Порядок смилаксовые (Smilacales)
Семейство филезиевые (Philesiaceae) (М. В. Баранова)
В семействе филезиевых 8 родов и 12-13 видов. Они распространены почти исключительно в южном полушарии. Только у одного монотипного рода гейтоноплезиум (Geitonoplesinm) ареал заходит в северное полушарие, на Филиппинские острова (остров Лусон). Представители семейства встречаются в Южной Америке - на Западе Аргентины и в Чили, к югу от 35° ю. ш. включая Фолклендские острова (филезия - Philesia, рис. 115, лапажерия - Lapageria, лузуриага - Luzuriaga), в Австралии на северо-западе (элахантера - Elachanthera) и на востоке (петермапния - Petermannia), на островах Океании (эвстрефус - Eustrephns, рис. 116, гейтоноплезиум) и в Африке, где на юго-востоке растет единственный африканский представитель семейства - бения сетчатая (Behnia reticulata). Такой разрыв ареала семейства указывает на его большую древность. Наибольшее число видов приурочено к тропической и субтропической зонам. В умеренной зоне встречается лишь южноамериканский и новозеландский род лузуриага (карта 7).
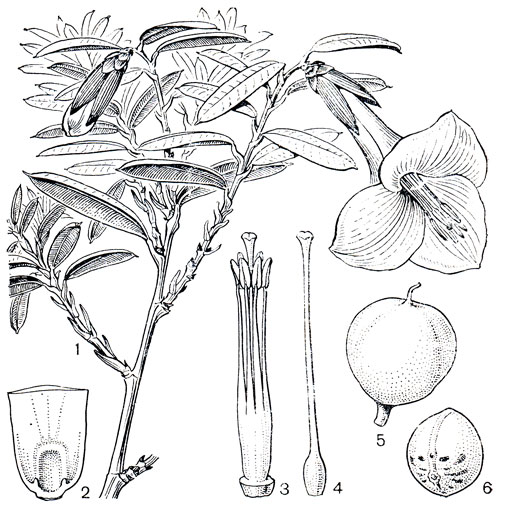
Рис. 115. Филезия самшитолистная (Phiiesia buxifolia): 1 - общий вид растения с цветком; 2 - основание лепестка с нектарником; 3 - андроцей; 4 - гинецей; 5 - плод; 6 - семя
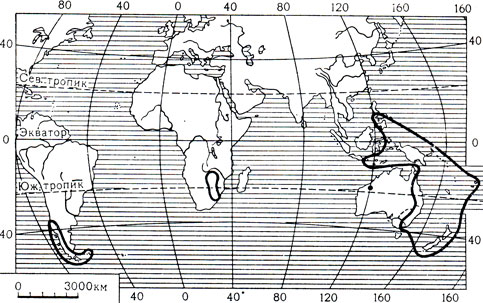
Карта 7. Ареал семейства филезиевых
Филезиевые - невысокие прямостоячие или стелющиеся полукустарники, лазающие кустарники или лианы с вечнозелеными кожистыми цельными листьями на коротких, иногда членистых (филезия) черешках или почти сидячие. Листья с 1-8 продольными, часто выпуклыми жилками и нередко развитыми поперечными, от чего жилкование имеет вид сетчатого (бения, лузуриага). Они варьируют от мелких линейных, длиной не более 1,5 см, до крупных широколанцетных и яйцевидных, длиной до 10 см (лапажерия). Любопытной особенностью некоторых представителей семейства (гейтоноплезиум цимозный - Geitonoplesium cymosum и петермапния усиковидная - Petermannia cirrhosa) является повернутая на 180° листовая пластинка. В результате нижняя эпидерма листа с устьицами и прилегающая к ней губчатая паренхима располагаются сверху, а верхняя эпидерма с палисадной тканью - снизу. Для защиты устьиц, оказавшихся на верхней поверхности листа, соседние с ними клетки эпидермы увеличиваются в объеме и прикрывают устьица сверху. Губчатая ткань таких листьев приобретает некоторое сходство с палисадной тканью. Ксилема филезиевых представлена трахеидами с лестничными перфорациями, промежуточными между трахеидами и сосудами образованиями и сосудами с лестничной или простой перфорацией.
Цветки пониклые, колокольчатые или звездчатые, актиноморфные, обычно на членистых цветоножках, собраны в верхушечные или пазушные соцветия, или цветки пазушные одиночные (филезия, лапажерия, лузуриага). Лишь у петерманнии соцветия внепазушные, супротивные листьям. Околоцветник филезиевых 6-членный, его сегменты свободные, равные или три наружных значительно уступают по размерам внутренним (филезия). Сросшийся околоцветник имеет только бения сетчатая. Сегменты околоцветника разнообразны по величине, форме, консистенции, окраске, от мелких, длиной не более 5 мм, как у эвстрефуса широколистного (Eustrephus latifolius, рис. 116), до крупных, длиной до 8 см, как у лапажерии розовой, от тонких ланцетных до сочных лопатчатых, яркие карминно-красные (лапажерия, филезия), розовые, белые (эвстрефус, лузуриага), зеленовато-желтые (бения). Тычинки прикреплены к основанию сегментов околоцветника или к его трубке (бения). Нити тычиной: свободные или сросшиеся по всей длине в коротную трубку (эвстрефус) или только в основании (филезия). Пыльники интрорзные или, редко, экстрорзные (петерманния), вскрываются одной или двумя (бения) продольными щелями или порами (лузуриага, эвстрефус). Пыльцевые зерна однобороздные, с простой, трех- или четырехлучевой бороздой, редко безапертурные (лапажерия, филезия); оболочка пыльцевых зерен мелкобугорчатая или шиповатая; пыльцевые зерна филезии и лапажерии покрыты крупными шипами. Гинецей филезиевых образован тремя сросшимися плодолистиками, синкарпный или паракарпный (лапажерия, филезия, петерманния); завязь верхняя, за исключением петерманнии, у которой она нижняя; у бении завязь в основании сжата и образует короткий гинофор (Э. Филлипс, 1951). Семязачатков в гнездах завязи несколько или много, кампилотропных. Цельное или трехлопастное, с чуть удлиненными лопастями рыльце едва выдается над тычинками.
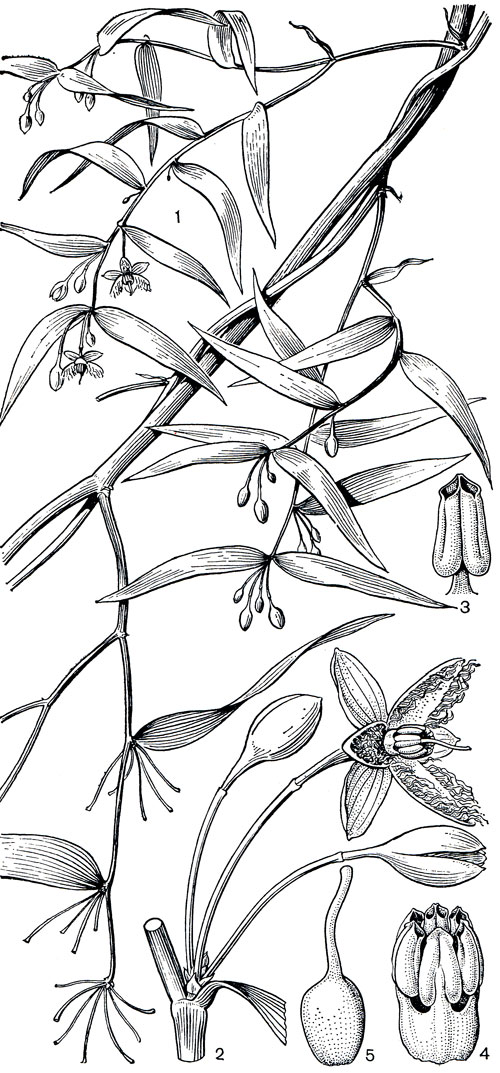
Рис. 116. Эвстрефус широколистный (Eustrephus latifolius): 1 - общий вид растения с цветками; 2 - цветок; 3 - зрелая тычинка со вскрывшимися пыльниками; 4 - андроцей; 5 - гинецей
Опыляют яркие карминно-красные цветки лапажерии и филезий колибри (К. Грант и В. Грант, 1968). Они собирают нектар, который выделяется и скапливается в ямчатых нектарниках в основании сегментов околоцветника, одновременно перенося пыльцу с цветка на цветок. Цветки лузуриаги, видимо, самоопыляются (Д. Мур, 1968). Бледноокрашенные мелкие цветки других филезиевых опыляют, очевидно, мелкие насекомые.
Плоды филезиевых - ягоды, от мелких и округлых, диаметром до 8 мм, до крупных продолговато-яйцевидных, длиной до 5 см (лапажерия). Интересно отметить, что у некоторых филезиевых (эвстрефус) после длительного и полного высыхания сочного перикарпия ягоды вскрываются по трем гнездам подобно локулицидной коробочке. Ярко окрашенные и сочные плоды привлекают птиц и мелких животных, которые, поедая плоды, распространяют семена. С яркой окраской плодов связаны местные названия растений. Так, "кораллами гор" называют в Чили красные ягоды лузуриаги укореняющейся (Luzuriaga radicans), оранжевые плоды Эвстрефуса широколистного известны в Австралии как "ягоды вомбата", а зеленоватые сочные ароматные и съедобные плоды лапажерии розовой в Чили продают под названием "чилийских огурцов". Семена округлые или слегка неправильной формы, с мелкоячеистой семенной кожурой, иногда с крупным белым ариллоидом (эвстрефус). Эндосперм обильный, а зародыш мелкий, прямой или слегка изогнутый.
Лиановидные филезиевые (эвстрефус, гейтоноплезиум, лапажерия, бения) - обитатели влажных тропических и субтропических лесов и кустарниковых зарослей, где они оплетают своими сизовато-зелеными, похожими на проволоку, сильно ветвистыми стеблями деревья и кустарники. Семена этих растений, разносимые птицами, иногда попадают на пни и стволы деревьев, прорастают там и растут на них как эпифиты, пока их корни не достигнут уровня земли. Филезиевые встречаются в светлых сухих лесах, на открытых склонах холмов и обнаженных скалах. Там они представлены невысокими прямостоячими или стелющимися полукустарниками. У многих из них в основаниях нижних ветвей на стеблях формируются воздушные стеблевые корни (филезия, лузуриага). Их развитие у стелющихся кустарников способствует широкому разрастанию растений и захвату ими большей площади питания. Способностью формировать стеблевые корни обладает и лиана лапажерия розовая. Однако формируются они только при полегании стеблей. На этом основано вегетативное размножение лапажерии - наиболее декоративного представителя семейства.
В настоящее время семейство филезиевых подразделяется на 3 подсемейства - лузуриаговые (Luzuriagoideae), филезиевые (Philesioideае с 2 родами - филезия и лапажерия) и петерманниевые (Petermannioideae с родом петерманния).
Наиболее известным растением в семействе филезиевых является лапажерия. Она описана в 1802 г. и названа в честь жены Наполеона I - Жозефины ла Пажери. Лапажерия розовая - единственный представитель рода, эндемик Чили - встречается в густых лесах на западных склонах Анд в центральных районах Чили (от 35 до 40° ю. ш.) и впервые была собрана в окрестностях города Консепсьон. Эта лиана с блестящими кожистыми листьями и ярко- малиновыми крупными цветками поразила своей красотой путешественников, впервые увидевших ее в Чили (табл. 11). Лапажерия разовая - национальный цветок Чили.
В 1854 г. лапажерия была привезена в Европу английским садоводом У. Лоббом. С 1866 г. лапажерию розовую и ее форму с белыми цветками (var. albiflora) выращивали в оранжереях Петербургского (Ленинградского) ботанического сада вплоть до зимы 1941 г., когда оранжереи сада были разрушены. В Англии в 1872 г., в садоводстве Вича, был получен межродовый гибрид филезии самшитолистной (Philesia buxifolia, рис. 115) с лапажерией розовой, названный филажерией Вича (Philageria veitchii). Филажерия - вечнозеленый кустарник с длинными слегка вьющимися ветвями и розовыми цветками. Лапажерия и филажерия признаны лучшими вьющимися декоративными растениями для оранжерей и зимних садов. Мясистые веретеновидные корни лапажерии используют в народной медицине при лечении болезней, связанных с нарушением обмена веществ.
В роде филезия всего 3 вида, которые растут в горах, на лесных болотах в южных районах Чили. Это невысокие прямостоячие кустарники, ветви которых густо покрыты мелкими кожистыми, внизу белыми листьями с четко выраженной центральной жилкой (рис. 115). Листья филезий, особенно филезии самшитолистной, очень сходны с листьями подбела многолистного (Andromeda polifolia), произрастающего на болотах в северном полушарии. Ярко-розовые, крупные, длиной до 5 см, цветки филезии очень красивы, и с 1847 г. ее выращивают в садах Англии как декоративное растение.
Род лузуриага объединяет 3 вида, два из которых встречаются в Южной Америке - Перу, Чили, Аргентине - и на Фолклендских островах, а один - лузуриага мелкоцветковая (Luzuriaga parviflora) - в Новой Зеландии, включая остров Стьюарт. Ветви этих стелющихся или прямых кустарников покрыты многочисленными мелкими листьями с сетчатым жилкованием. Клетки нижней эпидермы листьев лузуриаг четко квадратные или прямоугольные с сильно утолщенными радиальными стенками, отчего листья снизу кажутся белыми. Прочные и гибкие стебли лузуриаги укореняющейся местные жители используют для плетения корзин.
В кустарниковых зарослях Зимбабве, Свазиленда, Трансвааля и Наталя в Юго-Восточной Африке растет лиана бения сетчатая - единственный представитель рода. Растение имеет очень красивые с изящным сетчатым жилкованием кожистые листья.
Во влажных лесах Восточной Австралии и на островах Новая Гвинея и Новая Каледония часто встречается лазающий, сильно ветвящийся кустарник с подземным корневищем и утолщенными мясистыми корнями - эвстрефус широколистный - единственный вид рода. Его мелкие белые цветки в короткокистевидных соцветиях обильно покрывают ветви. Нитевидные выросты длиной до 1,5 см эпидермальной ткани внутренних сегментов околоцветника покрывают их края (рис. 116).
К роду эвстрефус наиболее близок монотипный род гейтоноплезиум. Ареал его единственного вида - гейтоноплезиума цимозного - очень широк и охватывает восточные районы Австралии, многие острова Океании - Новая Гвинея, Новая Каледония, Фиджи и Новые Гебриды, Соломоновы и Молуккские, Сумбава - и заходит в северное полушарие на Филиппинские острова (остров Лусон).
На северо-западе Австралии встречается малоизвестный монотипный род элахантера, а на юге-востоке - род петерманния с единственным видом петерманния усиковидная. Петерманния - вьющееся растение и обладает рядом признаков, таких, как супротивное по отношению к листу расположение соцветия, нижняя завязь, экстрорзные пыльники, которые послужили причиной для выделения этого рода некоторыми авторами (Дж. Хатчинсон, 1934, 1959) в самостоятельное семейство петерманниевые (Petermanniaceae).
Семейство стемоновые (Stemonaceae) (Т. В. Вельгорская)
Семейство Симоновых состоит из 3 родов и 30 видов, обитающих главным образом в тропиках и теплых областях Восточной, Южной, Юго-Восточной Азии и Северо-Восточной Австралии; один представитель этого семейства - крумия немногоцветковая (Croomia pauciflora, рис. 119) - произрастает на юго-востоке США.
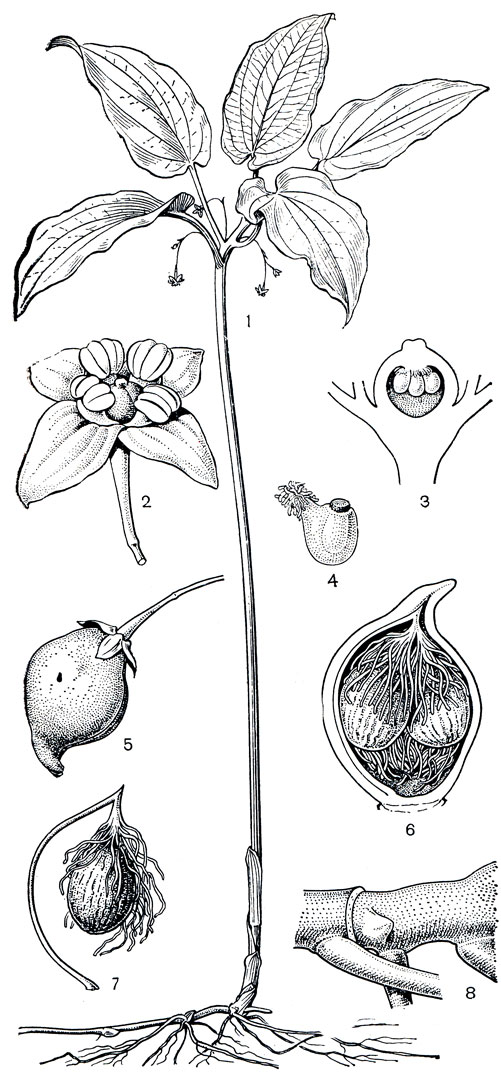
Рис. 119. Крумия немногоцветковая (Croomia pauciflora): 1 - общий вид; 2 - цветок; 3 - гинецей (видны висячие семязачатки); 4 - семязачаток; 5 - плод; 6 - продольный разрез плода; 7 - семя; 8 -часть корневища
Стемоновые - многолетние корневищные травы с прямостоячими стеблями или травянистые лианы с клубневидными корнями (виды стемоны - Stemona, см. рис. 118). Сосуды с лестничной перфорацией, обычно только в корнях, но иногда (у стемоны) они имеются и в стеблях. Листья очередные, супротивные или мутовчатые; широкая листовая пластинка с 5-11 (иногда до 15) дуговидно сходящимися главными и многочисленными поперечными, вторичными жилками (рис. 118, 7). Устьица аномоцитные. Цветки в малоцветковых пазушных соцветиях, обычно обоеполые, очень редко однополые (часть цветков у стемоны Куртиса - S. curtisii), актиноморфныеу 2-членные. Околоцветник венчиковидный, состоящий из 4 сегментов, расположенных в 2 кругах; сегменты свободные или иногда сросшиеся у ос-нования. Тычинок 4, в 2 круга или редко в 1 круг; тычиночные нити свободные или почти свободные; пыльники интрорзные, раскрывающиеся продольной щелью; у видов стемоны пыльники с большим надсвязником (рис. 118,2). Пыльцевые зерна однобороздные. Гинецей из 2 плодолистиков; завязь верхняя или редко полунижняя (у рода стихоневрон - Stichoneuron), 1-гнездная; столбик короткий или чаще рыльце сидячее (рис. 117); семязачатков 2 или много, они базальные и ортотропные, или апикальные и анатропные, или гемитропные (у крумии), битегмальные, крассинуцеллятные. Плод, за исключением стихоневрона,- коробочка, раскрывающаяся 2 створками. Семена удлиненные, ребристые, с маленьким зародышем и обильным эндоспермом.
