная цветочная стрелка несет на конце кисть из 6-20 поникших цветков на длинных изогнутых вниз цветоножках с пленчатыми прицветниками. Хотя цветоножки отходят с разных сторон стрелки, цветки все же обращены в одну сторону, так как сам стебель закручен спирально. Нижние цветки ландыша распускаются раньше верхних. В цветочной почке острые зубчики (верхушки сегментов околоцветника) плотно прижаты друг к другу, в распустившемся полушаровидно-колокольчатом цветке слегка отгибаются, а при отцветании они сильно загибаются. Тычинок 6; желтые продолговатые пыльники на сравнительно коротких нитях прикреплены при основании сегментов. Столбик короткий, с 3-раздельным рыльцем.
Цветки ландыша слабопротандричны. Они лишены нектарников и привлекают опылителей своим сильным ароматом и пыльцой. Опылители - пчелы, шмели, возможно мухи. Пчела, посещающая ландыш, касается прежде всего рыльца, которое вначале торчит чуть выше тычинок, и если она раньше уже успела вымазаться пыльцой, происходит перекрестное опыление. Затем насекомое, проникшее в цветок, ударяется о пыльники, вскрывающиеся интрорзно продольными щелями (пыльники вскрываются не все сразу, сначала 3 или 4, затем остальные), и обмазывается здесь новой пыльцой. При отсутствии насекомых-опылителей возможно самоопыление. Оно может произойти в тот момент, когда из расположенных немного выше рыльца пыльников осыпается пыльца, а рыльце еще не потеряло способность к восприятию. Так как весь цветок наклонен вниз, падающая пыльца легко может попасть на рыльце. Плод - почти шаровидная оранжево-красная ягода с 2-8 семенами. Семена распространяются птицами. Все растение ядовито.
Ландыш - важное лекарственное растение. Приготовляемые из него препараты укрепляют и регулируют сердечную деятельность. В русскую научную медицину ландыш как лекарственное средство был введен в конце прошлого столетия известным русским врачом С. П. Боткиным. Надземные части ландыша (цветки, листья, семена) содержат около 10 сердечных гликозидов. Цветки ландыша применяют в парфюмерии. Ландыш - декоративное растение, введенное в культуру еще с середины XVI в. Он очень хорош для теневых и полутеневых участков парков, скверов и садов. Садовые формы ландыша крупные, многоцветковые. Известны сорта с розовыми махровыми цветками, а также пестролистные. Во многих странах широко практикуется зимняя выгонка крупноцветковых сортов. Зимующие почки ландыша являются предметом международной торговли.
К ландышу близок монотипный восточно-азиатский род рейнекея (Reineckea, рис. 87, 7-9) - вечнозеленое полустелющееся растение, произрастающее в лесах в нижнем горном поясе и у подножий холмов в Китае (за исключением севера) и на всех островах Японии. У рейнекеи ползучее корневище с двумя рядами чешуй, с несколькими линейными или ланцентными листьями; безлистный цветонос заканчивается кистью светло-розовых цветков. В Японии и Китае, а также в Западной Европе рейнекею выращивают как бордюрное растение.
Во второе подсемейство иглицевых (Ruscoideae) входят 3 довольно специализированных, в основном древнесредиземноморских рода: даная (Danae), иглица (Ruscus) и семела (Semele). Они распространены от Азорских островов до лесных районов южного побережья Каспийского моря. Иглицевые очень своеобразные вечнозеленые кустарники и полукустарники или многолетние травы. Листья у них редуцированные, мелкие, чешуевидные, а филлокладии относительно большие, листовидные. Соцветия состоят из мелких цветков, сидящих на более или менее длинных цветоножках и расположенных на поверхности филлокладиев или в отдельном кистевидном соцветии (даная). Цветки однополые (с остатками редуцированного андроцея или гинецея) или обоеполые, но преимущественно функционирующие как однополые. Тычинок 6 или 3. Они срастаются в колонку, отходящую от основания цветка или от отгиба околоцветника. Пыльцевые зерна относительно более крупные, чем в подсемействе ландышевых. Завязь всегда верхняя, 1-2-, реже 3(неполно) - гнездная, с 2 семязачатками в каждом гнезде. Иногда эти 3 рода выделяют в особое семейство иглицевых (Ruscaceae, Дж. Хатчинсон, 1934, 1959; Т. Накаи, 1936; X. Хубер, 1969; П. Дальгрен, 1980).
Единственный вид рода даная - даная ветвистая (Danae racemosa, рис. 88, 10, 11) - вечнозеленый полукустарник с блестящими косоланцетными филлокладиями. Это в основном гирканский вид, который в третичный период был широко распространен на Кавказе и Малой Азии. Даная произрастает в северной горнолесной части Сирии, в лесной прикаспийской части Ирана и Юго-Восточного Закавказья, а также в отдельных небольших убежищах в восточной части самых южных склонов Большого Кавказа. Даная характерна для горных широколиственных лесов Талыша, где встречается до высоты 1200 м над уровнем моря, но изредка растет также и в низменных лесах. Этот полукустарник селится на скалистых берегах речек и по влажным тенистым обрывам с сочащейся водой, где свисает красивыми густыми шпалерами. Он имеет короткое ползучее горизонтальное корневище, от которого отходят длинные и толстые шнуровидные придаточ-ные корни. На верхушечных ветвях в пазухах филлокладиев расположены небольшие редкие кисти цветков. Белый околоцветник сростнолистный, мясистый, кувшинчатой формы, с 6 маленькими дельтовидными сегментами. Андроцей из 6 тычинок, сросшихся в колонку, по краям которой расположены 2-гнездные пыльники. Гинецей из 3 плодолистиков, не полностью сросшихся и образующих 3-гнездную (в верхней части 1-гнездную) завязь с неполными перегородками; в каждом гнезде 2 семязачатка. Эмбриологические исследования цветков данаи, проведенные Г. Е. Капинос и С. Д. Гусейновой (1971), показали, что они обоеполые только на первых этапах развития, а за тем развиваются как функционально однополые. По внешнему виду соцветия на кустах с функционально женскими цветками отличаются от соцветий на растениях с функционально мужскими цветками меньшим числом цветков и более короткими кистями. В раскрытом женском цветке рыльце находится на уровне пыльников или выше; пыльники существуют долго, хотя их пыльца совершенно стерильна. В мужских цветках рыльце короткое, завязь редуцирована или отсутствует вовсе. Цветет даная в мае - июне, а плоды дает в октябре. Ягоды красные, с 1-2 семенами. Даная - высокодекоративный полукустарник благодаря ветвистым ветвям, блестящим филлокладиям и красным плодам. В культуре известна с 1713 г. Встречается в ботанических садах и в декоративных насаждениях, в нашей стране на Черноморском побережье Кавказа. Данаю используют для изготовления венков, гирлянд. Филлокладии применяют в народной медицине. Вид нуждается в охране и в СССР занесен в "Красную книгу" (1975).
Карта 4. Ареалы родов иглица, даная и семела
Центральное место в подсемействе иглицевых принадлежит роду иглица (Ruscus, табл. 21, 2-4), состоящему из 7 видов, ареал которых почти совпадает с ареалом подсемейства. В Восточном Средиземноморье, на Южном берегу Крыма и в Западном Закавказье нередко можно встретить отдельные куртины или сплошные заросли иглицы понтийской (R. ponticus, рис. 88, 8, табл. 21, 2, 5), близкой к широко распространенной в западных частях ареала подсемейства иглице колючей (R. aculeatus). Это светолюбивый ксеромезофитный кустарничек с ветвящимися стеблями и небольшими кожистыми продолговато-яйцевидными или ланцетными филлокладиями, оттянутыми на верхушке в длинные остроконечия. Иглица понтийская обитает под пологом разреженных лесов или в кустарниках, нередко на скалах и песчаных местах. На Черноморском побережье и в прилегающих к нему районах с высокой влажностью и мягким температурным режимом произрастает иглица колхидская (R. colchicus, рис. 88, 1-7), близкая к западносредиземноморской иглице подъязычной (R. hypoglossum). Иглица колхидская самая мезофильная из всех иглиц. У нее неветвящиеся стебли и сравнительно крупные, обычно широкояйцевидные, тупые филлокладии. Иглица колхидская растет главным образом маленькими группами в тенистых ущельях под густым пологом смешанных и лиственных лесов, а также в самшитовых рощах, иногда под пологом кавказской пихты.
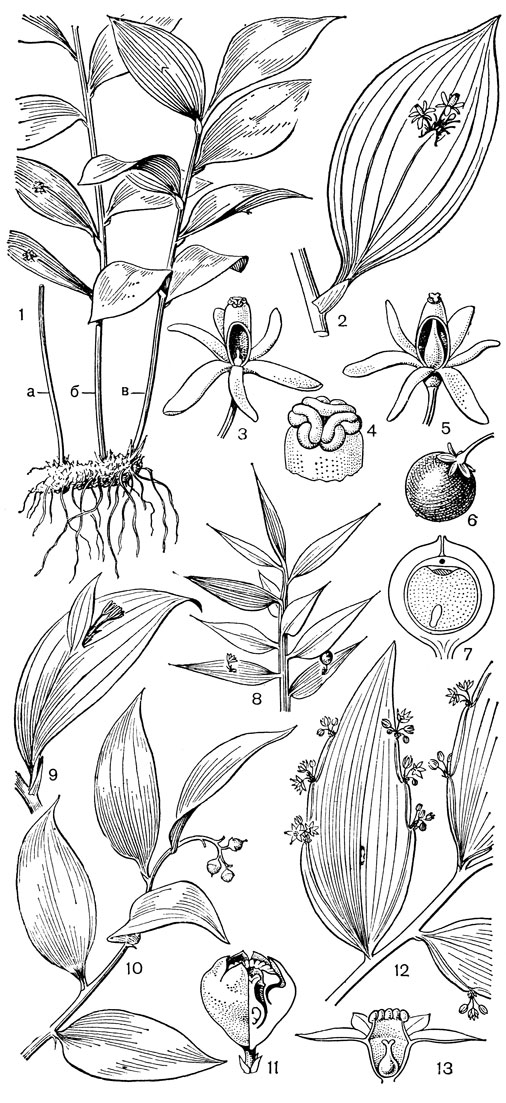
Рис. 88. Спаржевые. Иглица колхидская (Rusous colchicus): 1 - общий вид трехлетнего растения (а - побег 3-го года, б - побег 2-го года, в - побег 1-го года); 2 - филлокладий в пазухе чешуйчатого листа; 3 - мужской цветок; 4 - пыльники и верхняя часть трубки; 5. - женский цветок; 6 - плод; 7 - разрез плода. Иглица понтийская (R. ponticus): 8 - часть ветки. Иглица подъязычная (R. hypoglossum): 9 - филлокладий в пазухе чешуйчатого листа. Даная ветвистая (Dаnаё racemosa): 10 - часть ветки с филлокладиями и плодами; 11 - цветок, часть околоцветника удалена. Семела двуполая (Semele androgyna): 12 - часть ветки с цветками; 13 - цветок в продольном разрезе
При знакомстве с видами иглицы возникает вопрос: что они - кустарники, полукустарники или многолетние травы? Обычно принято относить иглицы к полукустарникам на том основании, что надземные побеги у них отмирают не ежегодно. Однако, как показали наблюдения М. И. Савченко и А. А. Дмитриевой (1962) в Батумском ботаническом саду, надземные стебли иглицы колхидской многолетних междоузлий не образуют (один из важнейших признаков для полукустарников), так как побеги возникают и отмирают непосредственно от корневища. К тому же рост надземного побега полностью завершается в первый вегетационный сезон. Развитие у этого вида протекает следующим образом. Ранней весной на поверхности почвы появляются почти бесцветные травянистые побеги, при основании которых имеются 2 наружные короткие и 3 внутренние более длинные чешуи. Из пазух маленьких листьев выходят почти бесцветные филлокладии, сначала тесно прилегающие к стеблю. Большинство листьев скоро засыхает и отмирает, часть остается в виде чешуй. По мере того как побег в течение весны растет, филлокладии зеленеют и становятся кожистыми. Самые нижние филлокладии располагаются мутовчато или супротивно, остальные очередно; филлокладий имеет хорошо выраженную главную жилку и 1-2 пары боковых. К концу лета начинается развитие соцветия, расположенного на нижней стороне в средней части филлокладия, в пазухе мелкого прицветного листа. Соцветия не развиваются только у самого верхнего и нижнего филлокладиев. Цветки распускаются постепенно в течение осени и зимы, а весной созревают плоды. Цветение заканчивается в конце лета, перед началом зацветания других молодых побегов, образовавшихся от корневища. Если смотреть на куст, то создается впечатление непрерывности цветения и плодоношения иглицы. На третий вегетационный период побег начинает буреть и от филлокладия остается лишь остов с сетью жилок. Следовательно, иглица колхидская ближе к многолетним травам. Распускание цветков идет от основания к верхушке. Цветки иглиц закладываются в бутонах как потенциально обоеполые, а затем развиваются как функционально однополые. Цветоножка у мужских цветков сравнительно длинная, а у женских почти равная цветку. Околоцветник 3-членный, раздельный, сегменты его внешнего круга продолговато-обратнояйцевидные, зеленые, внутреннего - ланцетные, зеленовато-фиолетовые. В функционально мужском цветке (рис. 88, 3, 4) андроцей представлен тычиночной колонкой (тычинки и пыльники, сросшиеся полностью); пыльники вскрываются поперечно. В функционально женском цветке гинецей состоит из крупной сидячей завязи с головчатым блестящим липким рыльцем; тычиночная колонка с деформированными пыльниками. Цветение и опыление иглиц изучено только у иглицы понтийской в Крыму, на склоне горы Аюдаг в дубово-можжевеловом лесу. Зацветает иглица понтийская осенью, в сентябре - октябре, и цветет всю зиму, до апреля, т. е. во время самой повышенной влажности воздуха и обилия дождей. Обычно отдельные цветки функционируют 8-10 суток. При выпадении дождя, росы или таянии снега поток или капли воды, обтекающие функционально мужские цветки, почти всегда содержат пыльцу. Брызги с мужских побегов могут попасть на филлокладии побегов с женскими цветками, на которых остается часть пыльцы, что и обеспечивает гидрогамное опыление. При относительно низкой влажности секреторная капля твердеет и рыльце становится неактивным. Семена иглицы колхидской могут прорастать только через 1,5-2 года.
Иглицы - декоративные вечнозеленые растения, пригодные для устройства бордюров в тенистых садах и парках с теплым климатом. В Англии иглицу колючую называют "метла мясника" (butcher's broom) из-за того, что мясники использовали ее в качестве метлы. Молодые побеги иглицы съедобны, а иглица колхидская представляет интерес в хозяйственном отношении - местное население заготавливает ее на корм скоту. Некоторые виды иглицы имеют разнообразное применение в народной медицине; ягоды их съедобны; семена находят употребление как суррогат кофе, а иногда их используют для изготовления бус.
По характеру расположения соцветий на филлокладиях к иглице довольно близок род семела (Semele), однако по общему строению цветка он имеет большее сходство с родом даная. В роде семела 5 видов, все они - эндемики Макаронезийской флористической области (большинство встречается только на острове Мадейра). Это высокие, вьющиеся кустарники, отличающиеся большим разнообразием филлокладиев. Цветки на коротких цветоножках, собраны большей частью пучками, расположены либо по краю филлокладиев, либо посередине, иногда сосредоточены около верхушки. Беловато-желтоватый околоцветник из 6 сегментов, сросшихся более чем наполовину в чашевидную трубку, по отгибу которой расположены 6 сросшихся тычинок. Столбик короткий, с 3 толстыми рыльцами; завязь 3-гнезднаяв У большинства видов цветки (рис. 88,13) кажутся обоеполыми, но при внимательном рассмотрении видно, что в них полностью развит либо гинецей, либо андроцей (их можно так же, как и цветки данаи, отнести к функционально мужским или функционально женским). Виды семелы цветут с мая по июль, некоторые до августа, а плодоносят с января по март, часть видов - до июня. Ягода красноватая или желто- красная, семена блестящие, бледно-желтые, почти шаровидные. Семела принадлежит к числу декоративных кустарников. Наиболее известна в культуре семела двуполая (S. androgynа, рис. 88, 12, 13).
С предыдущим подсемейством иглицевых тесно связано подсемейство собственно спаржевых (Asparagoideae), в состав которого входит 1 род спаржа (Asparagus), насчитывающий около 300 видов. Это самый большой род в семействе, широко распространенный в Старом Свете" Основное разнообразие видов сосредоточено в Африке; в Евразии спаржи особенно многочисленны в Средиземноморье и в Западной и Средней Азии, а также в Китае (24 вида). Представители рода спаржа - многолетние травы, полукустарники или лианы. Они характеризуются, подобно иглицевым, филлокладиями. Цветки спаржи маленькие, обоеполые, однополые или полигамные. Околоцветник состоит из 6 сегментов, почти свободных или более или менее сросшихся при основании, остающихся или опадающих; он может быть чашевидно-колокольчатый, или почти шаровидный, или воронковидный,- иногда сегменты отстоящие (венчик звездообразный). Тычинок 6; для собственно спаржевых особенно характерно, что тычинки между собой не срастаются; нити в разной степени прикреплены к основаниям сегментов околоцветника, они сравнительно тонкие или слегка расширенные; пыльники у спаржи крупные, оранжево-красные или красные. Завязь 3-гнездная, с 2 или более семязачатками в каждом гнезде. Имеются септальные железки. Евразиатские виды опыляются шмелями.
Большой род спаржа подразделяется на 3 довольно хорошо очерченных подрода. Наиболее многочисленным и своеобразным является подрод собственно спаржа, или аспарагус (Asparagus), виды которого произрастают как в засушливых, так и во влажных областях. Филлокладии у них линейные или шиловидные,, цветки двудомные. Все встречающиеся в нашей стране виды спаржи относятся к этому подроду. Сюда входят широко распространенная у нас на Кавказе, в европейской части и в Средней Азии (реже в Сибири) спаржа мутовчатая (A. verticillatus, рис. 89); на запад она доходит до островов Макаронезийской флористической области. Очень интересна спаржа туркестанская (A. turkestanicus) - эндемик среднеазиатских пустынь, обитающая на песках и щебнистой почве. У этого вида филлокладии отсутствуют. Из представителей этого подрода наибольшее значение имеет спаржа лекарственная (A. officinalis), произрастающая на лугах, в северных степях и в зарослях кустарников почти по всей Европе, на Кавказе и в Восточной Азии.
Многочисленные виды спаржи с обоеполыми цветками и шиловидными или линейными филлокладиями образуют подрод аспарагопсис (Asparagopsis). Несколько видов его широко культивируют в качестве декоративных растений. Отметим спаржу щетинистую (A. setaceus), известную в культуре под названием спаржи перистой (A. plumosus). Это изящный нежный африканский полукустарник, произрастающий во влажных тропических и субтропических лесах и в кустарниках по долинам рек, а также обитающий в саваннах и полусаваннах. Во влажных субтропиках Южной Африки встречается спаржа густоцветковая (A. densiflorus),, называемая в цветоводстве спаржей Шпренгера (A. sprengeri). Это одно из самых распространенных комнатных растений. Интересный вид - спаржа серповидная (A. falcatus) с длинным (до 15 м) лазающим и довольно колючим стеблем; обитает в Южной и Восточной Африке и в Юго-Восточной Азии, а также на острове Шри-Ланка; используется для декоративного оформления. У эндемика Мадагаскара спаржи Пуассона (A. poissonii, рис. 89, 10) имеются прямые ко-лючки, превышающие пучки филлокладиев на ветвях и особенно на стеблях.
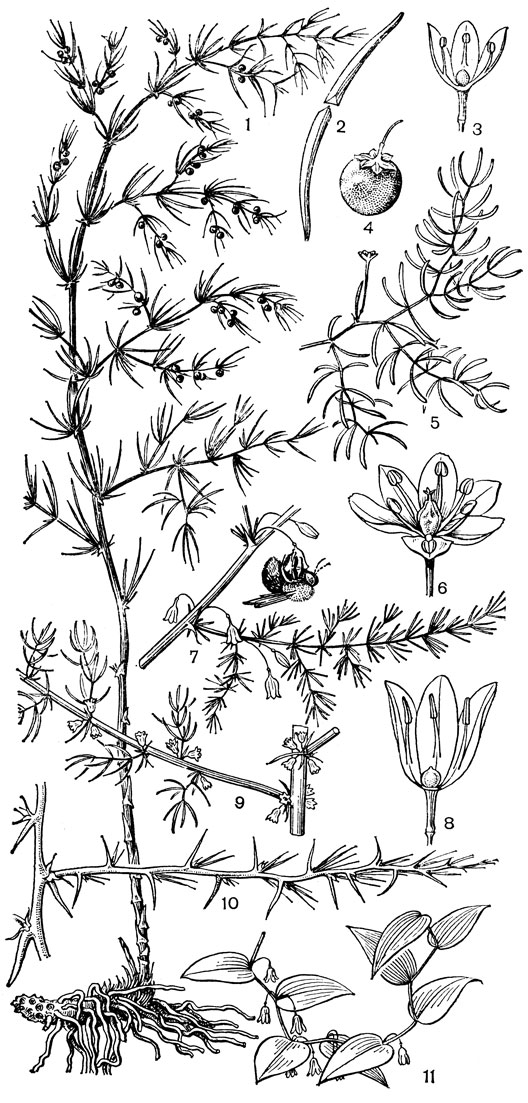
Рис. 89. Спаржа (Asparagus). Спаржа мутовчатая (A. verticillatus): 1 - общий вид растения с плодами; 2 - филлокладий; поперечный разрез; 3 - продольный разрез мужского цветка; 4 - плод. Спаржа цепляющаяся (A. scandens): 5 - часть растения; 6 - цветок. Спаржа прибрежная (A. litoralis): 7 - часть растения и опылитель - шмель; 8 - продольный разрез мужского цветка. Спаржа шобериевидная (А. schoberioides): 9 - часть растения с цветками. Спаржа Пуассона (A. poissonii): 10 - часть растения. Спаржа спаржевидная (A. asparagoides): 11 - часть растения
К подроду мирзифиллум (Myrsiphyllum) относятся 6 близких видов, обитающих в Южной Африке. Это ветвистые лианы с обоеполыми цветками и нежными листовидными плоскими филлокладиями, расположенными по одному в пазухах кроющих чешуй. При основании филлокладиев находятся сочленения, благодаря которым они легко осыпаются при засыхании. К этому подроду относится распространенная в культуре многолетняя спаржа спаржевидная (A. asparagoides, рис. 89, 11), растущая в Южной Африке в сухих полусаваннах и саваннах; встречается она и во влажных лесах южного побережья и в горных лесах до высоты 1300 м над уровнем моря.
Спаржа лекарственная - самый известный представитель рода. Как овощное растение ее культивируют на всех континентах, но особенно в Западной Европе и Северной Америке. В пищу употребляют еще не вышедшие на поверхность и поэтому еще белые и сочные побеги. Их отваривают в соленой воде и обжаривают в масле. Приготовленная таким образом спаржа является деликатесом. Помимо высоких вкусовых качеств, она содержит 1,6-1,7% белков, главным образом тех, в состав которых входит очень нужная для организма аминокислота лизин. Существует более 100 сортов спаржи, различающихся окраской верхушек съедобных молодых побегов, скороспелостью и другими признаками. Впервые в культуру спаржа лекарственная, по-видимому, была введена в Средиземноморье задолго до нашей эры. С конца XV в. спаржу начали выращивать во Франции, а затем и в других европейских странах. Хорошими вкусовыми качествами отличаются и побеги некоторых других видов, в частности средиземноморской спаржи остролистной (A. acutifolius), употреблявшейся в пищу еще в Древней Греции и до сих пор используемой как овощное и лекарственное растение в Италии и других странах. В Японии выращивают спаржу кохинхинскую (A. cochinchinensis) для пищевых целей (из корневищ приготавливают конфеты).
Употребление спаржи как лекарственного растения также относится к глубокой древности, о ней есть упоминания еще у Гиппократа. Многие виды спаржи используют в народной медицине в Южной Африке и других странах.
Семейство драценовые (Dracaenaceae) (Л. И. Иванина)
В семействе драценовых 9 родов и около 250 видов, распространенных в тропиках и субтропиках обоих полушарий и в умеренном поясе южного полушария. В Старом Свете драценовые встречаются главным образом в Африке, Юго-Восточной Азии и Новой Зеландии. Северная граница ареала в Азии доходит до Восточных Гималаев и Южного Китая (Юньнань и остров Тайвань); южная граница проходит через Новую Зеландию и прилегающие к ней южные острова Стьюарт, Окленд и Кэмпбелл. В Северной Америке представители двух родов - нолина (Nolina) и дазилирион (Dasylirion) - произрастают в аридных областях Мексики и на юге США (самая северная точка находится около 38° с. ш.). Чрезвычайно бедно представлены драценовые в Центральной Америке (драцена американская - Dracaena americana и несколько видов нолины) и в Вест-Индии (1 вид - драцена кубинская - D. cubensis). В Южной Америке ареал семейства разорван: в Бразилии встречается кордилина драценовидная (Cordyline dracaenoides); на юге континента, от провинции Вальдивия (около 40° ю. ш.), в Андах и на островах Чилоэ и Чонос, почти до Магелланова пролива и на Фолклендских островах обитает 1 вид - астелия карликовая (Astelia pumila).
Драценовые растут в самых различных местообитаниях - в дождевых и переменно-влажных тропических и сухих субтропических лесах, в редколесьях, на их опушках, в саваннах, жарких пустынях, вблизи морских побережий и в горах до субальпийского и альпийского поясов, а также встречаются на болотах.
Большинство драценовых - вечнозеленые древовидные растения, стебли которых имеют вторичный рост, происходящий за счет деятельности клеток меристематической зоны, расположенной по периферии ствола (а не за счет камбия, как у двудольных). Обычно это невысокие стройные деревья, но есть среди них и крупные деревья со стволами до 8 м в обхвате. Стволы большинства древесных растений совсем не ветвятся (верхушечная почка продолжает рост в течение всей жизни), но встречаются виды и с разветвленными в верхней части стволами. У таких растений верхушечная почка прекращает свой рост и заменяется двумя или несколькими боковыми почками. На концах стволов или их разветвлений драценовые несут пучки листьев (так называемые верхушечные розетки). Среди кустарников изредка встречаются эпифиты. Так, на островах Реюньон и Маврикий на стволах деревьев обитает кустарниковая кордилина маврикийская (Cordyline mauritiana, рис. 91). Наиболее обычны эпифиты среди многолетних вечнозеленых трав из подсемейства астелиевых (Astelioideae). Их длинные серебристые листья и крупные опушенные соцветия живописно свешиваются со стволов деревьев и скал в лесах островов Южной Азии и Новой Зеландии. Астелия карликовая на Фолклендских островах образует особую группировку, состоящую из зарослей низких плоских подушек или плотных ковров.
