Порядок наядовые (Najadales) (Н. Н. Цвелев)
Семейство апоногетоновые (Aponogetonaceae)
Среди водных растений тропиков Старого Света заметное место принадлежит видам апоногетона (Aponogeton, табл. 1, 1) - единственного рода семейства апоногетоновых, естественный ареал которого включает Африку к югу от Сахары, Южную Азию и Северную Австралию.
В роде апоногетон около 45 видов. Особенно много видов этого рода на Мадагаскаре - здесь встречаются 11 видов, эндемичных для этого острова и близлежащих Коморских островов. В ископаемом состоянии (олигоцен - около 25 млн. лет назад) отпечатки листьев одного из видов этого рода были найдены далеко за пределами тропиков - в Западном Казахстане (С. Г. Жилин, 1974).
По внешнему облику виды апоногетона напоминают обычные в наших водоемах рдесты (Potamogeton), однако уже в отношении жизненной формы имеются существенные отличия: все виды апоногетона - розеткообразующие растения с выходящими из пазух листьев розетки безлистными цветоносами и клубневидно утолщенным симподиальным корневищем, от которого отходят многочисленные тонкие корни.
Листья апоногетона обычно отчетливо разделены на короткое влагалище, черешок и цельнокрайнюю пластинку, форма которой варьирует от линейной до яйцевидной, но чаще всего бывает эллиптической или ланцетной. Вполне сидячие линейно-ланцетные листья имеет африканский апоногетон валлиснериевидный (A. vallisnerioides), а у африканского апоногетона ситникового (A. junceus) листовая пластинка редуцирована до средней жилки, в которую непосредственно переходит черешок. У многих видов или только погруженные, или только плавающие листья (последние обычно с более широкими и менее прозрачными пластинками), но не так уж редко и те и другие есть в пределах одного и того же вида и на одном и том же растении. Обычно листовые пластинки имеют утолщенную среднюю жилку и несколько дополнительных дуговидных жилок, соединенных друг с другом и со средней жилкой сетью поперечных анастомозов. Замечательны листья у разводимого в аквариумах апоногетона мадагаскарского (A. madagascariensis), более известного под названием апоногетона продырявленного (A. fenestralis), а также "растения-сетки" или "растения-кружева" (рис. 9, 1). Его листовые пластинки уже в самом начале своего развития теряют ткань между жилками, действительно становясь похожими на мелкоячеистую сеть или кружево. Эта особенность в какой-то степени предохраняет листовую пластинку от повреждений при обитании в быстро текущих ручьях и речках.
Колосовидные соцветия апоногетоновых располагаются на длинных ножках, выносящих их над поверхностью воды. У многих видов соцветия - простые колосья со спирально расположенными на довольно толстой оси цветками, у других они от основания делятся на 2, реже 3-10 ветвей, причем в этом случае цветки часто располагаются лишь на одной стороне каждой из ветвей (рис. 9, 3). У основания соцветия имеется окутывающее его покрывало, обычно быстро опадающее после того, как соцветие выходит из воды.

Рис. 9. Апоногетоновые. Апоногетон мадагаскарский (Aponogeton madagascariensis): 1 - общий вид; 2 - часть листовой пластинки; 3 - часть соцветия; 4 - цветок; 5 - тычинка; 6 - плод. Апоногетон изящный (A. gracilis): 7 - общий вид; 8 - листовая пластинка; 9 - часть соцветия; 10 - листочек околоцветника; 11 - тычинка; 12 - листовка
Цветки апоногетоновых обычно обоеполые, реже однополые, зигоморфные или актиноморфные, первично 3-членные, без прицветников. Сегменты околоцветника все одинакового строения, чаще всего их 2, реже 3 (тогда третий меньше двух других), 1 или они вообще отсутствуют. У многих видов сегменты околоцветника лепестковидные и окрашенные в белый, розовый или желтый цвет, но остающиеся при плодах и зеленеющие, у других они с самого начала больше похожи на прицветники. У апоногетона двуколосого (A. distachyon) и некоторых других видов единственный сегмент околоцветника сильно увеличен (рис. 10,4). Тычинок обычно 6, расположенных в 2 круга, с довольно длинными нитями и мелкими двугнездными пыльниками, реже (иногда в пределах одного и того же растения) число тычинок достигает 12, и тогда они расположены в 3-4 круга. Пыльцевые зерна однобороздные. Гинецей состоит обычно из 3, реже 2-9 почти свободных сидячих плодолистиков, с 2-8 семязачатками в каждом плодолистике. Каждый плодолистик на верхушке оттянут в короткий столбик, несущий низбегающее рыльце в виде продольной бороздки на внутренней стороне (рис. 9, 6, 12).
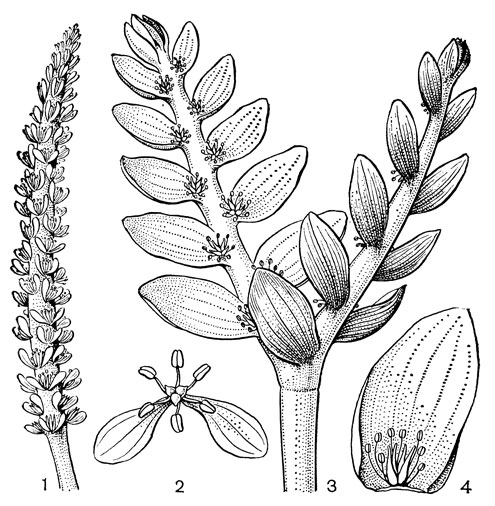
Рис. 10. Соцветия и цветки апоногетоновых. Апоногетон волнистый (Aponogeton undulatus): 1 - соцветие; 2 - цветок. Апоногетон двуколосый (A. distachyon): 3 - соцветие; 4 - цветок
Формирующийся из гинецея плод - многолистовка погружается в воду, где околоплодник постепенно разрушается, освобождая лишенные эндосперма семена, которые сразу падают на дно или некоторое время (до нескольких часов) плавают на поверхности. Прорастание семян происходит довольно быстро: от одного дня до нескольких недель.