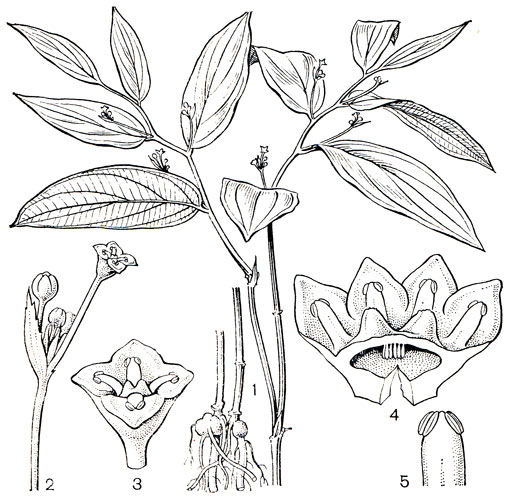
Рис. 117. Стихоневрон перепончатый (Stichoneuron membranaceum): 1 - общий вид; 2 - соцветие; 3 - цветок; 4 - продольный разрез цветка, видны висячие семязачатки; 5 - тычинка
Произрастают стемоновые в субтропических или тропических горных лесах, на горных склонах или в долинах, предпочитая тенистые места, заросли кустарников. Обыкновенно они растут на сухих песчаных или глинистых почвах, но иногда стемона клубневая (Stemona tuberosa, рис. 118,1-7) встречается и на сырых илистых почвах. Стихоневрон и некоторые виды стемоны встречаются на высоте 1100-1800 м над уровнем моря. В Японии виды крумии обычны на плантациях криптомерии японской (Cryptomeria japonica), а крумия немногоцветковая селится по отвесным лесистым берегам рек, выбирая места в густой тени обрывов, на рыхлых осушенных почвах.
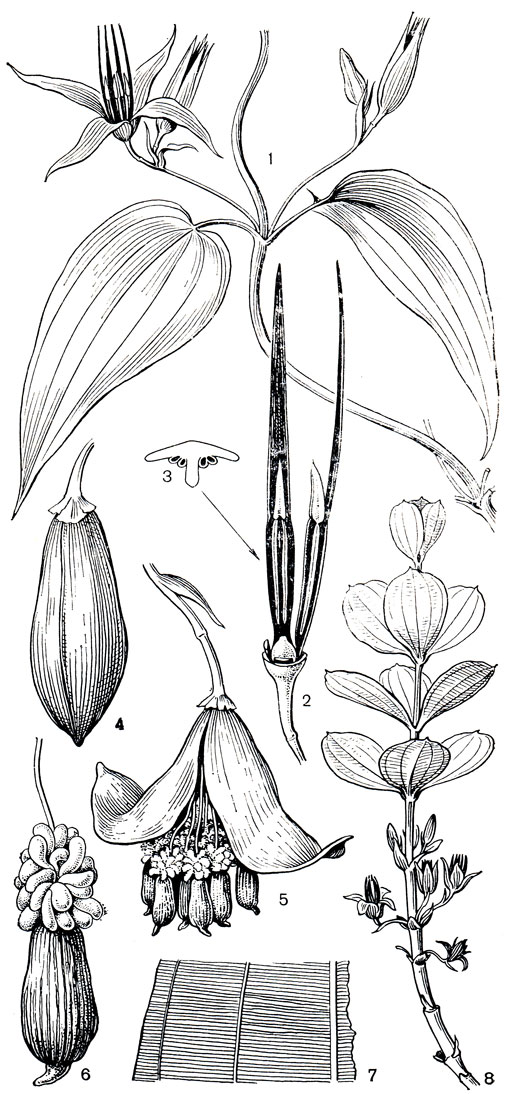
Рис. 118. Стемоновые. Стемона клубневая (Stemona tuberosa): 1 - часть растения с цветками; 2 - тычинки с надсвязниками; 3 - поперечный разрез тычинки; 4 - плод; 5 - раскрытая коробочка с семенами; 6 - семя; 7 - часть листовой пластинки. Стемона сидячелистная (S. sessilifolia): 8 - надземная часть растения
Цветут стемоновые в марте - июле. О биологии семейства сведений мало, но, скорее всего, стемоновым свойственна энтомофилия. В пользу этого предположения говорят строение цветка (большие надсвязники у видов стемоны, множество сосочков на внутренних стенках околоцветника у крумии), окраска (например, у стемоны клубневой пурпурно-красный, иногда голубой околоцветник и тычинки с ярко- желтыми надсвязниками и фиолетовыми пыльниками; оранжевые пыльники у крумии разночашелистиковой - Croomia heterosepala) и запах, чаще всего неприятный для человека, а у стемоны клубневой исключительно тяжелый, зловонный. По мнению П. Кунта (1904), цветкам стемоны яванской (Stemona javanica), возможно, свойственно самоопыление. Цветок этой стемоны устроен так, что при распускании околоцветник остается полуоткрытым; длинные лопасти надсвязников плотно смыкаются друг с другом и образуют подобие купола над гинецеем, тем самым предохраняя рыльце от попадания пыльцы с другого цветка.
Семена стемоновых снабжены ариллусами или ариллусовидными придатками - элайосомами, делающими их привлекательными для птиц, муравьев и других животных. Кроме того, возможно, что семена некоторых видов стемоны распространяются анемохорно; в раскрывшихся коробочках семена свисают на длинных нитевидных семяножках и, оторванные ветром, могут отлететь на какое-то расстояние от материнского растения. У крумии немногоцветковой, растущей по обрывистым берегам рек, семена с плотной кожурой могут распространяться и водным путем, но естественное возобновление этого вида крумии невысокое, что ведет к сокращению численности популяций растения, и это вызывает озабоченность американских ученых (П. Б. Томлинсон и Э. С. Айенсу, 1968).
Семейство стемоновые подразделяется на две трибы: крумиевые (Croomieae) и собственно стемоновые (Stemoneae).
Триба крумиевых характеризуется прямостоячими жесткими стеблями, пыльниками без надсвязников и висячими семязачатками. Известный японский ботаник Т. Накаи (1943) выделял эту трибу в самостоятельное семейство крумиевые (Croomiaceae), которое признается и некоторыми другими ботаниками. Крумиевые состоят из 2 родов - крумии (Croomia), названной в честь одного из первых американских ботаников Г. Б. Крума, и стихоневрона (Stichoneuron).
Род крумия (всего 3 вида) имеет дизъюнктивный ареал: 2 его вида - крумия японская (С. japonica) и крумия разночашелистиковая - растут в Японии на островах Хонсю, Сикоку и Кюсю, а третий вид - крумия немногоцветковая - обитает в юго-восточной части США, в штатах Флорида, Джорджия и Алабама. Такая разорванность ареала позволила знаменитому американскому ботанику прошлого века Эйсе Грею (1846) впервые высказать предположение о древнем флористическом родстве Северной Америки и Восточной Азии. Виды крумии - многолетние травы высотой 30-60 см, с ползучими, широко простирающимися корневищами, которые, однако, находятся в почве на небольшой глубине, что делает особи крумии уязвимыми - растения легко вырываются. Листья длиной 6-15 см и шириной 3-8 см расположены в верхней части гладкого, прямого стебля и имеют овальную или продолговато-овальную форму. В соцветиях обычно 1-4 небольших висячих цветка на слегка утолщенной наверху цветоножке длиной 5-8 мм, с маленьким прицветником. Сегменты околоцветника желтовато-зеленые или зеленые; у крумии разночашелистиковой около-цветник слегка зигоморфный, так как 1 из 4 сегментов несколько больше остальных. У крумии японской края околоцветника немного отогнуты. Внутри сегменты покрыты редкими тупыми сосочками, а у крумии японской и крумии немногоцветковой такими сосочками покрыты и тычиночные нити. Эти сосочки состоят из крупных клеток, цитоплазма которых содержит антоциан. Расположенные супротивно сегментам околоцветника 4 тычинки образуют 1 круг. Нектарников у крумии нет. При плодах сохраняется околоцветник. Семена с выступающими продольными ребрами, широко-овальные, длиной 4 мм, с короткой семяножкой, снабженной массивным пучком мясистого щетинистовидного образования.
Второй род трибы крумиевых - стихоневрон (только 2 вида) - произрастает в Восточной Индии (Ассам и Манипур), в Бангладеш, Таиланде, на полуострове Малакка (стихоневрон хвостатый - Stichoneuron candatum) и в Индонезии. Виды стихоневрона - прямостоячие, жесткие, гладкие или слегка опушенные травы с коротким корневищем и умеренно ветвистым стеблем. Листья супротивные, на коротких черешках, линейно-продолговатые. Соцветие пазушное, как бы лежащее вдоль главной жилки листа и несущее кисть очень мелких зеленовато-желтых цветков на маленьких цветоножках и с щетинковидными прицветниками. Околоцветник колокольчатый, с яйцевидными сегментами; тычинок 4, приросших к основаниям сегментов околоцветника, с короткими, толстыми, слегка загнутыми наружу нитями; рыльце 3-лопастное, сидячее (рис. 117). Плод сухой, односемянный. В литературе о плодах стихоневрона чаще всего сообщается, что "их никто не видел" (например, известный английский ботаник Г. Ридли в работе 1937 г. указывает, что на полуострове Малакка, как и в Гималаях, встречаются только стерильные особи стихоневрона). Но в гербарии Ботанического института им. В. JI. Комарова АН СССР в Ленинграде хранится уникальный гербарный материал, собранный еще в 1856 г. в Ассаме самим Дж. Д. Хукером и Т. Томсоном, в котором имеется половинка плода стихоневрона перепончатого (S. membranaceum, рис. 117).
Монотипная триба стемоновых состоит из рода стемона, в котором насчитывается 25 видов. Растут виды стемоны в Японии (2 вида: стемона японская - Stemona japonica и стемона сидячелистная - S. sessilifolia), в Центральном, Южном и Юго-Западном Китае (кроме широко распространенных стемоны клубневой, стемоны сидячелистной и других, здесь произрастают 2 эндемичных вида: стемона прямостоячая - S. erecta и стемона раскидистая - S. vagula), в Восточной Индии (Орисса, Ассам, Манипур), Бангладеш, в Таиланде, Бирме, на острове Шри-Ланка, в Индонезии (стемона яванская - S. javanica), на Филиппинских островах и на острове Новая Гвинея. Наконец, встречается стемона в Северо-Восточной Австралии (стемона австралийская - S. anstraliana). Виды стемоны - длинные вьющиеся или невысокие прямостоячие (высотой около 30 см у стемоны сидячелистной) травы или лианы с короткими корневищами и мясистыми клубневидными корнями, с облиственным стеблем. Листья супротивные или чаще мутовчатые, на черешках или сидячие, яйцевидные, округлые, с 3 - многими главными и многочисленными поперечными жилками. Цветки одиночные или собраны в малоцветковые пазушные соцветия, обоеполые, очень редко однополые. Сегменты околоцветника ланцетные, свободные, бледно-зеленые, с многочисленными пурпурно-красными жилками. Тычинки очень длинные (у стемоны клубневой длиной до 35 мм), с ланцетными или линейными надсвязниками и сросшимися у основания красными или пурпурно-фиолетовыми, иногда голубыми нитями; пыльники остроконечные, окрашенные в желтые или фиолетовые тона, рыльце сидячее. Семена продолговатые, с нитевидной, на конце волосистой семяножкой и снабжены беловатым или бесцветным ариллусом или ариллусовидным придатком.
Практическое значение стемоновых сравнительно невелико. Издавна китайская народная медицина, берущая свое начало в глубокой древности, использует клубневидные корни некоторых видов стемоны (главным образом стемоны клубневой) для лечения ряда заболеваний. Китайские кулинары также не обошли вниманием эти растения. Клубни стемоны клубневой, сваренные или тушенные со свининой,- довольно популярное блюдо. В корнях стемоны содержатся специфические алкалоиды - стемонин, стемонидин и изостемонидин. В Японии отвар из корней стемоны сидячелистной и стемоны японской используют как инсектицидное средство против кожных паразитов у крупного рогатого скота.
Семейство триллиевые (Trilliaceae) (А. Л. Тахтаджян)
В это семейство входят 4 рода и около 60 видов, распространенных в гумидных областях северного полушария. Монотипный японский род кинугаса (Kinugasa) растет на острове Хонсю. Небольшой евразиатский род парис (Paris), насчитывающий всего 5-6 видов, распространен в Средиземноморье, Европе, на Кавказе, в Сибири и Восточной Азии (включая Дальний Восток СССР). Род даисва (Daiswa), который современными авторами без достаточных оснований включается в род парис, насчитывает около 15 видов. Виды даисвы распространены в умеренной зоне Гималаев (на западе до Пенджаба и Кашмира), в Северо-Восточной Индии, Северной Бирме, континентальном Китае, на островах Хайнань и Тайвань, в Северном Индокитае. Четвертый и самый большой род триллиевых - это род триллиум (Trillium), насчитывающий около 40 видов, из которых 5 видов встречаются в Азии от Западных Гималаев до Японии и Камчатки, а остальные виды приурочены к Северной Америке.
Виды триллиевых - типичные мезофиты, предпочитающие влажные почвы. Они растут обычно в тенистых местах, в хвойных и лиственных лесах, на влажных местах среди кустарников и на горных лугах.
Триллиевые - многолетние травы с длинным или коротким (иногда почти клубневидным), горизонтальным или вертикальным моноподиальным корневищем, от верхушки которого ежегодно отходит один, иногда два, редко несколько однолетних побегов. Стебель всегда простой, прямостоячий, короткий или высокий (у некоторых видов даисвы высотой до 2 м), с несколькими короткими листовыми влагалищами у основания. Редко растение фактически бесстебельное, если не считать очень короткого подземного стебля, лишь иногда поднимающегося на несколько сантиметров над землей, как у триллиума черешчатого (Т. реtiolatum). Хотя стебель триллиевых в момент цветения всегда прямостоячий, у триллиума зеленолепестного (Т. chloropetalum) он перед созреванием плодов полегает и продолжает свое развитие в горизонтальном положении, что объясняется как дополнительное приспособление к мирмекохории. Листья триллиевых в мутовках по 3 (триллиум) или по 4-22 (остальные роды) в верхней части стебля, сидячие или на черешках, от линейных до яйцевидных, эллиптических, округлых или ромбических, у некоторых видов даисвы (например, у даисвы Фарже - Daiswa fargesii) у основания более или менее сердцевидные; жилкование листьев характеризуется большей или меньшей сетчатостью. Устьица аномоцитные. Сосуды только в корнях, с лестничной перфорацией. Цветки триллиевых всегда одиночные на верхушке стебля, обычно довольно крупные, большей частью на прямой или редко согнутой цветоножке (достигающей у некоторых видов даисвы длины более 1 м), обоеполые, актиноморфные, трехчленные или реже четырехчленные, с двойным околоцветником или иногда безлепестные. Сегменты околоцветника свободные. Чашелистики обычно ланцетные, как правило, зеленые, редко белые (кинугаса), сохраняющиеся и продолжающие увеличиваться в размерах до созревания плода. Лепестки от нитевидных до овальных или яйцевидных, опадающие или усыхающие после цветения, белые, желтые, желтовато-зеленые, зеленые, розовые, пурпурные, коричнево-пурпурные и буровато-красные. Тычинок от 6 до 22, чаще всего 6- 12 (у триллиума всегда 6), прикрепленные к основанию сегментов околоцветника; нити обычно короткие, тонкие, плоские; пыльники прикреплены к нити основанием, продолговатые или нитевидные, у некоторых видов париса и даисвы с хорошо развитым надсвязником (который у даисвы тибетской - D. thibetica достигает в длину 12-16 мм), раскрывающиеся продольной щелью. Пыльцевые зерна однобороздные. Гинецей паракарпный или реже синкарпный (скорее всего вторичносинкарпный), из 3-10 плодолистиков (у триллиума всего 3 плодолистика); завязь верхняя, 1-гнездная или 3-4-гнездная, с многочисленными анатропными семязачатками; столбики свободные или чаще более или менее сросшиеся. Плоды более или менее мясистые, ягоды (у триллиума к созреванию иногда высыхающие) или коробочковидные, раскрывающиеся локулицидно (даисва). Семена с эндоспермом и с очень маленьким недоразвитым зародышем, эллипсоидальные и слегка согнутые или более или менее шаровидные, у рода триллиум снабженные белым придатком, представляющим собой строфиолу (утолщение верхней части шва рядом с короткой семяножкой), у остальных родов придатки отсутствуют, но у даисвы семя окружено сочной саркотестой; только роды парис и кинугаса лишены как придатков, так и саркотесты и семя окружено сухой сетчатой оболочкой. Интересно отметить, что у представителей триллиевых самые крупные хромосомы среди всех изученных в этом отношении цветковых растений.