Листья у большинства драценовых кожистые, жесткие, иногда шершавые, у многих видов дазилириона по краю с крепкими колючками, мечевидные, часто линейные, реже продолговато-эллиптические и ланцетные, обычно плоские, сидячие или на черешках, при основании сильно расширенных. Они спирально расположенные и часто собранные пучками (розетками). У некоторых видов драцены чешуевидные листья сильно развиты, похожи на обычные и чередуются с ними. Листья у многих драценовых с выступающей центральной жилкой и многочисленными боковыми (до 40-50 на каждой половине листа), у некоторых кордилин жилкование напоминает перистое. У травянистых растений листья большей частью линейные, иногда продолговато-яйцевидные, у астелии с 3 главными жилками, сидячие, собранные в прикорневую розетку, остатки же старых оснований листьев плотно одевают верхнюю часть корневища. Сансевъеры (Sansevieria) имеют листья очень мясистые, от плоских до цилиндрических.
Цветки многочисленные, у большинства видов на цветоножках, нередко членистых, с 1 прицветником или прицветничком, иногда с 2 прицветниками и 1 прицветничком, собраны в верхушечные или пазушные, метельчатые, кистевидные или головчатые соцветия. У древесных растений соцветия бывают очень крупные, достигающие в длину 6 м, у травянистых растений они от сравнительно крупных до мелких, иногда редуцированных до одного цветка. Цветки мелкие или среднего размера, большей частью белые, кремовые или зеленоватые, лиловые или красноватые, актиноморфные, обоеполые или однополые. Околоцветник состоит из 6 сегментов, сросшихся в короткую трубку, достигающую половины их длины, или свободных (как у представителей трибы нолиновых - Nolineae). Тычинок 6, большей частью свободных; пыльники интрорзные, прикреплены к нити спинкой, открываются продольными щелями. У многих представителей семейства имеются септальные нектарники. Гинецей синкарпный, редко паракарпный; столбик нитевидный или короткий, утолщенный; рыльце 3-лопастное или почти головчатое. Завязь верхняя, у большинства видов 3-гнездная, но в роде дазилирион и у некоторых видов астелии 1-гнездная с париетальной плацентацией. Семязачатки анатропные, в каждом гнезде от 1 до 20 или 3 в 1-гнездной завязи. Плод - оранжевая, красная или голубая ягода, или плод сухой, нераскрывающийся, с сильно развитыми спинными крыльями, у миллигании (Milligania) - локулицидная коробочка. Семена от светло-коричневых до черных, в очертании продолговатые, изогнутые или яйцевидные, призматические, многогранные или округлые, с носиком или без него, иногда покров семени студенистый или мясистый, у видов коллоспермума (Collospermum) семена бороздчатые, покрытые длинными волосками, отходящими от семяножки.
Драценовые разделяются на 2 подсемейства: астелиевые (Astelioideae) и собственно драценовые (Dracaenoideae). В состав подсемейства астелиевых входит всего одна небольшая триба собственно астелиевых (Astelieae) с 3 родами.
Астелиевые значительно отличаются от всех остальных драценовых, и еще Б.Ш.Ж. Дюмортьв выделил их в отдельное семейство, признаваемое и некоторыми современными систематиками (Р. Дальгрен, 1981). Это многолетние вечнозеленые кустистые травы, образующие плотные дерновины, обычно с коротким корневищем, с многочисленными, преимущественно линейными листьями, собранными в розетку, и с более или менее высоким метельчатым соцветием. Они отличаются необычным серебристым опушением, образованным особыми чешуевидными волосками.
В роде астелия (Astelia) 25 видов, встречающихся в трех флористических царствах: в Палеотропическом (главным образом на островах Тихого океана и на Маскаренских островах), в Австралийском (в Северо-Восточно-Австралийской области) и в Голантарктическом (в Новозеландской и Чилийско-Патагонской областях). На островах, расположенных ближе к экватору, виды астелии обитают в высокогорьях, а на отдаленных от экватора островах - в нижнем поясе гор и на побережье. Так, на Новой Гвинее астелия альпийская (A. alpina, рис. 90) произрастает в субальпийских и альпийских кочкарниках, иногда сильно заболоченных, на высоте 3000-4500 м над уровнем моря. На острове Новая Каледония на склонах горы Гумбольдта встречается сравнительно крупное растение астелия новокаледонская (A. neocaledonica), обычно ведущая себя как эпифит, но здесь становящаяся наземным. Центром видообразования рода, очевидно, являются острова Новой Зеландии. Здесь произрастает 13 видов астелий (принадлежащих ко всем трем подродам), и все они - эндемики. Среди них астелия Соландера (A. solanderi, рис. 90) - сравнительно крупное растение с большим соцветием и листьями длиной до 2 м. Она цроизрастает как эпифит в нижнем влажном поясе в подокарпово-широколиственных лесах и как наземное растение в грасслендах. Густые дерновины очень маленького растения астелии линейной (A. linearis) образуют мощный плотный торф на болотах в горах Новой Зеландии, а также на островах Стьюарт и Окленд. Цветки астелии преимущественно однополые, душистые, с 3 септальными нектарниками. Мужские цветки немного больше женских, их околоцветник с короткой трубкой и 6 ланцетными или дельтовидными сегментами и 6 тычинками; пыльники продолговатые, прикрепленные спинкой, скоро опадающие. В мужских цветках обычно хорошо развита завязь, в которой заметны семязачатки. В женских цветках пыльники маленькие, без пыльцы. После цветения большинство мужских цветков засыхает, но на нескольких цветках семязачатки разрастаются и окрашиваются, цветоножки утолщаются, как у женских цветков. Таким образом, в некоторых соцветиях, внешне мало отличающихся от нормальных мужских соцветий, имеется значительное число функционально обоеполых цветков, которые дают потом полноценные семена. Растения, культивируемые на участках, как отмечают Люси Мур и Элизабет Эдгар (1970), своим сильным запахом привлекают медоносных пчел и крупных мух. Если цветущие мужские и женские растения находились на участке на расстоянии всего нескольких метров друг от друга, то семена у растений нормально завязывались, затем легко прорастали и давали хорошее поколение. Там, где производилась изоляция растений, семена не развивались совсем. Искусственное опыление давало хорошие результаты по количеству плодов и семян.

Рис. 90. Драценовые. Астелия альпийокая (Astelia alpina): l - общий вид; 2 - мужской цветок; 3 - женский цветок; 4 - семя; 5 - чешуйка (с нижней поверхности листа). Астелия Соландерa (A. solanderi): 6 - мужской цветок; 7 - женский цветок; 8 - поперечный разрез завязи; 9 - молодой плод; 10 - семя. Астелия жилковатая (A. nervosa): 11 - женский цветок; 12 - поперечный разрез завязи; 13 - семя. Астелия карликовая (A. pumila): 14 - мужской цветок; 15 - женский цветок; 16 - поперечный разрез завязи; 17 - плод; 18 - семя. Миллигания длиннолистная (Milligania longifolia): 19 - часть соцветия; 20 - лист; 21 - продольный разрез цветка; 22 - тычинка
Астелии используют в качестве декоративных растений. В культуре наиболее известны астелия Соландера и астелия Банкса (A. banksii).
К трибе астелиевых относится также род миллигания (Milligania, 4 вида) - эндемик Тасмании. Виды миллигании по общему облику и чешуйчато-волосистому опушению растений очень сходны с видами астелии (рис. 90), но отличаются обоеполыми цветками и локулицидными коробочками. Они произрастают на горных лугах и торфяниках.
Второе подсемейство - собственно драценовые (Dracaenoideae) - включает 3 трибы: собственно драценовые (Dracaeneae), нолиновые (Nolineae) и сансевъериевые (Sansevierieae).
Самой примитивной в подсемействе является триба собственно драценовых, заключающая 2 очень близких рода - кордилину и драцену. Для трибы характерны древесные растения, реже встречаются вечнозеленые травянистые многолетники. Цветки большей частью обоеполые. Завязь 3-гнездная. Плод - ягода, у кордилины по мере созревания плод становится сухим. Кордилина и драцена различаются между собой главным образом числом семязачатков в завязи и числом семян в плодах: у кордилины 4-20 (редко 2) семязачатков в гнезде, а у драцены в каждом гнезде лишь 1 семя-зачаток.
В роде кордилина (Cordyline) около 15 видов, большинство которых распространено в тропических и субтропических областях Старого Света. Их ареал тянется здесь от Восточных Гималаев, Индии и Китая (остров Хайнань) через Малезию и Полинезию, острова Фиджи и Новая Каледония до Австралия, Новой Зеландии и острова Стьюарт, а также включает Мадагаскар и Маскаренские острова. Только 1 вид встречается в Новом Свете - в Бразилии.
Белые и розовые цветки кордилины расположены в кистях, собранных в крупные метельчатые соцветия длиной до 2 м. Цветки в кисти распускаются не все сразу: у кордилины пограничной (С. terminalis) сначала нижние, затем средние и наконец верхние. Околоцветник у большинства кордилин сросшийся при основании в короткую трубку. В бутоне весь цветок выглядит как трубчатый. Когда цветок распускается, сегменты его более чем наполовину отгибаются наружу (рис. 91), и тогда хорошо обнажаются длинные окрашенные нити тычинок со сравнительно большими продолговатыми пыльниками, а между ними виден столбик с едва заметным рыльцем. В это время цветки вполне доступны насекомым. Но, как только они отцветают, сегменты околоцветника поднимаются кверху, и такие закрытые цветки становятся похожими на бутоны. Отмечено, что на островах Новой Зеландии кордилина южная (С. australis), развивающая огромные соцветия с тысячами цветков, выделяющими нектар, привлекает многочисленных насекомых (особенно из отряда двукрылых). Кордилина южная, выращенная из семян в Сухуми, начинала цвести на 5-6-й год; бутонизация продолжалась 15-25 дней. Цветки раскрывались в первую половину дня и сохранялись раскрытыми 5-8 дней, а соцветие в целом - от 20 до 30 дней. Их посещали пчелы, мелкие мухи и небольшие осы. Несмотря на обильное цветение,) около половины цветков осыпалось. Созревание плодов проходило около 4-5 месяцев. По сообщению некоторых авторов, жуки также играют большую роль в опылении новозеландских кордилины Банкса (С. banksii) и кордилины южной. Замечено, что распространению семян кордилины южной на островах Новой Зеландии способствуют скворцы обыкновенные (Sturnus vulgaris) и карпофаги новозеландские (Carpophaga novae-zelandiae), которые охотно поедают белые или голубые шаровидные ягоды и расселяют растения по острову. Плоды кордилины нераздельной (С. indivisa) также поедают карпофаги.
Наиболее широко распространена в тропиках Старого Света - от Индии до Австралии - кордилина пограничная (рис. 91), известная также под названием "дерево королей" ("tree of kings"). Это небольшое изящное деревце высотой до 3 м, с темно-кроваво-красными продолговатыми или продолговато-яйцевидными листьями. Его часто культивируют в тропиках и разводят в оранжереях и комнатах. Известны многочисленные декоративные садовые формы этого растения.- Отметим еще австралийские декоративные виды кордилины: кордилину сжатую (С. strict а) - кустарник с многоветвящимися стеблями и лиловыми цветками - и кордилину красную (С. rubra), встречающуюся в садоводствах.
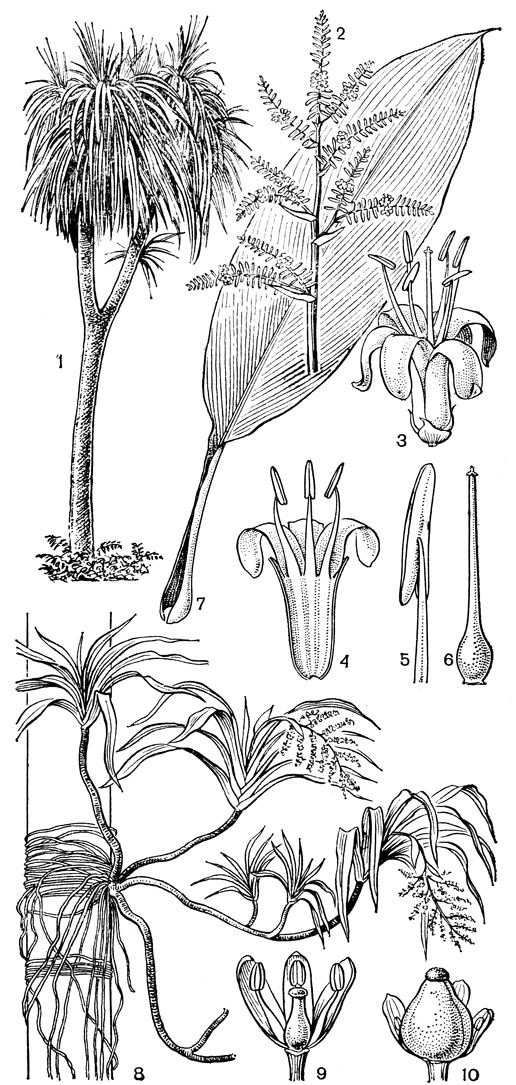
Рис. 91. Кордилина (Cordyline). Кордилина нераздельная (С. indivisa): 1 - общий вид. Кордилина пограничная (С. terminalis): 2 - общий вид; 3 - цветок; 4 - анализ цветка; 5 - тычинка; 6 - гинецей; 7 - лист. Кордилина маврикийская (С. mauritiana): 8 - общий вид; 9 - мужской цветок; 10 - женский цветок
Особого внимания заслуживают новозеландские кордилины. Все 5 видов, произрастающие здесь,- эндемики и известны как декоративные растения. Кордилина нераздельная (рис. 91) - характерный компонент субальпийских нотофагусовых лесов. Ствол ее достигает в высоту 8 м, на вершине разветвленный, с большим (длиной до 160 см) соцветием белых цветков; длинные (до 2 м) золотисто-бронзовые листья собраны пучками на ветках. По свидетельству путешественников, это одно из красивейших деревьев Новой Зеландии. Кордилина нераздельная давно распространена в культуре в Европе, в СССР ее можно встретить на Черноморском побережье Кавказа. Ее листья используют в сельском хозяйстве как подвязочный материал, а из волокон, получаемых из стебля, делают циновки и местную одежду, называемую "той". На открытых сырых равнинах и на опушках субтропического леса обитает кордилина южная, достигающая в высоту 12 и даже 20 м. Ее называют "капустным деревом". Впервые это название было дано Джеймсом Куком в связи с тем, что маорийцы используют в пищу молодые листья кордилины. Как декоративное растение ее разводят во многих странах, в СССР возделывают на Черноморском побережье Кавказа. В листьях кордилины южной более 40% сухой массы составляет волокно, из которого делают щетки, циновки, коврики и другие грубые плетеные изделия. В некоторых районах Грузии листья идут на изготовление влагоустойчивого и крепкого подвязочного материала, идущего главным образом для подвязки виноградных лоз.
Семена кордилины южной и других видов содержат до 33% жирного масла, в котором находится до 90% линолевой кислоты. Корневища кордилины кустарниковой (С. frutex), достигающие массы 4-6 кг, заключают до 20% сахара и их используют в пищу. Отвар корней кордилины сжатой и кордилины пограничной употребляют в народной медицине.
Другой род этой трибы - драцена (Dracaena) - состоит из 80 (по другим данным, от 40 до 150) видов, распространенных в основном в тропиках и субтропиках Старого Света. Большинство драцен произрастает в тропической и Южной Африке и на окружающих ее островах, включая Макаронезию, Мадагаскар и Маскаренские острова. Северная граница ареала проходит здесь через остров Мадейра (около 33° с. ш.), Эфиопию и остров Сокотра. Многие виды рода встречаются в Южной и Юго-Восточной Азии (на север часть видов доходит до южных склонов Восточных Гималаев и тропических районов Китая - до провинции Юньнань и острова Тайвань). Некоторые виды обитают в Новой Гвинее и в Квинсленде. В Новом Свете имеются всего 2-3 вида, произрастающих от Мексики до Коста-Рики, на острове Куба и в Бразилии.
У большинства драцен цветки белые или розовые, раскрывающиеся ночью, с приятным запахом, особенно у драцены душистой (D. fragrans), цветки которой издают аромат свежего сена или меда. Околоцветник достигает в длину 45 мм, сегменты околоцветника большей частью соединены в трубку до 1/3 длины венчика. Цветки выделяют сравнительно много нектара и привлекают различных насекомых - преимущественно ночных бабочек, которые играют основную роль в перекрестном опылении растений.
Среди драцен известны деревья, на стволах которых имеются кроваво-красные пятна смолянистого сока, так называемой "драконовой крови". Еще Диоскориду, известному врачу и ботанику, было известно о существовании на острове Сокотра драконова дерева, которое затем получило название драцены киноварно-красной (D. cinnabari, рис. 92). Это живописное дерево встречается там почти повсюду - на скалах и утесах, преимущественно на высоте 1300-1600 м над уровнем моря (табл. 22). Жители острова Сокотра называют смолистый сок, источаемый деревом, "кровью двух братьев" или "драконовой кровью". Согласно старинной легенде индийцев, дракон пристрастился к слоновой крови. Он обычно обвивался вокруг хобота слона и выпивал всю кровь. Но однажды умирающий слон упал и раздавил дракона, и их кровь перемешалась. Эту смешанную кровь назвали киноварью, а затем так стали называть красную землю, содержащую ртуть.
Драконово дерево Сокотры близкородственно драконовым деревьям Канарских островов, Сомали и Эфиопии. Знаменитое канарское драконово дерево (D. draco, рис. 92) произрастает на острове Мадейра (где сохранились лишь маленькие деревья на недоступных скалах в прибрежной полосе), на островах Тенерифе и Пальма и островах Зеленого Мыса. На Канарских островах оно растет в субтропическом поясе на высоте 100-400 м над уровнем моря и культивируется в долинах. На островах Зеленого Мыса леса почти вырублены и драконовы деревья сохранились преимущественно на склонах гор среди ксерофитной растительности, где старые экземпляры (в возрасте около 100 лет) достигают в высоту всего 5 м. Листья на крупных деревьях, изредка произрастающих в долинах, постоянно срезает местное население. Тревожит факт, пишет Кнут Бистрэм (1960), что молодые драконовы деревья безжалостно вырубают и их осталось очень мало, а на некоторых островах они совсем исчезли. Наиболее знаменитое дерево из окрестностей города Икод на острове Тенерифе погибло от урагана в 1868 г.

Рис. 92. Драцена (Dracaena). Драцена омбет (D. ombet): 1 - общий вид. Драцена киноварно-красная (D. cinnabari): 2 - общий вид; 3 - часть соцветия; 4 - цветок; 5 - продольный разрез цветка; 6 - продольный разрез плода; 7 - семя; 8 - лист. Драконово дерево (D. draco) на острове Тенерифе: 9 - общий вид

Таблица 22. Драцена киноварно-красная (Dracaena cinnabari), остров Сокотра, пик Фиери
Оно достигало в высоту около 23 м, а в обхвате 15 м. Его ствол, суженный к основанию конусом, был внутри совсем пустой (в XV в. там помещался алтарь). Знаменитый ученый-путешественник Александр Гумбольдт, который видел это дерево в 1799 г., предполагал, что ему 6 тыс. лет. Однако в настоящее время на основании наблюдения над ростом деревьев, как сообщает Карл Мегдефрау (1975), исследователи пришли к заключению, что этот возраст был сильно преувеличен и, возможно, самому старому дереву было не более 600 лет. Сейчас чемпионом среди драценовых считается драконово дерево, имеющее в обхвате около 8 м при высоте 21 м. Оно произрастает на том же острове в окрестности города Оротава (возможно, оно было посажено). Его возраст, как предполагают, не более 265 лет.
Цветение драконова дерева наступает в 8-11 лет, а иногда и в 25-30 лет. К этому времени ствол достигает в высоту нескольких метров, листья по всей его длине уже опали, на стволе остаются заметные рубцы от черешков. Цветение происходит не каждый год, а через несколько лет (так, за 185 лет дерево на Тенерифе цвело 11 - 13 раз, т. е. через 14, 2-16, 8 лет). После первого цветения ствол значительно увеличивается в толщину, в среднем дерево прибавляет в год на 2-3 см, рубцы исчезают, и оно начинает ветвиться в верхней части. У старых деревьев образуются воздушные корни и происходит выделение смолистого ярко-красного сока - "драконовой крови", которую на Сокотре и на Канарских островах получают главным образом путем подсочки. Этот сок содержит пигменты дракокармин и дракорубин. Из него получают лак, который идет на покрытие металла. Смолистый сок используют в народной медицине и для подкраски вина. "Драконова кровь" была обнаружена на Канарских островах и в пещерах доисторических людей. Предполагают, что ее применяли для бальзамирования.
Еще одно небольшое африканское драконово дерево, или драцена омбет (D. ombet, рис. 92), высотой до 12 м, произрастает в горах Южного Судана, на высоте 1300-1600 м над уровнем моря. В горах Сомали, почти на таких же высотах, обитает сомалийское драконово дерево,; или драцена расколотая (D. schizantha). Наконец, сравнительно недавно в горах Китая, в провинции Юньнань, было обнаружено, что драцена камбоджийская (D. cambodiana) также выделяет смолистый сок - "драконовую кровь".
Драцены в основном используют как декоративные растения. Большинство видов введено в культуру в прошлом веке из тропической Африки и с Маскаренских островов. Одно из любимых растений, выращиваемых в закрытых помещениях,- драцена душистая, распространенная в тропической Африке от Гвинеи и Сьерра-Леоне до Малави. В комнатах, оранжереях разводят нарядный гвинейский полукустарник с ложномутовчатым расположением листьев - драцену Годсефа (D. godseffiana) и другие виды. Известны разнообразные пестролистные формы тропической африканской драцены деременской (D. deremensis) и южноафриканской драцены из влажных субтропических лесов - драцены Хукера (D. hookeriana). С острова Маврикий введена в культуру драцена закривленная (D. reflexa), названная так за наклоненные вниз листья. Несколько особых разновидностей ее встречаются и в горных лесах Мадагаскара. Крупными цветками отличаются соцветия драцены зонтиконосной (D. umbraculifera), произрастающей на Мадагаскарских островах. На этих же островах обитает и драцена нарядная (D. concinna) с длинными блестящими желто-зелеными волнистыми листьями с красноватым краем.
Вместе с кактусовыми, различными видами юкки и агавы в пустынях, полупустынях и в сухих кустарниковых зарослях Северной Америки можно встретить виды нолины (Nolina), дазилириона (Dasylirion) и калибануса (Calibanus), входящих в состав трибы нолиновых. Нолиновые являются характерными представителями и эндемиками Мадреанской (или Сонорской) флористической области - центра развития ксерофитной североамериканской флоры. Нолиновые - многолетние травы или кустарники, нередко с большим подземным каудексом. У нолины Бигелова (Nolina bigelovii), произрастающей по сухим каменистым и пустынным склонам в Соноре, Западной Аризоне, Сиуатаке и в Южной Калифорнии, каудекс может достигать в ширину 1 м. Суккулентное растение нолина отогнутая (N. recurvata) имеет стебель, вздутый при основании в виде большой луковицы, который служит для сохранения воды; большой стебель может снабжать растение водой даже в течение всего года. Листья у большинства нолиновых несут черты ксерофильного или суккулентного растения. Они большей частью узколинейные, с хорошо развитой кутикулой, содержат много механических тканей, с колючками по краю. Нолиновые хорошо отличаются от драценовых двудомными или полигамно-двудомными цветками, а также очень маленьким кожистым венчиком, сегменты которого почти свободные. У нолины завязь 3-гнездная, с 2 семязачатками в каждом гнезде, а у дазилириона завязь 1-гнездная, с 3 или 6 семязачатками. Плоды этих двух родов приспособлены к распространению ветром. У дазилириона они не раскрывающиеся, односемянные, 3-гранные и у многих видов с 3 крыльями, на членистых плодоножках (плоды отваливаются вследствие отделения плодоножек). При ветре крылатые плоды легко могут передвигаться как по воздуху, так и по поверхности земли. На песке такие плоды меньше засыпаются. Плоды нолины шаровидные, раскрывающиеся, и под действием ветра они легко катятся по земле. Большинство видов дазилириона имеют высокие соцветия, что увеличивает дальность полета их плодов.
В роде нолина около 30 видов, распространенных в основном в Мексике, 9 видов - в южной части США, 1 вид встречается в Гватемале. Северная граница ареала рода проходит на западе США, в южной части горного штата Колорадо, а на юго-востоке - в штате Южная Каролина. Цветки у нолины диаметром 2-4 мм, раскрытые, слабодушистые, по 2-3 в пазухе небольших зубчатых прицветников на веточках 2-го и 3-го порядков, собранные большей частью в почти сидячие метелки или редуцированную кисть. Опыляются насекомыми. У нас на Черноморском побережье Кавказа в парках можно встретить мексиканскую нолину длиннолистную (N. longifolia, рис. 93). В Южной Мексике она произрастает в горах на высоте до 3000 м. Это небольшое вечнозеленое дерево высотой до 3 м с разветвленным на вершине и расширенным при основании стволом.
В роде дазилирион 20 видов. Большинство произрастает совместно с разными видами нолины, но ареал рода в целом более ограничен. В США встречаются только 5 видов. Многие виды растут на Мексиканском нагорье и в штате Коаула. В пустынях Аризоны, где жара летом достигает +52 °С, в формациях креозотового куста произрастает дазилирион Уилера (Dasylirion wheeleri, рис. 93). В районе Соноры и Чихуахуа дазилирион встречается совместно с нолиной мелкоплодной (Nolina microcarpa) и другими видами нолины. Этот вид испытывается в культуре на Южном берегу Крыма. В Центральной Мексике в горных пустынях на высоте 2200-2400 м растет дазилирион кисточковый (Dasylirion acrotrichium) с толстым древовидным стеблем и нередко без видимого ствола. Этот вид уже с начала прошлого столетия разводится в Западной Европе. На Черноморском побережье Кавказа в парках и садах, кроме этого вида, можно встретить дазилирион длиннейший (D. longissimum) из Восточной Мексики, дазилирион сизолистный (D. glaucophyllum) из Центральной Мексики со стволом высотой до 5 м и сизыми тусклыми листьямй, дазилирион злаколистный (D. graminifolium) из Восточной и Центральной Мексики, имеющий невысокий стебель и зеленые блестящие листья. В Северной Америке распространен в культуре еще дазилирион техасский (D. texanum) с небольшим стеблем, но с очень длинным соцветием.

Рис. 93. Драценовые. Дазилирион Уилера (Dasylirion wheeleri): 1 - общий вид; 2 - часть листа; 3 - часть мужского соцветия; 4 - мужской цветок; 5 - сегмент околоцветника с тычинкой; 6 - часть женского соцветия (с плодами); 7 - женский цветок; 8 - плод. Нолина длиннолистная (Nolina longifolia): 9 - общий вид; 10 - часть листа. Сансевьера полукустарниковая (Sansevieria suffruticosa): 11 - общий вид; 12 - поперечный разрез через лист. Сансевьера Роксбурга (S. roxburghiana): 13 - часть соцветия; 14 - анализ цветка; 15 - часть листа; 16 - поперечный разрез листа
В Мексике листья дазилириона и нолины употребляют для плетения корзин, шляп, а иногда и для покрытия крыш. По всей мексиканской территории, где встречаются большие заросли дазилириона, приготовляют из стеблей алкогольный напиток "сотол".