В экологическом отношении некоторые виды апоногетона не имеют узкой приуроченности, другие, напротив, обитают или только в быстро текущих речках и ручьях, или только в стоячих водоемах. Есть виды, например австралийский апоногетон шестилисточковый (A. hexapetalus), растущие в водоемах, существующих только 3-4 месяца в году, а затем сильно пересыхающих. При обитании в относительно глубоких водоемах соцветия могут не достигать поверхности воды и оставаться заключенными в покрывало. В этом случае одни виды, например австралийский апоногетон пузырчатый (A. bullosus), не образуют плодов, другие, например апоногетон мадагаскарский, опыляются клейстогамно, образуя более мелкие плоды с небольшим количеством семян.
Способ перекрестного опыления у видов апоногетона еще недостаточно изучен. Присутствие септальных нектарников между плодолистиками и часто довольно яркая окраска цветков, собранных в соцветия, свидетельствуют в пользу энтомофилии, хотя возможность опыления цветков ветром не может считаться исключенной. Возможно также опыление полупогруженных в воду соцветий пыльцой, высыпавшейся на поверхность воды. Самоопыление обычно исключается за счет протандрии (рыльца становятся восприимчивы к пыльце лишь после высыпания ее из пыльников того же цветка), а у апоногетонов Трупина (A. troupinii) и двудомного (A. dioecus) - за счет перехода к двудомности. У некоторых видов однополые цветки встречаются и в пределах одного и того же соцветия. Так, у апоногетона Лopии (A. loriae) нижние цветки в соцветии обоеполые, а верхние мужские - с рудиментами плодолистиков, у апоногетона голоцветкового (A. nudiflorus) нижние цветки соцветия женские, не имеющие околоцветника. Южноафриканский апоногетон Ремана (A. rehmannii) - апомиктический вид, в его соцветиях имеются только женские цветки, а у апоногетона волнистого (A. undnlatns) из Индии обычна вивипария: соцветия видоизменяются в молодые растения, которые опускаются на дно и укореняются.
Во всех тропических странах, где встречаются виды семейства апоногетоновых, местные жители употребляют в пищу в печеном или вареном виде их богатые крахмалом клубнеобразные корневища. Кроме того, многие виды апоногетона культивируют в качестве аквариумных растений. Среди них особенно популярен южноафриканский апоногетон двуколосый, введенный в культуру уже с 1780 г. и натурализовавшийся во многих водоемах Западной Европы, Австралии и Южной Америки. Несколько реже встречаются в культуре апоногетон волнистый, апоногетон пузырчатый из Австралии, мадагаскарский апоногетон улъвовидный (A. ulvaceus) и апоногетон мадагаскарский. Последний вид, замечательный своими сетчатыми листьями, культивируют уже более 100 лет, однако в аквариумах он живет недолго и очень требователен к температурным условиям.
Семейство шейхцериевые (Scheuchzeriaceae)
Единственный род семейства шейхцериевых - шейхцерия (Scheuchzeria) - представлен всего двумя очень близкими видами, часто принимаемыми за европейский и североамериканский подвиды одного вида - шейхцерии болотной (S. palustris), широко распространенной в холодных и умеренно теплых областях северного полушария. У южной границы своего ареала шейхцерия известна лишь из немногих изолированных местонахождений, обычно в горных районах (Пиренеи, Альпы, Карпаты, Северные Кордильеры), но также и на равнинах. Например, в Харьковской и Воронежской областях СССР она встречается на реликтовых сфагновых болотах песчаных надлуговых террас, в речных долинах вместе с многими другими северными видами (пушицей, росянкой, клюквой и др.).
Шейхцерия болотная (рис. 11, 2, 2) - небольшое (всего до 20 см высотой) многолетнее болотное растение с длинными (до 0,5 м), часто разветвленными симподиальными корневищами, которые играют заметную роль в закреплении сфагнового покрова на болотах, особенно так называемых сфагновых мочажин. В цветущем состоянии шейхцерия малозаметна, так как ее кистевидные соцветия с невзрачными цветками лишь едва возвышаются над сфагновым мхом, а линейные, очередные, двурядно расположенные на коротком стебле листья можно принять за листья какого-либо представителя осоковых или злаков. У основания листьев имеются довольно длинные открытые влагалища, отделенные от пластинки поперечным перепончатым выростом - язычком, а немного ниже верхушки листа на его верхней стороне хорошо заметна почти округлая ямка - водяная пора, служащая, подобно гидатодам, для выделения избыточной воды. В пазухах листьев шейхцерии расположены многоклеточные волоски, гомологичные внутривлагалищным чешуйкам многих других водных и болотных однодольных. Во всех вегетативных органах много воздухоносной ткани, позволяющей побегам шейхцерии возвышаться над поверхностью воды.