После этой общей характеристики семейства познакомимся с каждым из четырех родов в отдельности. Начнем с наиболее знакомого читателю рода парис, русское народное название которого "вороний глаз" основано на черных ягодах этого растения. Для видов этого рода характерны тонкие, длинные, горизонтальные корневища, ежегодно нарастающие своей верхушкой; невысокий стебель с 4-12 листьями и относительно небольшими цветками с 4 или реже 5 чашелистиками, 8-10, иногда 6 тычинками и 4- или реже 5-гнездной (изредка 6-гнездной) завязью с таким же числом свободных тонких столбиков с верхушечными рыльцами. Плоды представляют собой довольно типичные ягоды сизовато-черного цвета, ядовитые. Очень интересен цикл развития париса, в частности одного из наиболее распространенных его видов - париса обыкновенного (Paris quadrifolia). Это единственный вид этого рода, встречающийся в Средиземноморье, Европе и Западной Сибири и во многих отношениях самый изученный его представитель, которому посвящено довольно много исследований. Цикл развития париса обыкновенного особенно детально изучен С. С. Трофимовым (1956), который следующим образом описывает его. Весной, уже вскоре после таяния снега, из-под лесной подстилки появляется молодой побег париса со свернутыми в трубочку листьями, защищающими цветок. Уже через 10-15 дней листья на побеге расправляются и почти достигают нормальных размеров. Еще через некоторое время появляются первые цветущие экземпляры, а в мае наблюдается массовое цветение. В начале июня парис отцветает, а в начале августа дает зрелые ягоды. Осенью листья увядают, и при первых же заморозках отмирает весь надземный побег. От растения остается лишь подземное корневище с хорошо заметным следом прикрепления надземного побега и крупной шпоровидной почкой возобновления на верхушке. Ранней весной, когда показывается молодой побег, почка возобновления, прикрытая той же самой шпоровидной чешуей (низовым листом), что и сам побег, находится еще в покое. Но к моменту зацветания надземного побега почка возобновления трогается в рост и пробивается сквозь прикрывающую ее чешую. К моменту массового цветения почка увеличивается в полтора-два раза и состоит из 2 хорошо отличимых частей - плотного, утолщенного основания и несколько суженной верхушки. Верхушечная почка покрыта плотной беловатой чешуей. Если снять эту чешую, то в ней можно обнаружить целых 2 зачаточные почки: корневищную треугольной формы и стеблевую. Стеблевая почка, прикрытая двумя узкими чешуями - зачатками предлистьев,- заключает зачатки как листьев, так и сегментов околоцветника. Значительно сложнее строение расположенной рядом с ней корневищной почки. В ней под плотным чехликом находятся еще две почки: миниатюрная корневищная и лучше дифференцированная стеблевая с двумя чешуйками - предлистьями. В стеблевой почке, второй по счету, имеются не только зачатки листьев и околоцветника, но даже начавшие обособляться внутренние части цветка в виде бугорков. Вторая корневищная почка хотя и очень мала, но в ней уже обособлены стеблевая и корневищная части. Следовательно, в почке возобновления, трогающейся в рост во время зацветания молодого надземного побега, имеются зачатки 4 почек: конечной корневищной и 3 стеблевых. Интенсивный рост молодого корневища вороньего глаза с почкой возобновления начинается ко времени отцветания надземного побега, т. е. в начале лета. На этой стадии уже в самой верхушечной почке, третьей по счету, под прикрывающей ее чешуей можно легко отличить будущую стеблевую и корневищные почки. С боков к стеблевой почке прилегают два небольших выроста треугольной формы - будущие пленчатые предлистья, прикрывающие стеблевую почку и при развитии надземного побега остающиеся у его основания. Как указывает далее С. С. Трофимов, вот эта стеблевая почечка в большинстве случаев и является уем зачатком, из которого к осени сформировывается материнская почка, весной следующего года образующая надземный побег. В июне первое междоузлие корневища достигает половины нормального развития. Значительно увеличивается также верхушечная часть корневища, что в основном происходит за счет вытягивания второго междоузлия, пока еще прикрытого общим чехликом. Стеблевая почечка, обнаруживаемая после удаления этого чехлика, уже прекратила дальнейший рост. Так как молодое корневище париса обыкновенного всегда дает больше одного междоузлия (два или чаще три), то первая стеблевая почечка во всех случаях засыхает, что делает непонятным, почему она вообще образуется. Во второй стеблевой почечке высотой 1,5 мм, находящейся под внутренней чешуей, помимо зачатков листьев и сегментов околоцветника, хорошо видны зачатки 8 тычинок, имеющих вид сосочковидных бугорков, а также уже и гинецей. Со стороны корневищной почки стеблевая почка охватывается двумя предлистьями. В верхушечной (третьей) стеблевой почке уже хорошо заметны четыре зачатка будущих стеблевых листьев и куполовидный конус нарастания между ними. В течение второй половины июня молодое корневище достигает в длину 3,4-4 см. К этому времени полностью сформировывается первое междоузлие, на котором появляются небольшие корешки, и одновременно начинается активный рост второго междоузлия и верхушечная почка пробивает прикрывающий ее довольно плотный чехлик. Этот чехлик в дальнейшем сохраняется в виде тоненькой пленки у основания второго междоузлия, имея в своей пазухе редуцированную стеблевую почку. Вторая стеблевая почка в июне все еще продолжает увеличиваться и в ней хорошо видны зачатки листьев и часть цветка. Несмотря на это, из этой почки не всегда формируется та стеблевая почечка, которая даст надземный побег следующего года. Это происходит лишь в тех случаях, когда на молодом корневище формируются два сегмента. Тогда вторая стеблевая почка становится основной, начинает перетягивать к себе запасы питательных веществ, а самая верхушечная почечка, третья по счету, на короткое время задерживается в своем росте и развитии. При наличии на молодом корневище трех сегментов основная стеблевая почка образуется из почечки, находящейся в самой верхушечной корневищной почке, начинающей интенсивный рост с середины июня. В конце июня - в первой декаде июля уже можно безошибочно установить, будет ли корневище иметь два или три междоузлия. На корневищах с тремя междоузлиями вторая стеблевая почка прекращает свой рост, а затем засыхает, как и первая; в тех редких случаях, когда образуются четыре междоузлия, засыхают три почки. В зависимости от количества междоузлий на вновь образующемся корневище значительно меняется и процесс дальнейшего развития новой почки возобновления. При двух междоузлиях вторая стеблевая почка быстро увеличивается в размере, задерживая тем самым рост и развитие верхушечной пары почек. В ней на коротеньком стебельке имеются довольно хорошо обособившиеся зачаточные листья длиной до 2-3 мм, сегменты околоцветника, зачатки тычинок и гинецей, представленный четырьмя сосочковидными выступами. Конечная же почка временно приостанавливается в росте и поэтому оказывается у основания стеблевой почки. Она содержит, ломимо зачатка стеблевой почки с обособившимися зачатками листьев и сегментов околоцветника, еще две почки - корневищную и стеблевую. Следовательно, при двух междоузлиях на молодом корневище в конце июня - начале июля на оси первого порядка сохраняются зачатки двух стеблевых и одной корневищной почки. При трех же междоузлиях на корневище в конечной почке в это время обычно содержатся лишь два зачатка - корневища и стебля, но часто оба зачатка в виде слабо дифференцированных бугорков. В конце июля в основном заканчивается прирост корневища и на всех междоузлиях появляются корешки. Молодое корневище заканчивается почкой, заключенной в плотную пленчатую чешую и несколько изогнутой кверху. В крупных почках зачаточный побег будущего года вполне дифференцируется ко времени созревания плодов. -Он состоит из стебля высотой 4-5 мм с 2 предлистьями у основания и сложенных трубочкой листиков длиной до 8 мм, прикрывающих зачаточный цветок. В течение трех недель июля листья зачаточного побега увеличиваются в три раза. Крупнее становятся и сегменты околоцветника, а тычинки достигают в длину 3- 5 мм; гинецей уже дифференцирован на завязь и столбики с рыльцем. Ко времени засыхания надземного побега стеблевая почка, включенная в плотный пленчатый чехол, достигает в высоту 30 мм. Но, как отмечает С. С. Трофимов, несмотря на такое развитие стеблевой почки, в которой уже вполне обособилась даже часть цветка, в осеннем состоянии вороньего глаза поражает не стеблевая почка, а корневищная почка, т. е. почка возобновления будущего года. В ней имеются три зачатка стеблевых и один зачаток корневищной почки, что вполне обеспечивает формирование стеблевой почки в будущем году даже при наличии трех междоузлий на корневище. Осенью почки продолжают развитие до начала зимы. Подобным же образом происходит развитие у кавказского париса неполного (Paris incomplete), париса мутовчатого (P. verticillata) и других видов.
Сложность и своеобразие формирования молодого отрезка корневища и почки возобновления париса С. С. Трофимов связывает с присущим этому роду моноподиальным способом ветвления. У корневищных трав с симподиальным способом ветвления, например у купены (Polygonatum), дочерняя почка возобновления закладывается в пазухе низового листа только после сформирования корневища и обособления зачатка надземного побега. В предзимнем состоянии в этих почках обычно бывает несколько чешуй, будущих низовых листьев, прикрывающих конус нарастания. Ежегодный отрезок молодого корневища париса формируется при наличии зачатков двух-трех стеблевых почек. Но из закладывающихся стеблевых почек часть их недоразвивается и, оказавшись у основания второго или третьего междоузлия корневища, засыхает. Этой участи подвержена при двух сегментах на корневище половина почек, а при трех сегментах - две трети их. Поэтому С. С. Трофимов ставит вопрос о биологическом значении для вороньего глаза пазушных почек, неизбежно засыхающих при формировании ежегодного отрезка корневища. На этот вопрос он отвечает следующим образом. Если исходить из особенностей строения почки возобновления париса, у которого стеблевая и корневищная почки находятся под прикрытием общей чешуи и только в таком состоянии могут нормально развиваться, вполне можно допустить, что в прошлом корневище париса ежегодно образовывало только одно междоузлие. В таком случае из первого зачатка стеблевой почки могла формироваться материнская почка, на следующий год дающая надземный побег. Но в дальнейшем приспособительная эволюция париса пошла в направлении увеличения количества междоузлий, что лучше обеспечивало накопление запасов питательных веществ и формирование более разветвленной корневой системы.
Корневище париса обычно не ветвится, но нередко наблюдается ветвление путем заложения боковых почек в пазухе чешуй - низовых листьев у основания междоузлия. Боковая почка не имеет никакой связи со стеблевой почкой, остающейся в пазухе чешуи и потом засыхающей. Как отмечает С. С. Трофимов, вегетативная почка проходит сложный путь развития. В первый год она представляет собой плотный мясистый вырост, прикрытый в нижней части остатками чешуи и не имеющий ни зачатков листьев, ни каких-либо иных образований. Только на второй год на этом выросте закладывается стеблевая почка возобновления, а надземный побег, по-видимому, образуется только на второй год.
Таким образом, у париса продолжительность малого цикла развития, начинающегося с заложения почки возобновления и образования надземного побега, охватывает три вегетационных сезона. Заложенные летом дочерние почки перезимовывают в материнской почке. На будущий год из второй или третьей по счету дочерней почки сформировывается почка возобновления, которая лишь через год дает надземный побег с продолжительностью вегетирования около полугода (С. С. Трофимов, 1956).
Как же опыляются цветки париса? Как это ни странно, несмотря на широкое распространение этого рода, особенно париса обыкновенного, мы не имеем пока еще вполне ясного представления о биологии его опыления. В классической европейской литературе по биологии цветка париса обыкновенного мы найдем следующие сведения. Цветки лишены запаха и нектара. Они протогиничны: рыльца бывают готовы к восприятию пыльцы еще в то время, когда цветок только начинает раскрываться. Пыльники раскрываются лишь несколькими днями позже. Обычно считается, что темно-пурпурные завязи и рыльце привлекают мух, принимающих их за гнилое мясо. Но в действительности растение ничего не предлагает мухам, которые фактически остаются обманутыми.
Цветок-обманщик очень хорошо использует посетителей: когда пыльники раскрываются, насекомые почти неизбежно обмазываются пыльцой, которая переносится ими на другие растения. Так как рыльце на этой стадии еще продолжает функционировать, то легко может произойти и самоопыление. Однако чехословацкий ботаник Э. Дауманн (1959) пришел к выводу, что насекомые не посещают регулярно цветки этого растения и фактически оно не является энтомофильным. Наблюдения в природе и характер пыльцы и рыльца говорят о том, что опыление происходит при помощи ветра. На поверхности пыльцевых зерен Э. Дауманном были обнаружены остатки липкого вещества, которые, по его мнению, свидетельствуют о том, что у этого растения имеет место вторичная анемофилия. Что же касается самоопыления, то этот вопрос он оставляет открытым.
Близок к парису, но в некоторых отношениях хорошо от него отличается род кинугаса (Kinugasa) с единственным видом кинугаса японская (К. japonica), которая встречается в субальпийских хвойных лесах и в верхней полосе буковых лесов северных и центральных районов острова Хонсю на высоте от 1000 до 2100 м над уровнем моря, где растет вместе с такими замечательными растениями, как ранзания японская (Banzania japonica) и глауцидиум пальчатый (Glaucidium palmatum). В отличие от париса корневище у кинугасы толстое и очень короткое, восходящее, диаметром 1,5-2 см. Стебель высотой от 30 до 80 см несет (6) 7-10 (12) листьев. Листья обычно обратнояйцевидные или продолговатые, на верхушке внезапно суженные, в основании постепенно переходят в короткий черешок. Цветоножка короче листьев, шириной 3-8 см. Чашелистиков 7-10 (12), ланцетно-продолговатых, длиной 2-5 см. В отличие от париса чашелистики кинугасы лепестковидные, белые, с зеленой средней жилкой, позднее красноватые или зеленоватые. Лепестки линейные, на верхушке слегка расширенные, вдвое короче чашелистиков. Пыльники линейные, без надсвязника. Пыльцевые зерна более крупные, чем у париса. Завязь полушаровидная, в верхней части с очень узкими крыльями, 6-9-гнездная, на верхушке усеченная и с углублением, увенчанная желтоватым диском; столбиков 6-9, шиловидных, утолщенных, длиной 3-7 мм, на верхушке крючковидно загнутых, у основания сросшихся в очень короткую колонку. Плод - ягода, которая, в отличие от ягод париса, съедобна, по сообщению профессора X. Хара, кисло-сладкого вкуса. Семена сходны с семенами париса. Кинугаса цветет в июне - августе и, по всей вероятности, опыляется насекомыми.
Род даисва (Daiswa) отличается как от кинугасы, так и особенно от париса прежде всего своими плодами и семенами. В то время как у первых двух родов плод - типичная ягода и при созревании не раскрывается, у даисвы он представляет собой нечто среднее между ягодой и коробочкой. Он мясистый, иногда даже сочный, и красного цвета (например, у высокогорной даисвы фиолетовой - D. violacea), и поэтому многие авторы называют его ягодой. Но, с другой стороны, плоды даисвы при созревании раскрываются локулицидно, и ряд авторов считает их коробочкой. Иного типа также семена даисвы. В отличие от сухих семян париса и кинугасы семена даисвы имеют хорошо развитую сочную саркотесту оранжевого или ярко-красного цвета - приспособление к распространению птицами. Завязь даисвы яйцевидная или пирамидальная, на верхушке усеченная, с более или менее ясно выраженным диском на верхушке, по бокам ребристая и часто с узкими низбегающими крыльями вдоль ребер, а столбики относительно короткие и толстые, шиловидные, свободные или более или менее сросшиеся в обычно короткую колонку, с рыльцевой поверхностью на внутренней стороне. Корневище обычно толстое и короткое. Как по характеру корневища, так и по некоторым особенностям завязи (низбегающие крылья) даисва несколько сходна с кинугасой, но резко отличается от париса. Виды даисвы имеют довольно различный облик и размеры. Размеры колеблются от маленькой, распространенной в Восточных Гималаях и в Юго-Западном Китае высокогорной даисвы фиолетовой до очень высокой, распространенной в Северном Вьетнаме и на острове Хайнань даисвы хайнанъской (D. hainanensis), у которой одна только цветоножка длиной до 125-150 см, а само растение нередко превышает в высоту 2 м. Листьев в мутовке от 4 до 22. Они на черешках или реже сидячие, иногда очень крупные, обычно к основанию суженные, но иногда основание более или менее сердцевидное. Чашелистиков от 4-5 до 10, зеленых, сходных с чашелистиками париса. Лепестки такого же типа, как у париса, короче или длиннее чашелистиков, по направлению к верхушке обычно несколько расширенных, нередко желтых, оранжевых или пурпурных. Тычинок от 6 до 24 (у даисвы хайнаньской изредка даже до 36), обычно без надсвязника или с коротким надсвязником (который у даисвы хайнаньской достигает, однако, в длину 4,5 мм, а у даисвы тибетской - D. thibetica - даже до 12-16 мм). Число столбиков, а следовательно, и число плодолистиков также очень различно, варьируя от 3 у даисвы фиолетовой до 10 у даисвы юнънанъской (D. ynnnanensis). Самым распространенным видом является даисва многолистная (D. polyphylla), ареал которой почти совпадает с ареалом рода.
О биологии опыления видов даисвы мы знаем очень мало. Известно только, что они строго протогиничны и опыляются насекомыми. Но так как функционирование рыльца продолжается достаточно долго, то время от времени должно происходить также самоопыление. Во всяком случае, как показал японский ботаник Хироши Хара (1969), у даисвы многолистной нет самонесовместимости, и в культуре по крайней мере может происходить самоопыление.
Перейдем теперь к самому большому в семействе роду - триллиуму. Как и остальные три рода, это многолетние корневищные травы. Корневище, как правило, толстое, диаметром обычно от 2 до 5 см или больше, но у триллиума яйцевидного (Trillium ovatum) всего лишь от 0,5 до 1,5 см, а у триллиума крошечного (Т. pusillum) - 0,3-1 см. Оно большей частью короткое, от 2 до 5 см, реже до 8 см и больше.