Многолетние, вечнозеленые, травянистые, с большими суккулентными листьями представители рода сансевьера (Sansevieria, около 60 видов в Африке и Азии) выделяются в отдельную трибу сансевьериевых. По строению цветков и плодов этот род чрезвычайно близок к драцене и кордилине. Однако сансевьера резко отличается от родов трибы собственно драценовых по облику и строению семян (по общей форме зародыша и числу слоев внешнего интегумента).
Сансевьеры - корневищные растения с розетками прямостоячих мясистых листьев, достигающих в длину 2 м. Плоские или желобчатые, полуцилиндрические или цилиндрические листья служат им как бы резервуаром, в котором накапливается и сохраняется вода (благодаря богато представленной водозапасающей паренхиме), необходимая для переживания сухого периода в переменно-влажных и сухих тропиках. Для побегов некоторых видов - сансевьеры цилиндрической (S. cylindrica) и сансевьеры полукустарниковой (S. suffruticosa, рис. 93) - характерна резкая дифференциация на две различные части. Каждая часть побега сначала растет в горизонтальном направлении и несет чешуи, а затем изменяет направление на вертикальное и начинает образовывать нормальные листья. Основная ось побега располагается на поверхности земли и несет на себе поочередно то чешуи, то нормальные листья.
Значительное большинство видов распространено в тропической Африке. Они растут в саваннах, иногда образуя куртины под баобабами или на песчаных местах в пустыне Сомали; нередко виды сансевьеры густыми зарослями встречаются по открытым долинам рек рядом с молочаями. В Гвинее на южных склонах возвышенности Фута-Джаллона, рядом с водопадом Кинкон, по берегам рек и на выступах скал можно встретить совершенно бесстебельную, с плоскими листьями, украшенными оригинальным волнистым рисунком сансевьеру трехполосную (S. trifaciata), которую обычно неправильно называют сансевьерой гвинейской (S. guineensis). На Мадагаскаре произрастает небольшая сансевьера самбиранская (S. sambiranensis) с очень коротким соцветием. На скалах и песчаных склонах острова Шри-Ланка распространена сансевьера цейлонская (S. zeylanica). В Бирме и Восточной Индии обитают еще 3 вида.
Цветки у сансевьеры белые или розовато-лиловые, реже зеленые, обычно в пучках, в густом цилиндрическом или головчатом соцветии. Цветонос в нижней части с очередными прицветными листьями или чешуями. Каждый цветок с маленьким прицветничком на короткой цветоножке с сочленением в верхней части. Околоцветник с трубкой и с линейными, прямыми или часто закрученными лопастями, равными трубке или короче ее. На дне трубки цветка скапливается нектар, который выделяется септальными желёзками, находящимися у основания завязи. Кроме септальных нектарников, имеются еще внецветковые, расположенные на ветвях соцветия желёзки, выделяющие сахаросодержащую жидкость. Тычинки длиннее венчика, прикрепленные в нижней части трубки. Завязь 3-гнездная, с 1 семязачатком в каждом гнезде; столбик длинный, рыльце головчатое. Плод - шаровидная ягода, желтая или оранжевая, 1-3-семянная, бородавчатая. Семена сжатошаровидные, твердые. Цветки сансевьеры открываются в конце дня или ночью и опыляются преимущественно длиннохоботковыми насекомыми из отряда перепончатокрылых.
Сансевьера цейлонская и сансевьера трехполосная служат источником ценного технического волокна и культивируются во многих тропических странах. В Африке выращивают для получения волокна еще сансевьеру цилиндрическую, а в Индии - сансевъеру Роксборо (S. roxburghiana, рис. 93). Более 10 видов рода сансевьера (известных в садоводстве под названием "щучий хвост") культивируют как комнатные растения, а в теплых странах - в открытом грунте, где их иногда сажают вдоль изгородей. Наиболее распространены в культуре сансевьера трехполосная и сансевьера цейлонская.
Семейство текофилеевые (Tecophilaeaceae)
В этом семействе 6 родов и около 20 видов. Роды валлерия (Walleria, рис. 94), и цианелла (Cyanella, рис. 95) распространены в Южной и тропической Африке, в то время как роды конантера (Conanthera, рис. 96), текофилея (Tecophilaea, рис. 97) и зефира (Zephyra, рис. 96) эндемичны для Чили. Лишь один род одонтостомум (Odontostomum) растет обособленно от двух основных частей ареала семейства в Калифорнии. Семейство текофилеевые занимает в некоторых отношениях промежуточное положение между мелантиевыми и ирисовыми, но стоит ближе к последним. Некоторые текофилеевые, как текофилея цианокрокус (Tecophilaea cyanocrocus, рис. 97), даже внешне сходны с представителями ирисовых. Семейство это довольно разнородно, и его объем поэтому еще окончательно не установлен.
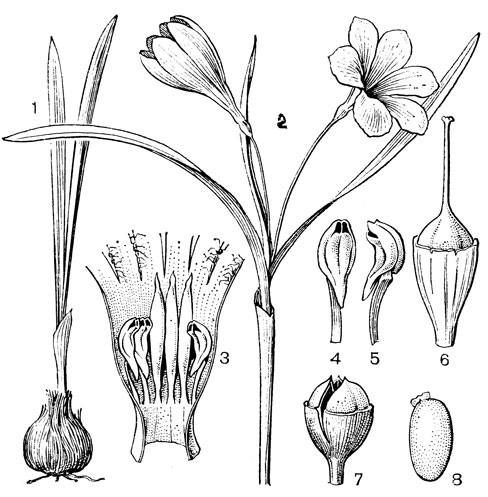
Рис. 97. Текофилея цианокрокус (Tecophilaea суаnоcrocus): 1, 2 - общий вид; 3 - цветок в развернутом виде; 4, 5 - тычинки; 6 - гинецей; 7 - плод; 8 - семя
Текофилеевые - многолетние травы с волокнистыми клубнелуковицами или реже (валлерия) клубнями. Стебли у них однолетние, прямостоячие, иногда колючие, или же они редуцированы, и соцветие возвышается непосредственно над приземными листьями. Листья приземные или реже стеблевые, от 1 до многочисленных, двурядные или в розетках, очень разной формы. Сосуды встречаются только в корнях и характеризуются лестничной перфорацией. Устьица аномоцитные. Цветки обоеполые, актиноморфные или слегка зигоморфные, собраны в простые кисти или в метелки. Сегментов околоцветника 6, обычно более или менее сросшихся. Тычинок 6, приросших к сегментам околоцветника; все тычинки фертильные или иногда 2-3 превращены в стаминодии; нити тычинок иногда более или менее сросшиеся у основания. Пыльники часто с надсвязником, прикреплены основанием, раскрываются верхушечными порами или короткой верхушечной щелью. Пыльцевые зерна обычно однобороздные, иногда с трехлучевой бороздой. Гинецей из 3 плодолистиков, синкарпный, с шиловидным или нитевидным столбиком и маленьким цельным рыльцем; завязь полунижняя, с 2 (одонтостомум) или многими анатропными семязачатками в каждом гнезде. Плод - трехбороздная локулицидная коробочка. Семена многочисленные, более или менее шарообразные или яйцевидные, с крупным зародышем, окруженным мясистым эндоспермом.
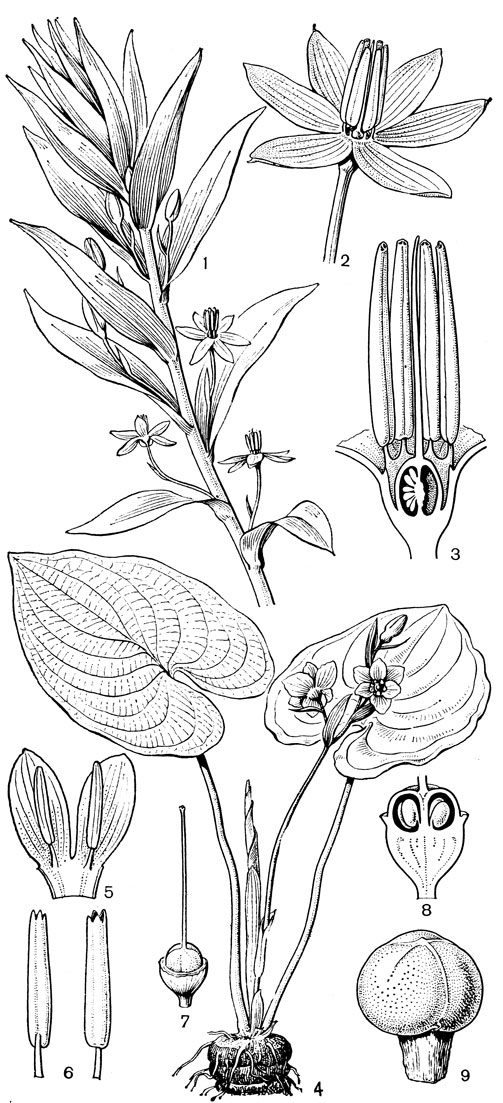
Рис. 94. Текофилеевые и цианастровые. Валлерия Макензи (Walleria mackenzii): 1 - часть растения; 2 - цветок; 3 - продольный разрез завязи. Цианаструм сердцевиднолистный (Cyanastrum cordifolium): 4 - общий вид; 5 - сегменты околоцветника с тычинками; 6 - тычинки; 7 - гинецей; 8 - продольный разрез завязи; 9 - рыльце
Текофилеевые подразделяются на 2 трибы - валлериевые и собственно текофилеевые.
Триба валлериевых (Wallerieae) состоит из единственного рода валлерия (Walleria, рис. 94), насчитывающего всего 3 вида, распространенных в тропической и Южной Африке. Как и все остальные текофилеевые, виды валлерии - типичные геофиты. Но, в отличие от других представителей семейства, подземные запасающие органы которых представляют собой типичные клубнелуковицы, у валлерии это глубокосидящие в почве картофелеподобные клубни, несущие волокнистые корни. От клубней отходят прямостоячие стебли, обычно снабженные крючковидно загнутыми колючими шипами. Листья все стеблевые, очередные, сидячие (у валлерии изящной - W. gracilis - стеблеобъемлющие), от линейных до ланцетных; у валлерии изящной, а иногда также у валлерии поникающей (W. nutans) нижняя сторона листа покрыта небольшими шипами; листья валлерии изящной на верхушке вытянуты в загнутый или спирально закрученный усик. Цветки пазушные, одиночные, на загнутых вниз тонких и длинных цветоножках; изредка в пазухах прицветничков развивается второй цветок. Сегменты околоцветника белые (валлерия изящная) или ярко окрашенные (от розовых до синих), у основания сросшиеся в очень короткую трубку; лопасти околоцветника отогнуты наружу, но после цветения складываются и закрывают завязь. Все 6 тычинок фертильные, приросшие к трубке околоцветника; нити тонкие, очень короткие, у основания сросшиеся; пыльники линейные, у валлерии поникающей и валлерии изящной на верхушке конусообразно сближенные; у всех трех видов валлерии они раскрываются верхушечными порами. Завязь шаровидная, с шиловидным столбиком, заканчивающимся маленьким рыльцем. Семена покрыты более или менее продольными рядами бородавок; на верхушке каждой бородавки сидит пучок очень коротких одноклеточных волосков, чем семена валлерии отличаются от семян всех остальных текофилеевых. Виды валлерии обычно произрастают на песчаной почве в редколесьях, кустарниковых зарослях или же в открытых грасслендах (валлерия изящная). Местные жители клубни валлерии употребляют в пищу как в сыром, так и в печеном виде. Бушмены из жареных клубней валлерии готовят специальную кашу.
В трибу текофилеевых (Tecophilaeeae) входят 5 родов. В отличие от валлерии подземный орган представителей трибы собственно текофилеевых - клубнелуковица.
У южноафриканской цианеллы (рис. 95, 8 видов) глубоко погруженная в почву клубнелуковица образует длинную тонкую шейку. Листья приземные, супротивные или в розетках, образующие в нижней части трубчатое влагалище (наружные влагалища лишены пластинок, часто перепончатые). Цветки у цианеллы в простых или ветвистых кистях, с цветоножками, обычно согнутыми, и с актиноморфным или слегка зигоморфным, чаще ярко окрашенным околоцветником, опадающим вместе с тычинками. Пыльники вскрываются апикальными порами, которые бывают часто утолщены. Столбик шиловидный. Семена яйцевидные, черные, блестящие.
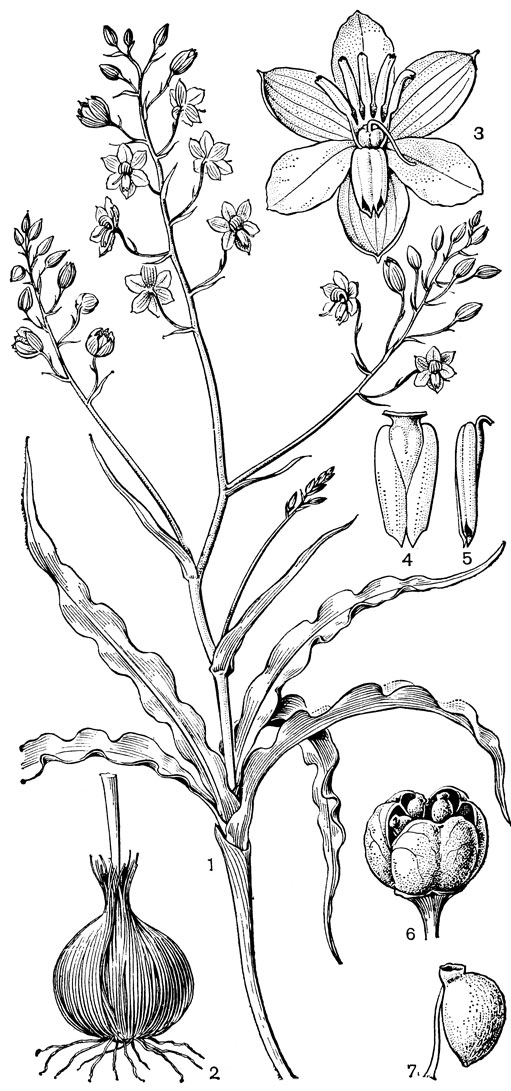
Рис. 95. Цианелла капская (Cyanella capensis): 1 - часть растения; 2 - луковица; 3 - цветок; 4, 5 - тычинки; 6 - плод; 7 - семя
Чилийский род конантера (рис. 96, 5 видов) характеризуется клубнелуковицей, покрытой волокнисто-сетчатой оболочкой, 2-3 ланцетными или линейно-ланцетными приземными листьями, синими цветками в рыхлых кистях, сегментами околоцветника, сросшимися у основания в короткую трубку. Тычинки очень короткие, короче околоцветника и прикреплены к его трубке, все фертильные и одинаковые. Пыльники вскрываются верхушечными порами. Такого же типа клубнелуковица у другого чилийского рода - текофилея (2 вида). Листьев у него несколько или только 1, приземные. Цветок 1 или несколько. Околоцветник колокольчатый, синий. Из 6 тычинок 3 фертильные и 3 превращены в стаминодии. Наконец, третий чилийский род - зефира, состоящий только из 1 вида, имеет такую же клубнелуковицу, как у первых двух видов, приземные линейные листья и синие цветки в кистях. Тычинки очень короткие, прикреплены к зеву трубки околоцветника, причем 3 из них фертильные, а 3 стерильные.
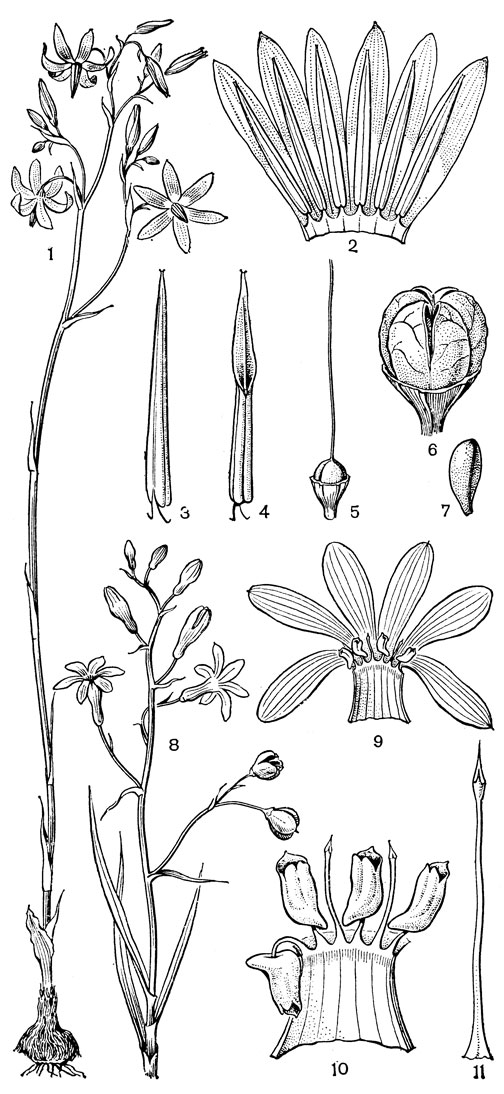
Рис. 96. Текофилеевые. Конантера двулистная (Conanthera bifolia): 1 - общий вид растения; 2 - часть цветка в развернутом виде; 3, 4 - тычинка; 5 - гинецей; 6 - плод; 7 - семя. Зефира прелестная (Zephyra amoena): 8 - часть растения; 9, 10 - часть цветка в развернутом виде; 11 - стерильная тычинка
Последний род в семействе текофилеевых - монотипный калифорнийский род одонтостомум. Клубнелуковицы у него сходны с таковыми чилийских родов, листья линейные, околоцветник с цилиндрической трубкой, снабженной 6 мелкими, узкими, линейными чешуями между тычинками. Тычинки все фертильные и одинаковые, причем нити соединены у основания в трубку. Пыльники вскрываются верхушечными порами. Завязь с 2 семязачатками в каждом гнезде.
Семейство цианастровые (Cyanastraceae) (А. Л. Тахтаджян)
В этом семействе всего один род цианаструм (Cyanastrum, рис. 94, 4-9), насчитывающий 6 видов, обитающих в лесах тропической Африки. Это типичные лесные геофиты с коротким четковидным корневищем, состоящим из толстых клубневидных члеников. Членики сосудов с лестничной перфорацией, приурочены к корням. Листья почти исключительно приземные, с замкнутым влагалищем и более или менее явственно расчлененные на черешок и пластинку; пластинка у некоторых видов ланцетная (цианаструм Буссе - С. bussei) или заостренно-эллиптическая (цианаструм хостолистный - С. hostifolium), но у остальных видов (например, у цианаструма сердцевиднолистного - С. cordifolium, рис. 94, 4-9) пластинка с явственным серцевидным основанием; в зависимости от формы листовой пластинки жилкование от параллельного до дугонервного, причем главные жилки соединены между собой поперечными жилками. Устьица парацитные. Характерно наличие в черешке и пластинке схизогенных каналов, содержащих масло. Цветки в кистях или метелках, заканчивающих цветонос, актиноморфные, обоеполые. Околоцветник из 6 синих лепестковидных сегментов в 2 неравных кругах; сегменты сросшиеся у основания в короткую трубку. Тычинок 6 в 2 кругах; нити короткие, у основания приросшие к сегментам околоцветника; пыльники прикреплены к нити основанием, раскрываются верхушечной порой или короткой щелью. Пыльцевые зерна однобороздные, иногда с трехлучевой бороздой. Гинецей из 3 плодолистиков, синкарпный; завязь полунижняя, 3-лопастная и 3-гнездная, с 2 анатропными семязачатками в каждом гнезде; столбик нитевидный, отходит из углубления между лопастями завязи и заканчивается коротким 3-лопастным рыльцем. Имеются септальные нектарники. Плод развивается только из одного плодолистика, обычно плоды односемянные, сухие. Семена без эндосперма, с относительно крупным зародышем и особым типом перисперма, развивающимся из тканей халазы (так называемый халазосперм) и содержащим крахмал и масло.
Хотя морфология цианаструма изучена относительно довольно хорошо, его биология, в том числе репродуктивная биология, остается фактически неизученной.
Семейство ирисовые, или касатиковые (Iridaceae) (Н. Н. Цвелев)
Наверное, всем известны такие декоративные растения, как касатик, или ирис (Iris, табл. 23, 24), шпажник, или гладиолус (Gladiolusг табл. 26, 3), и шафран, или крокус (Crocus, табл. 25). Все они принадлежат к семейству ирисовые, включающему еще целый ряд красивоцветущих растений. Ирисовые - довольно большое семейство, в которое входят около 1800 видов, принадлежащих к 75-80 родам. Ареал этого семейства очень велик: он охватывает почти всю сушу земного шара, исключая большую часть Арктики, крайний север таежной зоны Евразии, а также некоторые пустыни и участки равнинных тропиков с дождевыми лесами. Однако на протяжении этого обширного ареала ирисовые распределены очень неравномерно. В значительной по площади северной его части с умеренно теплым климатом представлены лишь немногие роды этого семейства, из которых дальше всего на север - до южных районов Арктики - проникают богатые видами роды ирис и сисиринхий, или голубоглазка (Sisyrinchium). Резко выделяются две области наибольшего разнообразия ирисовых: южноафриканская, где встречаются 45 родов и около 900 видов и центрально-южноамериканская с меньшим количеством родов и видов. Очень бедны представителями этого семейства Юго-Восточная и Южная Азия. В Австралии, Тасмании и Новой Зеландии ирисовые представлены 7 родами, из которых 3 эндемичных - изофизис (Isophisis), дипларрена (Diplarrhena) и патерсония (Patersonia) - очень оригинальны по морфологии. Особенно много эндемиков - 28 родов и около 450 видов - в Южной Африке (Капское флористическое царство), причем ареал многих из них (например, своеобразных полукустарников витсении - Witsenia и клаттии - Klattia) ограничен небольшим по площади участком в юго-западной части Капской земли. Преимущественное распространение ирисовых в южном полушарии позволяет связывать происхождение этого семейства с частями древней Гондваны, о чем свидетельствует, в частности, существование разрывов в ареалах некоторых родов (например, диэтеса - Dietes) между Южной Африкой, Австралией и Новой Зеландией. Американские роды значительно более обособлены от африканских и австралийских, однако один вид преимущественно американского рода сисиринхий - сисиринхий красивый (Sisyrinchium pulchellum) - распространен в Австралии, Новой Зеландии и Новой Гвинее.

Таблица 23. Ирисовые: 1 - ирис пестрый (Iris variegata); 2 - ирис водяной (I. pseudacorus); 3 - ирис Клятта (I. Klattii); 4 - ирис золотисторасписанный (I. chrysonrarhes)
Почти все ирисовые - многолетние травы, часто эфемероиды с мясистыми корневищами, клубнями и луковицами. Лишь представители 3 южноафриканских родов трибы нивениевых (Nivenieae): нивения (Nivenia), витсения и клаттия - являются своеобразными вечнозелеными полукустарниками с разветвленными одревесневающими стеблями высотой до 0,5 м, обладающими способностью к вторичному росту (рис. 98). Наибольшей высоты - до 1,8 м - достигают, однако, стебли некоторых травянистых растений из родов ирис и уотсония (Watsonia). Напротив, у видов крокуса, галаксии (Galaxia) и сирингодеи (Syringodea) стебель редуцирован до очень короткой стрелки, несущей цветок и расположенной во время цветения под поверхностью земли. Сосуды в стебле имеются только у сисиринхия, обычно же они находятся только в корнях, причем у видов триб аристеевых (Aristeeae) и нивениевых они имеют более примитивную, лестничную перфорацию.
Рис. 98. Ирисовые - полукустарники. Витсения маурa (Witsenia maura): 1 - цветущая ветвь; 2 - цветок. Клаттия разделенная (Klattia partita): 3 - цветущая ветвь; 4 - цветок
Листья ирисовых без черешков (рис. 99), расположены обычно двурядно и делятся на влагалищеобразное основание и пластинку с параллельным жилкованием. Наиболее распространенная форма листовых пластинок мечевидная, особенно характерная для корневищных родов, например ириса или сисиринхия. Такие пластинки имеют две боковые поверхности, которые фактически являются сторонами разросшейся нижней поверхности листа. Несколько реже встречаются листья с линейной или ланцетной пластинкой обычного для большинства однодольных строения: с развитыми верхней и нижней поверхностями (например, у видов очень близкого к ирису рода юнона - Juno, рис. 99, 2, табл. 26, 2). У крокусов такие листья очень узкие, с завернутыми вниз боковыми сторонами. Представители двух южноафриканских родов - тритониопсиса (Tritoniopsis) и анапалины (Anapalina) - имеют очень жесткие, кожистые листья с выступающими главными жилками. Пожалуй, наиболее специализированное строение листовых пластинок встречается у некоторых клубневых и луковичных эфемероидов. У них пластинки имеют цилиндрическую или четырехгранную форму (например, у восточносредиземноморского иридодиктиума сетчатого - Iridodictyum reticulatum, табл. 24, 4-5, известного также под названием ирис сетчатый - Iris reticulata), вероятно происходящую от мечевидной формы, свойственной более примитивным родам.
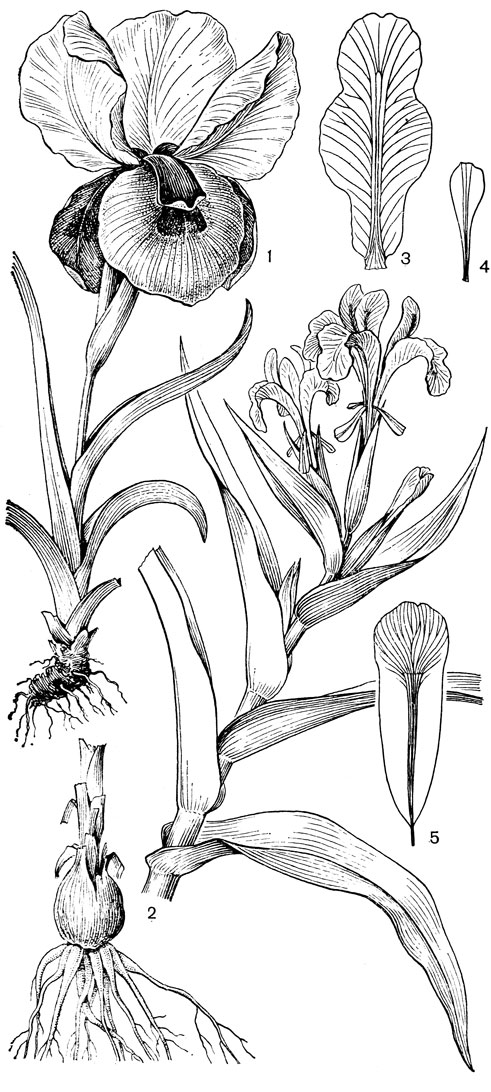
Рис. 99. Ирисовые. Ирис грузинский (Iris iberica): 1 - общий вид. Юнона великолепная (Juno magnifica): 2 - общий вид; 3 - наружный листочек околоцветника; 4 - внутренний листочек околоцветника; 5 - лопасть столбика

Таблица 24. Ирисовые: 1 - ирис японский (Iris japonica), оранжереи Ленинградского государственного университета; 2 - иридодиктиум Виноградова (I. winogradovii), парк Ботанического института АН СССР в Ленинграде; 3 - ирис сибирский (I. sibirica), там же; 4 - иридодиктиум сетчатый, сорт Гармония (I. reticulatum, cv. Harmony), Крым, Никитский ботанический сад; 5 - иридодиктиум сетчатый, сорт Кэнтеб (I. reticulatum, cv. Cantab), там же
Для большинства луковичных и клубневых ирисовых характерно присутствие влагалищеобразных низовых листьев, у некоторых видов шафрана достигающих значительной величины и окутывающих в виде чехла весь побег до его выхода на поверхность почвы. У всех ирисовых имеются также верховые, влагалищеобразные листья, прицветники и предлистья, образующие обертку или покрывало для одного или нескольких цветков. Они могут быть очень различными по величине, форме и консистенции у разных родов, а у капского полукустарника клаттии они ярко окрашены, выполняя функцию привлечения насекомых-опылителей.