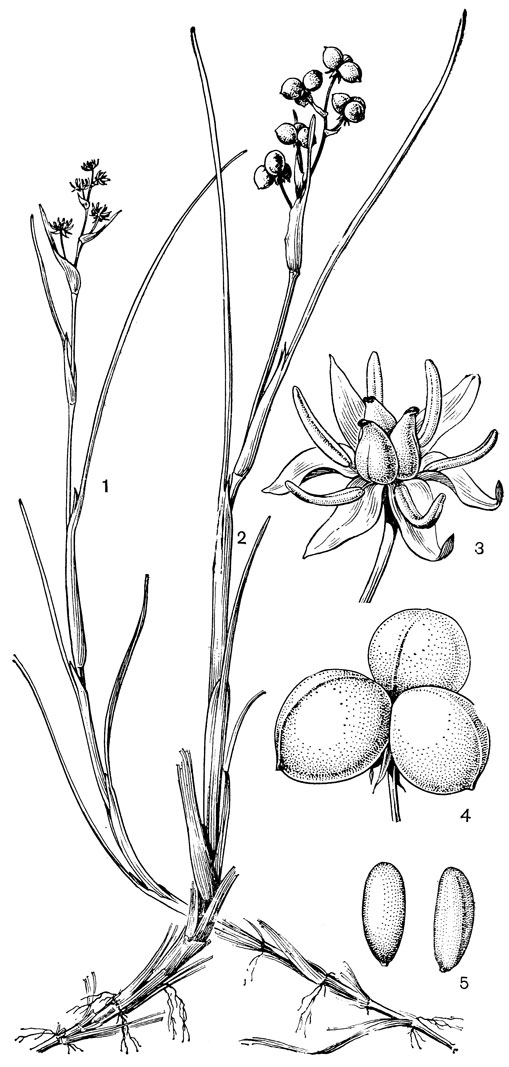
Рис. 11. Шейхцерия болотная (Scheuchzeria palustris): 1 - общий вид цветущего растения; 2 - общий вид плодоносящего растения; 3 - цветок; 4 - плод; 5 - семя
Заканчивающие собой облиственные репродуктивные побеги, кистевидные соцветия шейхцерии обычно состоят всего из 3-6(10) обоеполых актиноморфных цветков, расположенных на коротких ножках в пазухах прицветников. Околоцветник представлен 6 небольшими желтовато- или буровато-зелеными сегментами, расположенными в два 3-членных круга. Также расположенные двумя 3-членными кругами тычинки заметно выше околоцветника и состоят из довольно крупных красно-бурых пыльников на сильно удлиняющихся к концу цветения нитях. Связник пыльников вытянут на верхушке в заостренный надсвязник. Пыльцевые зерна безапертурные. Гинецей составляют 3, реже 2 или 4-6 лишь едва сросшихся у основания, а при плодах свободных плодолистиков. Каждый плодолистик несет 2, реже больше анатропных семязачатка и несколько суживается к расположенному на его верхушке сидячему дисковидному рыльцу, покрытому довольно длинными сосочками. При плодах репродуктивные побеги шейхцерии становятся более заметными, так как несут относительно крупные желто-зеленые плоды - много листовки, части которых сильно вздуты и вскрываются щелью по швам плодолистиков.
Шейхцерия принадлежит к числу ветроопыляемых растений, причем возможность самоопыления уменьшается за счет протандрии - более раннего созревания пыльников по сравнению с рыльцами того же цветка. Широкоэллипсоидальные, довольно крупные семена распространяются главным образом гидрохорно, так как обладают хорошей плавучестью благодаря наличию воздухоносной ткани в оболочке. Кроме того, шейхцерия быстро размножается вегетативным путем, так как ветви корневища быстро теряют связь с материнским растением. Возможен также перенос водой во время половодья небольших частей сфагнового покрова болота вместе с корневищами шейхцерии и других болотных растений.
Семейство ситниковидные, или триостренниковые (Juncaginaceae)
Представители этого небольшого, включающего всего 5 родов и около 20 видов, семейства мало бросаются в глаза из-за своего невзрачного облика, но интересны как группа, переходная от относительно примитивных, энтомофильных семейств однодольных, подобных частуховым, к анемофильным и гидрофильным семействам высокоспециализированных водных растений (рдестовым, дзанникеллиевым и др.). Ареал этого семейства охватывает почти все внетропические области как в северном, так и в южном полушарии, а также горные районы тропиков. Почти по всему ареалу семейства распространен род триостренник (Triglochin) с 15 видами, из которых 2 встречаются и на территории СССР. Остальные роды имеют более ограниченные ареалы: цикногетон (Cycnogeton) и маундия (Maundia) - в Австралии и Тасмании, тетронциум (Tetroncium) - во внетропической части Южной Америки и лилея (Lilaea) - вдоль Тихоокеанского побережья Америки от Юго-Западной Канады до Чили включительно.
Большинство видов ситниковидных - розеткообразующие многолетники с коротким, реже более длинным и разветвленным корневищем, двурядно расположенными линейными (иногда щетиновидными) листьями и колосовидными или кистевидными соцветиями на длинных безлистных ножках. К однолетникам, также имеющим собранные в розетку листья и безлистные ножки соцветий, принадлежат несколько австралийских видов триостренника, в том числе триостренник капканный (Triglochin calcitraра, рис. 12, 4). Обычно высота однолетников не превышает 10 см, а триостренник мельчайший (Т. minutissimum) представляет собой карликовое растеньице высотой 1,5-3(4) см с щетиновидными листьями и соцветиями с 4-11 цветками. Плоские, реже полуцилиндрические листья ситниковидных у основания расширены и образуют свободные (не сросшиеся краями) влагалища, а в месте перехода их в пластинки имеется поперечный перепончатый вырост - язычок. В пазухах листьев заметны характерные для многих болотных и водных однодольных внутривлагалищные чешуйки с выделяющими слизь железками.