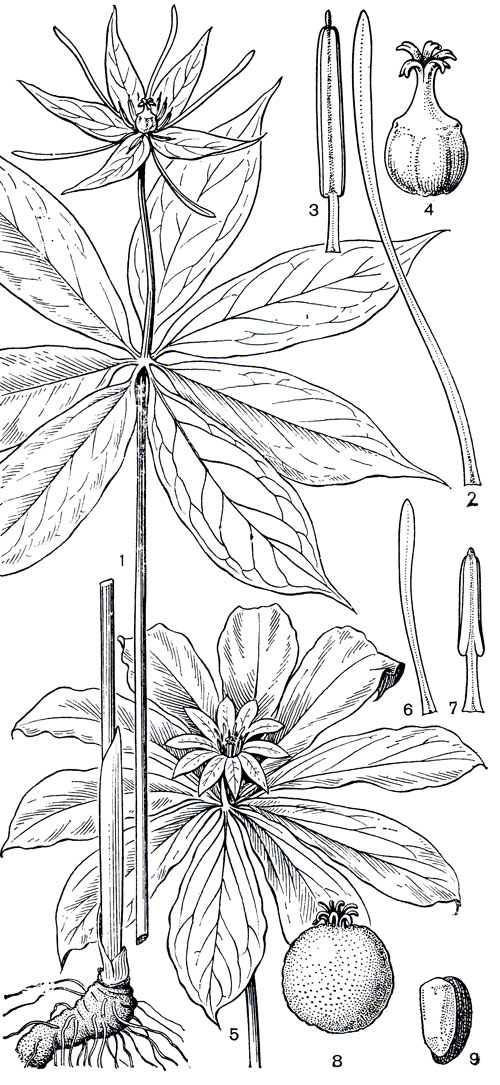
Рис. 120. Триллиевые. Даисва многолистная (Daiswa polyphylla): 1 - общий вид; 2 - лепесток; 3 - тычинка; 4 - гинецей. Кинугаса японская (Kinugasa japonica): 5 - часть растения с цветком; 6 - лепесток; 7 - тычинка; 8 - плод; 9 - семя с присохшим остатком части пульпы
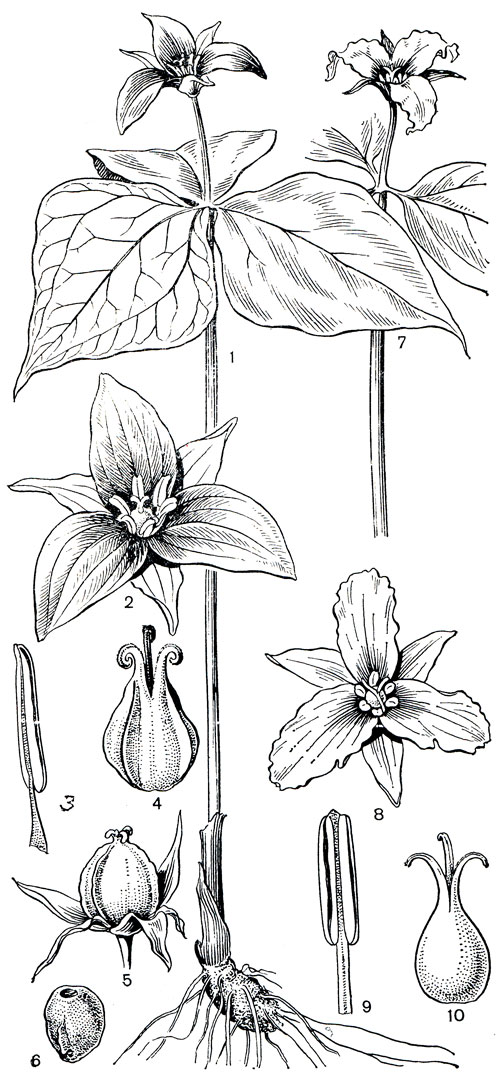
Рис. 121. Триллиевые. Триллиум прямостоячий (Trillium erectum): 1 - общий вид; 2 - цветок; 3 - тычинка; 4 - гинецей; 5 - плод; 6 - семя. Триллиум волнистый (Т. undulatum): 7 - часть растения с цветком; 8 - цветок; 9 - тычинка; 10 - гинецей
Корневище более или менее прямостоячее или реже горизонтальное (например, у триллиума отогнутого - Т. recurvatum), снабженное многолетними корнями и большой верхушечной почкой. Оно растет очень медленно (годичный прирост всего лишь 1-2 мм) и обычно живет не более 15 лет. Когда корни достигают определенной длины, они укорачиваются и втягивают корневище вниз, углубляя его в почву. Так как контрактильны только молодые корни, втягивание происходит лишь на растущем конце корневища, в результате чего корневище сохраняет прямостоячее положение. Верхушечная почка возобновления имеет много общего с почкой возобновления париса и других родов триллиевых. Она довольно большая, более или менее удлиненная и коническая, в диаметре лишь немного меньше самого корневища и покрыта тонкими бурыми чешуями. Верхушечная почка содержит не только стеблевую почку для следующего вегетационного сезона, а также две или больше стеблевых почек для последующих сезонов. Обычно первая из этих стеблевых почек не развивается дальше. У некоторых видов на корневище возникают боковые отростки, которые со временем отделяются от материнского растения и дают начало новым особям. Ранней весной из верхушечной почки корневища появляется над землей побег обычно с 3 (очень редко в виде исключения 2 или 4-8) листьями, сложенными в виде конуса вокруг бутона. Стебли высотой от 10 до 60 см, но иногда гораздо короче (у триллиума нивалъного - Т. nivale - всего от 3 до 15 см), а триллиум черешчатый (Т. petiolatum) фактически бесстебельный. Удлинение стебля сопровождается раскрыванием цветка и увеличением размеров листьев. Мейоз и развитие женского гаметофита начинается еще до раскрытия цветка.
Цветки триллиума сидячие (как у триллиума зеленоватолепестного - Т. chloropetalum - и триллиума черешчатого) или чаще на цветоножке длиной от 0,5 до 12 см. Чашелистиков 3, обычно зеленых, при плодах увеличивающихся. Лепестков 3 (редко 4-8), крупнее чашелистиков, от линейно-ланцетных до яйцевидных, белых, желтых, зеленых, розовых или пурпурных. Тычинок 6 в 2 кругах; нити короткие, плоские; пыльники линейные, обычно без надсвязника, редко с коротким надсвязником (триллиум зеленый - Т. viride). Завязь яйцевидная или шаровидная, одногнездная, но у триллиума яйцевидного (Т. ovatum) вторично 3-гнездная (в результате срастания постенных плацент в центре полости завязи), более или менее ребристая. Столбики более или менее свободные и с рыльцевой поверхностью на внутренней стороне, редко столбики сросшиеся почти до верхушки.
Как и у всех остальных триллиевых, цветки триллиума протогиничны. Они опыляются пчелиными (пчелами и шмелями) или мухами. Цветки, опыляемые пчелиными, имеют нектар (выделяемый септальными нектарниками) и приятный запах, иногда даже запах лимона или розы. Сюда относятся, например, триллиум сидячий (Т. sessile), триллиум поникший (Т. cernuum), триллиум крупноцветковый (Т. grandiflorum), триллиум желтый (Т. lnteum). Цветки, опыляемые мухами, например цветки триллиума прямостоячего (Т. erectum) или триллиума клиновидного (Т. cnneatnm), лишены нектара и характеризуются неприятным для человека запахом (но привлекательным для мух). Лепестки таких цветков темно- или коричнево-пурпурные, т. е. цвета гниющего мяса. У некоторых видов триллиума, например у триллиума крупноцветкового, наряду с перекрестным опылением наблюдается запасное самоопыление. У некоторых видов, как триллиум зеленоватолепестный и триллиум отогнутый (Т. recurvatum), перекрестное опыление заменилось самоопылением. В результате перехода к самоопылению у некоторых видов триллиума, например у триллиума Смолла (Т. smallii), встречающегося в Японии, на Курильских островах и на острове Сахалин, лепестки обычно отсутствуют. Отсутствуют лепестки, как мы видели, и у некоторых видов париса и даисвы.
Плоды триллиума двух типов - раскрывающиеся, коробочковидные, как у триллиума яйцевидного, триллиума зеленоватолепестного и большинства остальных видов, и нераскрывающиеся, сочные, ягодовидные или даже настоящие ягоды, как у триллиума Смолла, триллиума камчатского (Т. camschatense), триллиума волнистого (Т. undulatum), триллиума крупноцветкового или триллиума прямостоячего. Коробочковидные плоды варьируют от шаровидных до яйцевидных. У триллиума яйцевидного вдоль плода тянется 6 ясно выраженных ребер или килей, но у большинства других видов с коробочковидными плодами они едва выражены или отсутствуют. Вполне зрелые коробочковидные плоды приобретают желтый или реже белый цвет, а мезокарпий у них становится губчатым или мучнистым. Семена внутри таких плодов имеют клейкую поверхность и как бы склеены в одну общую массу. Эти коробочковидные плоды не являются еще настоящими коробочками, так как не имеют специальных приспособлений для раскрывания и при созревании открываются просто в результате чисто механического давления на околоплодник, производимого увеличившимися в размерах семенами, а также в результате постепенного разрушения ткани самого околоплодника. При этом местами наименьшего сопротивления оказываются основания плодов и линии вдоль швов, т. е. краев плодолистиков. Так, плоды триллиума зеленоватолепестного раскрываются вдоль основания и одного из швов, в то время как плоды триллиума яйцевидного раскрываются самыми разными способами, в том числе иногда септицидно, но только в самой верхней части плода. В отличие от настоящих коробочек раскрывание плодов происходит скорее неправильно. Как только плоды раскрываются в достаточной для этого степени, семена падают на землю. Обычно семена падают все вместе, но иногда отдельными группами, каждая из которых соответствует семенам, прикрепленным к одной плаценте. В отличие от плодов первого типа плоды второго типа мясистые, нераскрывающиеся и красного цвета. Они также ребристые или килеватые в разной степени. От плодов первого типа они отличаются главным образом наличием мякоти (пульпы), которая образуется из ткани плацент и в меньшей степени из мезокарпия. Плаценты при созревании становятся очень большими, желтеют и очень сочные. Их кисловатый сок содержит сахар и масла, причем последние приурочены только к определенным клеткам. Пульпа и плод в целом издают слабый запах, похожий на запах земляники.
В отличие от остальных трех родов триллиевых семена рода триллиум характеризуются наличием мясистого придатка, который образуется сначала как небольшое утолщение в верхней части шва рядом с короткой семяножкой. В морфологии семян такие придатки называют строфиолой. Придатки эти белого цвета, а поверхность самого семени светло-коричневая с темным пятном на микропилярном конце. По исследованиям Р. Берга (1958), придатки семени, представляющие собой настоящие элайосомы, привлекают муравьев, растаскивающих семена. При этом; не вполне ясно, имеет ли место у видов триллиума с плодами типа ягоды наряду с мирмекохорией также эндозоохория. Прямые наблюдения, к сожалению, отсутствуют, но, по мнению Р. Берга, это мало вероятно. Против возможности эндозоохории Берг выдвигает лишь один довод - загибание вниз цветоножки некоторых видов триллиума с плодами типа ягод, что явно уменьшает экспозицию плодов для пролетающих мимо птиц. Но полностью отрицать эндозоохорию все же нельзя, вполне возможно, что триллиумы диплохорны, т. е. распространяются двумя способами. Однако, как отмечает Берг, плоды типа ягод у мирмекохорных видов триллиума представляют собой загадку. Ведь муравьев привлекают только семена, а не плоды. Для чего же тогда мясистость плодов? По мнению Берга, первоначально плоды триллиума были ягодами и распространялись птицами. Однако сомнительно, чтобы раскрывающиеся коробочкообразные плоды триллиума возникли из ягоды. Скорее могло быть обратное. Легко представить себе происхождение ягод из раскрывающихся плодов с сохранением мирмекохории как запасного способа распространения. Ведь большинство лесных птиц, принимающих активное участие в распространении эндозоохорных плодов, обитает в верхних древесных ярусах леса, в то время как триллиумы - это растения травяного полога, где всюду снуют муравьи.
Среди триллиевых целый ряд очень декоративных растений. Это прежде всего виды рода триллиум, особенно триллиум крупноцветковый. Очень декоративны также кунигаса японская и некоторые виды даисвы. Однако лишь немногие из триллиевых вошли в культуру. Некоторый интерес представляют триллиевые и как лекарственные растения. В этом отношении наиболее известен парис обыкновенный, в плодах и корневищах которого содержатся сапонин (паристифин), гликозид паритин, а в корневищах - также алкалоиды. Ягоды и корневища очень ядовиты.
Семейство смилаксовые (Smilacaceae) (Р. А. Удалова)
Семейство объединяет 3 рода и 200-300 видов, распространенных очень широко по всему земному шару в тропических, субтропических и отчасти умеренных областях. Самый большой род смилакс (Smilax) содержит более 200 (по мнению некоторых авторов до 300) видов, произрастающих в обоих полушариях. Основное видовое богатство его сосредоточено в Юго-Восточной Азии, к северу виды смилакса распространены до Китая, Японии и Дальнего Востока СССР, где в Приморском крае растет смилакс прибрежный (S. riparia). Встречаются смилаксы и на островах Тихого океана. Из 11 видов, произрастающих на Филиппинских островах, 8 являются эндемичными. К западу ареал рода простирается до стран Ближнего Востока и Средиземноморья. Наиболее широкое распространение, от Юго-Восточной Азии до берегов Средиземного моря, имеет смилакс шероховатый (S. aspera) - один из трех видов, встречающихся в Европе. Средиземноморско-западноазиатский вид смилакс высокий (S. ехcelsa) растет на Кавказе в припойменных лесах из ольхи бородатой и бука восточного. Два вида смилакса встречаются в Южной и Юго-Западной Африке. В западном полушарии большое разнообразие видов смилакса представлено в Южной и Центральной Америке и в США, где в южных и восточных штатах произрастает более 20 видов. Второй род - гетересмилакс (Heterosmilax, рис. 123) - объединяет около 5 видов, распространенных в Восточной Индии, Таиланде, на полуострове Малакка, в Китае, Японии и на островах Тихого океана. Третий, наиболее примитивный род - рипогонум (Khipogonum) - состоит из 7 видов, произрастающих в Северной и Восточной Австралии, на Новой Гвинее и в Новой Зеландии.
Смилаксовые, как правило, корневищные вьющиеся или лазающие кустарники с древеснеющими стеблями, у большинства покрытыми колючками, часто с крючками. Встречаются в семействе и травянистые формы, как, например, эндемик тропических вечнозеленых лесов Таиланда смилакс незабудкоцветковый (Smilax myosotiflora). Немногие виды смилакса имеют прямостоячие стебли, среди них восточноазиатский вид смилакс круглолистный (S. cyclophylla), неветвящийся стебель которого достигает в высоту лишь 10-22 см. Характерные для семейства лианы имеют тонкие, диаметром 1,5-5 мм, упругие и прочные стебли длиной около 3-5 м. Отдельные виды, как, например, широко распространенный в Индии смилакс овалънолистый (S. ovalifolia), развивают стебли длиной до 10 м, а смилакс высокий - до 15 м.
Смилаксовые растут в лесах, преимущественно по опушкам. Их тонкие колючие стебли обвивают деревья, перебрасываются с одного дерева на другое, в результате чего создаются трудно проходимые заросли. Широко распространенный в Мексике и Центральной Америке смилакс густой (S. spissa) образует колючие непроходимые заросли по побережьям, поднимаясь на высоту лишь до 300 м, но наиболее характерны смилаксовые для горных лесов. В Юго-Восточной Азии они встречаются в листопадных муссонных лесах. Таким примером является таиландский вид смилакс многоколючковый (S. polyacantha), растущий в диптерокарповых, дубово-диптерокарповых лесах и лесах из тектоны на высоте 500-800 м. В Мексике большинство видов встречается по крутым склонам известковых гор в дубовых и дубово-сосновых лесах на высоте от 1800 до 2600 м над уровнем моря. Максимальной высоты (до 2900 м) достигает смилакс жесткий (S. rigida) в горах Непала. Смилаксовые, как правило, нетребовательны к почве. В Китае виды смилакса произрастают в смешанных жестколистных лесах, зарослях бамбука на бедных песчаных и гранитных субстратах. Виды рипогонума в Австралии растут иногда в колючих зарослях кустарников - скрэбах, к таким видам относится рипогонум Илей (Rhipogonnm elsey), встречающийся на юге Квинсленда. Единственный новозеландский вид рипогонум лазающий (R. scandens) является неотъемлемым элементом подокарповых лесов (рис. 123).
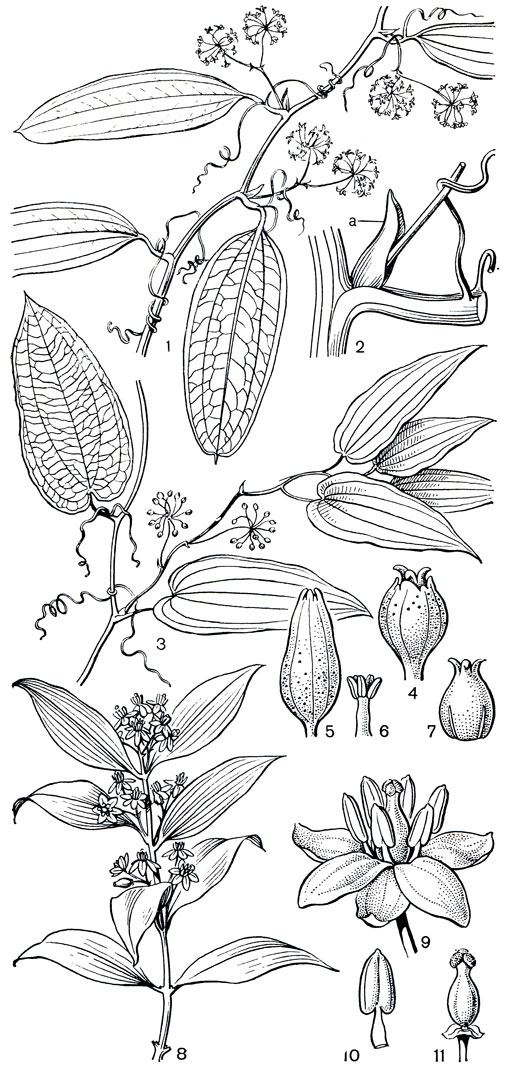
Рис. 123. Смилаксовые. Смилакс перевернутый (Smilax inversa): 1 - часть ветки с соцветием мужских цветков; 2 - профилл и основание соцветия в пазухе листа (а - профилл). Гетеросмилакс многотычинковый (Heterosmilax polyandra): 3 - часть ветки с соцветием женских цветков; 4 - женский цветок; 5 - мужской цветок; 6 - тычинки; 7 - гинецей (видны два стаминодия). Рипогонум лазающий (Rhipogonum scandens): 8 - часть ветки; 9 - цветок; 10 - тычинка; 11 - гинецей
Листья смилаксовых очередные, более или менее кожистые, по форме от округлых и яйцевидных до продолговатых, удлиненных и стреловидных. У рипогонума они располагаются супротивно. Нижние листья недоразвиты и имеют вид чешуй. Пластинка листа покрыта густой сетью жилок, среди которых выделяются от 3 до 7 главных жилок. Для листьев смилаксовых характерно хорошо развитое влагалищное основание. У индийского смилакса прямокрылого (Smilax orthoptera) оно разрастается по краям в своеобразные крылья (рис. 122,6). Влагалище заканчивается парой усиков, которые отсутствуют у рипогонума. Смилаксы большей частью листопадные растения. У одних видов лист сочленен с черешком в его средней части и опадает вместе с частью черешка; у других листовая пластинка сочленена с черешком в его верхней части и опадает отдельно от черешка. У смилаксовых в листьях, стеблях и корнях имеются сосуды с лестничной перфорацией.