Актиноморфные или зигоморфные и тогда двусторонне-симметричные цветки ирисовых обычно имеют относительно крупные размеры (исключая капский род микрантус - Micranthus) и собраны в верхоцветные соцветия, которые, однако, могут редуцироваться до одного цветка (у крокуса, некоторых видов ириса и в других родах). У шпажника и многих других родов трибы иксиевых (Ixieae) цветки собраны в односторонние колосовидные соцветия, причем каждый цветок окружен оберткой из прицветника и двукилевого предлиста, а у некоторых видов южноафриканского рода бобартии (Bobartia) цветки собраны довольно густыми головками.
Околоцветник цветка ирисовых венчиковидный, трехмерный и состоит из 2 кругов сегментов, которые могут бытр до основания свободными (например, у сисиринхия) или сросшимися в более или менее длинную трубку. Особенно большой длины - часто свыше 10 см - трубка околоцветника достигает у крокуса (рис. 100) и близкой к нему сирингодеи, где она выполняет функцию вынесения цветка над поверхностью почвы при сильно редуцированном стебле. У обоих этих родов завязь во время цветения находится довольно глубоко под поверхностью земли.
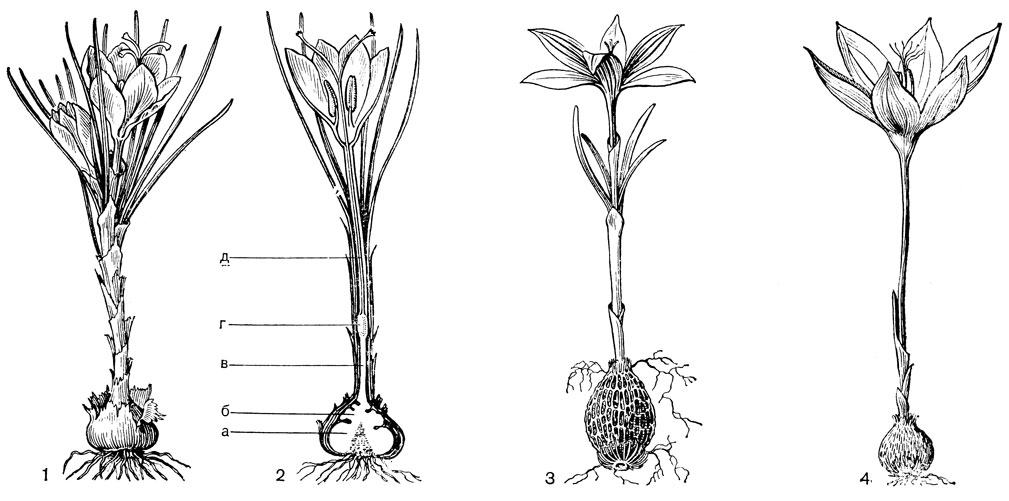
Рис. 100. Крокус, или шафран (Crocus): 1 - шафран посевной (С. sativus); 2 - он же на продольном разрезе через растение (а - старый клубень, б - молодой клубень, в - стрелка, г - завязь, д - трубка околоцветника); 3 - шафран сетчатый (С. reticulatus); 4 - шафран прекрасный (С. speciosus)
Сегменты обоих кругов околоцветника часто отличаются друг от друга по форме и величине, что можно видеть уже на примере хорошо всем известного ириса (рис. 99, 2), имеющего очень сложное строение в связи с высокой специализацией к энтомофилии. Все сегменты околоцветника у видов этого и родственных родов сужены у основания в ноготок. Доли внутреннего круга направлены вверх и играют роль привлечения насекомых-опылителей; доли наружного круга обычно более широкие, отогнуты в стороны и являются посадочной площадкой для них. Уже у некоторых видов ириса сегменты внутреннего круга становятся небольшими и очень узкими, а у американской тигридии (Tigridia, табл. 26,1) и австралийской патерсонии они вообще малозаметны. Гораздо реже - только у австралийско-южноамериканского рода либертии (Libertia) - имеется обратное соотношение: наружные сегменты околоцветника становятся очень мелкими, как бы образующими чашечку. Соцветия некоторых видов этого рода (рис. 101) внешне очень похожи на соцветия стрелолиста (Sagittaria). У наиболее примитивных родов семейства, например у сисиринхия, все 6 сегментов околоцветника вполне свободны и сходны друг с другом по форме и величине.
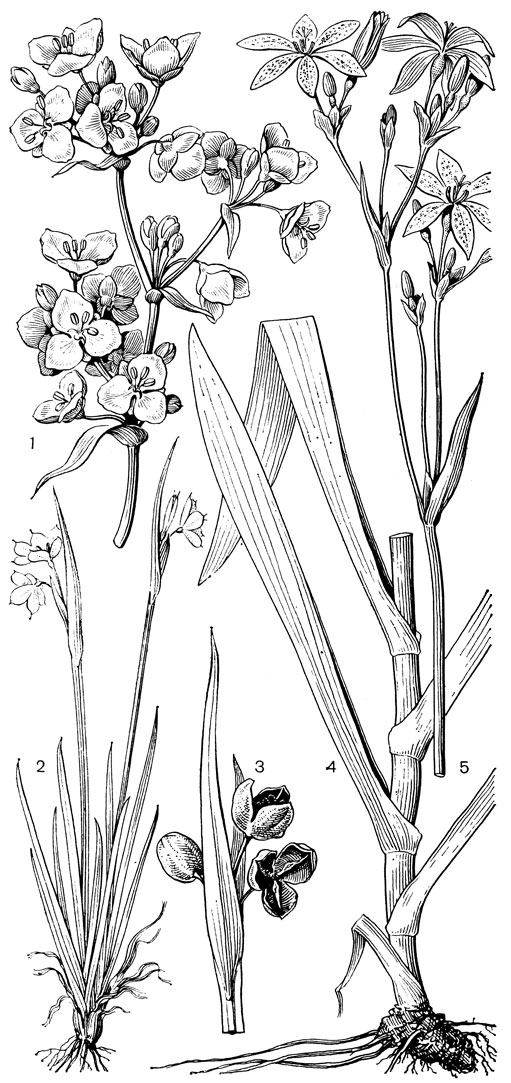
Рис. 101. Ирисовые. Либертия чилийская (Libertia chilensis): l - соцветие. Сисиринхий горный (Sisyrinchium montanum): 2 - общий вид; 3 - соцветие при плодах. Беламканда китайская (Belamcanda chinensis): 4 - нижняя часть растения; 5 - соцветие
В пределах трибы иксиевых (в принимаемом нами более широком ее объеме) есть роды с сильно зигоморфными цветками, по строению нередко напоминающими цветки норичниковых или губоцветных. Из видов флоры СССР двусторонне-симметричные цветки у шпажника (рис. 102, 5), еще более зигоморфные они у южноафриканских родов антолиза (Antholyza), аномалесия (Anomalesia), кентросифон (Kentrosiphori) и др. У кентросифона верхний сегмент околоцветника сильно увеличен, образуя верхнюю губу, а остальные, значительно более мелкие, образующие нижнюю губу, имеют у основания мешковидное вздутие (рис. 103, 6). Околоцветник антолизы имеет длинную узкую трубку и еще более похож по строению на венчик некоторых губоцветных.
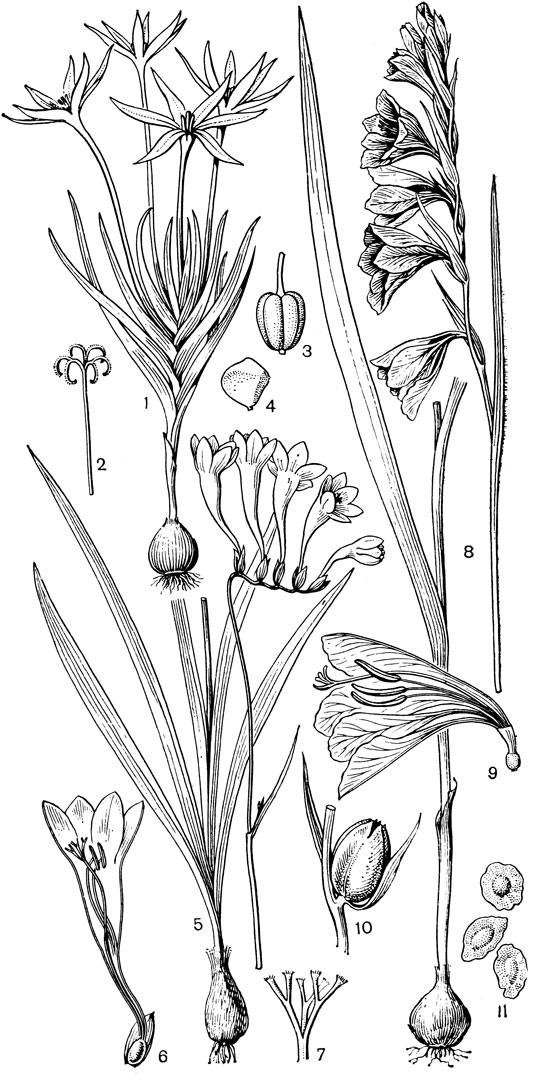
Рис. 102. Ирисовые. Лаперузия ароматнейшая (Lapeirousia odoratis- sima): 1 - общий вид; 2 - верхняя часть столбика; 3 - завязь; 4 - семя. Фреезия переломанная (Freesla retracta): 5 - общий вид; 6 - продольный разрез через цветок; 7 - ветви столбика. Шпажник черепитчатый (Gladiolus imbricatus): 8 - общий вид; 9 - цветок в продольном разрезе: 10 - плод; 11 - семена
В семействе ирисовых и даже в пределах одного только рода ирис можно встретить почти все возможные варианты окраски цветков. Нередко сегменты околоцветника очень пестро окрашены, например у некоторых тигридий или ирисов. У степного ириса карликового (Iris pumila) в одной популяции можно встретить цветки разнообразной окраски, от чисто-желтых до сине-фиолетовых. Существенно различаются роды ирисовых по времени цветения каждого цветка. Так, у родов из триб ирисовых (Irideae) и сисиринхиевых (Sisyrinchieae) цветение одного цветка продолжается менее суток, а у родов из триб иксиевых и шафрановых (Сrосеае) - по нескольку суток.
Лишь у рода кампинема (Campynema) с 2 видами в Тасмании и Новой Каледонии и монотипного новокаледонского рода кампинеманта (Campynemanthe), отнесенных А. Л. Тахтаджяном (1980) к семейству ирисовых, имеется 6 тычинок; у всех остальных ирисовых 3 тычинки. У многих родов с актиноморфными цветками, в том числе у сисиринхия, нити тычинок более или менее срастаются друг с другом в трубку, окружающую завязь. В зигоморфных цветках шпажника или антолизы тычинки обычно тесно сближены и располагаются под верхней долей околоцветника, играющей роль верхней губы. В качестве исключения цветки австралийской дипларрены имеют только 2 развитые тычинки (и 2 ветви столбика), а третья видоизменена в стаминодий. Как правило, тычиночные нити прикреплены к основанию сегментов околоцветника, а в цветках, околоцветник которых имеет более или менее длинную трубку,- у основания отгиба, где обычно находятся и вместилища нектара в виде небольших ямок. Пыльцевые зерна ирисовых очень разнообразны по строению, однако чаще всего имеют однобороздную оболочку.
Гинецей ирисовых синкарпный и образован 3 плодолистиками. Завязь почти всегда нижняя и 3-гнездная, с многочисленными (очень редко немногими) семязачатками. Единственными исключениями являются тасманский изофизис с верхней завязью и средиземноморский гермодактилус (Germodactylus) с одногнездной завязью, в отношении других деталей строения цветка близкий к ирису. Лишь у родов из триб аристеевых и нивениевых столбик заканчивается почти цельным рыльцем (рис. 104, 3). Обычно же он делится на 3 ветви, строение которых связано со способом опыления и имеет большое, систематическое значение. У ириса и представителей многих других родов трибы ирисовых ветви столбика лепестковидно расширены и делятся на 2 направленные вверх лопасти - "гребешки". Близ основания этих лопастей на нижней поверхности ветвей столбика располагаются клапановидные рыльца, воспринимающая пыльцу поверхность которых находится на их обращенной наружу стороне. Каждая ветвь столбика образует как бы свод над верхней поверхностью наружных сегментов околоцветника. У африканского рода феррария (Ferraria) ветви столбика над клапановидными рыльцами перисто рассечены (рис. 103, 8). Крупные, бахромчатые по краю ветви столбика у патерсонии как бы замещают здесь редуцированный внутренний круг околоцветника (рис. 104, 7). У видов шафрана 3 ветви столбика могут, в свою очередь, многократно ветвиться, причем рыльца располагаются по краю особых впадин на верхушках конечных ветвей.
По строению плодов ирисовые довольно однообразны: все они имеют плод - многосемянную синкарпную коробочку, вскрывающуюся локулицидно - по средней жилке каждого из трех плодолистиков. Нередко при вскрывании коробочки ее створки расходятся лишь в верхней части, в других случаях (например, у некоторых ирисов), напротив, створки остаются спаянными у верхушки коробочки. Семена могут быть самой различной формы, определяющейся их расположением в коробочке и способом распространения. В частности, они могут иметь мясистый ариллусоподобный придаток или крыловидную кайму. Как и у многих других семейств однодольных, в семенах ирисовых обильный эндосперм и маленький зародыш.
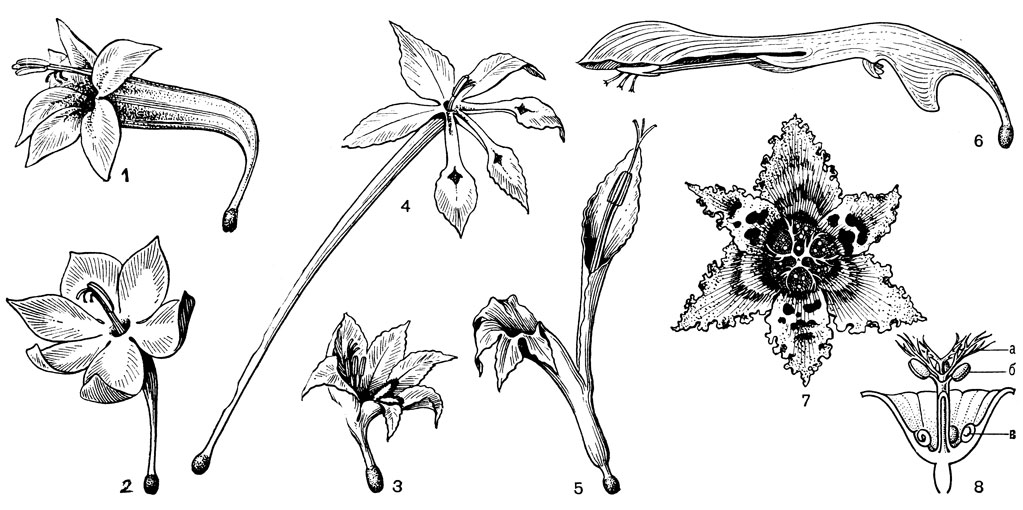
Рис. 103. Цветки ирисовых: 1 - уотсония Шлехтера (Watsonia schlechteri); 2 - уотсония окаймленная (W. marginata); 3 - бабиана складчатая (Babiana plicata); 4 - бабиана трубкоцветковая (В. tubiflora); 5 - бабиана раскрытая (В. ringens); 6 - кентросифон мешковидный (Kentrosiphon saccatus); 7 - феррария волнистая (Ferraria undulata); 8 - цветок феррарии волнистой в продольном разрезе (а - ветви столбика, б - пыльники, в - вместилища нектара)
В экологическом отношении ирисовые являются преимущественно растениями открытых пространств, от болот и берегов водоемов до степей и пустынь. Среди них нет настоящих лесных видов, хотя некоторые, обычно обитающие на лесных полянах и опушках, могут заходить и в разреженные участки леса (например, ирис безлистный - Iris aphylla - в дубравах юга европейской части СССР). На болотах и по берегам водоемов растут некоторые виды ириса (например, широко распространенный в Евразии ирис болотный - I. pseudacorus, табл. 23, 2), африканской уотсонии и сисиринхия; на болотистых местах растет и своеобразный капский полукустарник витсения маура (Witsenia maura, рис. 98, 1) и многие виды других родов. Целый ряд ирисовых приурочен к лугам различных типов, как равнинным, так и высокогорным, хотя доминантами или эдификаторами они здесь обычно не являются. Это преимущественно корневищные виды из родов ирис, сисиринхий и других, хотя среди луговых ирисовых встречаются и клубневые (например, многие виды шпажника). Еще более многочисленны ирисовые в травянистых и кустарниковых группировках ксерофитной растительности низкогорий, где особенно много видов эфемероидов с клубнями и луковицами, предки которых, по-видимому, имели корневища.
В частности, очень заметную роль играют многие ирисовые в районах Южной Африки с засушливым летним периодом и зимними осадками. Из ирисовых флоры СССР, обитающих в песчаных степях Казахстана и крайнего юга Сибири, а за пределами СССР - в Центральной Азии, следует особенно отметить ирис тонколистный (I. tenuifolia), нередко являющийся эдификатором растительных группировок. Это растение с очень узкими (шириной 1-2,5 мм) и жесткими листьями образует крупные и плотные дерновины.
Из представленных в семействе жизненных форм наименее специализированной можно считать жизненную форму примитивного и во многих других отношениях (лестничная перфорация сосудов, свободные сегменты околоцветника одинакового строения, почти цельное рыльце и т. п.) южноафриканского рода аристея (Aristea): прямостоячее полурозеточное растение со слабо облиственным, лишь в соцветии разветвленным стеблем и мочковатой корневой системой без каких-либо запасающих подземных органов стеблевого или листового происхождения (рис. 104, 1). Мочковатая корневая система и у южноафриканских полукустарников из трибы нивениевых (рис. 98).

Рис. 104. Ирисовые. Аристея крылатая (Aristea alata): 1 - общий вид; 2 - цветок; 3 - столбик с рыльцем и тычинки; 4 - раскрывшаяся коробочка; 5 - семя. Патерсония сизая (Раtersonia glauca): 6 - верхушка стебля с оберткой и цветком; 7 - столбик с рыльцем и тычинки; 8 - плод
Среди довольно многочисленных корневищных ирисовых виды сисиринхия (рис. 101, 2) имеют тонкие и обычно короткие корневища с относительно небольшими запасами питательных веществ и приурочены к более или менее влажным местообитаниям-лугам и болотам. Сложную систему толстых симподиальных корневищ образуют большинство видов ириса, многие из которых встречаются в областях с засушливым климатом. Такие корневища состоят из укороченных годичных побегов, покрытых рубцами от отмерших листьев. Первоначально они часто развиваются у поверхности земли, но затем втягиваются в землю с помощью контрактильных (сокращающихся) корней. По величине и форме годичных звеньев корневищ, как по годичным кольцам деревьев, можно не только определить возраст растения, но и выяснить, насколько благоприятными для него были отдельные годы. Обычно корневища ирисов живут до 8-13 лет.
По особенностям побегообразования виды ирисов делятся на плотнокустовые, рыхлокустовые и длиннокорневищные. Примером плотнокустового вида может служить упомянутый выше ирис тонколистный, а длиннокорневищного - обитающий на песках приречных террас Юго-Восточной Европы ирис песчаный (Iris arenaria) с обычно. удаленными друг от друга надземными побегами.
У ирисовых - клубневых и клубнелуковичных геофитов - клубни, как правило, образуются из нижних междоузлий стебля. Только у некоторых видов тигридии отмечается образование клубневидно утолщенных корней. Клубни стеблевого происхождения обычно закладываются уже на стадии проростка. Так, у проростка шпажника утолщается второе междоузлие над семядолей. Имеющиеся у цветущего растения шпажника или шафрана клубень и клубнелуковица являются результатом роста в предшествующий год. В то время как эти органы постепенно уменьшаются за счет расходования содержащихся в них питательных веществ, утолщается самое нижнее междоузлие цветущего побега, давая новый клубень, накапливающий питательные вещества для следующего года. У основания нового клубня образуются контрактильные корни, постепенно втягивающие его в землю. Клубни и клубнелуковицы обычно бывают одеты отмершими низовыми листьями, которые позднее расщепляются на волокна, образуя параллельно- или сетчато-волокнистый чехол вокруг клубня.
Луковицы, состоящие из более или менее утолщенного основания стебля - донца и мясистых чешуй листового происхождения, имеются, например, у близкородственных роду ирис и часто объединяемых с ним эфемероидных родов ксифиум (Xiphium), юнона и гинандририс (Ginandriris). У представителей последнего клубневидное донце луковицы одето чешуевидными с мясистым основанием листьями, которые постепенно становятся сухими и сетчато-волокнистыми. У распространенных в Средней и Юго-Западной Азии родов юнона и ксифиум стеблевая часть луковицы может быть слабо развита и основную ее часть составляют 3-5 (редко больше) мясистых чешуевидных листа. У также близкого к ирису и распространенного в Юго-Западной Азии рода иридодиктиум (Iridodictyum) основу небольшой луковицы составляет всего одна мясистая чешуя, сросшаяся своими боковыми сторонами. В ходе цветения и плодоношения эта чешуя становится сухой и сетчато-волокнистой, замещаясь вновь развившейся молодой чешуей.
Многие ирисовые, как корневищные, так и клубневые и луковичные, могут образовывать горизонтальные ползучие подземные побеги - столоны, дающие начало новым особям. Довольно широко распространен (например, у многих видов шпажника) и другой способ вегетативного размножения - образование придаточных клубеньков и луковичек у основания клубня или луковицы под их наружными оболочками. В садоводческой практике культивируемые ирисы легко размножают с помощью небольших отрезков их толстых корневищ.
Очень интересная жизненная форма геофита- эфемероида с редуцированным стеблем представлена среди ирисовых у видов древнесредиземноморского рода шафран и у южноафриканских родов сирингодея и галаксия. У них очень короткая и несущая всего один цветок стрелка во время цветения находится глубоко в земле вместе с завязью цветка, который выносится на поверхность земли очень длинной и узкой трубкой околоцветника (рис. 100). Удлинение стрелки, главным образом за счет роста ее клеток, происходит только при созревании плода, который раскрывается уже на поверхности земли. Среди многочисленных видов шафрана имеются как ранневесенние виды с листьями, появляющимися к концу цветения, так и виды, цветущие осенью, причем листья у них могут появляться одновременно с цветком (как у культивируемого шафрана посевного - Crocus sativus) или на следующий год вместе с плодом (например, у крымско-кавказского шафрана прекрасного - С. speciosus, табл. 25, 3). Шафран и галаксия очень сходны по облику, и прежде их относили к одной и той же трибе (или подсемейству) шафрановых. Однако это сходство оказалось результатом конвергенции. В настоящее время доказано, что галаксия очень тесно связана с родом морея (Моrаеа) из трибы ирисовых, замещающим близкий род ирис в тропической и Южной Африке, а шафран и сирингодея значительно ближе к родам трибы иксиевых. Средиземноморско-африканский род ромулея (Romulea) может быть принят за переходный от иксиевых, имеющих развитые надземные стебли, к бесстебельным родам, подобным шафрану. Цветочные стрелки у видов этого рода нередко ветвятся и уже во время цветения растения достигают поверхности почвы, а цветки имеют короткую трубку околоцветника.

Таблица 25. Ирисовые: 1 - шафран крымский (Crocus tauricus), Южный Крым, Ангарский перевал; 2 - шафран Королькова (S. korolkowii), парк Ботанического института АН СССР в Ленинграде; 3 - шафран прекрасный (С. speciosus), Украина; 4 - шафран сузианский (С. susianus), Крым, Никитский ботанический сад
Цветки значительного большинства ирисовых опыляются с помощью насекомых. Однако некоторые африканские виды из родов уотсония (рис. 103), бабиана (Babiana, рис. 103, 105), кентросифон, аномалесия, хазманта (Chasmanthe) и анапалина имеют орнитофильные цветки: довольно крупные и обычно сильно зигоморфные, без запаха, но обильно выделяющие нектар. Их опылители - птицы из семейства нектарниц - просовывают головку внутрь цветка, касаясь ею сначала рылец, а затем глубже расположенных тычинок. Особенно специализированы в направлении орнитофилии цветки кентросифона мешковидного (Kentrosiphon saccatus), у которого нижняя губа околоцветника сильно укорочена и имеет мешковидный выступ, заполненный легко доступным для птиц нектаром (рис. 103, 6), длинная верхняя губа прикрывает в виде крыши тычинки и столбик с рыльцами.

Рис. 105. Бабиана раскрытая (Babiana ringens): верхняя часть растения с опылителем - птицей из семейства нектарниц
Энтомофильные цветки имеют очень различное строение в зависимости от особенностей насекомых-опылителей. Последних, как и опылителей-птиц, привлекает главным образом нектар, который вырабатывается специальными нектарниками, расположенными на завязи в местах срастания краев плодолистиков. Образующийся нектар может заполнять нижнюю часть трубки околоцветника, если последняя имеется, но чаще всего он находится в специальных вместилищах на внутренней стороне листочков или лопастей околоцветника близ места прикрепления к ним тычинок. У шафрана и других родов с узкой и длинной трубкой околоцветника нектар просачивается по трубке до расположенных у основания расширенной части околоцветника вместилищ.