Соцветия всегда заканчивают собой укороченные побеги с розетками листьев и не имеют у основания предлистьев. Прицветники в соцветии также отсутствуют. Цветки у разных родов семейства довольно сильно различаются по строению. У триостренника и цикногетона они обоеполые, актиноморфные и 3-членные. Очень мелкие и расположенные двумя кругами 6 сегментов околоцветника имеют оригинальное строение: с внутренней (верхней) стороны они сильно вогнуты и напоминают створку раковины. В каждом таком сегменте располагается почти сидячий двугнездный пыльник, прикрепленный к основанию листочка. Таким образом, тычинки располагаются здесь двумя кругами, но, в отличие от близкого семейства шейхцериевых, не чередуются с сегментами околоцветника, а противостоят им, что позволило некоторым авторам принимать сегменты околоцветника у ситниковидных за прицветники (брактеи), а весь цветок - за соцветие из нескольких однополых цветков. Шаровидные пыльцевые зерна имеют гладкую оболочку, как и у представителей многих других анемофильных семейств и родов.
Гинецей у видов триостренника состоит из 6 расположенных мутовчато плодолистиков, которые обычно полностью срастаются друг с другом своей брюшной (вентральной) частью. Лишь у триостренника заостренного (Т. mucronatum) верхушки плодолистиков остаются свободными и переходят в острие. Существенное различие имеется в числе развитых плодолистиков: у широко распространенного триостренника морского (Т. maritimum) все 6 плодолистиков развиты и несут по одному анатропному семязачатку, а у остальных видов этого рода 3 вполне развитых плодолистика чередуются с 3 стерильными и более или менее редуцированными. У представителей близкого к триостреннику рода цикногетон все 6 плодолистиков одинаково развитые и до основания свободные. У обоих родов плодолистики в верхней части более или менее оттянуты и заканчиваются длиннососочковыми рыльцами, сливающимися у видов с полностью сросшимися плодолистиками в одно дисковидное рыльце.
У очень обособленного и даже выделявшегося некоторыми авторами в особое семейство австралийского рода маундия число сегментов околоцветника варьирует от 2 до 4, вследствие чего цветки становятся более или менее зигоморфными. Тычинок у маундии обычно 6, с сидячими пыльниками. Этот род отличается от триостренника и по положению семязачатка в плодолистике: он здесь висячий и ортотропный.
Тетронциум магелланский (Tetroncium magellanicum, рис. 12, 7-10), в отличие от других представителей семейства, имеет не только однополые, но и двудомные цветки. Как соцветия, так и цветки мужских особей очень сходны по строению с соцветиями и цветками рдестовых (ср. рис. 13 и 14). Они также имеют 4 сегмента околоцветника и 4 тычинки с сидячими пыльниками, прикрепленными к основанию сегментов. Женские цветки в соцветиях женских особей обращены вниз и состоят из 4 сегментов околоцветника и 4 рогообразных, близ верхушки свободных плодолистиков.
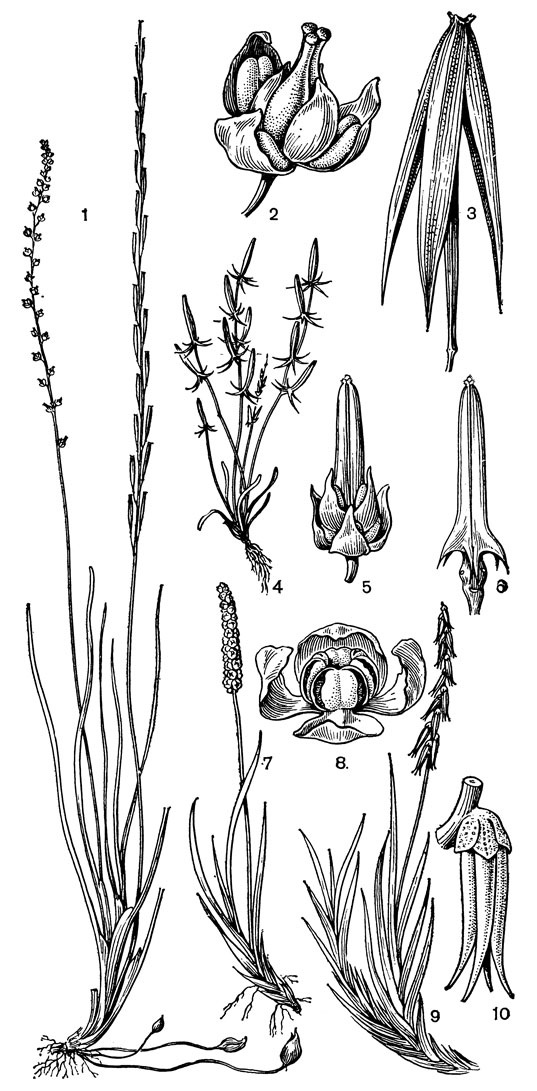
Рис. 12. Ситниковидные. Триостренник болотный (Triglochin palustre) 1 - общий вид; 2 - цветок; 3 - зрелый плод. Триострен ник капканный (Т. calcitrapa): 4 - общий вид; 5 - цветок; 6 - незрелый плод. Тетронциум магелланский (Tetroncium magellanicum): 7 - цветущий побег мужского растения; 8 - мужской цветок; 9 - цветущий побег женского растения; 10 - женский цветок
Еще оригинальнее цветки представителей американского рода лилеи, выделявшегося многими авторами в самостоятельное семейство. Их колосовидные соцветия могут быть двух типов: одни - на довольно длинных цветоносах, выступающих из листовых влагалищ, несут только женские цветки без околоцветника, состоящие из одного плодолистика с одним семязачатком и относительно коротким столбиком; другие - отчасти скрытые во влагалищах листьев и состоящие из обоеполых цветков, кроме одного или немногих конечных мужских цветков. Каждый обоеполый цветок имеет один прицветникообразный сегмент околоцветника, который срастается с основанием единственной тычинки, и один плодолистик. Кроме того, у основания цветоносов, во влагалищах листьев обычно имеются еще 1-2 женских цветка, которые кажутся сидячими, но в действительности заканчивают собой самостоятельные, сильно укороченные пазушные ветви. Эти цветки лишены околоцветника, а их одноплодолистиковый гинецей имеет необычайно длинный, иногда до 14 см, прямой нитевидный столбик с булавовидным рыльцем. Такой длинный столбик, по-видимому, необходим для того, чтобы рыльце всегда возвышалось над поверхностью воды, так как лилея растет на болотах и в мелких водоемах.