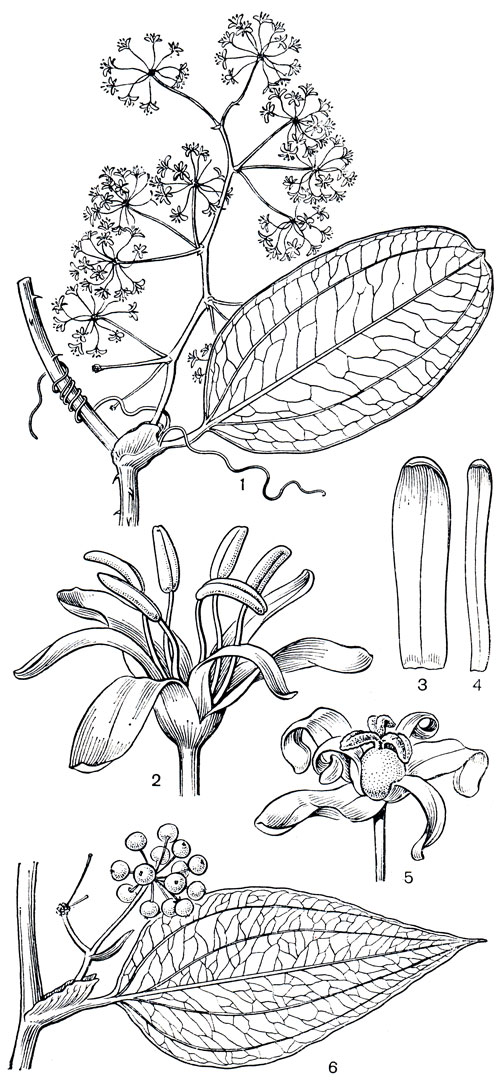
Рис. 122. Смилаксовые. Смилакс сиамский (Smilax siamensis): l - часть ветки с листом и соцветием мужских цветков; 2 - мужской цветок; 3 - наружный сегмент околоцветника; 4 - внутренний сегмент околоцветника; 5 - женский цветок. Смилакс прямокрылый (S. orthoptera): 6 - часть ветки с листом и плодами
Цветки смилаксовых обычно однополые и двудомные и лишь у рипогонума обоеполые, собраны в сложное пазушное кистевидное или колосовидное соцветие, состоящее из парциальных соцветий зонтиков. У многих видов, например у смилакса перевернутого (S. inversa, рис. 123), число зонтиков сокращается до одного-двух. Сегментов околоцветника 6; они свободные у смилакса и рипогонума и сросшиеся у гетеросмилакса. Сегменты внутреннего круга более узкие. Тычинок в мужских цветках 6, а у гетеросмилакса от 3 до 12. Тычиночные нити большей частью свободные и длинные, у смилакса голого (S. glabra) и рипогонума очень короткие, сильно утолщенные, а у гетеросмилакса они срастаются в колонку. Пыльники продолговатые, эллиптические или шаровидные, большей частью одногнездные, интрорзные. Пыльцевые зерна 1-бороздные, иногда с трехлучевой бороздой или безапертурные, с очень мелкими шипиками. Гинецей синкарпный, из 3 плодолистиков. Столбик короткий с тремя удлиненными лопастями или очень толстыми, почти приросшими (у рипогонума). Завязь верхняя, 3-гнездная, в каждом гнезде от 1 до 2 анатропных или гемианатропных висячих семязачатков.
Мелкие и невзрачные, большей частью зеленоватые или белые цветки смилаксовых имеют сильный запах. У некоторых видов он неприятен и напоминает запах падали или гниющих фруктов. Такие цветки опыляются мухами. Приятно пахнущие цветки посещают пчелы и жуки. По П. Кнуту (1904), смилакс травянистый (S. herbacea) опыляется настоящими складчатокрылыми осами, наездниками, жуками, а смилакс безусиковый (S. ecirrhata) также борборидами (Borboridae) и мухами-журчалками (Syrphydae). Насекомых привлекает в цветках как пыльца, так и нектар. Нектарники находятся в мужских цветках в основании тычиночных нитей, а в женских - в основании завязи. По данным Ф. Дельпино (1903), биологическое значение имеют также экстрафлоральные желёзки, находящиеся на кончике самых молодых листьев.
Плоды смилаксовых - сочные ягоды красного, черного или голубовато-сизого цвета с 1 - 6 семенами. Семена с твердым эндоспермом и маленьким зародышем. Распространение их осуществляют птицы. Г. Ридли (1930) указывает, что плоды смилакса округлолистного (S. rotundifolia) поедает американская ворона (Corvus brachyrhyncos), а плоды новозеландского рипогонума лазающего - ворона глаукопис (Glaucopis wilsoni). Американский странствующий дрозд использует в пищу плоды североамериканского вида смилакса сизого (S. glauca). Распространяют семена также дятлы. Сумчатые животные поссумы используют в пищу плоды рипогонума.
Семейство смилаксовых делится на два подсемейства: рипогоновые (Rhipogonoideae) с единственным родом рипогонум и смилаксовые (Smilacoideae) с родами смилакс и гетеросмилакс.
Некоторые виды смилакса с древних времен использовались в медицине. Утолщенные корни и корневища этих растений получили название "сарсапариль". Любопытно, что в русском языке это название имеет до десятка различных орфографических вариантов: сарсапарель, сассапарель, сассапариль и др. Все они восходят к испанскому zarzaparilla - сарсапарилья, составленному из двух слов: zarza - ежевика или вообще колючий кустарник и parilla (уменьшительное от раrrа) - вьющаяся лоза. Корни сарсапариля содержат крахмал, кристаллы оксалата кальция.
Основное действующее вещество - это сапонины стероидного характера: париллин, сарсапонин. Видами, дающими сарсапариль, являются смилакс, или сарсапариль медицинский (S. medica), родом из Мексики и сарсапариль полезный (S. utilis), известный также под названием "гондурасский" или "ямайский" сарсапариль. В китайской фармакопее широко используют сарсапариль китайский (S. china).
Семейство диоскорейные (Dioscoreaceae) (И. В. Грушвицкий)
К семейству диоскорейных относится 6 родов и более 700 видов, подавляющее большинство которых (вероятно, около 650) составляют род пантропического распространения диоскорею (Dioscorea). Из других родов диоскорейных только американский род рейания (Rajania) представлен относительно большим числом (25) видов; остальные, распространенные в восточном полушарии, олиготипны или монотипны. Род тамус (Tamus) включает 4 вида, стеномерис (Stenomeris) - 2, аветра (Avetra) и трихопус (Trichopus) представлены каждый одним видом. Эти роды характеризуются весьма ограниченными ареалами, в отличие от ареала рода диоскорея, границы которого почти везде совпадают с пределами распространения семейства в целом. На широчайшей площади ареала диоскореи, в какой-то степени захватывающего все пять материков, ареалы остальных родов представляют небольшие островки, занимающие (все вместе взятые) гораздо меньшую площадь, чем ареал одной лишь диоскореи клубненосной (Dioscorea bulbifera, рис. 128).
Диоскорейные - многолетние травянистые и кустарниковые растения весьма своеобразного габитуса. В подавляющем большинстве случаев это вьющиеся растения с подземным запасающим клубнем. Поражает кажущееся несоответствие между большой массой этого хранилища запасных веществ и всего одним тонким и недолговечным, чаще всего однолетним вьющимся побегом, как правило не успевающим за один сезон существования подняться по своей опоре на большую высоту. Листья диоскорейных очередные, реже супротивные, достигающие иногда больших размеров, характеризуются пальчатым жилкованием с анастомозами между жилками 1-го порядка, чем создается сетчатость, как известно характерная главным образом для двудольных. Цветки большей частью мелкие, собранные в соцветия разных типов, двудомные, реже обоеполые, актиноморфные, 3-членные. Околоцветник из 6 сегментов, расположенных в 2 круга, большей частью сросшихся в короткую трубку. Тычинок 6, также расположенных в 2 круга, причем 3 внутренние иногда превращаются в стаминодии или вообще не развиваются. Нити тычинок прикреплены к трубке околоцветника; пыльники интрорзные или реже экстрорзные, у стеномериса и аверы с длинным надсвязником; пыльцевые зерна большей частью однобороздные. Гинецей синкарпный, с 3-гнездной нижней завязью и свободными или более или менее сросшимися столбиками. В каждом гнезде завязи развивается по 2, реже по 3-4 и совсем редко по многу анатропных семязачатков. Плод диоскорейных обычно коробочка, иногда нераскрывающаяся крылатка (рейания), редко ягода (тамус). У многих видов семейства крылатые семена. Маленький зародыш заключен в роговидный эндосперм.
Семейство диоскорейных состоит из 2 подсемейств: диоскорейные (Dioscoreoideae) и трихоподовые (Trichopodoideae), последние с единственным представителем трихопусом цейлонским (Trichopus zeylanicum). Подсемейство собственно диоскорейных включает две трибы - стеномерисовых (Stenomerideae) и диоскорейных (Dioscoreae).
К трибе стеномерисовых принадлежат 2 маленьких рода - стеномерис, представленный 2 близкими видами, и монотипный род аветра. Для этой трибы характерны относительно крупные и обоеполые цветки. Оба вида стеномериса распространены в Малезии: стеномерис диоскореелистный (Stenomeris dioscoreifolia, рис. 124) - на Филиппинских островах, а стеномерис борнейский (S. borneensis), кроме Филиппин (Минданао), также на островах Калимантан и Суматра и в южной части полуострова Малакка. Это обитатели в основном низинных, реже низкогорных тропических дождевых лесов, растения тепло- и влаголюбивые.
От короткого горизонтального корневища стеномерисов отрастает длинный побег с гибким, прочным стеблем, несущим очередные листья. Побеги обвивают опору влево. Сползанию же этих гладких стеблей по опоре, очевидно, препятствуют сильно развитые, иногда удлиненные подушечки в основании листовых черешков. Сами листья крупные (длиной до 25, шириной до 19 см), сердцевидные, с длинно заостренным кончиком и с жилкованием как у диоскорей (рис. 124). Цветки стеномерисов, расположенные в пазушных цимоидного типа метельчатых соцветиях, висячие, хорошо приспособлены к перекрестному опылению насекомыми. Их цветоложе разрастается в кувшинчатую околоцветниковидную камеру-урночку, заключающую в себе репродуктивные части цветка. Основанием для этой урночки служит 3-гнездная нижняя завязь с многочисленными семязачатками; короткий столбик в глубине цветка заканчивается 3 двураздельными рыльцами. По краям зеленой урночки расположены 6 ярко окрашенных (желтых у стеномериса диоскореелистного и красных у второго вида) сегментов околоцветника, образующих при распускании цветка довольно крупную (диаметром до 3,5 см) розетку, обращенную вниз. Очень своеобразно строение и расположение 6 тычинок. Уплощенные тычиночные нити, прикрепленные близ узкого зева околоцветника, направлены не из цветка, а в глубь цветка - внутрь урночки, что определяет расположение интрорзно раскрывающихся пыльников у ее внутренних стенок. Наконец, надсвязники пыльников заходят в глубь урночки до самых рылец и, срастаясь между собой и со столбиком, образуют удобную посадочную площадку для насекомого (рис. 124,5), нагруженного пыльцой с другого растения. Осуществив опыление, насекомое спускается по внутренней стенке урночки и захватывает при этом пыльцу из пыльников. После опыления и оплодотворения гинецей сильно разрастается, образуя длинную (до 30 см и более), темно-бурую и блестящую коробочку, вскрывающуюся створками вдоль всей своей длины и освобождающую многочисленные (до 100) крылатые семена, распространяемые токами воздуха.
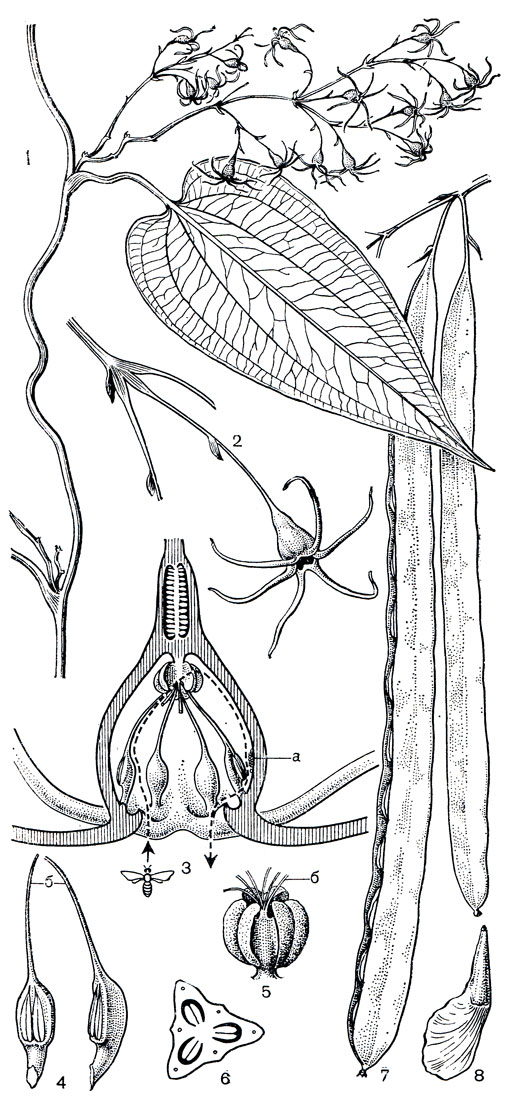
Рис. 124. Стеномерис диоскореелистный (Stenomeris dioscoreifolia): 1 - ветвь с соцветием; 2 - часть ветви с цветком; 3 - цветок в разрезе (а - пыльца, выпавшая из пыльника на внутреннюю стенку урночки; прерывистая линия - путь насекомого опылителя в цветке); 4 - тычинки (б - надсвязники); 5 - столбик с рыльцами (б - обрывки приросших к столбику надсвязников); 6 - завязь в поперечном разрезе; 7 - плоды; 8 - семя
А ветра вечнозеленая (Avetra sempervirens) является эндемиком восточного побережья Мадагаскара, где она довольно обычна в прибрежных лесах, в частности встречается и в окрестностях города Антананариву. У аветры (рис. 125), как и у стеномерисов, запасающим органом является корневище, листья на тонком упругом стебле очередные, довольно крупные, но побеги обвивают опору не влево, а вправо. Отличается аветра и по строению и расположению цветков, а также по характеру плодов. Цветки аветры довольно крупные, длиной до 3 см, чисто-белые. Они повисают на длинных тонких цветоножках на фертильных побегах и расположены поодиночке или в коротких кистях (по 2 и до 10), имеющих в основании мелкие скученные прицветники. Приспособление аветры к перекрестному опылению сходно с тем, что уже описано у стеномерисов. Многочисленные посетители ее цветков из мелких насекомых, проникая в суженный зев околоцветника, проползают в находящейся глубже зева маленькой "опылительной камере" сперва над рыльцем, а затем, перед тем как покинуть цветок, под пыльниками. В результате пыльца из одного цветка обязательно переносится на рыльце другого, возможно, и цветка другой особи аветры.
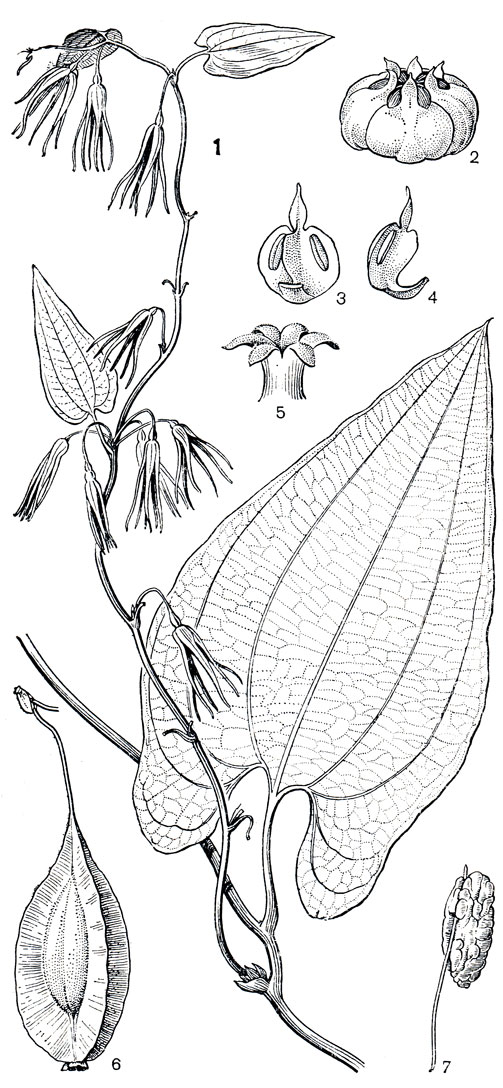
Рис. 125. Аветра вечнозеленая (Avetra sempervirens): 1 - ветвь с цветками; 2 - цветок (околоцветник удален); 3-4 - тычинки с надсвязниками; 5 - столбик с рыльцами; 6 - плод; 7 - семя (с глубокоруминированным эндоспермом) на длинной семяножке
Цветение аветры приурочено к концу относительно сухого периода, а односемянные невскрывающиеся плоды-крылатки созревают уже в дождливый сезон. В насыщенном влагой воздухе семя в ряде случаев начинает"прорастать в плоде, еще висящем на ветви, или в ветреную, штормовую погоду плоды отрываются от ветви, относятся ветром и прорастают после сгнивания перикарпия уже на почве.
Триба диоскорейных охватывает подавляющее большинство видов семейства. За небольшим исключением это вьющиеся растения с клубнями, в то же время растения двудомные, с мелкими однополыми цветками. Впрочем, у некоторых видов женские цветки содержат недоразвитые тычинки, а мужские - рудиментарный гинецей. У тычинок в этой трибе отсутствуют надсвязники, столь характерные для трихоподовых.
Центральное место в трибе диоскорейных, как и во всем семействе, занимает род диоскорея, многочисленные виды которого распространены во всех тропических областях мира, исключая лишь некоторые обширные крайне засушливые территории Африки (Сахара) и Евразии (Аравийский полуостров). Меньшим числом видов представлены диоскореи в субтропиках и, наконец, единичными видами в теплоумеренных областях (Восточная Азия, Кавказ, Балканы, Северная Америка). На каждом материке свои виды; единственным исключением, как уже отмечалось, является диоскорея клубненосная, встречающаяся на обширных пространствах и в Африке, и в тропической Азии.