Дополнительными средствами привлечения опылителей служит яркая окраска цветков, а иногда и листочков обертки, а также запах, свойственный многим ирисовым. Довольно сильным и приятным запахом обладают некоторые виды, опыляемые длиннохоботковыми ночными бабочками, особенно бражниками, например лаперузия ароматнейшая (Lapeirousia odoratissima), цветки которой имеют очень длинную трубку околоцветника (рис. 102, 1). Напротив, неприятным запахом падали при очень короткой трубке околоцветника обладают цветки опыляемой короткохоботковыми мухами феррарии волнистой (Ferraria undulata) из Южной Африки (рис. 103, 8). Интересно, что они и внешне напоминают падаль: доли околоцветника у них по краю волнистые и зеленоватые, а в остальной части пестрые темно-коричневые и беловатые. Цветки опыляемых ночными бабочками видов гесперанты (Hesperantha) открываются только вечером и закрываются на день. Трубку околоцветника средней величины обычно имеют виды, опыляемые крупными перепончатокрылыми и длиннохоботковыми мухами. Нередко виды одного и того же рода приспособлены к разным опылителям. Например, в южноафриканском роде бабиана бабиана складчатая (Babiana plicata) с короткой трубкой околоцветника опыляется перепончатокрылыми и мухами, бабиана трубкоцветковая (В. tubiflora) с очень длинной трубкой - длиннохоботковыми бабочками, бабиана раскрытая (В. ringens) с двугубым околоцветником оригинального строения - птицами (рис. 103, 105).
Цветки с наиболее просто построенным околоцветником из свободных сегментов, например у сисиринхия или аристеи, опыляются различными короткохоботковыми насекомыми. У этих родов, как и у большинства других родов ирисовых, самоопылению препятствует протандрия: пыльники вскрываются одновременно с раскрыванием околоцветника, а ветви рыльца расходятся в стороны, открывая воспринимающую пыльцу поверхность, обычно при уже пустых пыльниках. У шпажника (рис. 102, 9) и многих других ирисовых с зигоморфными цветками все 3 тычинки располагаются в верхней части более или менее двугубого околоцветника, и насекомые (обычно крупные перепончатокрылые), вползая в цветок, касаются их спинкой. Лопасти рыльца в начале цветения не только сомкнуты друг с другом, но и отклонены кверху, так что посещающие цветок насекомые их не задевают. При уже пустых пыльниках лопасти рыльца раскрываются и наклоняются книзу, так что вползающее в цветок насекомое касается спинкой прежде всего их, а затем уже пыльников. Однако в конце цветения, если перекрестное опыление не состоялось, у шпажника становится возможным самоопыление: рыльце поникающего цветка, отгибаясь книзу, касается пыльцы, выпавшей из пыльников на нижние сегменты околоцветника. У некоторых видов шпажника отмечена и гетеростилия, при которой рыльца у короткостолбиковых особей не возвышаются над верхушками тычинок и не могут быть опылены насекомыми.
У видов шафрана (рис. 100) 3 основные ветви столбика, в свою очередь, ветвятся, а рыльца находятся на более или менее вогнутых верхушках их конечных разветвлений. Насекомые (обычно шмели и другие крупные перепончатокрылые), вползая в цветок, прежде всего касаются спинкой выступающих над пыльниками ветвей столбика, и пыльца со спинки как бы счищается краями воронкообразных рылец. Затем насекомое соприкасается спинкой с пыльником, щель которого обращена наружу, и получают часть пыльцы из него. Выползая из цветка, насекомое вновь касается спинкой ветвей столбика, но пыльца со своего цветка на рыльца не попадает, так как последние обращены своей вогнутой поверхностью кверху. Цветки шафрана факультативно протандричны, возможно, в зависимости от погодных условий. Кроме того, они легко самоопыляются, что гарантирует образование плодов в случае неблагоприятных погодных условий, воздействию которых так часто подвергаются весеннецветущие и осеннецветущие виды. У многих видов имеется даже специальный механизм, обеспечивающий самоопыление в конце цветения. Трубка околоцветника удлиняется, прикрепленные в его расширенной части тычинки продвигаются вверх, и ветви не изменяющего свою длину столбика оказываются расположенными между пыльниками. Пыльники к этому времени тоже изменяют свое положение, поворачиваясь щелью внутрь цветка. Таким образом, пыльца из пыльников может попасть на рыльца при раскачивании цветка ветром; кроме того, самоопылению в этом случае способствуют посещающие цветок насекомые, которые, прикасаясь спинкой к пыльникам, вытряхивают пыльцу на рыльца. У культивируемого шафрана посевного рыльца всегда находятся значительно выше пыльников, однако этот вид всегда стерилен и, вероятно, возник в культуре в результате гибридизации нескольких видов.
У некоторых ирисовых в ходе сопряженной эволюции цветков и опыляющих их насекомых выработались сложные приспособления для более совершенного перекрестного опыления. Особенно интересен способ обеспечения перекрестного опыления у видов ириса и близких к нему родов, имеющих сходное строение цветка (рис. 106). Верхняя поверхность отклоненных в сторону и вниз наружных сегментов околоцветника выполняет здесь функцию посадочной площадки для насекомых-опылителей, причем у многих более специализированных видов на ней имеется бородка волосков, которая направляет движение насекомых внутрь цветка. Над каждым наружным сегментом околоцветника в виде низкого свода располагается лепестковидно расширенная двулопастная ветвь столбика с направленным вниз клапановидным рыльцем. Позади рыльца к нижней поверхности этой ветви прилегает пыльник одной из 3 тычинок. Таким образом, цветок ириса как бы делится на три самостоятельных отделения, каждое из которых очень сходно с двугубым цветком многих других растений. В начале цветения вследствие более или менее выраженной протандрии клапановидные рыльца прилегают своей передней, воспринимающей пыльцу поверхностью к нижней поверхности ветви столбика. Вползающее в это время в щель между ветвью столбика и наружным сегментом околоцветника насекомое касается лишь нижней поверхности уже вскрывшегося продольной щелью пыльника, откуда на его спинку попадает часть пыльцы. Когда клапановидное рыльце созревает, оно свешивается вниз воспринимающей пыльцу поверхностью навстречу насекомому, которое, прежде всего касается спинкой его, а затем уже расположенного глубже пыльника. Выползая из цветка, насекомое еще раз касается спинкой пыльника, получая еще одну порцию пыльцы, а затем внутренней поверхности рыльца, которое под его давлением отгибается наружу и вверх. Поскольку внутренняя поверхность клапановидного рыльца не может воспринимать пыльцу, опыление пыльцой того же цветка в этом случае исключается и вообще может иметь место лишь в том случае, если насекомое последовательно посетит два или все три отделения одного и того же цветка, что бывает очень редко. У некоторых видов ириса направляющие движение насекомого расширенная ветвь столбика и наружный сегмент околоцветника настолько плотно пригнаны друг к другу, что приоткрыть их может только очень сильное насекомое, подобное шмелям, которые являются наиболее обычными опылителями цветков ириса. Протандрия у ирисов нередко определяется только морфологическим строением: рыльца созревают одновременно с вскрыванием пыльников, но некоторое время остаются прижатыми воспринимающей поверхностью к ветви столбика, что исключает возможность их опыления.
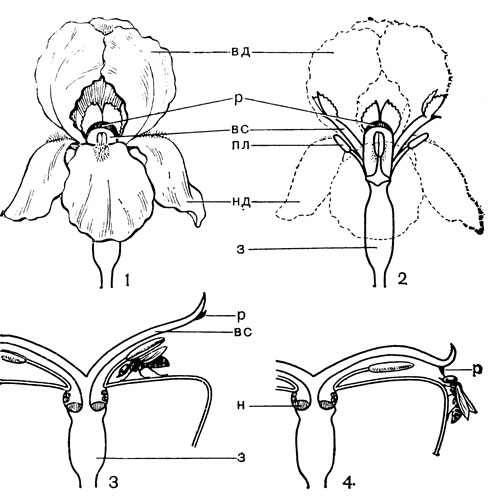
Рис. 106. Схема строения цветка ириса (1 и 2) и способ его опыления (3 - в начале цветения, 4 - в конце цветения): нд - наружные доли околоцветника, вд - внутренние доли околоцветника, вс - двулопастные ветви столбика, р - рыльца, пл - пыльники, н - нектар, з - завязь (по Г. И. Родионенко)
Многие ирисовые не имеют специальных приспособлений для распространения семян. Коробочки растрескиваются, створки их расходятся в стороны, и семена постепенно выпадают (барохория). При наличии относительно высоких стеблей и ветра семена и таким способом могут распостраняться довольно далеко; кроме того, уже выпавшие семена могут переноситься водными потоками или вместе с почвой на ногах животных и человека. Так;, семена широко распространившегося в Европе, в том числе и в СССР, но происходящего из Северной Америки сисиринхия горного (Sisyrinchium montanum, рис. 101, 2) часто разносятся на ногах человека, вследствие чего этот вид быстро расселяется вдоль троп. Выпавшие из коробочек семена болотных и прибрежных ирисов (например, обычного в СССР ириса водяного), имеющие несмачивающийся эпидермис и пористую оболочку обычно распространяются водой, особенно во время весеннего половодья (гидрохория). Очень толстый слой пористой ткани, напоминающей пробку, имеется у семян ириса виннокрасного (Iris vinicolor) из бассейна Миссисипи. Семена другого прибрежного вида - ириса щетинистого (I. setosa) могут плавать по воде, сохраняя всхожесть до 200 и более суток. Многим представителям семейства свойственна анемохория - распространение семян с помощью ветра. В разной степени крылатые семена отмечаются у некоторых ирисов, у видов шпажника и уотсонии, а также у представителей некоторых других родов. У ирисовых отмечается также мирмекохория - распространение семян с помощью муравьев. В частности, у семян ряда видов ириса и средиземноморского гермодактилуса имеются ариллусоподобные придатки, поедаемые муравьями.
Семейство ирисовых достаточно четко делится на 2 основных подсемейства: собственно ирисовых (Iridoideae) и иксиевых (Ixioideae), различающихся по продолжительности цветения отдельных цветков, строению соцветия и ряду других признаков. Первое из этих подсемейств представлено трибами аристеевых (Aristeae), нивениевых (Nivenieae), сисиринхиевых (Sisyrinchieae), тигридиевых (Tigridieae) и ирисовых (Irideae), а второе - трибами иксиевых (Ixieae) и шафрановых (Сrосеае). Кроме того, в особые монотипные подсемейства изофизисовых (Isophysidoideae) и геосирисовых (Geosiridoideae) могут быть выделены тасманский род изофизис и мадагаскарский род геосирис (Geosiris), систематическое положение которых все еще остается не вполне определенным из-за их недостаточной изученности. Изофизис, впервые отнесенный к ирисовым Дж. Хатчинсоном, обладает таким примитивным признаком, как верхняя завязь, и, вероятно, является остатком предковой для всех ирисовых группы, еще сохранившей много общего с другими семействами порядка лилейных. Геосирис, напоминающий по облику представителей семейства бурманниевых, был включен в семейство ирисовых еще Б. Байоном (1890), а в последнее время вновь присоединен к ним Р. Торном (1976) и А. Л. Тахтаджяном (1980). Это небольшое сапрофитное растение с корневищем и стеблем, покрытым чешуевидными листьями и заканчивающимися одним или несколькими зигоморфными цветками. Кроме того, А. Л. Тахтаджян (1980) перевел из семейства гипоксисовых (Ilypoxidaceae) в ирисовые роды кампинему и кампинеманту, выделяя их в подсемейство кампинемантовые (Campynemanthoideae). К сожалению, оба эти рода все еще очень мало изучены.
Из подсемейства собственно ирисовых наиболее примитивной может быть признана триба аристеевых, из 5 родов которой аристея встречается в тропической и Южной Африке, патерсония - в Австралии, Тасмании и Новой Зеландии и 3 рода в Андах Южной Америки. Все роды трибы - травянистые растения, для которых, как и для родов следующей трибы, характерно присутствие наиболее примитивных в семействе сосудов с лестничной перфорацией. Виды аристеи имеют мочковатую корневую систему, свободные листочки околоцветника одинакового строения, свободные тычинки и почти цельные рыльца (рис. 104, 2-3). Остальные роды - корневищные травы с более или менее срастающимися в трубку листочками околоцветника. Интересная особенность патерсонии - значительная редукция внутренних листочков околоцветника (рис, 104, 6).
Триба нивениевых близка к предыдущей трибе и нередко с ней объединяется, но принадлежащие сюда 3 южноафриканских рода: нивения, витсения и клаттия являются полукустарниками, обладающими способностью к вторичному росту (рис. 98). Сегменты околоцветника у основания сросшиеся, рыльце едва 3-лопастное, корневая система мочковатая. Как и аристеевые, прежде присоединялась к трибе сисиринхиевых, но отличается от нее по строению рылец и пыльцевых зерен, а также по жизненной форме.
Из 6 родов трибы сисиринхиевых бобартия встречается в Южной Африке, дипларрена - в Австралии, тапейния (Tapeinia) - на юге Чили, либертия и ортозантус (Orthosanthus) - в Австралии и Америке. Наиболее известный род - сисиринхий, включающий около 100 видов, распространен в Северной и Южной Америке, за исключением сисиринхия красивого, обитающего в Австралии, Новой Гвинее и Новой Зеландии. Некоторые виды культивируются в качестве декоративных растений и дичают, а один из них - сисиринхий горный (рис. 101, 2) с небольшими голубовато-фиолетовыми цветками и коротким тонким корневищем полностью натурализовался во многих странах Европы, в том числе и на территории европейской части СССР (Прибалтика, Ленинградская и Московская области, запад Украины). В Андах некоторые виды сисиринхия поднимаются до 5000 м, а сисиринхий горный заходит в американскую Арктику, включая Гренландию. Для трибы сисиринхиевых характерны актиноморфные, быстро отцветающие цветки со свободными или почти свободными сегментами околоцветника, неразветвленные и нелепестковидные ветви столбика, тычинки, чередующиеся с ветвями столбика, и довольно тонкие корневища.
К трибе тигридиевых, включающей и ципуровые Хатчинсона, принадлежат 12 родов, распространенных главным образом в Центральной и Южной Америке, а также на юге США. Это луковичные и клубнелуковичные, редко корневищные растения с актиноморфными цветками, имеющими свободные или лишь немного сросшиеся у основания сегменты околоцветника. В отличие от предыдущей трибы, тычинки у них супротивны часто разветвленным ветвям столбика, и поэтому они более тесно примыкают к трибе ирисовых. Однако у тигридиевых нектароносные вместилища находятся на внутренних сегментах околоцветника, а у ирисовых - на наружных.
Наиболее известный род трибы - тигридия с 15 видами, большая часть которых является мексиканскими эндемиками. У видов этого рода крупные и красивые цветки с чашеобразным или колокольчатым околоцветником и сросшимися в трубку тычиночными нитями. Многие из них очень декоративны, в особенности культивируемая уже с начала XVI в. тигридия павония (Tigridia pavonia, табл. 26, 2), называемая "тигровым цветком" из-за пестроокрашенного околоцветника, достигающего иногда в ширину 10-15 см. Сегменты околоцветника у нее снаружи обычно фиолетовые с желтыми полосками и пурпурными пятнами, внутри более крупные наружные сегменты светло-красные, а внутренние желтые с красными полосками и пятнами. Имеются садовые разновидности и с другой окраской цветков.
Около 15 родов трибы ирисовых распространены главным образом в Африке и внетропических областях Евразии. Лишь род ирис заходит в умеренно теплые и субтропические области Северной Америки. Во многом, исключая расположение нектароносных вместилищ, эта триба сходна с тигридиевыми, но представители входящих в нее родов, как правило, имеют брлее высокоспециализированные цветки часто с лепестковидно расширенными ветвями столбика. У представителей многих родов цветки сходны по строению и способу опыления с описанным выше цветком ириса.
Самый крупный по количеству видов род трибы и всего семейства - ирис, в который входит более 250 видов. Его ареал охватывает почти всю сушу между Северным полярным кругом и Северным тропиком. Наибольшее число видов приходится, однако, на страны Средиземноморья, Юго-Западную и Среднюю Азию, где встречаются также близкородственные луковичные и клубнелуковичные роды юнона, кси фиум, иридодиктион, гинандририс и др. Все виды ириса в узком понимании этого рода - корневищные растения, очень различные по высоте стеблей и плотности дерновин. Цветки могут быть всегда одиночными на очень коротком (высотой до 5 см) стебле, например у ириса карликового (Iris pumila), или многочисленными на стеблях высотой до 150 см. Трубка околоцветника также варьирует от почти отсутствующей (у ириса щетинистого) до очень длинной (у ириса карликового).
Из большого числа культивируемых видов ириса, многие из которых, возможно, имеют гибридное происхождение, наиболее известны ирисы германский (I. germanica), пестрый (I. variegata, табл. 23, 1), бледный (I. pallida), флорентийский (I. florentina), бузинный (I. sambucina), тусклоцветковый (I. sqnalens) и др. Существуют общества любителей ирисов и специальные журналы, посвященные этим растениям. Число садовых разновидностей ирисов необыкновенно велико и продолжает расти. По данным Г. И. Родионенко (1981), в настоящее время зарегистрировано более 30000 сортов ирисов.
Из дикорастущих ирисов флоры СССР наиболее известны желтоцветковый ирис болотный (I. pseudacorus, табл. 23, 2), ирис сибирский (I. sibirica, табл. 24, 3) с синими цветками и довольно узкими листьями, лесостепной ирис безлистный (I. aphylla), степной ирис карликовый, очень узколистный и густодерновинный ирис тонколистный (I. teniiifolia), ирис вильчатый (I. dichotoma) с вильчато разветвленным соцветием и др. Много оригинальнейших видов ириса в низкогорьях Закавказья, Турции и Ирана, например ирис грузинский (I. iberica) с очень крупным одиночным цветком (рис. 99, 1).
Стоит упомянуть еще о том, что именно цветок ириса был в дореволюционной Франции эмблемой французского королевского двора, его "геральдической лилией". Эти эмблемы наносились на королевские одежды, доспехи и другие предметы, принадлежавшие королям. Во время сражений французские солдаты несли впереди голубой флаг короля с тремя золотыми цветками ириса, символизирующими святую троицу.
Встречаются в культуре и другие роды трибы ирисовых. Из наиболее близких к ирису и часто объединяемых с ним родов часто культивируется в садах ранневесенний эфемероид - иридодиктиум сетчатый. Изредка культивируют виды рода морея (Моrаеа), замещающего близкий род ирис в Африке к югу от Сахары. Представители небольшого южноафриканского рода галаксия внешне очень сходны с шафраном, они также имеют стрелкообразный подземный стебель с одним цветком, однако этот род значительно ближе к таким родам трибы ирисовых, как гомерия (Homeria) и морея. Довольно обособленное положение в трибе занимает восточноазиатский род беламканда (Belamcanda), один вид которого - беламканда китайская (В. chinensis) заходит на крайний юг советского Дальнего Востока, где растет на песчаных местах близ морского побережья (рис. 101, 4, 5). По строению цветков это растение напоминает американские тигридии и также имеет пестрый околоцветник: красно-бурый или желтоватый с темно-пурпурными пятнами. Подобно многим другим ирисовым, беламканду китайскую культивируют в качестве декоративного растения.
В принимаемом нами более широком объеме триба иксиевых включает 30-32 рода, распространенных преимущественно в Южной Африке. Только род шпажник выходит за пределы Африканского континента, встречаясь в значи-тельной части Европы, а также в Средней и Юго-Западной Азии. В отличие от представителей всех предыдущих триб иксиевые имеют длительно цветущие цветки и обычно колосообразные соцветия. Околоцветник у них всегда более или менее сростнолистный, ветви столбика простые или двураздельные (у фреезии - Freesia, уотсонии и некоторых других родов), но не лепестковидно расширенные, как у многих родов трибы ирисовых. Более типичные роды иксиевых (например, иксия - Ixia) имеют актиноморфные или едва зигоморфные цветки. Роды с сильно зигоморфными цветками более или менее двугубого строения, с тычинками, смещенными под верхнюю губу околоцветника, иногда выделяют в особые трибы шпажниковых (Glaclioleae) и антолизовых (Antholyzeае). Все иксиевые - клубневые или клубнелуковичные растения, исключая небольшой африканский род схизостилис (Schizostylis) с короткими и толстыми корневищами.
Среди иксиевых много красивоцветущих растений, имеющих большое декоративное значение. Из них на первом месте стоит шпажник, или гладиолус, около 250 видов которого распространены главным образом в Африке к югу от Сахары. Немногие евразиатские виды с розовыми или розовато-фиолетовыми цветками, например шпажник обыкновенный (Gladiolus communis) и шпажник черепитчатый (G. imbricatus, табл. 26, 3), довольно красивы (рис. 102, 8, 9) и широко культивировались в качестве декоративных растений в садах Европы. Однако с начала прошлого века их стали вытеснять еще более красивоцветущие африканские виды (шпажник пурпурный - G. cardinalis, шпажник печальный - G. tristis и др.), а также гибриды между ними и европейскими видами. В настоящее время в культуре наиболее распространен гибридогенный шпажник гибридный (G. hybridus), имеющий цветки самой разнообразной окраски и представленный множеством садовых разновидностей. В Западной Европе и США работают общества любителей гладиолусов, регистрирующие все новые и новые сорта этого замечательного по своей декоративности растения.

Таблица 26. Ирисовые: 1 - тигридия павония (Tigridia pavonia), парк Ботанического института АН СССР в Ленинграде; 2 - юнона бухарская (Juno bucharica), Средняя Азия; 3 - гладиолус, или шпажник черепитчатый (Gladiolus imbricatus), Карпаты
Все более широко входит в культуру и другой род иксиевых - фреезия с 20 преимущественно южноафриканскими видами. Наибольшей популярностью пользуется фреезия переломанная (Freesia refracta), названная так за круто изогнутый под соцветием стебель (рис. 102, 5-7). Подобно шпажнику, это клубневое растение с простым, реже ветвистым стеблем. Околоцветник из узкотрубчатого основания довольно резко расширяется в колокольчатую верхнюю часть и варьирует по окраске от белого до фиолетового. Цветки имеют сильный приятный запах. Фреезия введена в культуру с конца прошлого века, но уже имеются многочисленные садовые разновидности, из которых особенно выделяются так называемые суперфреезии со стрелками высотой до 1 м и цветками диаметром свыше 5 см.
Менее известны в СССР, но довольно широко культивируются за его пределами представители рода иксия, 44 вида которых распространены главным образом в засушливых районах Южной Африки. Много красивоцветущих декоративных растений в роде лаперузия (Lapeirousia) с 30 видами, из которых лаперузия ароматнейшая (L. odoratissima) из Анголы имеет белые, сильно душистые цветки с очень длинной трубкой околоцветника (рис. 102, 1-4), а также в родах уотсония с 70 видами, тритония (Tritonia) с 40 видами и крокосмия (Crocosmia) с 7 видами.
К трибе шафрановых принадлежат 3 рода: ромулея с приблизительно 90 видами, распространенными в Средиземноморье, в горных районах тропической Африки и в Южной Африке, сирингодея с 10 южноафриканскими видами и шафран с 80 видами, распространенными в Южной Европе, Средиземноморье и Юго-Западной Азии. Эти растения габитуально очень отличаются от иксиевых, однако имеют с ними настолько близкое родство, что их нередко к ним присоединяют. Для шафрановых особенно характерна сильная редукция стебля и соцветия. Ромулея, у которой одноцветковые цветоносы уже во время цветения достигают поверхности земли, а цветки имеют очень короткую трубку околоцветника, занимает промежуточное положение между иксиевыми и остальными шафрановыми. Обычно этот род принимают за предковую группу по отношению к шафрановым - очень высокоспециализированным геофитам. Можно отметить, что у шафрана, подобно другому евразиатскому роду ирис, значительно более крупные хромосомы, чем у ромулеи, сирингодеи и родов трибы иксиевых.
Хотя виды ромулеи также встречаются в культуре, наибольшее хозяйственное значение из родов трибы имеет шафран. Один из видов этого рода - шафран посевной (Crocus sativus) принадлежит к числу древнейших культивируемых растений и в диком состоянии неизвестен (рис. 100, 1-2). Возможно, это гибрид близких дикорастущих видов, что подтверждается его постоянной стерильностью. Высушенные рыльца этого растения очень ароматичны, имеют сладковатый вкус и с глубокой древности использовались как пряность, а также в парфюмерии и медицине. Многие рецепты лекарств древних авторов содержат шафран в качестве составной части. Кроме того, рыльца шафрана - отличный краситель для пищевых продуктов: 1 г шафрана окрашивает 100 л воды в желтый цвет. Рыльца шафрана очень богаты каротиноидами, содержат также глюкозид пикрокрозин и эфирное масло. Шафран посевной принадлежит к осеннецветущим видам, поэтому его рыльца собирают на плантациях обычно в октябре, причем на 1 кг сухой массы идут рыльца со 150 000 цветков. Наибольшим спросом шафран пользуется в арабских странах, однако его основные плантации в настоящее время находятся в Испании, куда он был завезен маврами, и Южной Франции. Культивируют его и на юге СССР, где испытывали для культуры и другие, близкородственные виды шафрана, в частности распространенный в Крыму и Малой Азии шафран Палласа (С. pallasii).
В качестве декоративных растений чаще используют весеннецветущие виды шафрана с желтыми цветками: шафран желтый (С. flavus), золотистоцветковый (С. chrysanthus) и узколистный (С. angustifolius), а также виды с лиловыми, голубыми и белыми цветками, обычно шафраны весенний (С. vermis) и Томаса (С. to- masianus). Из дикорастущих видов на степных склонах юга европейской части СССР ранней весной можно встретить шафран сетчатый (С. reticulatus) с бледно-лиловыми снаружи и 3 фиолетовыми полосками цветками (рис. 100, 3). Очень красивы осеннецветущие виды с лилово-розовыми цветками - шафран банатский (С. banaticus), заходящий на Карпаты в пределах СССР, и шафран прекрасный, распространенный в Крыму, на Кавказе и в Юго-Западной Азии (рис. 100, 4). У этих растений вполне развитые листья и плоды появляются только на следующее лето. В настоящее время существует много садовых разновидностей шафранов, часто имеющих гибридное происхождение.
Хозяйственное значение ирисовых прежде всего заключается в их высокой декоративности. Ирисы, гладиолусы и крокусы культивируют сейчас почти во всех внетропических странах. Все шире используют в садово-парковом хозяйстве многих стран мира и другие красивоцветущие ирисовые из родов фреезия, тигридия, иксия, тритония, беламканда, морея, лаперузия и др. Другие стороны использования человеком представителей этого семейства менее существенны. Очищенные от коры и высушенные на воздухе корневища ирисов германского, бледного и флорентийского используют в Средней и Южной Европе как лекарственное и антисептическое средство под названием "фиалкового корня" (из-за приятного запаха, напоминающего запах фиалки). В Северной Италии эти виды культивируют уже более 250 лет. Наконец, клубни и луковицы многих ирисовых, например тигридии, шафрана, тритонии, использует в пищу местное население, главным образом в печеном виде.
Семейство гемодоровые (Haemodoraceae) (Л. И. Абрамова)
Семейство гемодоровых представлено 15-16 родами и примерно 75 видами, распространенными в тропиках и субтропиках Австралии, Африки и Америки.