Плод ситниковидных обычно состоит из свободных или сросшихся друг с другом, невскрывающихся, односемянных частей, число которых соответствует числу фертильных плодолистиков. У триостренника морского плод распадается на 6 частей, а у других видов этого рода плод, начиная снизу, распадается на 3 части, которые некоторое время остаются соединенными с центральным столбиком плода, образованным тремя стерильными плодолистиками. У наиболее широко распространенного вида триостренника болотного (Triglochin palustre, рис. 12, 3) 3 части зрелого плода заканчиваются внизу длинным острием - отсюда как русское, так и латинское название этого рода. У лилеи плод состоит только из одной односемянной части. Семена всех ситниковидных лишены эндосперма и имеют прямой зародыш.
Все ситниковидные - влаголюбивые растения, обычно растущие на болотах и болотистых лугах, по берегам водоемов. Мелкие однолетние виды триостренника нередко обильно разрастаются на приречных отмелях и на месте пересохших водоемов. Уже маундия и лилея могут расти на дне мелководных водоемов, а австралийский цикногетон высокий (Cycnogeton procerus) - настоящее водное растение, обитающее в довольно глубоких водоемах, хотя его соцветия всегда возвышаются над водой. Высота его обычной водной формы может достигать 1 м и более; однако на месте пересохших водоемов встречается и наземная форма этого вида, значительно меньших размеров и с более узкими листьями. Многие виды триостренника, в том числе триостренник морской, галофильны и встречаются как на внутриконтинентальных солончаках, так и на засоленных местах морских побережий. К галофильным растениям принадлежит и лилея, часто растущая по берегам соленых озер.
Цветки ситниковидных опыляются с помощью ветра, причем самоопылению обычно препятствует протогиния, а у тетронциума - двудомность цветков. У видов триостренника отмечается скена не только женской и мужской фаз в развитии цветка, но и двух мужских фаз: в первой из них вскрываются тычинки наружного круга, а во второй - внутреннего круга. Несмотря на отсутствие нектарников, цветки триостренника посещают мелкие насекомые (вероятно, ради пыльцы), которые могут принимать участие в их опылении.
Основным способом распространения диаспор - односемянных частей плода - у ситниковидных является гидрохория. У всех видов семейства части плодов обладают хорошей плавучестью и способны переноситься потоками воды на большие расстояния. Однако у многих видов существенное значение имеет и экзозоохория. Части плодов триостренника морского и других видов этого рода могут переноситься с комочками почвы на ногах животных и человека. У некоторых видов есть специальные приспособления для такого распространения. Так, у триостренника болотного обращенные в сторону и вниз длиннозаостренные части плода запутываются в шерсти зверей и перьях птиц при соприкосновении с ними, а у триостренника капканного на каждой части плода есть еще шипообразный вырост (рис. 12, 6).
Многолетним ситниковидным свойственно также вегетативное размножение с помощью корневищ и луковичек. У тетронциума ветви корневища теряют связь с материнским корневищем. Триостренник болотный осенью образует в пазухах нижних листьев тонкие горизонтальные подземные побеги до 10 (15) см длиной, несущие на верхушке луковицеобразную зимующую почку, состоящую из немногих видоизмененных листьев. Материнское растение за зиму обычно отмирает, а из ставших самостоятельными луковичек образуются новые особи. Близкий вид - триостренник луковичный (Triglochin bulbosum) также образует луковички, но не на длинных столонах, а у основания материнского растения.
Водный цикногетон высокий образует довольно крупные клубни, употребляемые в пищу местным населением. Триостренник морской - хорошее пастбищное кормовое растение; кроме того, его вегетативные побеги можно использовать в пищу в качестве салата, но только после ошпаривания кипятком, уничтожающим не очень приятный запах, свойственный всем видам триостренника.
Семейство рдестовые (Potamogetonaceae)
К семейству рдестовых, включающему 2 рода, принадлежат многие обычные по всей территории СССР водные растения разнообразного облика с возвышающимися над поверхностью воды, реже плавающими колосовидными соцветиями. Содержащий около 100 видов род рдест (Potamogeton, рис. 13 и 14) распространен в пресных, реже солоноватых водоемах обоих полушарий, но преимущественно вне тропиков, а нередко присоединяемый к нему род гренландия (Groenlandia) представлен всего одним видом - гренландией густолистной (G. densa, рис. 13, 6-9), известной лишь из относительно немногих местонахождений в Евразии и Северной Африке.