Н. И. Вавилов (1935), а вслед за ним английский ботаник Г. Беркилл (1960) центром происхождения рода диоскорея считали Восточную Азию. Именно здесь сосредоточены представители наиболее древней секции стенофора (Stenophora), для которой характерно образование в качестве запасающего органа корневища. Именно отсюда в отдаленные времена их предки мигрировали на запад до Европы и оставили на этом пути, как следы переселения, ареалы таких современных внетропических видов, как диоскорея японская (Dioscorea nipponica, рис. 126), диоскорея кавказская (D. caucasica) и диоскорея балканская (D. balcanica). Позднее и другим путем (через Берингию) виды секции мигрировали на северо-восток - в Северную Америку, где поныне сохранился также корневищный вид диоскорея мохнатая (D. villosa), распространенная от Мексики и Флориды до Канады. Но большая часть видов этой древней секции и ныне сосредоточена в Японии и Китае, а некоторые заходят в горы тропиков Южной и Юго-Восточной Азии, примером чего является индийская диоскорея дельтовидная (D. deltoidea), распространенная в Северо-Западных Гималаях, в высокогорных лесах на высоте от 1500 до 3000 м. Диоскорея балканская и диоскорея кавказская, древние реликтовые виды, имеют в настоящее время очень не-большие ареалы. Диоскорея балканская встречается только в пограничных районах севера Албании и юго-запада Югославии, а диоскорея кавказская (рис. 126) - эндемик Западного Закавказья. Это - травянистая лиана преимущественно ксерофитных лиственных (дубовых, дубово-грабовых) лесов и кустарниковых зарослей, образовавшихся после вырубки дубовых лесов. Этот вид распространен от уровня моря до высоты 1600 м.
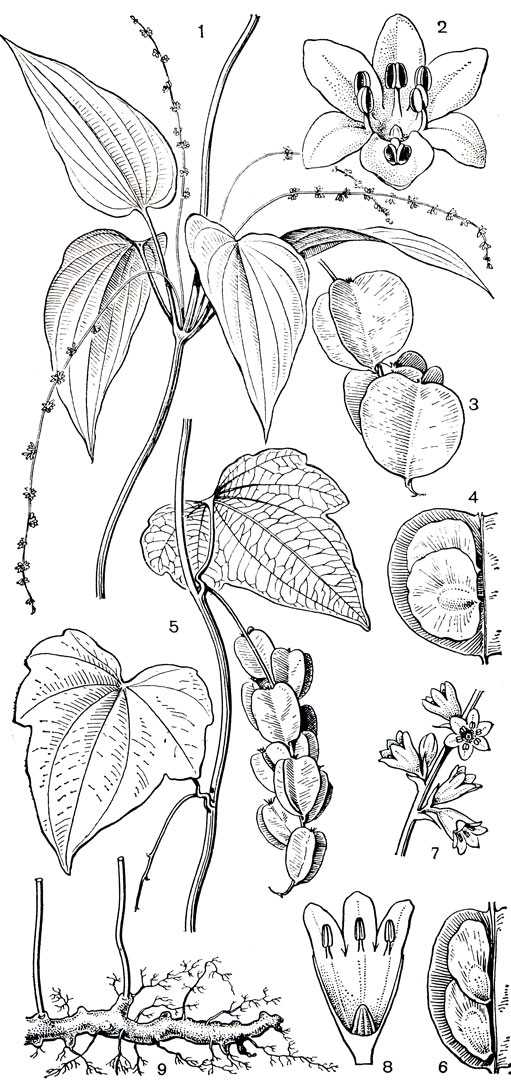
Рис. 126. Диоскореи. Диоскорея кавказская (Dioscorea caucasica): 1 - ветвь мужского растения; 2 - мужской цветок; 3 - плоды; 4 - часть коробочки с удаленной створкой (видны крылатые семена). Диоскорея японская (D. nipponica): 5 - ветвь женского растения с плодами; 6 - часть коробочки с удаленной створкой (видны крылатые семена); 7 - часть соцветия мужского растения; 8 - мужской цветок; 9 - корневище. В мужских цветках обоих видов (2 и 8) виден недоразвитый гинецей
Диоскорея японская (рис. 126) является эндемиком Восточно-Азиатской области. В пределах СССР этот вид распространен в Приморском крае, на юге Хабаровского края и на юго-востоке Амурской области; диоскорея японская приурочена к осветленным вторичным растительным сообществам - низкорослым зарослям из дуба монгольского и кустарниковым зарослям, развившимся после пожаров и вырубки коренных лесов. Встречается она, но реже, и в долинных широколиственных лесах, а также в горных кедрово-широколиственных лесах, но не поднимается в горы выше 500 м.
Для видов диоскорей других секций характерно развитие подземного, иногда частично или почти полностью надземного запасающего клубня. В подавляющем большинстве случаев это вьющиеся растения, обвивающие свою опору то левосторонне, то правосторонне. Особенность эта весьма стабильна: то или другое направление обвивания опоры постоянно не только для каждого вида, но и для каждой секции рода. Вьющиеся надземные побеги диоскорей обычно не поднимаются на большую высоту, как у других лиан тропического леса. Основная причина в том, что эти облиственные побеги недолговечны. У большинства видов они отмирают к концу первого же вегетационного периода, т. е. существуют от 6 до 12 месяцев. В начале нового сезона вегетации из почки на клубне развивается новый вьющийся побег. Эта особенность развития многих тропических диоскорей, вероятно, обусловлена приспособленностью к условиям относительно высокой освещенности. Очень многие из них растут в более или менее нарушенных лесах и в осветленных участках девственного тропического леса, в частности в "окнах", образующихся при падении лесных гигантов, сокрушающих при этом и более низкие деревья. Хотя тропические диоскореи и поднимаются в горы почти до 3000 м (как в провинции Юньнань Китая и в Гималаях), они значительно более разнообразны и обильны в низменных, в частности в прибрежных, лесах на больших островах и островках. У ряда видов - обитателей тропических дождевых лесов развиваются весьма длинные вьющиеся побеги со стеблями деревенеющими и колючими в основании, побеги, взбирающиеся до верхнего полога леса. Так, у африканской диоскореи Манжено (D. mangenotiana) измеренный побег оказался длиной более 40 м. Но даже у этой диоскореи толщина стебля не превышала 4 см, типичным же у видов рода является стебель в поперечнике всего около 1 см.
В составе рода имеются и низкорослые, карликовые виды диоскорей с короткими надземными побегами, прямостоячими и не нуждающимися в опоре, или с побегами, стелющимися по земле. Как считает Г. Беркилл (1960), предками этих видов должны быть типичные диоскореи с вьющимися побегами, но в ходе эволюции они приспособились к жизни в открытых растительных сообществах, особенно в скальных местообитаниях и в травянистых саваннах под влиянием яркого света, угнетающего рост, вызывающего сокращение длины междоузлий и подавляющего тем самым способность побега виться. Особенно интересна в этом отношении мадагаскарская диоскорея шестигранная (D. hexagona). Она обитает как в саваннах, так и в сухих лесах западных склонов гор и, соответственно, представлена здесь двумя формами роста. В лесу и в зарослях кустарников среди саванны стебель ее ветвистый и в верхней части вьющийся, с довольно широкими эллиптическими листьями, в то время как на открытых местах, в низкотравной саванне, стебель прямостоячий, высотой не более 40 см и с узкими ланцетными листьями на очень коротких (длиной 1-2 мм) черешках. Два вида, весьма сходных с этой, как правило, невьющейся мадагаскарской диоскореей, распространены в аналогичных условиях в кампосах Бразилии. Скальные карликовые диоскореи характерны и для Анд, распространяясь на север до Мексики. Наконец, два вида из этой своеобразной группы произрастают в Южной Европе, в Испанских (Восточных и Центральных) Пиренеях на известковых скалах, в расщелинах, на оползнях и каменистых осыпях. Диоскорея пиренейская (D. pyrenaica) - низкорослое травянистое растение высотой всего 5-10 см. Двудомное, как все диоскореи, она характеризуется четко выраженным половым диморфизмом: мужские растения более крупные, а женские - настоящие карлики. У тех и других в почве развивается слегка продолговатый маленький клубень, увенчанный у основания стебля скоплением чешуевидных листочков (рис. 127). От верхушки одиночного, короткого и прямого стебля отходят от 3 до 5 тонких восходящих веточек с маленькими сердцевидными листьями, из пазух которых развиваются или мужские соцветия в виде простой кисти, или же женские малоцветковые (с 1-3 цветками) простые колосья. Этот и близкий к нему также пиренейский вид настолько отличаются от обычных, широкоизвестных вьющихся диоскорей, что до сих пор некоторые ботаники относят их к особому роду бордерея (Borderea). Но с этим трудно согласиться, в частности и потому, что на противоположном конце земного шара, в южноамериканских Андах и Мексике в сходных условиях произрастают габитуально такие же карлики, но, несомненно, относящиеся к другим секциям рода диоскорея.

Рис. 127. Диоскорея пиренейская (Dioscorea pyrenaica): 1 - цветущее женское растение; 2 - щенский цветок с недоразвитым андроцеем; 3 - завязь в поперечном разрезе; 4 - ветвь с плодом; 5 - мужское растение с соцветиями; 6 - мужской цветок; 7 - мужской цветок в разрезе; 8 - тычинка
Внутреннее строение стебля у вьющихся диоскорей и в целом у вьющихся диоскорейных, как это отмечено недавно анатомом из Ганы Эдвардом Айенсу (1972), характеризуется рядом своеобразных черт, отличающих их от вьющихся растений из всех других семейств. У расположенных в два круга проводящих пучков тяжи ксилемы и флоэмы совершенно необычно переплетены так, что на поперечном срезе стебля, а также черешка флоэма оказывается разбитой на несколько отстоящих друг от друга участков, примыкающих к ксилеме. Далее, некоторые ситовидные трубки настолько необычно широки в поперечнике, что их трудно не принять за сосуды. Наконец, в узлах стебля ситовидные трубки одного и другого междоузлия соединяются не прямо, а через так называемые гломерулы, или узловые сплетения, состоящие из нескольких расширенных рядов вытянутых живых тонкостенных клеток (рис. 128,4) с ситовидными полями, различимыми лишь при электронно-микроскопическом наблюдении. Ксилемные гломерулы узла также аномальны, они состоят из множества мелких трахеид. В то же время индийский специалист Ц. Карник (1971) доказал, что эти узловые сплетения являются местом заметной химической активности - постоянных изменений в содержании различных биологически активных веществ и их превращений.
Листья у диоскорей очередные, реже супротивные и даже мутовчатые. У некоторых видов листорасположение изменяется на протяжении побега, как это можно видеть у диоскореи кавказской; ее нижние листья собраны в мутовки, вышерасположенные - очередные или почти супротивные. Типичный лист в роде диоскорея простой и цельный (иногда с волнистыми краями), реже лопастный. У небольшого числа видов листья сложные - тройчатые и даже пятипальчатосложные. Есть основания считать сложные листья в эволюционном отношении производными от простых. Этот переход, очевидно, происходил независимо и параллельно в разных секциях рода и сопровождался увеличением общей ассимиляционной поверхности листа в целом. Размеры листовой пластинки (листочка у сложных листьев) колеблются в значительных пределах как у видов рода, так и на одном побеге. Листья чаще всего на длинных черешках с развитыми листовыми подушечками в местах сочленения как с листовой пластинкой, так и со стеблем. Это, как известно, типичный признак листьев тропических лиан и деревьев, позволяющий ориентировать листовую пластинку в наиболее выгодном положении по отношению к свету. В основании черешка у некоторых видов, особенно из секции стенофора, могут развиваться выросты, которые можно считать прилистниками. Но их образование непостоянно, и у разных видов они выполняют различные функции: у одних они увеличивают ассимиляционную поверхность, у других становятся колючками, у третьих укрепляют соединение черешка со стеблем. По форме листовые пластинки простых листьев у диоскорей обычно сердцевидные, с сильно выступающими ушками; жилкование у них дуговидное, до кончика листа доходит чаще всего 3 из 5, 7, 9 первичных жилок. Краевые жилки обеспечивают прочность этих "ветровых" листьев, чему, очевидно, служат и анастомозы между первичными жилками, образуемые жилками 2-го порядка и создающие общее сетчатонервное жилкование.
Кончик листа у ряда диоскорей сильно вытянут в капельное острие. У западноафриканской диоскореи длиннохвостой (Dioscorea macronra), распространенной в тропических дождевых лесах, оно достигает иногда в длину 8 см при длине листовой пластинки 15 см (рис. 128). Подобные капельные острия, свисающие от листа вниз, обеспечивают сток воды с его поверхности и воды, выделяемой гидатодами. Однако у ряда тропических диоскорей, в том числе у диоскореи длиннохвостой и диоскореи клубненосной, они проявляют интересную особенность внутреннего строения. Всю длину кончика листа пересекает здесь внутри сложная железистая система из полостей (кармашков) с щелевидным выходом на поверхность капельного острия (рис. 128,6). Железистый эпителий полостей выделяет в них слизь, в которой поселяются азотфиксирующие бактерии. Установлено также, что содержание азота в капельных остриях выше, чем в листовой пластинке. Наблюдалось, наконец, что листья диоскорей, в частности диоскореи клубненосной, постоянно посещаются муравьями, и высказывалась мысль, что эта диоскорея является мирмекофильным растением. Но выяснение истинных связей между названными организмами еще требует дальнейших исследований.
Среди интересных особенностей вьющихся побегов тропических диоскорей весьма своеобразной является способность к образованию в пазухах листьев воздушных клубней. Они характерны для множества видов этого рода из разных секций, но не образуются у примитивных трихоподовых, а также у диоскорей из древней секции стенофора. Воздушные клубни - это видоизмененные, укороченные и утолщенные пазушные побеги. Из обычных для диоскорей 3 пазушных почек, как правило, лишь одна (реже 2) развивается в воздушный клубень. У диоскореи клубненосной (рис. 128, 1) воздушные клубни небольшие, диаметром 5-7,5 см; они почти шаровидные, с поверхности бурые и покрытые бородавчатыми выростами. Эта диоскорея редко приносит плоды, и вегетативное размножение воздушными клубнями у нее в большой степени заменяет размножение семенами. Ко времени созревания воздушные клубни подсыхают и становятся плавучими. Опадая с растения, они или сразу попадают в текучие воды, или оказываются на почве, где их могут подхватить и унести ливневые потоки. Наконец, реки могут выносить их в море на волю морских течений. Предполагают, что именно так диоскорея (по-видимому, именно клубненосная) появилась на острове Кракатау вскоре после страшного извержения 1883 г., уничтожившего на нем почти всю растительность. Переносом воздушных клубней диоскореи занзибарской (D. sansibarensis) речными водами объясняют широкое распространение этого африканского вида, особенно обильного по речным долинам. Мясистые, богатые запасными веществами воздушные клубни могли бы стать легкой добычей растительноядных животных, если бы не защитное накопление в них токсических веществ, делающих их тошнотворными на вкус и ядовитыми.
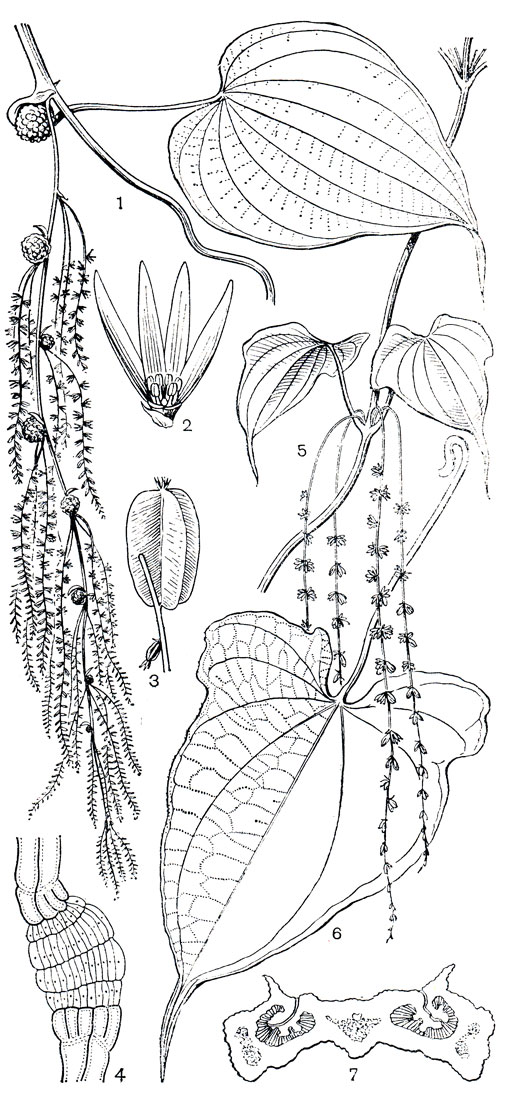
Рис. 128. Диоскореи. Диоскорея клубненосная (Dioscorea bulbifera): 1 - ветвь мужского растения с соцветиями и воздушными клубнями; 2 - мужской цветок; 3 - женский цветок и плод; 4 - флоэмная гломерула с промежуточной зоной из мелкоклетной проводящей ткани. Диоскорея длиннохвостая (D. macroura): 5 - ветвь мужского растения с соцветием; 6 - лист с капельным острием; 7 - капельное острие в поперечном разрезе (видны железистые вместилища слизи)
Все виды диоскореи, кроме представителей секции стенофора, развивают подземные клубни как запасающий орган, обеспечивающий развитие с каждым годом новых и новых недолговечных вьющихся побегов. В сравнении с корневищами клубни являются более продуктивным запасающим органом, способным накапливать большие количества питательных веществ и воды. Отсюда и возможность произрастания диоскорей в областях с продолжительной засухой - в саваннах Африки, Мадагаскара, Бразилии, в чилийских степях, в сухих субтропиках Средиземноморья. Вопрос о происхождении клубней продолжительное время был дискуссионным. В настоящее время можно считать, что у разных видов они формируются из различных частей молодого проростка: в одних случаях из гипокотиля, в других - из эпикотиля, в третьих - из первых двух междоузлий побега. Так или иначе, но клубни диоскорей следует считать по происхождению органами стеблевыми, а не корневыми. Различают 2 основных типа формирования системы зрелых клубней. У одних видов вслед за небольшим первичным утолщением в зоне гипокотиля (или эпикотиля) от него отходят тонкие горизонтальные столоны, на концах которых и формируются клубни (рис. 129), существующие всего один год. Такие клубни характерны для культивируемых диоскорей, в частности для диоскореи съедобной (D. esculenta), распространенной в природе от Индии до тихоокеанских островов. У других диоскорей образуется всегда один многолетний, из года в год увеличивающийся в размерах и массе клубень (рис. 129). Примером может служить южноафриканская диоскорея лесная (D. sylvatica), у которой клубень развивается из зоны гипокотиля. Окончательная форма этого многолетнего клубня, как указывает исследователь из Зимбабве Эйли Арчиболд (1967), в большой степени зависит от почвенных условий местообитания этого вида. На глинистой почве клубень дисковидный, с почкой возобновления ниже уровня почвы; на каменистых осыпях он может быть подземным и характеризуется наличием вмятин, где продолжающемуся росту клубня препятствуют крупные камни; на мелкой почве клубень частично выступает над поверхностью почвы и покрыт разноцветными "плитками"; наконец, на песках дюн клубень подземный, по форме напоминающий толстый блин с лопастями по краям. Но во всех случаях с возрастом срединная осевая часть подобных многолетних клубней деревенеет.