Большинство родов ограничено в своем распространении одним континентом. В Австралии - наибольшее число родов гемодоровых. Здесь растут анигозантос (Anigozanthos, рис. 107, табл. 27, 7), коностилис (Conostylis), гемодорум (Haemodorum, рис. 108), трибонантес (Tribonanthes), флебокария (Phlebocarya). Гемодорум встречается, кроме Австралии, на островах Новая Гвинея и Тасмания. Для Тасмании характерен узкоэндемичный гемодорум двуряднолистный (Haemodorum distichopliyllum). На юге Африки, в Капской области, произрастают эндемики - вахендорфия (Wachendorfia) и дилатрис (Dilatris). Африканский род барберетта (Barberetta) обитает на востоке Южно-Африканской Республики. Из Южной и Центральной Америки известны монотипных рода - ксифидиум (Xiphidium, рис. 108), хагенбахия {Hagenbachia), пиррориза (Pyrrorhiza) и шикия (Schiekia). В Северной Америке встречаются 2 рода - лофиола (Lophiola) и монотипный лахнантес (Lachnanthes). Они распространены вдоль Атлантического побережья от Флориды до штата Массачусетс (США) и полуострова Новая Шотландия (Канада), где и проходит северная граница распространения семейства гемодоровых.

Таблица 27. Гемодоровые, триллиевые и такковые: 1 - анигозантос желтоватый (Anigozanthos flavidus), оранжереи Ботанического института АН СССР в Ленинграде; 2 - даисва хайнаньская (Daiswa hainanensis), там же; 3 - триллиум камчатский (Trillium kamschatcense), сад Ленинградского государственного университета; 4 - вороний глаз, или парис обыкновенный (Paris guadrifolia), Ленинградская область; 5 - такка цельнолистная (Тасса integrifolia), оранжереи Ботанического института АН СССР в Ленинграде
Ограниченность ареалов и труднодоступность многих местопроизрастаний отдельных представителей гемодоровых объясняет слабую изученность некоторых из них. Барберетта золотистая (Barberetta aurea), описанная еще в 18G8 г., только к 1971 г. была подробно исследована на живом материале. В состав семейства входит такое замечательное растение, как анигозантос (рис. 107). Форма цветка у представителей этого рода очень причудлива и напоминает лапу животного. Опушение придает цветкам бархатистость, а концы сегментов околоцветника, чуть загнутые внутрь, похожи на когти. Этим внешним сходством объясняется местное австралийское название рода "кенгуровая лапа". Анигозантос Менглза (Anigozanthos manglesii) - эндемик Юго-Западной Австралии. В 1960 г. он стал ботанической эмблемой штата Западная Австралия.
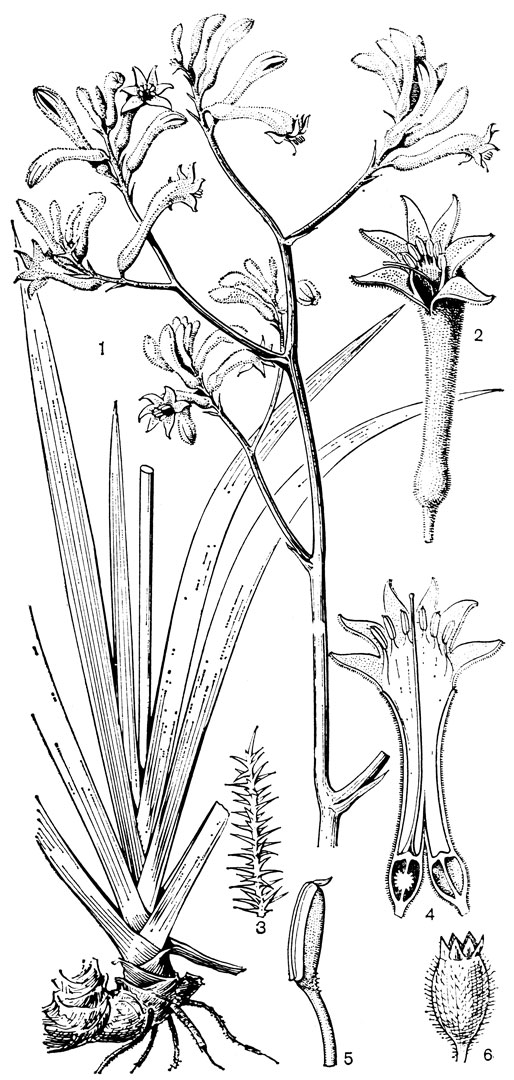
Рис. 107. Анигозантос желтоватый (Anigozanthos flavidus): 1 - общий вид растения; 2 - цветок; 3 - волосок с поверхности околоцветника; 4 - цветок в разрезе; 5 - тычинка; 6 - плод
Многие представители гемодоровых являются обычными компонентами лесов. В вечнозеленых эвкалиптовых лесах Австралии растут многочисленные виды анигозантоса и коностилиса. Гемодорум щитковидный (Haemodorum corymbosum, рис. 108, 1) встречается в австралийских прибрежных саванновых лесах вместе с мелалеукой (Melaleuca) и банксией (Banksia). Обильные заросли образует барберетта золотистая вместе с циртантусом (Cyrtanthus) на влажных лесных просеках Южной Африки.
Некоторые виды коностилиса и тасманийский гемодорум двуряднолистный считаются типичными обитателями верещатников, а лахнантес и лофиола заселяют влажные сосняки Северной Америки. Гемодоровые приспособились к большому разнообразию экологических условий. С одной стороны, это болота, болотистые земли и влажные берега ручьев,- типичные для североамериканских лахнантеса, лофиолы и видов южно-африканской вахендорфии. С другой стороны, сухие склерофильные леса Юго-Западной Австралии обычны для большинства видов коностилиса. Особые климатические условия в тех географических областях, где встречаются некоторые представители семейства, например в тропиках Северной Австралии с ее сильно различающимися по количеству осадков сезонами года, требуют приспособленности к резкой смене режимов увлажнения. Растения гемодорума щитковидного в сухой сезон подвергаются действию сильной засухи, когда серые глиноземы, на которых они растут, высыхают и сжимаются, а в период влажного сезона они оказываются погруженными в воду почти на 10 см. Даже в культуре анигозантос нуждается в обилии воды только в период активной вегетации и цветения, а в период покоя излишнее увлажнение приводит к загниванию корневищ.
Гемодоровые - травянистые многолетники с короткими, иногда клубневидными корневищами. Листья у них двурядные, линейные или мечевидные, с влагалищным основанием, унифациальные. Пластинка листа обычно сжата с боков, как у ирисов. Листья образуют приземную розетку, из которой выходит облиственный стебель, несущий слабо развитые стеблевые листья, иногда редуцированные до чешуек, и заканчивающийся соцветием. Основание стебля или чаще корневище короткое и толстое, может приобретать вид луковицы благодаря сохраняющимся вокруг него буреющим влагалищам отмерших листьев. Такие ложные клубнелуковицы имеются у вахендорфии и трибонантеса. Влагалища листьев служат хорошей защитой корневищ от пожаров в сухой период года и, как пишет австралийский ботаник Д. Грин (1960), молодые побеги некоторых видов коностилиса после пожаров не выглядят пострадавшими и даже активнее растут, создавая большую вегетативную массу. Корни пучковатые, волокнистые, иногда довольно толстые и губчатые, нередко окрашенные в красный цвет. У некоторых представителей семейства корневища образуют столоны. Сосуды обычно с простой, реже (у трибонантеса, дилатриса и лофиолы) с лестничной перфорацией, находятся в корнях, а у лахнантеса сосуды есть и в надземных частях. Гемодоровые имеют очень различные размеры. Одни довольно крупные, как растения из родов вахендорфия, высотой 1-2 м; другие, как, например, эндемичный для острова Тасмания гемодорум двуряднолистный, не превышают 4-8 см. Растения могут быть голыми или опушенными. Опушены в основном цветоносы и особенно околоцветники, реже опушение распространено и на другие части растения. Цветки в различного рода соцветиях, обоеполые, актиноморфные или иногда слабозигоморфные. Околоцветник состоит из 6 лепестковидных сегментов, расположенных в 1 или 2 круга. Сегменты свободные или сросшиеся в трубку (анигозантос, коностилис, трибонантес), короткую или длинную, прямую или согнутую. Цветовая гамма околоцветника необычайно разнообразна, от белых цветков у ксифидиума до почти черных у гемодорума колосистого (Haemodornm spicatnm) и анигозантоса грязно-бурого (Anigozanthos fnliginosus). Тычинок 6 или 3, очень редко 1; иногда 1 или 2 превращены в стаминодии; 6 тычинок характерны для австралийских родов флебокария, аигозантос, коностилис и трибонантес, для североамериканского рода лофиола. Остальные представители семейства имеют по 3 тычинки, а у монотипного рода пиррориза из Южной Америки - только 1 тычинка и 2 стаминодия. Тычиночные нити длинные, свободные или сросшиеся с околоцветником. Пыльники интрорзные, открывающиеся продольной щелью. Гинецей из 3 плодолистиков. Столбик нитевидный, короткий или длинный, с маленьким 1-3-головчатым рыльцем. Завязь верхняя у вахендорфии и ксифидиума; полунижняя - у флебокарии и коностилиса; нижняя - у лахнантеса, дилатриса и др. Иногда завязь бывает в цветках полунижняя, а в плодах становится нижней. Обычно завязь 3-гнездная, за исключением флебокарии, с париетальной плацентацией. У барберетты гинецей псевдомономерный; фертилен только 1 плодолистик, а 2 других стерильны и рано редуцируются, так что завязь у нее тоже становится как бы одногнездной и уплощается с одной стороны. Только 1 гнездо функционирует в завязи дилатриса щитковидного (Dilatris corymbosa), в 2 других зародыши недоразвиты. Семязачатки ортотропные или гемитропные, от одиночных в каждом гнезде (барберетта) до многочисленных (ксифидиум, анигозантос). У гемодорума К. Ван Стенис (1954) обнаружил по 2 семязачатка в гнезде, в то время как некоторые авторы ошибочно указывают 3, принимая за третий выступ утолщенную плаценту, к которой прикреплены семязачатки. Септальные нектарники гемодоровых часто слабо развиты. Плод - обычно локулицидная коробочка. В редких случаях он ореховидный, нераскрывающийся (флебокария, барберетта), может иметь кожистую или мясистую консистенцию. Семена обычно черные (у барберетты - оранжевые) и у разных родов различаются величиной и формой. Зародыш в семени маленький, с обильным эндоспермом.
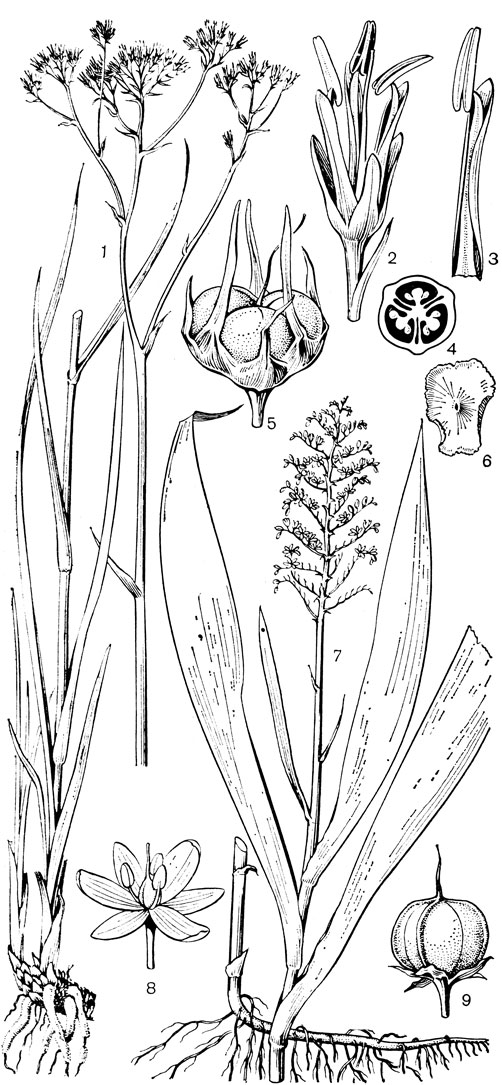
Рис. 108. Гемодоровые. Гемодорум щитковидный (Haemodorum corymbosum): l - общий вид; 2 - цветок; 3 - сегмент околоцветника с тычинкой; 4 - поперечный разрез завязи; 5 - плод; 6 - семя. Ксифидиум голубой (Xiphidium coeruleum): 7 - общий вид; 8 - цветок; 9 - плод
Большинство гемодоровых опыляется насекомыми, которых привлекает нектар, выделяемый септальными нектарниками. Цветки растений рода гемодорум обычно пахучие. У вахендорфии и барберетты венчиковидный околоцветник воронковидный, желтого цвета, имеет оранжевые пятна, четко выделяющиеся на 3 верхних сегментах, что может привлекать насекомых. Такую же роль у лиловых цветков дилатриса щитковидного может играть оранжевая окраска крупных пыльников двух верхних тычинок. Цветки вахендорфии посещают пчелы и другие насекомые. Длиннотрубчатые цветки анигозантоса опыляют только птицы. По мнению ряда исследователей, посещение цветков птицами стимулируется наличием обильного нектара и яркой, чаще всего красной, окраской околоцветника. В то же время значи-тельная длина трубки и опушенность внутренних частей цветка отпугивает насекомых. Пыльники у цветков анигозантоса расположены открыто в верхней части околоцветника на расстоянии примерно 80 мм от нектара. По-этому посещающие их маленькие медососы (Acanthorhynchus) уносят пыльцу на спинке, а у более крупных медососов (Anthochaera саrunculata) пыльца прилипает к перьям на шее и голове. С. Хоппер и А. Бербидж (1978) пришли к выводу, что в зависимости от размера цветков и расположения в них пыльников (авторы изучали два вида - анигозантос Менглза и анигозантос приземистый - Anigozantlios humilis) пыльца попадает на разные части тела птицы. Этим достигается частичная изоляция видов в природе.
Мясистая консистенция и пурпурно-коричневая окраска коробочек ксифидиума привлекают птиц. Высохший, остающийся при плодах околоцветник дилатриса щитковидного, вероятно, играет роль легкого крыла при переносе их ветром. У многих представителей семейства плоды имеют густое опушение из волосков, что может способствовать их переносу при прикреплении к движущимся предметам. Семена гемодоровых бывают двух типов. Одни из них дисковидно сплющены, часто с крыловидным краем, гладкие с поверхности; для них не исключено распространение с помощью ветра. Поверхность других неровная и покрыта шипиками и бородавчатыми выростами, они могут легко прилипать к одежде людей и шерсти животных.
Семейство гемодоровых делится на две трибы - гемодоровые (Haemodoreae) и коностилисовые (Conostylideae), различающиеся главным образом типом околоцветника и числом тычинок. Некоторые авторы считают их подсемействами. У видов трибы собственно гемодоровых сегменты околоцветника обычно расположены в 2 круга, трубка почти отсутствует, тычинок 3 (реже 6, 1), пыльцевые зерна однобороздные. В эту трибу входит 10-11 родов, самый многочисленный из них гемодорум (около 20 видов). Виды гемодорума распространены в Австралии, особенно в большом числе на северо-западе страны. На востоке континента они распространены до Нового Южного Уэльса, а на западе до южного побережья, встречаются на Тасмании, островах Меланезии и Новой Гвинее. Цветки гемодорума актиноморфные, довольно мелкие, полузакрытые в фазу цветения, голые, с различающимися по величине сегментами околоцветника. Наружные сегменты более короткие и несколько прикрывают внутренние в нижней части цветка. Края внутренних сегментов завернуты внутрь и охватывают тычиночные нити. Тычинки длиннее сегментов и, так же как и столбик, выступают из цветка. Цветки собраны в метельчатое соцветие, которое иногда за счет укороченных цветоножек приобретает вид колоса или головки. Завязь нижняя и полунижняя.
Совершенно иной тип цветка имеет южноафриканская вахендорфия (5 видов). Цветки ее слабозигоморфные, относительно крупного размера, с венчиковидным околоцветником воронковидной формы, обычно желтого цвета. Тычиночные нити длинные, тычинки и столбик выступают из цветка. Снаружи околоцветник покрыт железистыми волосками. Асимметрия цветка выражена в большой степени и не столько проявляется в разных размерах сегментов, сколько в неодинаковом отклонении вбок тычинок и столбика. Особое своеобразие цветкам вахендорфии пирамидальной (Wachendorfia thyrsiflora) придают выделяющиеся на общем желтом фоне оранжевые пятна в основании трех верхних сегментов околоцветника. Завязь у всех видов верхняя. Для вахендорфии характерно образование ложной клубнелуковицы. Такой же тип корневища имеет барберетта золотистая, вегетативное размножение которой изучили английские ботаники О. Хиллиард и Б. Бартт (1971). У барберетты цветущий стебель выходит из подземного корневища, закладывающего два зачатка столонов, которые после окончания вегетации начинают расти в стороны от родительского растения и, разрастаясь, образуют каждый новую ложную клубнелуковицу. Столоны сгнивают, и тем самым нарушается связь между молодыми и старым растениями. Такое активное увеличение числа особей вполне объясняет численность природных популяций барберетты. Подобным образом происходит вегетативное размножение и у вахендорфии.
Интересным в морфологическом плане представляется и монотипный род ксифидиум (рис. 108), распространенный в Южной и Центральной Америке, от Боливии и Бразилии до Мексики и на острове Куба. У ксифидиума голубого (Xiphidium coeruleum, рис. 108, 7) цветки мелкие, актиноморфные, широко раскрывающиеся. Пыльники прикреплены к свободным тычиночным нитям, которые короче сегментов околоцветника, столбик короткий, завязь верхняя. Коробочка в период созревания становится мясистой и меняет оранжевую окраску на пурпурно-коричневую. Вегетативное размножение у растений ксифидиума происходит за счет придаточных почек, образующихся на стебле под соцветием. Опадая на землю, они дают начало новым побегам.
У представителей трибы коностилисовых сегменты околоцветника почти створчатые, расположены в один круг, трубка хорошо выраженная, часто длинная, иногда искривленная, тычинок 6, пыльцевые зерна 2-8-поровые. Все роды, принадлежащие к этой трибе, австралийские; из них самый многочисленный - коностилис (около 23 видов). Этот род распространен на небольшой территории в Юго-Западной Австралии. Цветки различных видов варьируют по форме и размерам околоцветника. Как показали исследования Д. Грина (1960), у видов коностилис обнаружены трубчатые, колокольчатые и свободнолепестные цветки. Последние характерны для коностилиса короткоплодного (Conostylis breviscarpa). Тычинки и столбик имеют одинаковую величину с сегментами околоцветника или короче их. В качестве особой закономерности в структуре цветков можно отметить то, что в более открытых типах цветка тычинки и столбик короче сегментов околоцветника, а в трубчатых они длиннее сегментов. Плохое завязывание семян у коностилиса компенсируется существованием нескольких форм вегетативного размножения (Д. Грин, 1960): отпрысковый (пролиферирующий) тип роста, когда стебель может многократно ветвиться и образовывать в узлах пучки листьев и цветущие стрелки, и отводковый тип, при котором развиваются столоны, дающие начало новым побегам. Некоторые растения, имеющие короткие корневища, образуют плотные дернины. Как выяснилось, многие виды коностилиса обладают этими формами роста в различных комбинациях. У анигозантоса - второго по количеству видов в трибе коностилисовых (10 видов) - цветок зигоморфный, с длинной, несколько искривленной и расщепленной трубкой.
Растения, принадлежащие к семейству гемодоровых, часто используют в садово-парковой культуре и выращивают в открытом грунте и теплицах, поскольку в основном их относят к тропическим видам. В Австралии анигозантос обычно применяют для создания естественных ландшафтов в частных садах и общественных парках. Особенно широкое распространение получил анигозантос желтоватый (Anigozanthos flavidus, рис. 107, табл. 27, 1) благодаря вегетативному размножению корневищами, сравнительно хорошему семенному воспроизведению и устойчивости к ряду заболеваний, которым подвержены другие виды. В настоящее время активно проводятся опыты по получению гибридов и полиплоидов этого хорошо зарекомендовавшего себя вида за счет скрещивания его с другими дикими видами, отличающимися красивыми цветками,- анигозантосом Менглза, анигозантосом красивейшим и др. Гибридизация позволяет получать новые декоративные формы с самыми разнообразными цветовыми сочетаниями в окраске околоцветника. Более длительного периода цветения в местных условиях добиваются, выращивая растения в теплицах. В оранжереях можно встретить таких представителей семейства, как гемодорум, вахендорфия, ксифидиум. Ксифидиум в Центральной Америке выращивают в грунте, и за крупные веерообразно расположенные листья в Эквадоре его называют "пальмой севера". Высокие декоративные качества некоторых видов гемодоровых являются причиной их исчезновения в дикой природе, поэтому в Австралии они находятся под особой защитой. Анигозантос Менглза сохраняется в Королевском парке (город Перт), основную часть которого составляет естественный лесной массив. За незаконный сбор растений взимается крупный денежный штраф.
Многие гемодоровые содержат красный сок в корневищах, клубнелуковицах, а иногда и в корнях, цвет которого обусловливает содержание полифенольного красного пигмента. Сок лахнантеса и вахендорфии используют для приготовления краски. В корнях и побегах лахнантеса находятся вещества, применяемые в фармакологии. Из гемодорума щитковидного в Квинсленде (Австралия) добывают яд. Жареные корневища некоторых других видов этого рода австралийские аборигены употребляют в пищу, а из длинных жестких листьев получают прочные волокна для плетения сумок.
Семейство гипоксисовые (Hypoxidaceae) (В. Н. Гладкова)
В семейство входят 8 родов и около 150 видов трав, большинство которых растет в южном полушарии и тропической Азии. Особенно богата родами гипоксисовых Южная Африка. Здесь обособились такие некрупные по числу видов эндемичные роды, как родогипоксис (Rliodohypoxis, 6 видов), эмподиум (Empodium, 10 видов), павридия (Pauridia, 1 вид), спилоксена (Spiloxene, 23 вида), саниелла (Saniella, 1 вид). В Африке достигает наибольшего видового разнообразия и самый крупный в семействе род - гипоксис (Hypoxis, 100 видов), виды которого растут также на Маскаренских островах, в Японии и Южной и Юго-Восточной Азии, доходя до острова Тайвань, Филиппин, Австралии, Новой Зеландии и Тасмании, на обоих континентах Америки (от Атлантического побережья США через Мексику до Уругвая), в Вест-Индии. Остальные 2 рода - молинерия (Molineria, 5 видов) и куркулиго (Curculigo, 5 видов) - обитатели тропических областей обоих полушарий. Куркулиго представлен в Южной Азии, доходит до Филиппин и Явы, встречается на Сейшельских островах, в тропической Америке и Вест-Индии, один вид заходит в тропическую Африку. Молинерия имеет центр разнообразия видов в Юго-Восточной Азии, заходит на Новую Гвинею и на Азиатский континент до Восточных Гималаев (штат Сикким, Индия) и Юго-Западного Китая (Юньнань), а в Африке не встречается совсем.
Гипоксисовые - лишенные надземных стеблей травы с прикорневыми или розеточными листьями. Многие из них - геофиты, надземные части которых появляются только на короткое время дождей. Виды гипоксисовых либо имеют вертикальное утолщенное корневище, которое на своей верхушке ежегодно формирует новые листья, либо образуют настоящую туникатную клубнелуковицу, которая нарастает ежегодно на сморщенной клубнелуковице предыдущего года. Удлиненное вертикальное корневище характерно для родов гипоксис, куркулиго, молинерия; более плотное и сжатое корневище у видов родогипоксиса, саниеллы. Клубнелуковицу формируют некоторые американские и австралийско-новозеландские виды гипоксиса, африканские роды эмподиум, павридия, спилоксена. Покровы из старых листовых влагалищ, окружающие подземные органы, могут либо срастаться с клубнем, либо полностью или частично отделяться от него. Иногда эти покровы отсутствуют, клубнелуковица бывает оплетена только корнями. Корни гипоксисовых неветвящиеся, обычно короткие, но иногда проникающие в почву на глубину до 15 см и достающие влагу из ее глубоких слоев. Листья сидячие или черешковые, часто с выдающимися жилками, иногда складчатые, от удлиненно-яйцевидных до ланцетных и линейных, цельнокрайные или редко с отогнутыми назад зубцами по краю. Сосуды с лестничной перфорацией, имеются только в корнях. В пластинке листа у многих видов имеются слизистые каналы, расположенные над сосудистым пучком или рассеянные в мезофилле, в котором встречаются также клетки с рафидами - пучками игловидных кристаллов.
Кистевидные или неправильно зонтиковидные соцветия или одиночные цветки выносятся на пазушной стрелке разной величины, иногда очень короткой. У молинерии соцветие головчатое, с множеством прицветников. Цветки актиноморфные, обоеполые. У одного африканского вида куркулиго верхние цветки в соцветии могут быть функционально мужскими. Околоцветник желтый, белый или красный, состоит из 6 почти одинаковых или различающихся (родогипоксис) сегментов, у основания сросшихся в трубку (павридия, родогипоксис, саниелла) или свободных (остальные роды); сегменты околоцветника простертые (цветки напоминают звездочки). Тычинок 6 или 3 (павридия), прикрепленные к основанию околоцветника. По строению тычинок роды семейства распадаются на две группы. У молинерии, эмподиума, родогипоксиса и саниеллы нить соединяется с пыльником на его спинке с наружной стороны (рис. 109, 110), связник при этом сильно утолщен и тычинка имеет разный облик с наружной и внутренней стороны. У остальных родов пыльник симметричен в поперечном сечении, а нить присоединяется к нему в выемке между его базальными долями (качающийся подвижный пыльник). Открываются пыльники продольными щелями интрорзно, экстрорзно или латрорзно. У некоторых видов (эмподиум) выражен надсвязник, иногда пыльники бывают расщепленными наверху (куркулиго). Пыльцевые зерна однобороздные, с тонкой сетчатой экзиной. Завязь у гипоксисовых нижняя, 3-гнездная или 1-гнездная (эмподиум, спилоксена), с немногими или многочисленными анатропными или гемитропными семязачатками. Столбик с головчатым или 2-3х-лопастным рыльцем. Завязь некоторых видов гипоксисовых (виды родогипоксиса, куркулиго, эмподиума) имеет характерный, свойственный и некоторым другим однодольным носик, или клювик. Это длинное цилиндрическое образование на верхушке завязи (рис. 109, 6, 7), благодаря которому завязь и остальные части цветка оказываются отделенными друг от друга, околоцветник выносится вверх, а завязь остается скрытой в листовых влагалищах и иногда даже под землей, получая, таким образом, дополнительную защиту от воздействия неблагоприятных условий. Клювик похож на узкую трубку венчика, но он сплошной и возник, по-видимому, в результате срастания околоцветника со столбиком (Г. Нел, 1914). У некоторых видов в клювике можно также обнаружить и следы тонких тычиночных нитей. Плод - коробочка, большей частью с сохраняющимся околоцветником, неправильно разламывающаяся на части или открывающаяся близ верхушки опоясывающей поперечной щелью или продольными щелями. У куркулиго и некоторых других видов плод мясистый, нераскрывающийся. Семена темные, шаровидные, хрупкие, с обильным эндоспермом, с выступающим клювовидным рубчиком и сохраняющейся ножкой, покрыты, включая ножку, темной оболочкой. Оболочка семени почти гладкая (у некоторых куркулиго, гипоксисов) или, чаще, с выраженной скульптурой (шиповатой, струйчатой, бугорчатой), характер которой является систематическим признаком для разграничения видов. У эмподиума семена снабжены карункулой.