Рдестовые начинают свое развитие, закрепляясь на дне водоемов с помощью более или менее длинных симподиальных корневищ, от узлов которых отходят простые, реже разветвленные корни с длинными корневыми волосками. Обычно они остаются прикрепленными к грунту, но отдельные побеги и части побегов могут отделяться от родительской особи и свободно плавать в воде, продолжая свое развитие. Рдестовые - многолетние растения. В оптимальных условиях, например в тропиках и субтропиках, они вегетируют в течение всего года, а во внетропических областях, где водоемы нередко замерзают и даже промерзают до дна, они перезимовывают с помощью корневищ и нижних частей побегов или за счет образующихся в пазухах листьев и на верхушках стеблей зимующих почек - сильно укороченных вегетативных побегов с тесно сближенными мелкими листьями и нередко утолщенной осью.
В отличие от преимущественно тропического семейства апоногетоновых, представители которого внешне похожи на рдестовых, последние всегда имеют удлиненные, часто сильно разветвленные стебли с расставленными, а не сближенными в прикорневую розетку листьями. У рдестов почти все листья расположены очередно за исключением двух почти супротивных листьев в местах отхождения соцветий, а у Гренландии все листья попарно (иногда по 3) сближены, располагаясь почти супротивно. Стебли рдестовых обычно цилиндрические, однако у рдеста сплюснутого (Potamogeton compressus) и родственных ему видов они сильно сплюснутые с боков и лентовидно расширенные, почти не отличающиеся по ширине от его линейных листьев.
По строению расположенных у основания листьев прилистниковидных придатков, обычно принимаемых за прилистники, все рдестовые можно разделить на три очень неодинаковые по объему группы. К первой принадлежит значительное большинство рдестов, составляющих подрод рдест (Potamogeton); у них прилистники срастаются друг с другом, образуя охватывающий стебель футляр, расположенный в пазухе листа и расщепленный с противоположной ему стороны. У видов этой группы листья не имеют влагалищ. Вторую группу образуют относительно немногочисленные виды рдестов из подрода колеогетон (Coleogeton), в том числе широко распространенный в СССР рдест гребенчатый (Potamogeton pectinatus). У этих видов прилистники почти по всей своей длине прирастают к нижней части листьев, образуя довольно длинные трубкообразные влагалища, которые могут быть расщепленными со стороны, противоположной листу (у рдеста гребенчатого), или замкнутыми (у рдеста нитевидного - P. filiformis). Существуют немногие переходные между обеими группами виды с короткими влагалищами листьев, к ним принадлежат дальневосточный рдест Маака (Р. maackianus) и североамериканский рдест Роббинса (P. robbinsii), у которого верхние свободные части прилистников выступают над основанием листовой пластинки в виде двух длинных и острых зубцов. К третьей группе принадлежит только род Гренландия, у представителей которого два почти свободных прилистника имеют лишь листья, расположенные у выхода ножек соцветия, а остальные листья вообще лишены прилистников. Кроме прилистников, в пазухах листьев большинства рдестов расположены еще очень мелкие внутривлагалищные чешуйки с выделяющими слизь железками.
Форма листьев у рдестовых очень изменчива. У большинства видов сидячие листья, от узколинейных, почти нитевидных (у рдеста волосовидного - P. trichoides, или рдеста нитевидного) до штирокоэллиптических (у рдеста блестящего - P. lucens, рис. 13, 1) или широкояйцевидных со стеблеобъемлющим основанием (у рдеста пронзеннолистного - P. perfoliatus, рис. 13, 3). Длинные черешки имеются у листьев с плавающими на поверхности воды эллиптическими или ланцетными пластинками. Наиболее широко распространенный вид с такими листьями - рдест плавающий (P. natans, рис. 14, 2), у которого многочисленные плавающие листья и относительно немногие подводные листья с очень узкими, иногда почти полностью редуцированными пластинками. Более широкие подводные листья при также всегда имеющихся плавающих листьях у близкого вида рдеста узловатого (P. nodosus); у рдеста злаковидного (P. gramineus, рис. 14, 7) листья уже преимущественно подводные, ланцетные, с очень короткими черешками, а плавающие листья с эллиптическими пластинками и длинными черешками часто вообще отсутствуют. У рдеста альпийского (P. alpinus, рис. 14, 9) плавающие короткочерешковые листья уже мало отличаются от подводных и также могут отсутствовать. Многие рдестовые имеют цельнокрайние листья, но не так уж редки и виды с зубчатыми по краю листьями, а у широко распространенного в СССР рдеста курчавого (P. crispus) листья по краю обычно курчавоволнистые. Жилкование листьев у рдестовых дуговидное или параллельное, причем у узколистных видов с линейными пластинками количество жилок имеет большое систематическое значение.
Колосовидные соцветия рдестовых расположены на безлистных ножках, выходящих из верхушек побегов или из пазухи одного из двух почти супротивно сближенных листьев. У значительного большинства видов соцветия во время цветения возвышаются над поверхностью воды, поддерживаемые или розеткой плавающих листьев, или утолщенными за счет воздухоносной ткани ножками. При плодах соцветия обычно вновь погружаются в воду, а у рдестов из подрода колеогетон соцветия, часто имеющие очень расставленные мутовки цветков, и во время цветения лежат на поверхности воды. У Гренландии соцветия образованы лишь немногими, часто только двумя цветками (рис. 13, 7), в отличие от обычно многоцветковых соцветий рдестов.