Рис. 129. Типы клубней у диоскорей: 1 - одногодичные подземные клубни; 2 - многолетний подземный клубень; 3 - многолетний полунадземный клубень
В сухих субтропиках Юго-Западной Африки, в области Намиб-Кару, к югу от реки Оранжевой, на высоте от 170 до 1300 м встречается другой представитель рода с тем же типом клубнеобразования - это знаменитая диоскорея слоновая, или слоновая нога (D. elephantipes),- одно из чудес здешней своеобразной природы. Особенностью этого вида является формирование с возрастом огромного, в основном (на 3/4 высоты) надземного клубня, мозаично одетого снаружи толстыми и прочными многоугольными плитками из пробки (рис. 129,5). Щели между плитками углубляются по мере разрастания клубня. Покров этот напоминает поверхность панциря черепахи, откуда бытующее название этой диоскореи "спящая черепаха". Считают, что у старых растений клубень достигает в высоту 60 см, но рекордные размеры в свое время указал английский ботаник У. Хукер. Описанный им экземпляр имел клубень высотой (над землей) 2 м 10 см, диаметром 90 см и массой около 315 кг. В Кейптаунском ботаническом саду в открытом грунте произрастала диоскорея слоновая с клубнем высотой 1,5 м и диаметром 5 м. Какой же возраст могут иметь такие гиганты, если даже для растения с клубнем высотой всего 30 см с учетом ряда признаков, в частности числа слоев пробки, был установлен примерный возраст в 100 лет!
Образование пробки здесь, как и у некоторых других однодольных (панданусов, некоторых пальм, агав, кордилин), своеобразно и отличается от этого процесса у двудольных. В этом случае паренхимные клетки все более глубокой закладки многократно делятся периклинально и затем суберинизируются. Образуется так называемая ярусная пробка. Что касается утолщения клубня, то оно обязано деятельности камбиеподобной ткани - специализированной первичной меристемы утолщения. Богатая крахмалом запасающая ткань лопастей клубня использовалась готтентотами в пищу, с чем связано еще одно название этой диоскореи - "готтентотский хлеб". Диоскорея слоновая обитает в колючих и суккулентных кустарниковых зарослях каменистых склонов на мелкой сухой почве, обвивая молодым побегом свой отмерший в предыдущем году побег или ветви ближайших кустарников, среди которых можно видеть виды родов алоэ (Aloe), толстянки (Crassula) и др. С этими суровыми условиями связано не только образование диоскореей мощного водозапасающего клубня, но и уменьшение размеров сизоватых от воскового налета листьев, в сухие дни складывающихся вдоль, а также полное отмирание листоносных побегов на засушливый период. Наконец, боковые побеги по опадении листьев, а также оси соцветий по отцветании остаются на клубне в виде острых твердых шипов, как защита от травоядных животных. У других диоскорей шипы на клубнях имеют корневое происхождение. В ряде случаев в шипы превращаются отходящие от клубня короткие придаточные корни после одревеснения их внутренних тканей и сбрасывания первичной коры. Иначе они образуются, например, у диоскореи съедобной, у которой в шипы превращаются боковые ответвления скелетных корней 1-го порядка; в этом случае в нескольких сантиметрах от поверхности клубня постепенно формируется колючий защитный чехол. Защитным приспособлением от животных у ряда диоскорей является также накопление в их клубнях токсических веществ, в частности алкалоида диоскорина. У некоторых диоскорей клубни особенно ядовиты. Отмечают, например, что кусок сырого клубня индо-малезийской диоскореи щетинковолосистой (D. hispida) размером с яблоко может вызвать смерть неразборчивого в еде человека. В Индии истолченные клубни этой диоскореи используют для приготовления ядовитых приманок для тигров. Малезийская диоскорея рыбаков (D. piscatorium) получила свое название в связи с использованием ее клубней местными рыбаками как средства для глушения рыб. В Африке клубни диоскореи занзибарской и диоскореи кустарниковой, или горькой (D. dumetorum), используют для получения стрельного яда.
Как было уже сказано, все виды диоскореи двудомны. Мы уже видели, как мужские и женские растения у диоскореи пиренейской отличаются даже габитуально. Но это исключение, и у большинства диоскорей различия между полами проявляются в основном в репродуктивной сфере. Прежде всего отметим, что мужские растения диоскорей намного обильнее в природе, чем женские того же вида, что их особи формируют больше соцветий, а сами соцветия крупнее и содержат значительно больше цветков, чем женские особи. В образовании такого изобильного, казалось бы расточительного, числа мужских цветков можно усмотреть "плату" за нелегкое обеспечение опыления и формирование плодов. Мужские соцветия у диоскорей достигают иногда больших размеров. Так, повисающие соцветия этого пола у диоскореи клубненосной иногда имеют длину до 1 м. Увеличение числа цветков при этом определяется также отчасти принадлежностью этих соцветий к типу верхоцветных - тирсов, у которых цветки на осях скучены в цимоидах. Женские же соцветия, как правило, не образуют на своих осях скоплений цветков и относятся к типу бокоцветных. Это или кисти, или колосовидные соцветия. Цветки, как правило, мелкие и невзрачные, диаметром всего от 2 до 5 мм, а у одного мадагаскарского вида даже 1 мм. Исключением являются цветки южноамериканской диоскореи крупноцветковой (D. macrantha) с диаметром цветка до 2,7 см. Околоцветник у большинства видов чаще всего чашечковидный, зеленоватый и только у немногих видов имеет более или менее заметную окраску - у некоторых видов он почти белый, у диоскореи клубненосной цветки красноватые, а у диоскорей (особенно американских), обитающих в сухих местах, цветки темноокрашенные, иногда пурпурные и даже почти черные.
Все диоскореи энтомофильны, о чем свидетельствует строение их цветков. Прежде всего цветки, особенно мужские, издают аромат. Так, приятным запахом, сходным с ароматом ладана - смолы некоторых бурзеровых (Burseraceae),- отличаются мужские цветки диоскореи клубненосной. Лопасти или сегменты околоцветника раскрываются не полностью, особенно у мужских цветков, которые даже в разгар цветения остаются почти сомкнутыми; шире раскрываются они у женских цветков. Тычинки, продуцирующие клейкую пыльцу, нередко скрыты в глубине цветка. У ряда видов из 6 тычинок полноценно развиваются лишь 3; тычинки внутреннего круга превращаются в стаминодии, расширенные в верхней части и загораживающие вход в цветок. У других видов внутренний круг тычинок вообще не развивается, но тогда тычиночные нити наружных тычинок раздваиваются и образуют те же 6 пыльников, что у видов с 6 фертильными тычинками. Прекрасным примером приспособления к насекомоопылению являются цветки диоскореи волосистоцветковой (D. hirtiflora), распространенной в тропической Африке. В мужских цветках здесь только сегменты наружного круга околоцветника слегка расходятся; сегменты внутреннего круга остаются соединенными на верхушке переплетающимися волосками. Ниже между сегментами остаются узкие длинные щели для прохода мелких посетителей. Последние находят путь между довольно крупными стаминодиями и крупными, также стерильными рыльцами в глубину цветка, где, как яйца в птичьем гнезде, на коротких искривленных нитях сидят пыльники трех фертильных тычинок. Опылителями диоскорей, скорее всего, являются мелкие летающие, очевидно, в основном ночные насекомые, сходные по размерам с трипсами (отряд пузыреногих).
Плод у диоскорей - 3-гнездная и 3-лопастная коробочка, в каждом гнезде которой развивается по одному или по два семени. У многих видов коробочка крылатая; в крылья здесь превращаются выступающие ребра плода. Значение этих крыльев достаточно загадочно, поскольку коробочка диоскорей - плод вскрывающийся, освобождающий крылатые семена. Раскрывается коробочка створками сверху вниз вдоль края крыла, после выпадения семян коробочка еще долго продолжает висеть на растении. Предполагают, что крылья плода могут каким-то образом содействовать освобождению и распространению семян. Крылатые семена диоскорей распространяются токами воздуха. У разных видов рода крылья имеют различный характер. У большинства видов с их почти шаровидными коробочками легкие плосковатые семена окружены очень тонким и бумажистым крылом равномерно по всей окружности. Выскальзывая из коробочки, такие семена могут парить в спокойном воздухе, постепенно снижаясь и отлетая от растения. У диоскорей с несколько продолговатыми коробочками характер крыла и способ распространения семян отличается. В этом случае, наподобие крылатки ясеня, крыло развивается только с одной стороны; у одних видов - с верхнего, у других - с нижнего конца семени. Семена такого строения, выпадая из плода, отлетают при порывах ветра, вращаясь вокруг своей оси, или опадают поблизости при полном безветрии. Интересно отметить, что у низкорослых, карликовых диоскорей способность семян к распространению токами воздуха в ходе эволюции была утеряна. Утрачены крылья, например, у диоскореи пиренейской. Семена, выпадающие из ее округлых коробочек, не относятся за пределы специфического местообитания этого карлика на каменистых осыпях, а попадают в какую-либо расщелину поблизости.
Из всего семейства диоскорейных именно диоскореи представляют наибольшее значение для человека. Клубни ряда диоскорей - один из древнейших видов пищи у народов тропических областей. Даже их возделывание, не говоря об уходящем в древность собирательстве, развилось независимо в тропических Африке и Азии, по-видимому, более 5000 лет назад. Американские индейцы возделывали местный вид диоскорею трехнадрезную (D. trifida) задолго до Колумба. Одним из наиболее древних центров культуры диоскорей явилась Западная Африка. Именно отсюда вошло в широкий обиход название - синоним для диоскорей в целом - "ямсы", как видоизменение их местных африканских названий. Именно здесь окончание сбора урожая ямсов издавна сопровождалось празднеством, эквивалентным Новому году. Здесь и независимо в других областях культивирования диоскорей (острова Малезии, Новая Гвинея, Новая Каледония, Фиджи) развился традиционный ритуал фестиваля, своего рода конкурса на самый крупный из выращенных клубней. Такие клубни-гиганты массой до 60 кг, иногда до 100 кг и больше и длиной до 2 м и даже до 6 м сохранялись до следующего урожая как "жилище ямсового духа". Но и в наше время ямсы являются главным пищевым продуктом примерно для 400 млн. человек, т. е. для каждого третьего жителя тропиков мира. Наиболее широко культивируемыми ради съедобных клубней видами являются в Африке диоскорея округлая, или "белый ямс" (D. rotundata), и диоскорея кайенская, или "желтый ямс" (D. cayenensis), а в Азии и на островах Тихого океана - диоскорея крылатая (D. alata) и диоскорея съедобная (D. esculenta). При этом диоскорея округлая и диоскорея крылатая являются культиварами, не встречающимися в природе в диком виде. Также только культивируемыми растениями представлена диоскорея супротивная (D. opposita), более известная как "китайский ямс". Клубни этих и других экономически гораздо менее важных видов, содержащие большое количество крахмала, а также протеин, витамины С и В и минеральные элементы, используются в пищу в вареном и жареном виде, а также для получения муки. Ямсы очень перспективная культура для будущего тропического земледелия.
Другой, не менее перспективной областью применения диоскорей оказалась медицина, и объектами использования здесь являются другие виды диоскореи - виды, содержащие в своих корневищах или клубнях биологически активные и в то же время токсические вещества. Ядовитые подземные и воздушные клубни диоскорей издавна применялись в туземной медицине как средство против ревматизма, кожных болезней, против укусов змей и так далее. Научное название "диоскорея" дано в честь прославленного врача древности Диоскорида (I в. н. э.) и было закреплено за родом Карлом Линнеем. Новая история медицински значимых диоскорей открылась после второй мировой войны, когда с особой остротой встала проблема гормонального препарата кортизона с его широким спектром терапевтического действия при тяжелых заболеваниях и в связи с крайне низкой обеспеченностью сырьем (единственным источником для получения кортизона - гормона коры надпочечников - были железы убойного скота). Новым сырьем для производства этого гормона должны были стать растения, содержащие стероидные соединения - "полуфабрикаты" для последующего синтеза кортизона. Наиболее перспективными в этом отношении и оказались диоскореи. На содержание в диоскореях такого "полуфабриката" диосгенина было исследовано 125 видов этого рода - 64 американских, 33 азиатских, 28 африканских. В итоге оказалось, что 60 видов содержат это соединение в заметном количестве, а из них выделилось несколько с особо повышенным его содержанием. Таковыми оказались прежде всего три мексиканских вида: диоскорея колосоцветковая (D. spiculiflora), диоскорея сложная (D. composite) и диоскорея обилъноцветущая (D. floribuncla) - с максимальным содержанием диосгенина в клубнях 15, 13 и 10% соответственно. Мексика и страны Центральной Америки стали основными поставщиками сырья для промышленного производства диосгенина. Ценным источником сырья стала также индийская диоскорея дельтовидная. В связи с последовавшими массовыми заготовками сырья всех названных видов их запасы в природе в последнее время сильно сократились. Выход из положения находят в создании плантаций для культивирования отобранных и созданных селекцией форм этих ценных растений. В СССР ведутся успешные опыты по введению диоскореи дельтовидной в культуру изолированных тканей.
Ценными лекарственными растениями являются и наши отечественные диоскореи. Корневища с корнями диоскореи кавказской и диоскореи японской, содержащие стероидные сапонины, стали в последнее время сырьем для производства лечебных препаратов диоспонина и полиспонина. Из трех видов диоскореи, произрастающих дико в СССР, два включены в Красную книгу как нуждающиеся в охране. В первую очередь - это диоскорея кавказская, запасы которой быстро сокращаются в результате усиленного сбора корневищ как лекарственного сырья и очень медленного восстановления зарослей после заготовок. Решение об ограничении заготовок сбором только по лицензиям должно сопровождаться расширением масштабов выращивания в культуре. Другой вид - диоскорея тонконогая (D. tennipes) - произрастает в СССР в единственной точке - в долине одного ключа на острове Кунашир, вблизи горячего сернистого родника. В этом случае определена необходимость полной охраны путем создания специального заказника. В мерах охраны, несомненно, нуждаются и многие тропические диоскореи и диоскорейные из других родов, имеющие маленькие ареалы и редко встречающиеся в их границах. Так, все более редкой, а в некоторых районах и совершенно исчезающей становится в Южной Африке диоскорея слоновая, и не только из-за беспощадного сбора растениеводами-любителями, но и по вине ангорских коз, которых здесь разводят во все большем количестве.
На островах Карибского бассейна можно встретить весьма похожих на диоскореи представителей другого рода из той же трибы диоскорейных - виды рода рейания (Bajania), названного в честь известного английского ботаника Джона Рея. Рейанию можно бы назвать кубинским родом, поскольку из общего числа (25 видов) 15 являются кубинскими. К последним относится наиболее широко распространенная рейания сердцевидная (R. cordata), произрастающая, кроме Кубы, и на других Больших Антильских и Малых Антильских островах. Виды рейании - вьющиеся растения с тонкими, более или менее высоко взбирающимися надземными побегами и с подземными клубнями. Сходство с диоскореями обнаруживается и в строении очередных листьев с характерным дугонервным жилкованием. Наиболее существенным отличием от рода диоскорея является у рейании тип плода - невскрывающаяся односемянная крылатка с одним крыловидным выростом и с сохраняющимся остатком околоцветника.
Наконец, в Средиземноморской и Макаронезийской областях и в Западной Европе распространены виды третьего рода трибы диоскорейных - рода тамус (Tamus). Из 4 видов этого рода наиболее широко распространен тамус обыкновенный (Т. communis). Будучи в основном растением средиземноморским, он заходит далеко на север в Западной Европе, в целом же распространен от Атлантического побережья Европы на западе до Восточного Закавказья и Ирана на востоке и от Ирландии на севере до Атласских гор на юге; в СССР встречается на Кавказе и в Крыму. Тамус обычен в средиземноморских маквисах, а в тепло-умеренной части ареала приурочен к лиственным и смешанным лесам, особенно к опушкам леса и к кустарниковым зарослям. Тамус обыкновенный - травянистая лиана. Его одногодичные неветвящиеся надземные побеги обвивают ветви кустарников и стволы невысоких деревьев, взбираясь по ним иногда на высоту до 6 м. Очередные листья, многообразные по форме, от цельных глубокосердцевидных или треугольных до почти трехлопастных, на длинных черешках и с капельными остриями,) подчас образуют выраженную листовую мозаику. Старые экземпляры с мощным клубнем могут развивать одновременно большое число (до 20 и более) вьющихся побегов, которые своей листвой, как шапкой, покрывают служащие им опорой кустарники. В почве разрастается многолетний удлиненный клубень. Первое время он расположен в ней почти горизонтально, позднее втягивается отходящими корнями и приобретает углубленное и почти вертикальное положение. При многолетнем росте в длину и вторичном утолщении с годами клубень становится достаточно мощным. Так, у одного измеренного 30-летнего растения клубень имел длину 52 см, толщину 20 см и массу 10 кг. Интересно, что число цветков у мужских растений у этого двудомного вида, как и у диоскорей, преобладает над числом цветков, образуемых женскими растениями. В длинных обращенных к свету мужских метелках может быть до 200 цветков, в то время как повисающие короткие и рыхлые кисти женских растений малоцветковые, несут до 20-25 цветков, но чаще и того меньше, так что соотношение между теми и другими становится как 60 : 1. Цветки тамуса обыкновенного мелкие (диаметром 3-4 мм), незаметные, желтовато-зеленые, со слабым ароматом. В то же время они продуцируют нектар, привлекающий разнообразных мелких насекомых-опылителей. Постоянными посетителями цветков тамуса, по наблюдениям в Европе, являются мелкие представители рода эмпис (Empis, семейство толкунчики из двукрылых).