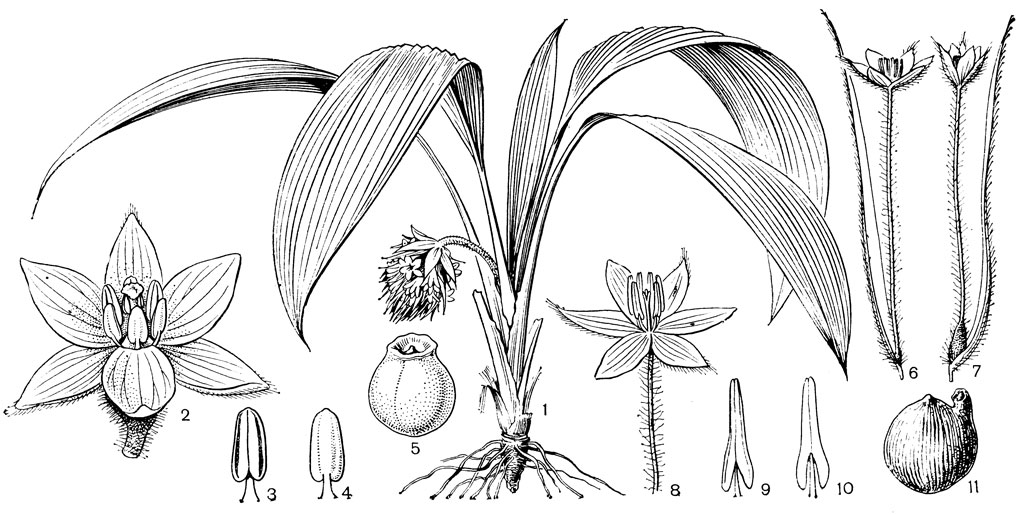
Рис. 109. Гипоксисовые. Молинерия головчатая (Molineria capitulata): 1 - общий вид; 2 - цветок; 3, 4 - тычинка в двух положениях; 5 - плод. Куркули го орхидный (Curculigo orchioides): 6, 7 - цветок с завязью, снабженной длинным клювом: 8 - цветок без завязи; 9, 10 - тычинки в двух положениях; 11 - семя с выступающими ножкой и рубчиком
Многие гипоксисовые являются ксерофильными растениями и встречаются на открытых солнечных участках мелкотравных пространств, часто выжигаемых, на неглубокой почве на скалистых или каменистых местах, на кремнеземах и на песках, на задернованных участках горных склонов, поднимаясь в горы на высоту свыше 3000 м над уровнем моря. Реже они растут в тенистых или влажных местах, на прибрежных, иногда засоленных болотах, на влажных песчаных участках, по травянистым берегам рек, среди дерновинных злаков и осок и даже в мелководных пересыхающих водоемах. Некоторые виды встречаются большими скоплениями и составляют существенную часть травянистых формаций. Спилоксена и павридия растут в области зимних дождей, и именно в этот период происходит их основной рост и развитие: летом эти растения находятся в покоящемся состоянии. Саниелла и родогипоксис свой жизненный цикл завершают летом, произрастая в области летних дождей; они цветут и плодоносят в ноябре - феврале. Виды гипоксиса и некоторых других родов, произрастающие на открытых местах с дефицитом влаги, запасают воду в клубнях. Клубни их в периферической зоне содержат мягкую ткань, пронизанную слизистыми каналами и заполненную желтым соком. У некоторых видов гипоксиса они достигают в диаметре 10 см. Клубни прочно удерживают растения в почве при сильных ветрах и способствуют переживанию ими случающихся пожаров. Растения влажных мест обычно развивают небольшие клубни без водозапасающей ткани или не имеют их вообще, как некоторые влаголюбивые виды спилоксены. От условий произрастания зависят также размер листьев и развитие на них опушения как защиты от излишней транспирации. Сильно опушенными и очень узкими, почти нитевидными бывают листья многих ксерофитных видов гипоксиса.
Гипоксисовые перекрестноопыляемые растения. У родогипоксисов (рис. 110, 1) внутренние сегменты околоцветника, в отличие от внешних, имеют длинные ноготки, которые изогнуты внутрь цветка так, что закрывают зев околоцветника и цветок кажется слепым, недоступным для насекомых. Но именно благодаря такому изгибу простертые пластинки внутреннего и наружного кругов околоцветника оказываются расположенными на разных уровнях и над основанием каждого наружного сегмента околоцветника и между ноготками внутренних возникают маленькие латеральные отверстия, через которые и проникают насекомые. Интересные наблюдения, касающиеся опыления, приводит во "Флоре Южной Африки" Р. Марлот (1915). Цветки гипоксиса звездчатого (Нуpoxys stellata), опыляемые жуками, варьируют по окраске от желтых до белых и имеют нередко более темную, каштановую или черную середину, а в самом центре у некоторых экземпляров расположено еще зеленоватое пятно с металлическим отливом. Зеленый цвет определяется здесь, однако, не зеленым пигментом, который отсутствует, а особым строением эпидермы, клетки которой в центре цветка вытянуты в конические папиллы, заполненные бесцветными неправильной формы гранулами (из жиров) с большой преломляющей способностью. В результате интерференции лучей света, проникающих в зернистую массу, и возникает зеленый оттенок цветка. Этот оттенок привлекает жуков, причем особенно тех видов, у которых надкрылья также имеют серо-зеленый оттенок.
Цветки многих гипоксисов открываются только в солнечные и светлые дни, благоприятные для опыления. В открытые цветки проникают мелкие пчелы и другие некрупные насекомые, привлекаемые отделяемой в основании цветка сладкой жидкостью. Насекомые ползают как по рыльцу, так и у основания тычинок, перенося пыльцу. У некоторых гипоксисов, например американского гипоксиса сидячего (Н. sessilis), бывают клейстогамные неоткрывающиеся цветки.
Распространение семян определяется положением цветка на растении. Если завязь без клюва и цветок развивается на прямом цветоносе, сформировавшаяся из него коробочка обычно раскрывается поперечной щелью, ее верхняя часть и увядший околоцветник падают в сторону и семена постепенно выпадают при сотрясении цветоножки ветром или пробегающими мимо животными. У видов, имеющих завязь с клювом, спрятанную в листовом влагалище близко к поверхности почвы, хрупкие коробочки по мере созревания приподнимаются над влагалищем на чуть удлиняющемся цветоносе и здесь распадаются на части, падающие на землю вместе с семенами. У родогипоксиса отогнутого (Rhodohypoxis deflexa), по наблюдениям О. М. Гильярда и Б. Л. Бёрта (1978), коробочка сначала растрескивается поперечной щелью на верхушке, а затем оставшаяся ее часть еще расщепляется тремя закручивающимися вниз створками, освобождая семена.
У некоторых видов, имеющих длинный клюв на завязи, коробочки заведомо спрятаны в поверхностных слоях ила или в почве. У растений влажных местообитаний свободное рассеивание семян утратилось и выработались приспособления к их захоронению поблизости от материнского растения. Ножка цветка или стрелка у таких растений изгибается вниз, и завязь, а затем плод, легко разламывающийся и тонкостенный, лежат на поверхности болотистой почвы. Это наблюдается у гипоксиса стелющегося (Hypoxis decumbens, рис. 110, 7) и других американских видов гипоксиса, у павридии, видов спилоксены, родогипоксиса и др. Сочные и сладковатые на вкус коробочки молинерии, возможно, поедаются некоторыми грызунами, которые их уносят, теряя по дороге и тем самым рассеивая семена.
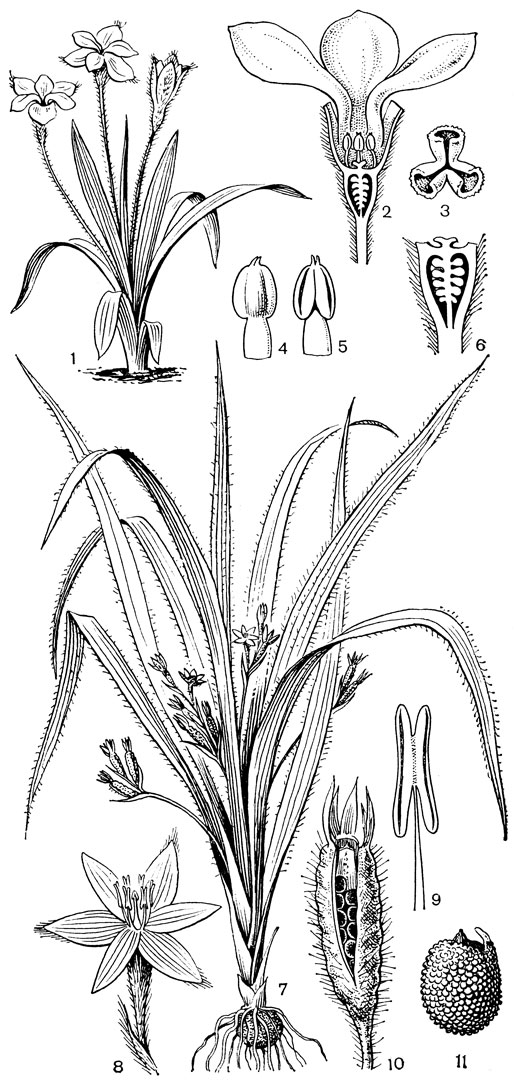
Рис. 110. Гипоксисовые. Родогипоксис Баурa (Rhodohypoxis baurii): 1 - общий вид; 2 - цветок в разрезе; 3 - рыльце; 4, 5 - тычинка в двух положениях; 6 - продольный разрез завязи. Гипоксис стелющийся (Hypoxis decumbens): 7 - общий вид; 8 - цветок; 9 - тычинка; 10 - зрелая коробочка; 11 - семя
Семена гипоксисовых очень легки и, не смачиваясь, плавают в воде, которая тоже может способствовать их распространению.
Семейство гипоксисовых не отличается большим разнообразием форм. Наиболее часто встречающимися и широко распространенными по всему ареалу семейства являются виды рода гипоксис (Hypoxis) - невысокие, обычно опушенные травы с желтыми или белыми цветками в соцветиях, внешне напоминающие гусиный лук. Они развивают от 6 до 20 листьев, от почти нитевидных или линейных у большинства видов до довольно крупных у некоторых видов, достигающих в длину 50 см и в ширину 6 см. Листья, как правило, с шелковистым, войлочным или рассеянным опушением, полоски бывают звездчатыми или кустистыми. Цветки появляются раньше листьев или одновременно с ними.
Второй по числу видов род - спилоксена (Spiloxene) - ограничен в своем распространении юго-западом Капской области. Это клубнелуковичные невысокие травы с довольно мясистыми, узкими или сложенными пополам, голыми листьями и соцветиями оранжевых, желтых или белых цветков. У некоторых видов, растущих в тенистых местах или на болотах, листья тонкие и более широкие, а клубнелуковица заменена системой толстых боковых корней.
К африканскому роду родогипоксис (Rhodohypoxis) также принадлежат мелкие травы с удлиненно-клубневидными корневищами, одетыми щетинисто-волокнистыми беловатыми остатками старых листьев. Цветки белые, розовые или красные, одиночные или в парах, на длинных превосходящих по длине листья цветоносах. Виды родогипоксиса часто встречаются крупными популяциями, распределенными по микроместообитаниям. Миллионы экземпляров насчитывают на травянистых суховатых участках по скалистым склонам гор в популяциях родогипоксиса Баура (R. baurii, рис. 110, 1). Сплошной ковер из простертых горизонтально листьев, над которыми рассыпаны звездочки розовых или красных цветков, образует на болотистых местах родогипоксис отогнутый (R. deflexa). Явную приверженность к проточной воде обнаруживает родогипоксис миллоподобный (R. milloides), поселяясь вблизи протекающих водных источников. Мириадами экземпляров встречается в пересыхающих водоемах родогипоксис красноватый (R. rubella); цветущие экземпляры торчат из влажного галечника, заливаемого во время дождей, или стоят в мелкой воде, высыхаемой ко времени плодоношения, и тогда только пучки зеленых листьев видны на голой почве. В Драконовых горах виды родогипоксиса поднимаются до 3200 м над уровнем моря. Окраска венчика у них варьирует в пределах вида и, кроме того, может меняться с возрастом у отдельных экземпляров от чисто-белой через розовую до ярко-красной. Популяции из таких разноцветных растений представляют собой необычное и красочное зрелище. Иногда популяции распадаются на отдельные цветовые пятна, что определяется присущим родогипоксису вегетативным размножением с помощью подземных столонов. Столоны развивают на своем апикальном конце новые растения, и после этого ось столона загнивает и отмирает, отделяясь от материнского растения. Виды родогипоксиса представляют большой интерес для садоводов как перспективные для культуры декоративные растения.
Саниелла (Saniella) - род, недавно выделенный из родогипоксиса, представлен во флоре Южной Африки единственным видом - саниеллой весенней (S. verna). Это горные растения, похожие на белый шафран, с подземным вертикальным корневищем и одиночными бело-желтыми цветками. Околоцветник у них приподнят на 2-5,7 см на длинном клюве завязи.
Эмподиум (Empodium) также чисто южноафриканский род, произрастающий от Капской области до Наталя и Южного Трансвааля. Относящиеся к роду некрупные травы с туни- катной клубнелуковицей растут одиночно или небольшими дерновинками, формируя ежегодно от одного до нескольких листьев и по 2-3 желтых цветка. Длинные пыльники с надсвязником, семена с карункулой - характерные признаки рода. Эмподиум однолистный (Е. monophyllum) развивает всего по одному листу на клубнелуковице и цветет до его появления.
Куркулиго (Curculigo) и молинерия (Molineria) - корневищные травы с черешковыми крупными листьями и желтыми или белыми цветками. У куркулиго цветки большей частью одиночные, а завязь часто с клювиком, пыльники симметричные в поперечном сечении, семена со струйчатой или почти гладкой тестой. Молинерия очень близка к куркулиго и иногда объединяется с ним. Она характеризуется густыми, иногда головчатыми соцветиями с прицветниками, закругленными на спинке пыльниками, мясистыми нераскрывающимися плодами и бородавчатыми или гладкими блестящими семенами. Оба рода широко распространены в тропической зоне, но представлены небольшим числом видов.
Гипоксисовые выращивают как декоративные растения. Наиболее известна и широко распространена в культуре молинерия головчатая (М. capitulata, рис. 109, 7), выращиваемая ради красивых пальмовидных листьев в горшках в оранжереях или на открытом воздухе в теплых странах. Культивируют также виды гипоксиса и особенно родогипоксиса, пригодного для создания красивоцветущих участков на сухих каменистых почвах в садах и парках. Некоторые родогипоксисы и представители других родов, растущие на пастбищах, имеют небольшое значение и как кормовые растения: их поедают овцы и козы.
Семейство веллозиевые (Velloziaceae) (А. Л. Тахтаджян)
С гемодоровыми и особенно с гипоксисовыми тесно связано и имеет общее с ними происхождение семейство веллозиевых. В этом семействе 6 родов и около 260 видов (Л.В.Смит и Е. С. Айенсу, 1974, 1976). Они распространены в тропической и Южной Африке, на Мадагаскаре, на юге Аравийского полуострова (ксерофита аравийская - Xerophyta arabica) и в аридных областях тропической Америки (главным образом в Бразилии, а также в Панаме, Перу, Боливии и Аргентине). Роды ксерофита (28 видов) и очень близкая к ней талботия (Talbotia), монотипный южноафриканский род, распространены только в Старом Свете, а остальные 4 рода, в том числе самые крупные в семействе веллозия (Vellozia, 122 вида; рис. 111) и барбасения (Barbacenia, 103 вида), не выходят за пределы Неотропического царства (карта 5).
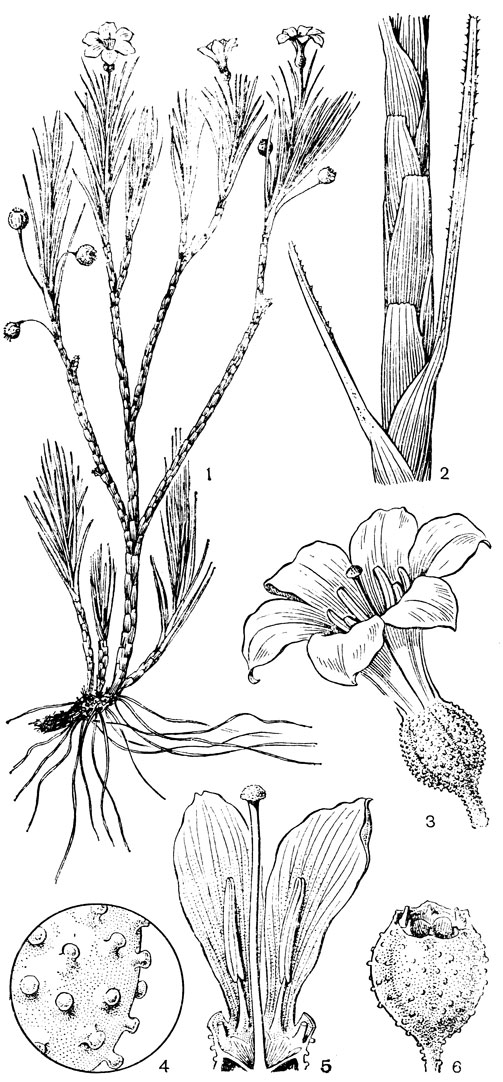
Рис. 111. Веллозия прутьевидная (Vellozia scoparia) 1 - общий вид растения; 2 - часть стебля с листьями; 3 - цветок; 4 - наружняя эпидерма завязи; 5 - продольный разрез цветка; 6 - молодой плод
Карта 5. Ареал семейства веллозиевых
Веллозиевые имеют довольно своеобразный облик, резко отличающий их от знакомых читателю северных однодольных. Стебли веллозиевых деревянистые, простые или, реже, ложнодихотомически ветвящиеся, густо покрытые остающимися старыми листьями или только их волокнистыми влагалищными основаниями, а также длинными воздушными корнями, полностью окруженными листовыми основаниями. Они очень разных размеров, и некоторые бразильские веллозии превышают в высоту 6 м. Листья ксероморфные, кожистые, линейные или узколанцетные, на верхушке большей частью заостренные, дорсивентральные, с хорошо выраженной средней жилкой, молодые скучены у верхушки стебля или ветвей в виде розеток, но старые загибаются вниз и у многих опадают. Устьица обычно парацитные, иногда тетрацитные. Сосуды как в корнях, так и в листьях, но у талботии также в стеблях.
Цветки одиночные, расположены в пазухах верхних листьев, обычно крупные и ярко окрашенные, обоеполые или редко функционально однополые и двудомные (западноафриканский род барбасениопсис - Barbaceniopsis), актиноморфные. Сегментов околоцветника 6, в 2 рядах, большей частью вполне одинаковых, в нижней части вместе с основаниями тычиночных нитей обычно более или менее сросшихся в трубку, но почти свободных у бразильской веллозии Бурлемаркса (V. burlemarxii). Тычинок от 6 до многочисленных (многие виды веллозий). В тех случаях, когда тычинок более 6, они более или менее соединены в 6 плоских пучков, или фаланг, что особенно характерно для большинства видов рода веллозия (интересная параллель с родом гетиллис - Gethyllis - из семейства амариллисовых). Иногда, как у веллозии отклоненной (Vellozia patens), нити соединены лишь у самого основания и поэтому кажутся свободными. Хотя специалист по семейству веллозиевых бразильский ботаник Нануза де Менезес (1980) считает, что фаланги представляют собой результат срастания, но более вероятно, что они возникли скорее в результате расщепления. Пыльники прикреплены основаниями или серединой, латрорзные или реже интрорзные, очень редко экстрорзные, раскрывающиеся продольной щелью. Пыльцевые зерна веллозиевых относительно примитивные, однобороздные, у рода веллозия соединены в тетрады. У ксерофиты и барбасении между тычинками и околоцветником имеются своеобразные, похожие на корону амариллисовых придатки, которые обычно срастаются с тычиночными нитями, что делает эти нити как бы широкими и плоскими; у ксерофиты эти коронковидные придатки, приросшие к основаниям сегментов околоцветника; у веллозии придатки, когда они имеются, прикреплены к основаниям тычиночных нитей с вентральной стороны. Происхождение этих придатков не вполне ясно. Одни авторы (Н. де Менезес, 1970, 1980) считают их выростами тканей околоцветника, в то время как другие (Р. Ноер де Алак, 1969) отстаивают их тычиночное происхождение. Гинецей из 3 плодолистиков, 3-гнездный, морфологически паракарпный, с простым столбиком, увенчанным 3-лопастным или цельным (головчатым или удлиненным) рыльцем; завязь обычно нижняя, редко полунижняя. Семязачатки многочисленные на интрузивных плацентах. Плод - деревянистая коробочка, септицидная или раскрывающаяся неправильно. Семена многочисленные, мелкие, с твердой кожурой, маленьким зародышем и обильным эндоспермом.
Большинство веллозиевых произрастают в более или менее сухих местообитаниях, чаще всего на гранитных или кварцитовых скалах, а также на сухих песчаных или каменистых почвах. Часто наблюдается очень узкая приуроченность к химическому составу почвы. Так, некоторые виды веллозии встречаются только на почвах, богатых железом (гематитом). Некоторые веллозиевые, особенно виды африканского рода ксерофита, исключительно засухоустойчивы. Некоторые виды ксерофиты, как ксерофита низкая (X. humilis) и ксерофита Шницлайна (X. schnizleinia), а также бразильская нануза складчатая (Nanuza plicata) в течение нескольких месяцев выдерживают полное обезвоживание листьев. Как только совершенно сухое и, казалось, уже мертвое растение получает влагу, оно очень скоро полностью оживает, зеленеет и продолжает расти. В сухое время года многие веллозиевые могут довольствоваться тем ничтожным количеством влаги, которую доставляет роса. Но среди веллозиевых встречаются и менее ксерофильные, почти мезофильные растения, а талботия является даже на-стоящим мезофитом.
Цветки веллозиевых опыляются как насекомыми (пчелами), так и птицами (колибри). По исследованиям М. Сазимы (1979), у видов барбасении и у одного вида веллозии опыление осуществляется колибри. У остальных изученных видов веллозии опыление производится пчелами.
Семейство понтедериевые (Pontederiaceae) (Т. Д. Сурова)
Представители этого небольшого семейства, насчитывающего 8-9 родов и немногим более 30 видов,- обитатели различных пресноводных водоемов и болот в странах с преимущественно жарким климатом. Южная Америка, в особенности многочисленные реки и озера бассейна реки Амазонки,- центр видового разнообразия понтедериевых. Роды понтедерия (Роntederia), гидротрикс (Hydrothrix) и евристемон (Eurystemon) являются исключительно американскими растениями. Большинство видов родов эйхорния (Eichhornia) и гетерантера (Неteranthera) также жители Американского континента. Только 2-3 вида дикорастущей гетерантеры встречаются в Старом Свете. Род эйхорния представлен здесь одним видом - эйхорнией плавающей (Eichhornia natans), произрастающей в Африке и на острове Мадагаскар. Другой вид этого рода, носящий название водный гиацинт (Е. crassipes), распространенный в тропических и субтропических областях почти всего земного шара (отсутствует только в Европе), является по происхождению американским растением. Вывезенное более 100 лет назад из Южной Америки, оно быстро натурализовалось, заполонило реки и озера многих стран, препятствуя судоходству и засоряя рисовые поля. В странах Старого Света из семейства понтедериевых встречаются также роды монохория (Monochorea) и шоллеропсис (Scholleropsis). Ареал рода монохория, объединяющего 3-4 вида, охватывает тропические и субтропические районы Африки, Австралии, Юго-Восточной и Восточной Азии, заходит на север вплоть до советского Дальнего Востока, где 2 его вида обитают в Приморье и на среднем Амуре. Шоллеропсис - монотипный род, эндемик острова Мадагаскар. Иногда к этому семейству относят еще один близкий к понтедерии род - реусию (Reussia). Однако на основании морфолого-систематических исследований, проведенных американским ботаником P. М. Лoyденом (1973), представляется, по-видимому, более целесообразным включить реусию в состав рода понтедерия в качестве подрода.
Понтедериевые - типичные гидрофиты. Большинство из них полупогруженные растения, растущие на мелководьях у берегов рек, озер, прудов, каналов и канав, по окраинам рисовых полей. Некоторые виды гетерантеры и оба вида бразильского рода гидротрикс - полностью погруженные в воду растения. Их рост, цветение, опыление и плодоношение происходят под водой. Водный гиацинт быстро растет и развивается не только в прикрепленном состоянии, но и свободно плавает по поверхности воды. Многие виды понтедериевых легко переносят недостаток кислорода в почве, образуют обширные заросли на болотах и топях и, в свою очередь, способствуют зарастанию и заболачиванию озер и прудов.
Представители семейства понтедериевых - крупные, высотой до 1 м, или средних размеров многолетние, редко однолетние травы с симподиальным стеблем. У многолетних понтедериевых мясистые, ползучие или иногда укороченные симподиально ветвящиеся корневища. Корневища покрыты остатками влагалищ старых, отмерших листьев. Длинные ползучие корневища с пучками тонких придаточных корней, отходящих от каждого узла, легко обламываются, а их обрывки разносятся ветром, течением и лодками на значительные расстояния. Укореняясь, они дают начало новым растениям. Таким образом осуществляется расселение понтедериевых. Плавучести корневищ способствуют имеющиеся в них многочисленные воздухоносные полости. Такими полостями богаты ткани почти всех органов понтедериевых. Основная их функция - обеспечение нормального газообмена у этих растений. Сосуды с простой или лестничной перфорацией приурочены к корням, редко имеются также в стебле. У большинства видов понтедериевых, как и у многих других водных растений, на одной особи формируются листья двух типов: подводные (погруженные) и надводные (воздушные), имеющие различную форму (гетерофиллия). Погруженные листья обычно более узкие, линейные или узколанцетные. Их черешки укорочены и сближены таким образом, что образуется подобие прикорневой розетки. Воздушные листья супротивные или в мутовках по 3-4 на длинных мясистых черешках. Они либо плавают на воде, либо возвышаются над ее поверхностью. Черешки влагалищные (нижняя часть черешка охватывает стебель), с пленчатым язычком или без него. Необычна форма черешков у водного гиацинта. Они очень толстые, шаровидно вздутые. Вероятно, форма черешков имеет приспособительное значение. Замечено, что почти у всех свободно плавающих особей водного гиацинта черешки имеют характерные вздутия, тогда как последние отсутствуют часто у экземпляров, растущих в грунте. Содержащие большое количество воздуха мясистые черешки водного гиацинта, как поплавки, удерживают растение на поверхности воды. Форма воздушных листьев у понтедериевых чрезвычайно разнообразна и сильно варьирует даже в пределах одного вида. В их очертаниях наблюдаются все переходы от ланцетных до широкояйцевидных или округлых с сердцевидным, стреловидным или почковидным основанием.