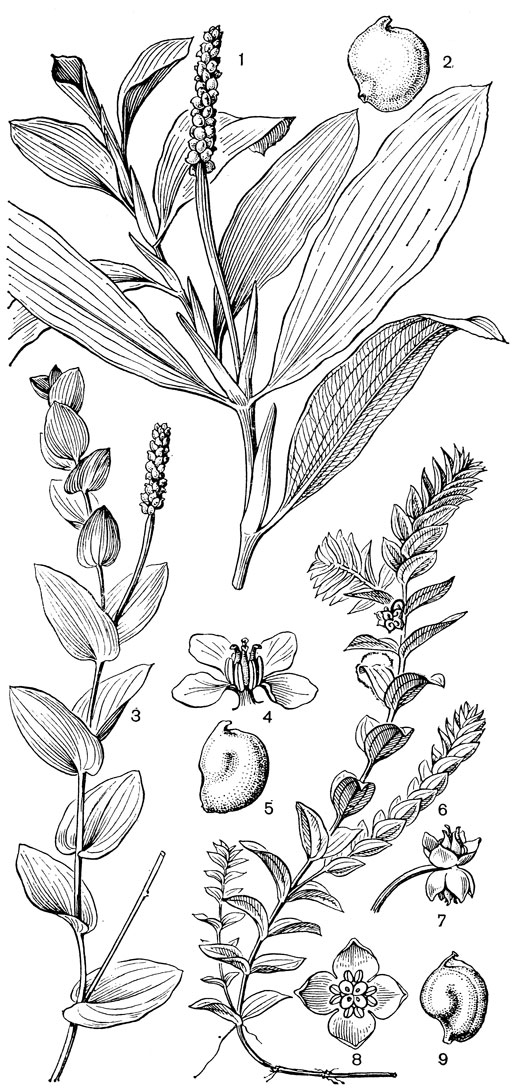
Рис. 13. Рдестовые. Рдест блестящий (Potamogeton lucens): 1 - общий вид; 2 - плодик. Рдест пронзеннолистный (Р. perfoliatus): 3 - общий вид; 4 - цветок; 5 - плодик. Гренландия густолистная (Groenlandia densa): 6 - общий вид плодоносящего побега; 7 - соцветие из двух цветков; 8 - цветок; 9 - плодик
Цветки всех рдестовых очень сходны по строению. Они не имеют прицветников, всегда обоеполые, актиноморфные и четырехмерные. Их интересной особенностью является необычное расположение тычинок: сидячие или почти сидячие пыльники прикреплены здесь к ноготкам сегментов околоцветника, составляя с ними одно целое и располагаясь непосредственно над ними (рис. 13 и 14). Прежде многие авторы принимали сегменты околоцветника в цветке рдестов за выросты сильно разросшихся связников пыльников, однако в настоящее время эта точка зрения оставлена, так как было доказано, что тычинки и сегменты околоцветника закладываются разными зачатками и срастаются лишь в ходе дальнейшего развития. Пыльцевые зерна шаровидной формы. Гинецей состоит из 4, редко 2-5 свободных плодолистиков, обычно чередующихся с тычинками. Лишь у рдеста волосовидного в цветке развит только 1 плодолистик, а 3 остальных редуцированы. Каждый развитый плодолистик имеет 1 семязачаток и оттянут на верхушке в более или менее длинный носик, на котором, обычно вдоль его вентральной борозды, располагается покрытое сосочками рыльце. Плод состоит из 1-4 костянковидных, реже (у Гренландии) орешковидных односемянных частей - плодиков, детали строения которых имеют большое систематическое значение. У некоторых видов, например у дальневосточного рдеста гребнеплодного (P. cristatus), плоды имеют шипообразные выросты вдоль киля.
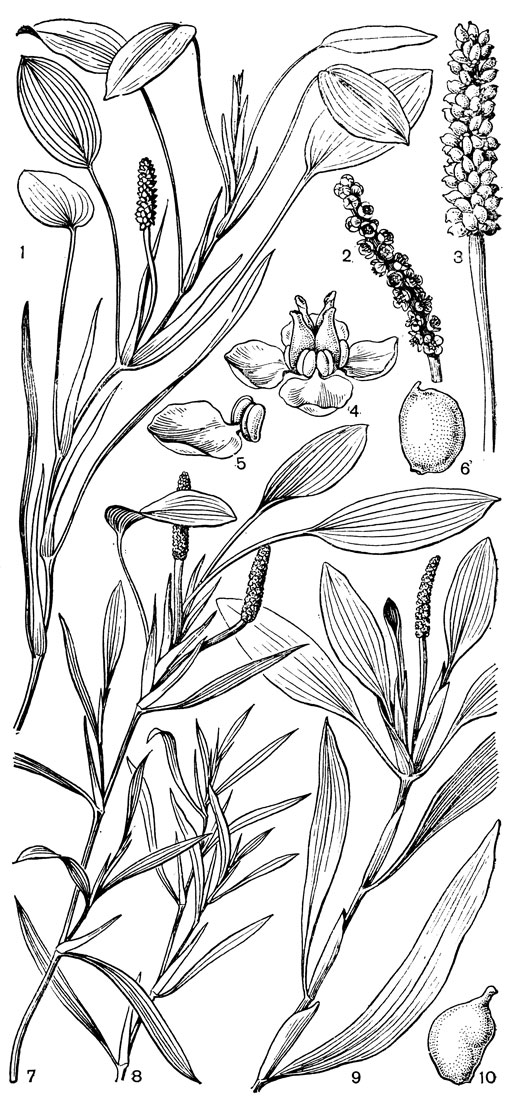
Рис. 14. Рдестовые. Рдест плавающий (Potamogeton natans): 1 - общий вид; 2 - соцветие; 3 - соцветие при плодах; 4 - цветок; 5 - листочек околоцветника с противолежащей ему тычинкой; 6 - плодик. Рдест злаковидный (P. gramineus): 7 - общий вид побега с плавающими листьями и соцветием; 8 - побег без плавающих листьев. Рдест альпийский (P. alpinus): 9 - общий вид; 10 - плодик
Все рдестовые - пресноводные растения, и лишь немногие из них (например, рдест гребенчатый) способны развиваться в приморских лагунах и солоноватых озерах. Некоторые широко распространенные виды могут расти на разных глубинах как в стоячих, так и в быстра текущих водоемах; другие более требовательны к условиям обитания. Так, относительно редко встречающийся в СССР рдест длиннейший (P. praelongus), как правило, растет в довольно больших и глубоких озерах. Рдест узловатый обычен для рек с относительно большой скоростью течения, а Гренландия густая - для ручьев и речек с довольно быстрым течением и чистой водой. При пересыхании водоемов многие виды рдестов погибают, но рдесты злаковидный и альпийский могут некоторое время существовать на бывшем дне водоема, образуя своеобразную карликовую наземную форму. Рдест злаковидный очень часто цветет и плодоносит, не образуя плавающих листьев, причем наличие или отсутствие плавающих листьев у него далеко не всегда определяется глубиной водоема. За счет образования отчленяющихся боковых побегов и зимующих почек рдесты способны быстро размножаться, нередко полностью заполняя небольшие водоемы. Это особенно относится к таким наиболее обычным в СССР рдестам, как плавающий, пронзеннолистный, гребенчатый, рдест Берхтолъда (P. berchtoldii), злаковидный, блестящий и др. Некоторые виды рдестов поднимаются в горы до 3000 м и выше.