Зрелые плоды тамуса - кораллово-красные, блестящие и ядовитые ягоды, резко выделяющиеся осенью на фоне желтеющей или буреющей отмирающей листвы этой лианы. Заключенные в них шаровидные семена твердые, как кость, в связи с отложением в оболочках клеток эндосперма в качестве запасного материала целлюлозы. В твердый эндосперм погружен маленький недоразвитый зародыш. Отсюда крайне замедленное прорастание семян: проходит от 2 до 3 лет, пока, попав в почву, семя прорастет. Очень своеобразно распространение этих семян. Они относятся к категории клейких; собственно, очень липкой является мякоть ягоды. При повреждении экзокарпия, например, улитками семена в клейкой массе мякоти выступают наружу и прилипают к перьям птиц или шерсти животных или, опав на почву, приклеиваются к опавшим листьям, которые затем уносятся ветром. Но для прорастания им необходимо теперь углубиться в почву, только здесь семена смогут пройти длительное дозревание, и это достигается их самозарыванием на глубину около 4-5 см вследствие чередования набухания и сморщивания семенной кожуры. Продолжает углубляться в почву и развивающийся клубень, так что у зрелого растения основание клубня нередко оказывается на глубине 10-20 см и даже более.
Клубни тамуса обыкновенного издавна применяют в разных странах в народной медицине. В клубнях содержатся биологически активные вещества, в том числе стероидные сапонины. Между прочим, из надземных побегов и листьев тамуса съедобного (Т. edulis), произрастающего на островах Макаронезии, был выделен тамусгенин, более, чем диосгенин диоскорей, приближенный к строению кортизона. К сожалению, использование побегов этого растения как сырья для производства гормональных препаратов типа кортизона крайне затруднено из-за примеси к тамусгенину большого разнообразия других стероидных генинов.
Обратимся в заключение ко второму подсемейству диоскорейных - к трихоподовым. К нему, как уже отмечалось, относится всего один монотипный род трихопус (Trichopus), единственный вид которого трихопус цейлонский (Т. zeylanicus) кроме Шри-Ланки распространен на юге Индии, в штате Мадрас, а также на полуострове Малакка. Это невысокое (всего около 25 см) дернистое травянистое растение (рис. 130) обитает в низинных тропических дождевых лесах по берегам рек, в условиях постоянной высокой влажности. В Индии трихопус встречается в горных тропических лесах на высоте около 1000 м. Запасающим органом здесь, как и у стеномерисовых, является корневище. У трихопуса оно тонкое, толщиной всего с карандаш, покрытое очень острыми чешуйчатыми листьями и образующее множество крепких проволоковидных корней. Корневище это горизонтальное и очень короткое (1-4 см), слегка восходящее на верхушке и отмирающее после опадания чешуек на противоположном конце. От корневища, иногда ветвящегося, отрастают 5-7 (иногда 20) прямых стеблей высотой до 12 см. Каждый из этих слегка угловатых стеблей несет только по одному листу, черешок и пластинка которого продолжают линию стебля. Листовые пластинки длиной около 10 см и шириной до 4 см весьма вариабельны по форме: от линейно-ланцетных до треугольно-яйцевидных и сердцевидно-стреловидных с клиновидным или глубокосердцевидным основанием (с сильно развитыми ушками) и с 5-9 дуговидными жилками, из которых только 3 достигают притуплённой коротко заостренной верхушки. Край листа волнистый. Листовые пластинки вытянуты вверх, и эпидерма на обеих сторонах листа этого тенелюбивого растения, в отличие от остальных диоскорейных, состоит из глубоко извилистостенных клеток. На верхушке стебля, сразу под черешком, развивается сдвинутая в боковое положение укороченная фертильная ветвь с двурядно расположенными тесно примыкающими друг к другу чешуйчатыми прицветниками. В целом эта веточка очень напоминает многоцветковый колосок злаков из рода костер (Bromus). В пазухах этих защитных чешуек-прицветников развиваются пазушные почки, которые пробуждаются по очереди, выбрасывая единовременно один или два цветка на длинных (до 7 см), тонких, на концах поникающих цветоножках. Только после опыления цветка выбрасывается новый из пазухи другой чешуйки, что уже является своеобразным приспособлением к перекрестному опылению. Столь своеобразный, неповторимый в семействе габитус был выработан в эволюции, очевидно, как неотеническая форма. Надземный побег ближайшего предка трихопуса, бывшего, очевидно, вьющимся растением, приобрел способность к переходу в зрелое состояние, т. е. к цветению и плодоношению на фазе ювенильного растения-предка и развитие даже второго листа подавляется ради более успешной деятельности листа первого и единственного. Таким образом, трихопус является как бы "взрослым проростком".
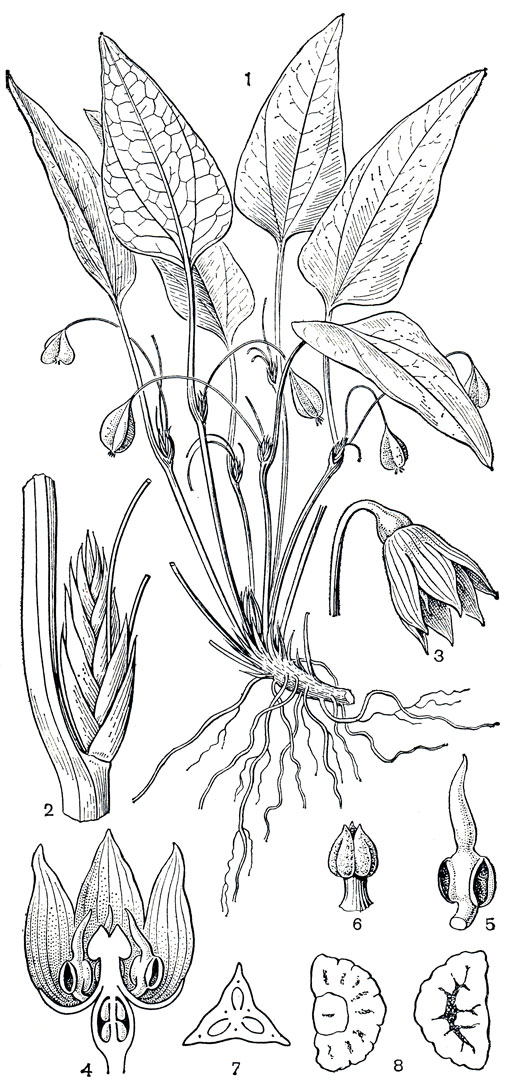
Рис. 130. Трихопус цейлонский (Trichopus zeylanicus): 1 - общий вид растения с плодами; 2 - укороченная фертильная ветвь; 3 - цветок; 4 - цветок в разрезе; 5 - тычинка с надсвязником; 6 - столбик с рыльцами; 7 - завязь в поперечном разрезе; 8 - плоское семя (вид с двух уплощенных сторон)
Обоеполые, актиноморфные цветки трихопуса имеют сростносегментный колокольчатый околоцветник шириной до 1,5 см. Трубка его зеленоватая, а лопасти тускло-пурпурные. Как у стеномерисовых, пыльники 6 тычинок снабжены надсвязниками, протягивающимися к расходящимся от очень короткого столбика рыльцам. Сами пыльники с широкими связниками смыкается край к краю и образуют как бы потолок над камерой, в которую должны заползти мелкие насекомые-опылители. При таком строении цветка механизм перекрестного опыления здесь должен быть сходным с характером опыления у аветры.
Плод у трихопуса - 3-гранная, обратнояйцевидная, суккулентная, ягодовидная коробочка с 3 утолщенными крыльями, при созревании раскрывающаяся на 3 створки. Распространение этих сочных плодов с заключенными в них плоскими семенами осуществляется, по предположению Г. Беркилла (1951, 1960), водой. Сильные дождевые потоки отламывают плоды от их тонких плодоножек и относят в сторону от материнского растения.
Семейство такковые (Тассасеае) (Т. В. Вельгорская)
Семейство такковые представлено одним родом такка (Тасса), насчитывающим около 10 видов, из которых 9 - обитатели тропиков Старого Света и только 1 вид - такка Паркера (Т. parkeri) произрастает в тропической Южной Америке (Венесуэла, Гайана, Колумбия и Северная Бразилия). Ареал такки леонтолепестковидной (Т. leontopetaloides) простирается от западного побережья тропической Африки до острова Пасхи, включая Мадагаскар, Сейшельские и Маскаренские острова, Юго-Западную и Южную Азию, остров Шри-Ланка, Малезию, Северную Австралию и острова Океании. Остальные 8 видов рода обитают преимущественно в Южной и Юго-Восточной Азии: в Восточной Индии, Бангладеш, Таиланде, Южном Китае, на полуострове Малакка, островах Суматра и Калимантан, в западной части острова Ява, доходят на востоке до Соломоновых островов.
Такковые - многолетние травы с ползучими или клубневидными корневищами, снабженными сосудами примитивного типа. Молодые части растений, как правило, бывают опушены мельчайшими волосками, исчезающими по мере взросления органа. Размеры растений обычно невелики, от 40 до 100 см, но некоторые виды (например, такка леонтолепестковидная) достигают иногда в высоту 3 м. Количество листьев и соцветий на каждом растении, как правило, невелико. Листья все прикорневые, крупные, на более или менее длинных ребристых мясистых черешках, цельные (такка цельнолистная - Т. integrifolia, рис. 131, табл. 27,5) или сильно расчлененные (такка палъчатонадрезная - Т. palmatifida). Обоеполые актиноморфные цветки на коротких цветоножках собраны в зонтиковидное верхушечное соцветие, окруженное покрывалом из 4 прицветников, в большинстве случаев расположенных в 2 круга. За исключением видов секции пальмотакка (Palmotacca) и такки Паркера в соцветиях имеются, кроме того, длинные (до 25 см) нитевидные поникающие прицветники.
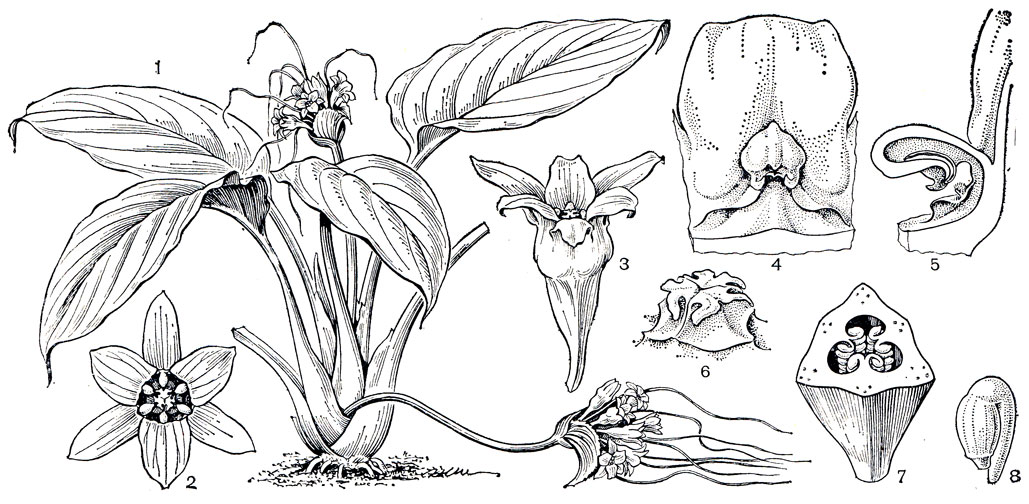
Рис. 131. Танка цельнолистная (Тасса integrifolia): 1 - общий вид; 2 - вид цветка сверху; 3 - вид цветка сбоку; 4 - тычинка; 5 - вид тычинки сбоку; 6 - рыльце; 7 - завязь; 8 - семязачаток
Околоцветник из 6 свободных сегментов, расположенных в 2 круга (сегменты внутреннего круга темнее окрашены), или сросшийся в короткую трубку, несколько венчиковидный. Тычинок 6, они расположены в 2 круга и при-креплены к околоцветнику короткими шлемовидными нитями (рис. 131,5-5). Пыльники интрорзные; оболочка пыльцевых зерен однобороздная. Гинецей паракарпный; столбик короткий, с 3 частолепестковидными и загнутыми над столбиком рыльцевыми ветвями (рис. 131,6); завязь нижняя с многочисленными анатропными семязачатками; эндосперм нуклеарный. У некоторых видов имеются нектарники или нектарниковидные желёзки. Плод - ягода, и только у одного вида - такки подорожниковой (Т. plantaginea) - локулицидная коробочка,. что позволило некоторым ботаникам выделить этот вид в самостоятельный род схизокапса (Schizocapsa). Семена многочисленные, с маленьким зародышем и обильным эндоспермом. Зрелые семена длиной до 5 мм, светло- или темно-коричневые, иногда с ариллусовидным мясистым образованием у рубчика; ребристая семенная кожура снабжена папиллами.
Род такка на основании морфологических признаков подразделяется на 3 секции. Виды секции атакка (Atacca) имеют цельные листья и длинные нитевидные прицветники. К этой секции относят также и южноамериканскую такку Паркера, у которой, однако, нет прицветников или 1-2 очень коротких. В секцию палъмотакка (Palmotacca) входят виды со сложными расчлененными листьями, без нитевидных прицветников. И наконец, представители третьей палеотропической секции эвтакка (Eutacca) имеют рассеченные листья и очень длинные нитевидные прицветники. Монограф семейства голландский ботаник Э. Дрене (1972) объединяет виды этой секции в один - такка леонтолепестковидная.
Произрастают виды такки в самых разнообразных экологических условиях. Они селятся на открытых и сильно затененных местах, в саваннах, в зарослях кустарников и в дождевых лесах. Их можно встретить на морских побережьях и в горных тропических лесах, иногда на высоте до 2100 м над уровнем моря (такка Шантръе - Т. chantrieri). Растет такка на самых разных почвах: на песчаных и каменистых субстратах, на гравии, суглинках, глинистых почвах, на известняках и вулканических породах. На побережьях островов Суматра и Ява такка образует растительные группировки с казуариной (Casuarina), панданусами (Pandamis); в лесах Северной Австралии такка леонтолепестковидная произрастает совместно с видами эвкалипта (Eucalyptus); на Маршалловых островах - с аргузией (Argusia) и веделией (Wedelia).
Цветет и плодоносит такка почти круглый год. Цветки ее не обладают ярко выраженными приспособлениями для привлечения опылителей, которые так характерны для большинства тропических растений (цвет, форма и т. д.). Тона их тусклые, темные, коричневато-пурпурно-зеленоватые или грязно-фиолетовые. У такки нет резких сильных запахов, и неизвестно также, выделяют ли нектар иногда имеющиеся нектарники или нектарниковидные желёзки. Таким образом, соцветия такки обладают совокупностью тех признаков, которые, по мнению К. Фэгри и Л. ван дер Пейла (1966), присущи сапромиофильным растениям, опыляемым мелкими насекомыми, чаще всего падальными или навозными мухами. Пищи для насекомых-опылителей в таких цветках нет, и цветок "обманывает" мух, стремящихся проникнуть в него. Возможно, мух привлекает слабый запах разлагающегося протеина, и мухи вползают внутрь цветка через небольшое отверстие, привлеченные блеском железистых эпидермальных клеток на "донышке" цветка. Не найдя внутри ничего для себя интересного, мухи стремятся быстрее покинуть цветок, но короткие шлемовидные тычинки и короткий столбик с ветвистыми долями рыльца мешают им беспрепятственно сделать это. В поисках выхода из ловушки насекомое неизбежно соприкасается с поверхностью пыльников или рыльца и, наконец, нагруженное пыльцой, выбирается из цветка, чтобы снова, соблазнившись блеском железистых клеток, попасть в кратковременный плен другого цветка. Не исключено также, что большие прицветники, образующие покрывало вокруг соцветия, могут служить убежищем для этих маленьких мух, а длинные сочные нитевидные при-цветники, возможно, и пищей для насекомых.
Способы распространения плодов и семян такковых весьма разнообразны. Плоды растущих на морских побережьях видов такки могут переноситься морскими течениями. Губчатая семенная оболочка обеспечивает плавучесть семян на многие месяцы. Во всяком случае, этим можно объяснить расселение некоторых видов на многочисленных островах Тихого океана, отдаленных друг от друга подчас на значительные расстояния. По сообщению Г. Ридли (1930), в распространении семян такки принимают участие и птицы, склевывающие плоды. Во влажном тропическом климате опавшие плоды такки быстро сгнивают и освободившиеся семена с мясистым ариллусовидным придатком у рубчика растаскиваются муравьями. Человек также играет немаловажную роль в распространении этого растения, возделывая некоторые виды такки далеко от мест их естественного обитания.
Издавна люди используют в качестве ценного пищевого продукта мучнистые клубни такки леонтолепестковидной, содержащие очень большое количество (до 25%) крахмала. Клубни очищают от кожуры, натирают и несколько раз тщательно промывают горячей и холодной водой, чтобы удалить содержащееся в них горькое и ядовитое вещество таккалин. Полученный крахмал обычно используют для выпечки хлеба, изготовления пастилы, халвы, для пудингов. Иногда крахмал такки смешивают с другими ингредиентами, например с соком молодого кокосового ореха, и используют для изготовления целебного напитка. В африканских странах употребляют в пищу и мякоть ягод. Правда, едят эти ягоды в основном дети. В Полинезии из волокон стеблей такки леонтолепестковидной делают шляпы и рыболовные сети. Мягкие листья и соцветия такки Шантрье употребляют в пищу, а из корневищ приготовляют лекарство. В Малезии клубневидные корневища такки пальчатой (Т. palmata) используют как средство от укуса змей. Такка подорожниковая, растущая в смешанных лесах по берегам рек в Южном Китае и Таиланде, находит применение в китайской народной медицине. Разводят некоторые виды такки и в качестве декоративных растений: фиолетовые и коричнево-пурпурные тона соцветий прекрасно сочетаются с яркой зеленью крупных листьев.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'