Цветки большинства понтедериевых собраны в более или менее крупные пирамидальные соцветия типа кисти, колоса или метелки, которые венчают возвышающиеся над водой цветоносные побеги. У некоторых видов гетерантеры и шоллеропсиса соцветия редуцированы до 2-1 цветка. У основания соцветия имеется покрываловидное влагалище. Оно полностью окружает формирующееся соцветие, защищая бутоны от неблагоприятных воздействий среды. Ко времени расцветания ось соцветия сильно удлиняется и у многих видов значительно выступает за пределы "покрывала". Как показали анатомические исследования, покрываловидное влагалище представляет собой лишенный пластинки видоизмененный черешок - филлодий.
У всех понтедериевых, за исключением нескольких видов гетерантеры и гидротрикса, растущих под водой, образуются открытые (хазмогамные) цветки. Околоцветник обычно состоит из 6 сегментов в 2 кругах (рис. 112). Они почти свободные, как у монохории (рис. 113), или чаще сросшиеся у основания в более или менее длинную трубку. Околоцветник очень яркий, блестящий, фиолетовый или (редко) белый. Тычинок обычно 6, в 2 кругах. Иной облик имеют цветки у рода шоллеропсис. Они желтые, с четырехчленным околоцветником и 4 тычинками, из которых 3 фертильные, а одна превращена в стаминодий. Как правило, тычиночные нити у понтедериевых частично прирастают к трубке околоцветника, пыльники вскрываются интрорзно продольной щелью или (редко) порой. Гинецей синкарпный (у понтедерии - псевдомономерный). Завязь верхняя, с многочисленными анатропными семязачатками или, как у понтедерии, с одним висячим семязачатком. Семена с обильным мучнистым эндоспермом. Образ жизни подводных растений из родов гетерантера и гидротрикс сказался на строении и способе опыления их цветков. У этих видов цветки, как правило, нераскрывающиеся, клейстогамные. Для них характерна в различной степени выраженная редукция андроцея. У гетерантеры развиваются обычно 3 тычинки вместо 6, как у подавляющего числа остальных понтедериевых. У гидротрикса из 3 закладывающихся тычинок формируется только 1 фертильная, а другие (внутренние) превращены в стаминодии.
Рис. 112. Схема опыления трех морфологических форм цветков понтедерии сердцевидной (Pontederia cordata): 1 - законное опыление; 2 - незаконное опыление (а - форма с длинным столбиком, тремя средними и тремя короткими тычинками; б - форма со средним столбиком, тремя длинными и тремя короткими тычинками; в - форма с коротким столбиком, тремя длинными и тремя средними тычинками)
Цветки понтедериевых, формирующиеся в соцветиях, возвышающихся над водой, прекрасно приспособлены к перекрестному опылению. Они слегка зигоморфные. Три верхних сегмента околоцветника образуют подобие паруса, три нижних слегка отогнуты и представляют удобную посадочную площадку для мелких насекомых. На внутренней поверхности центрального верхнего сегмента отчетливо выделяется крупное желтое пятно на синем или фиолетовом фоне. Это метка, указатель нектара для насекомых-опылителей. Они посещают цветки понтедериевых ради нектара, который в большом количестве накапливается на дне трубочки околоцветника. Вначале предполагали, что нектар выделяется базальными частями околоцветника. Однако сейчас точно установлено, что с секрецией нектара связаны септальные нектарники, развивающиеся в нижней части завязи, где стенки плодолистиков срастаются не полностью. Подобные нектарники распространены во многих родах порядка лилейных, бромелиевых и некоторых других однодольных растений. Для большинства понтедериевых характерна триморфная гетеростилия. На различных экземплярах одного и того же вида имеются цветки трех морфологических форм: цветки с длинным столбиком, 3 короткими тычинками и 3 тычинками средней длины; цветки со средним столбиком, 3 короткими и 3 длинными тычинками; цветки с коротким столбиком, 3 длинными тычинками и 3 тычинками средней длины (рис. 112). Рыльце длинностолбчатой формы находится на высоте, точно соответствующей длине самых длинных тычинок в среднестолбчатой и короткостолбчатой формах. Длина столбика среднестолбчатых форм соответствует длине тычинок среднего размера в длинностолбчатых и короткостолбчатых формах. Рыльце короткостолбчатой формы приподнято над завязью на высоту, равную длине самой короткой тычинки в длинностолбчатых и среднестолбчатых формах. Эти три формы различаются и по другим признакам: размерами пыльцы, ее окраской и рисунком экзины, окраской околоцветника, размерами рылец, опушением тычиночных нитей и плодов. Триморфизм такого рода - явление, менее распространенное среди растений, чем диморфная гетеростилия. Аналогичные понтедериевым триморфные формы цветков известны еще у дербенниковых (Lythraceae) и кисличных (Oxalidaceae). Опыление у таких растений будет эффективным только в тех случаях, когда на рыльце каждой формы попадет пыльца от двух других форм, причем с тычинок, длина которых соответствует длине столбика опыляемого рыльца. Наряду с таким легитимным, или законным, способом опыления в популяциях возможно и всегда имеет место, хотя и в очень незначительной степени, иллегитимное, или незаконное, опыление. К последнему относится как самоопыление, так и опыление рылец пыльцой из тычинок несоответствующей длины. При иллегитимном опылении вступает в действие механизм самонесовместимости: нормальное оплодотворение происходит редко, семян производится значительно меньше, чем в случае легитимного опыления. Одним из факторов, препятствующих иллегитимному опылению, является различие в размерах пыльцы, которые находятся в соответствии с длиной тычинок. Некоторые исследователи полагают, что крупные пыльцевые зерна из длинных тычинок развивают более длинную пыльцевую трубку, достаточную для проникновения через длинный столбик. Кроме того, было показано, что более крупные пыльцевые зерна быстрее прорастают, чем мелкие. Это важное обстоятельство для среднестолбчатых и длинностолбчатых форм, так как их столбики вянут раньше, чем медленно растущая трубка мелких пыльцевых зерен успеет достигнуть их завязи. Именно поэтому при искусственном опылении длинно- и среднестолбчатых форм мелкой пыльцой из самых маленьких тычинок семян образуется значительно меньше, чем во всех других случаях незаконного опыления. Цветки понтедериевых опыляются различными маленькими бабочками - парусниками, белянками, нимфалидами. Среди опылителей встречаются пчелы, шмели, осы. Доставая нектар со дна трубочки, насекомое неизбежно касается тычинок разной длины частями своего тельца. Таким образом пыльца из крупных, средних или мелких тычинок оказывается на различных сегментах брюшка насекомого. Перелетая на другой цветок, насекомое соприкасается испачканным пыльцой участком тела с рыльцем соответствующей длины и производит опыление.
Интересный способ приспособления к перекрестному опылению обнаружен у монохории копьевидной (Monochoria hastata) и монохории влагалищной (М. vaginalis, рис. 113). Этим растениям также свойствен полиморфизм (диморфизм) цветков, но он не сопровождается гетеростилией. Цветки этих монохорий голубые, с 6-лопастным околоцветником и 6 тычинками, из которых 5 маленьких стерильных с желтыми пыльниками и одна крупная голубая фертильная тычинка, на тычиночной нити которой имеется один или два выроста типа шпорца. Завязь 3-гнездная, с длинным столбиком и слегка лопастным рыльцем. Диморфизм обусловлен сгибанием фертильной тычинки в правую либо в левую сторону и сгибанием столбика соответственно в противоположном направлении. Сгибание тычинки происходит всегда в сторону где находится шпорец, либо если их два, то по направлению более крупного из них. Эти два типа цветков ("тычинка вправо - столбик влево" и "тычинка влево - столбик вправо") находятся на одном соцветии. Однако наблюдения показали, что они открываются неодновременно. Голубой пыльник вскрывается верхушечной щелью. Верхушка пыльника и рыльце находятся на одном уровне, но с разных сторон цветка. Благодаря такому расположению частей цветка самоопыление маловероятно. Если насекомое, как правило в данном случае пчела, посещает цветок монохории, верхушка пыльника и рыльце касаются разных сторон его тельца. При посещении цветка той же формы опыление не происходит, но оно будет успешным, если насекомое сядет на цветок с противоположным расположением тычинки и столбика. Очень близкий тип диморфизма цветков найден у гетерантеры почковидной (Heteranthera reniformis). Аналогичное строение репродуктивного аппарата известно у некоторых других растений, например у видов рода кассия из семейства бобовых.

Рис. 113. Монохория влагалищная (Monochoria vaginalis): 1 - общий вид растения; 2-3 - две морфологические формы; цветков (а - стерильные тычинки, б - фертильная тычинка, в - столбик); 4 - взаиморасположение фертильной тычинки и столбика в форме 'тычинка влево - столбик вправо' (г - 'шпорец'); 5 - взаиморасположение фертильной тычинки и столбика в форме 'тычинка вправо - столбик влево'; 6 - плод
Продолжительность жизни цветка у понтедериевых невелика. Как правило, он распускается рано утром и к вечеру увядает. Большинство цветков в кисти цветут одновременно, но при неблагоприятных условиях часть цветков может раскрываться на второй и даже на третий день. После отцветания, независимо от того, произошло опыление или нет, цветки закрываются, околоцветник скручивается, цветоножки и ось соцветия сгибаются, погружая завязи в воду. Дальнейшее развитие и созревание семян происходит под водой.
В семействе понтедериевых встречаются плоды двух типов. У представителей рода понтедерия плод односемянный, ореховидный, у остальных - трехстворчатая многосемянная локулицидная коробочка. После созревания плоды всплывают. Их плавучести способствует богатая воздухоносными полостями ткань околоцветника. Ею плоды окружены в период всего своего существования. Заключенные в остатки околоцветника, плоды понтедериевых могут без ущерба для жизни находиться в воде более 15 дней. Этого вполне достаточно для преодоления довольно больших расстояний. Менее обычным способом распространения плодов у понтедериевых является перенос их летающими и наземными животными. Это свойственно некоторым видам рода понтедерии, чьи плоды снабжены крепкими цепляющимися щетинками. Было давно замечено, что в изолированных популяциях видов этого семейства с триморфными цветками часто подавлено развитие семян. Объясняется это тем, что такие популяции возникают нередко в результате вегетативного размножения случайно попавших в какое-либо удаленное место особей одной-единственной флоральной формы.
Вегетативное размножение занимает значительное место в жизненном цикле всех понтедериевых. Оно осуществляется при помощи корневищ и столонообразных побегов. Вегетативное размножение обеспечивает поддержание необходимой для процветания вида численности популяций, особенно тех понтедериевых, у которых по какой-либо причине не развиваются семена. Поразительна скорость, с которой размножаются вегетативным способом представители этого семейства. Например, одна особь водного гиацинта за 50 суток в состоянии образовать до 1000 вегетативных отпрысков. Растущие в грунте понтедериевые образуют таким образом обширные, но локальные заросли. Буйное размножение водного гиацинта в сочетании со свободно плавающим образом жизни, присущим большинству представителей этого вида, ведет к формированию крупных плавучих островков, которые порождают преграды для навигации, рыбной ловли и ирригации.
Родина водного гиацинта - Южная Америка. Немногим более 100 лет назад о существовании этого красивого растения знали только местные жители и специалисты. Водный гиацинт был впервые ввезен в США из Венесуэлы в 1884 г. и демонстрировался как декоративное растение на выставке хлопка. Посетители выставки охотно покупали растение и высаживали его в своих водоемах. Гиацинты быстро размножались и распространялись по рекам и каналам всей округи. Количество их катастрофически росло. Из прелестного декоративного растения водный гиацинт превратился в злостный сорняк, засоряющий водоемы и препятствующий судоходству. Аналогичная история произошла спустя ряд лет, когда растение по той же причине с помощью человека попало в другие страны. Водный гиацинт буквально заполонил многие реки, озера и более мелкие водоемы в странах Африки, Азии, Австралии. Появились сообщения о том, что водный гиацинт натурализовался и на территории нашей страны. В одичавшем виде особи этого растения встречаются местами на Каракумском канале в Туркмении. Плотный ковер из водных гиацинтов иногда полностью останавливает навигацию, нарушает нормальную аэрацию воды, способствует заболачиванию. Это отрицательно сказывается на жизни многих обитателей водоемов, в том числе и ценных пород рыб.
Науке известны примеры, когда неумеренное распространение коварных пришельцев из других семейств высших растений было остановлено насекомыми-паразитами. К сожалению, такого врага у водного гиацинта не нашлось. Одно время предполагали, что неограниченному росту этих растений могут помешать животные. В Африке возлагали большие надежды на гиппопотамов. Однако и гиппопотамы оказались не в состоянии уменьшить численность гиацинтов. Не дали ощутимых результатов механические способы борьбы типа скашивания, выдергивания и т. п. Только использование гербицида 2, 4-Д, распыляемого с самолетов или специальных судов до начала массового цветения гиацинтов, позволило очистить почти полностью от этого сорняка крупные реки и водохранилища. Однако проблема борьбы с водным гиацинтом не решена окончательно. Выяснилось, что хотя данный химический препарат вызывает гибель почти 100% взрослых особей, сам способ борьбы, как это ни парадоксально, способствует созданию оптимальных условий для прорастания семян и не исключает возможности вторичной "инфекции". Дело в том, что к началу распыления гербицида в популяциях всегда имеются особи, успевшие отцвести и завязать плоды. Вместе с отмирающими растениями плоды либо прибиваются к берегу, либо погружаются на дно, где они благополучно заканчивают свое развитие. Попавшие на берег плоды оказываются погруженными в толщу гниющих остатков растений. Образующийся в результате гниения гиацинтов торфоподобный слой может быть толщиной до 2-3 м. Эта жидкая грязь - идеальное место для прорастания семян водного гиацинта. Они накапливаются здесь тысячами и дают многочисленные всходы. Было замечено, что в тех местах, где прибрежный песок не был покрыт разлагающимися остатками растений, молодые всходы водного гиацинта отсутствовали. Таким образом, семена не только концентрируются в местах скопления гниющих остатков водных гиацинтов, но и получают дополнительные условия, улучшающие и ускоряющие их всхожесть - условия, обычно в природе не встречающиеся.
Одновременно с проблемой борьбы с водным гиацинтом встал вопрос о возможности использования его зеленой массы, производимой в большом количестве. Повсеместно зеленую массу применяют как удобрение и на корм скоту. Недавно в Индии разработан способ получения бумаги из этого растения. В северных странах водный гиацинт украшает оранжерейные водоемы. Его выращивают в ботанических садах всего мира. Водный гиацинт - одно из самых любимых растений аквариумистов. В странах Южной и Юго-Восточной Азии водный гиацинт засоряет рисовые поля.
Другим злостным сорняком рисовых полей является монохория влагалищная. В этих условиях растение ведет себя как однолетник: оно гибнет в период высыхания полей, но возобновляется из семян после очередного затопления поля. В постоянно влажных условиях такого чередования не наблюдается и растение может достигать большого возраста. Зеленые части растения монохории влагалищной местное население употребляет в пищу как овощ, а сок листьев и корней находит применение в медицине.
Употребляют в пищу и другой вид монохории - монохорию копьевидную. В культуре в декоративных целях иногда выращивают гетерантеру почковидную (Heteranthera reniformis) и понтедерию сердцевидную (Pontederia cordata).
Семейство филидровые (Philydraceae) (Е. А. Толмачева)
К семейству филидровых относятся всего 4 рода с 5 видами. Они распространены в Восточной и Юго-Восточной Азии, на острове Гуам, Андаманских островах, в Новой Гвинее и Австралии (где встречаются все 4 рода - карта 6).
Карта 6. Ареал семейства филидровых
Филидровые - обитатели постоянно или периодически увлажняемых или даже затопляемых земель. Самый широко распространенный в семействе род - филидрум (Philydrum, рис. 114). Его представители обитают на болотах, в канавах, на намывных песках, но иногда, например в Индокитае, он обычен на рисовых полях и у рыбных прудов. Австралийский род филидрелла (Philydrella) распространен в низменностях у морских побережий, на намывных почвах, сильно увлажняемых зимой и пересыхающих летом. В прибрежных гористых массивах Восточной Австралии на высотах от 600 до 1500 м над уровнем моря обитают ортотилакс гладкий (Orthotilax glaberrimus) и гелъмголъция аиролистная (Helmgoltzia acorifolia). Они занимают берега ручьев в дождевых лесах, образуя весьма своеобразные сообщества. Ортотилакс заселяет преимущественно бурые и каштановые почвы с базальтовой подстилкой. В Новой Гвинее на таких же высотах обитает гелъмголъция новогвинейская (Н. novoguineensis), где она растет на осветленных пространствах в лесах и на морском берегу, на скалах или как эпифит.
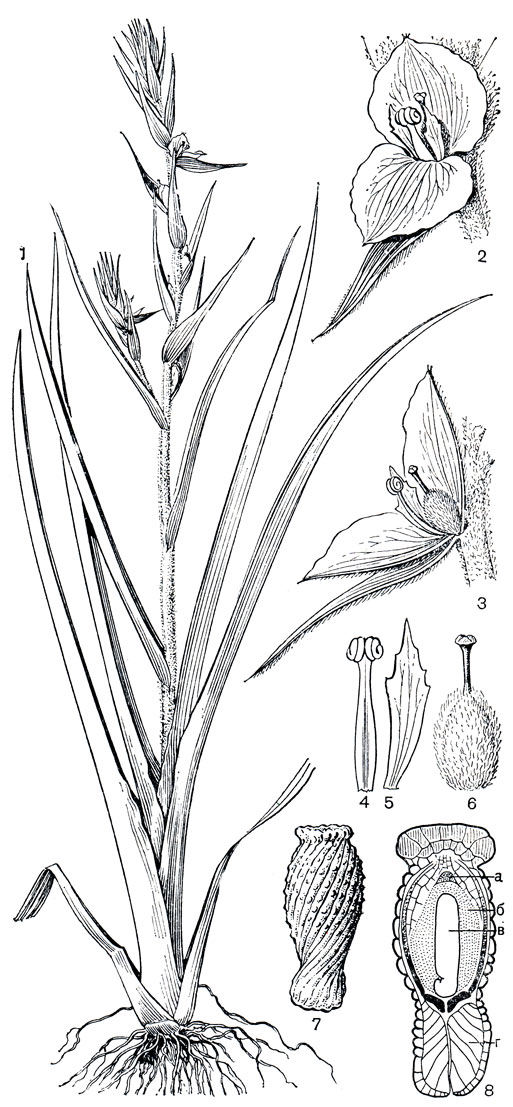
Рис. 114. Филидрум шерстистый (Philydrum lanuginosum): 1 - общий вид растения; 2 - раскрытый цветок; 3 - цветок с удаленным лепестком внутреннего круга околоцветника (вид сбоку); 4 - тычинка со спирально закрученными пыльниками (вид спереди); 5 - лепесток внутреннего круга околоцветника; 6 - гинецей; 7 - общий вид зрелого семени; 8 - продольный разрез семени (а - халазальный и б - микропилярный эндосперм, в - зародыш, г - семенная крышечка)
По внешнему облику филидровые напоминают представителей ирисовых, лилейных и коммелиновых. Это многолетние травы высотой 0,5-1,5 м. Единственный миниатюрный представитель семейства - филидрелла карликовая (Philydrella pygmaea) редко превышает в высоту 15 см, но встречаются экземпляры в половину и даже в одну треть этой величины. Подземным органом у большинства видов является корневище, короткое у филидрума и более мощное и длинное у гельмгольции и ортотилакса. У филидреллы развивается подземный клубень, состоящий из дискообразно утолщенного междоузлия, несущего зачатки листьев, частично укрывающих его. С помощью этого клубня растение переживает летнюю засуху. Корни филидровых короткие, слабоветвящиеся, иногда покрытые густыми, ржавыми волосками. В проводящих пучках корня рядом с трахеидами находятся примитивные сосуды с многочисленными лестничными перфорациями.
Листья филидровых очень сходны а листьями ирисовых. Они в основном прикорневые, с хорошо развитым влагалищем, линейные или мечевидные. Консистенция листьев варьирует от более мягкой у филидрума до кожистой у гельмгольции и ортотилакса, у которых хорошо выражена средняя жилка. У всех видов жилкование параллельное. Устьица расположены с обеих сторон листа. Филидрелла и филидрум имеют устьица с двумя побочными клетками, которые значительно мельче, чем другие клетки эпидермы. У гельмгольции и ортотилакса 4 и более побочных клеток в полярном и латеральном положении, отличающихся только слегка от других клеток эпидермы. В некоторых случаях имеются 4 побочные клетки и у филидрума. Стебель, включая ось соцветия, более или менее опушен у разных видов.
Цветки сидят в пазухах прицветников и собраны в простые или метелковидно ветвистые колосья. Прицветники довольно крупные, овальные и шиловидно заостренные, целиком охватывающие бутон, но отгибающиеся при раскрытии цветка. Цветок сильно зигоморфный. Венчиковидный околоцветник светло-желтого, белого или розоватого цвета, состоящий из 4 сегментов - 2 (внешних) крупных (1-1,5 см), медианных (адаксиального и абаксиального) и 2 (внутренних) очень маленьких (2-5 мм), напоминающих стаминодии и у основания в той или иной степени сросшихся с тычиночной нитью. Верхний (адаксиальный, или передний) сегмент считается морфологически соответствующим адаксиальному лепестку и 2 соседним чашелистикам (иногда на нем наблюдаются 3 зубца), а нижний (абаксиальный) соответствует одному чашелистику. Единственная тычинка по своему положению принадлежит к внешнему кругу типичного 2-кругового андроцея однодольных. Тычиночная нить, приросшая к основанию нижнего лепестка, а также к основанию завязи, образует подобие колонки. Степень срастания лепестков варьирует у отдельных родов. Пыльники интрорзные и открываются 2 продольными щелями. Пыльца с одной широкой дистальной бороздой и сетчатой скульптурой. У филидрума она собрана в тетрады, у остальных родов - одиночная. Гинецей состоит из 3 плодолистиков, синкарпный или реже паракарпный, со столбиком, заканчивающимся головчатым, иногда неявственно 3-лопастным рыльцем. Семязачатки мно-гочисленные, анатропные.
Плод - обычно локулицидная коробочка. У основания плода часто сохраняются остатки околоцветника в виде усохших пленчатых лепестков. Если у филидрума, филидреллы и ортотилакса сухой плод раскрывается 3 щелями по средней линии плодолистиков, то о раскрывании плода гельмгольции сведения противоречивы. Некоторые исследователи называют его нераскрывающимся. К. Скоттсберг (1933) описывает плод гельмгольции новогвинейской как белую ягоду с кожистым тугим перикарпием. Многочисленные мелкие семена, по форме луковицеобразные или веретенообразные, имеют спирально исчерченную или продольно штриховатую семенную кожуру. Характерно наличие семенной крышечки, образованной из внутреннего интегумента. Маленький, слабо дифференцированный зародыш занимает около одной трети от всей массы мясистого эндосперма, состоящего из крупных крахмальных зерен.
Биология опыления филидровых изучена недостаточно. Первые исследования опыления культивируемых растений провел К. Скоттсберг (1932). Он установил, что еще в бутоне до открытия цветка пыльник начинает освобождаться от пыльцы. Тычиночная нить удлиняется, и пыльник снизу прижимается вплотную к рыльцу, где постоянно находятся комочки пыльцы. Позже околоцветник раскрывается. Прорастание пыльцы на собственном рыльце и регулярное оплодотворение цитологически подтверждены У. Хаманном (1962). У цветущего филидрума одновременно открыты 1-2, редко 3 цветка. Каждый цветок открыт только один день, и, как правило, следующий бутон открывается на следующий день. После опыления околоцветник снова закрывается вокруг тычинки и завязи. Прицветник во время распускания цветка отогнут вниз, а затем он опять выпрямляется и охватывает будущий плод, как перед этим бутон. У. Хаманн наблюдал, что время опыления может изменяться. Иногда опыление происходит только при раскрывании или уже после закрытия цветка. Поэтому, считая самоопыление филидрума несомненным, У. Хаманн предлагает не пользоваться в данном случае понятием клейстогамии, о которой говорил К. Скоттсберг, а просто говорить о самоопылении. В природе, однако, не исключена возможность параллельного перекрестного опыления. Филидрум цветет в течение всего года, и в разных частях ареала, несмотря на отсутствие запаха, нектара и богатой пыльцы, а также несмотря на бледную окраску околоцветника, могут оказаться некоторые дополнительные возможности для перекрестного опыления. У филидреллы самоопыление установлено на гербарном и фиксированном материале. Филидрелла произрастает в районах периодической засухи и цветет только в октябре, после окончания дождей, когда почва еще насыщена влагой. Культивируемые растения изредка дают семена, и самоопыление весьма вероятно.
Являясь обитателями хорошо увлажняемых земель и образуя сообщества вблизи водопадов, во влажных оврагах и по краям ручьев, филидровые обладают легкими плавучими семенами, которые прорастают через неделю после набухания. По-видимому, вода - основной фактор для распространения их семян. Прорастание семян наземное.
Внутри семейства выявляется довольно тесное родство между отдельными родами. Т. Каруэль (1881) объединял в один род ортотилакс и гельмгольцию, а современные австралийские ботаники О. Эванс и Л. Джонсон (1966) считают выделение ортотилакса в особый род недостаточно обоснованным. Эти роды действительно имеют между собой много общего по внешнему облику, форме листьев и соцветий. Гельмгольция и ортотилакс более примитивны чем филидрум и филидрелла, хотя их уровень специализации во многом не одинаков. Каждый род имеет особенности, свойственные только ему: у филидрума - пыльца в тетрадах и спирально закрученные пыльники; у филидреллы - подземный клубень, почковидно выгнутый пыльник и упрощенное строение семенной кожуры; у ортотилакса - удлиненные веретенообразные семена; у гельмгольции - закрытые плоды. Близость между ортотилаксом и гельмгольцией особенно большая: прямые пыльники, форма внешнего круга околоцветника, ветвящееся соцветие, строение устьичного аппарата, характер корневища.
В экономическом отношении филидровые не представляют сколько-нибудь заметного интереса. Однако, как изящные и легко возделываемые растения, их выращивают на улицах Сиднея и во многих ботанических садах Европы. В теплицах филидрум цветет уже в первый год после посева. Его выращивают как однолетник и пересевают семенами. При подходящих условиях культивирования достигается богатое семенное возобновление. В Берлинском ботаническом саду отдельные экземпляры филидрума достигают двухметровой высоты и могут перезимовывать корневищами.
Узнать цены и подробную информацию о доставке грузов самолетом в Санкт-Петербург.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'