Опыление цветков у большинства рдестов осуществляется с помощью ветра. В безветренные дни часть пыльцы из вскрывшихся пыльников может высыпаться на вогнутую поверхность сегментов околоцветника и позднее разноситься ветром уже отсюда. Обычно рдесты растут большими колониями, что существенно облегчает опыление. У большинства видов самоопылению препятствует более или менее выраженная протогиния - более раннее созревание плодолистиков, причем у некоторых видов, например у рдеста курчавого, две фазы цветения: женская и мужская отчетливо отделены друг от друга по времени. Имеются наблюдения, что цветки некоторых рдестов, находясь под водой, способны самоопыляться клейстогамно. У видов подрода колеогетон, по-видимому, и у некоторых других опыление постоянно происходит на поверхности воды, куда высыпается пыльца. Длинные и гибкие соцветия рдестов гребенчатого и нитевидного никогда не возвышаются над водой, а плавают на ее поверхности, где и опыляются. Опылителями рдестов могут быть также улитки и некоторые насекомые, поедающие пыльцу. Виды рдестов, опыляющихся на поверхности воды, имеют более длинные сосочки на рыльцах, чем анемофильные виды.
Плодики большинства рдестовых плавают очень короткое время или даже совсем не могут плавать. Лишь у рдеста плавающего и близких к нему видов плодики долго не тонут и могут распространяться водой на большие расстояния. Важную роль в распространении плодиков рдестов играют также рыбы и водоплавающие птицы, поедающие их ради мясистого околоплодника. Орешкообразные плодики Гренландии, по-видимому, хорошо плавают и распространяются только при помощи водных потоков.
Отметим, что среди рдестов немало плохо плодоносящих или даже совсем не плодоносящих видов. Некоторые, например рдест узловатый, не плодоносят в северной части своего ареала. Поэтому вегетативные способы размножения рдестов, о которых уже говорилось выше, играют не меньшую, а, наверное, даже большую роль, чем размножение с помощью репродуктивных органов.
Рдестовые имеют некоторое значение в рыбном хозяйстве, так как в их зарослях рыбы мечут икру и мальки находят себе защиту. Однако большие заросли рдестов во многих случаях препятствуют движению мелких судов и лодок. При очистке водоемов рдестовые можно использовать как ценное удобрение для полей. Богатые крахмалом клубенькообразные утолщения корневищ рдеста гребенчатого и некоторых других видов можно употреблять в пищу.
Семейство руппиевые (Ruppiaceae)
К семейству руппиевых принадлежит всего один род руппия (Ruppia, рис. 15), 8-10 видов которого широко распространены во внетропических областях обоих полушарий, а отчасти также в горных районах тропиков. Руппии - обычно растущие большими колониями водные растения, напоминающие по облику узколистные рдесты (Potamogeton). Однако, в отличие от последних, они обитают только в солоноватых водоемах разной степени солености. Там, где такие водоемы отсутствуют (например, в большей части таежной зоны Евразии, исключая морское побережье), там нет и руппий. Встречаются они и высоко в горах (например, в Андах на высоте до 4000 м над уровнем моря), если там есть соленые озера.
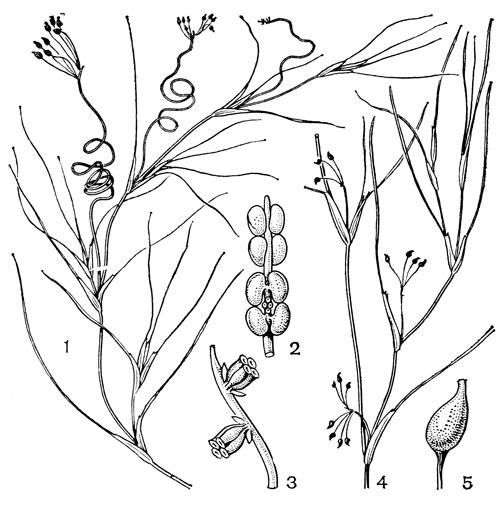
Рис. 15. Виды руппии (Ruppia). Руппия усиконосная (R. cirrhosa): 1 - общий вид; 2 - соцветие с двумя цветками в мужской фазе цветения; 3 - оно же в женской фазе цветения. Руппия морская (R. maritima): 4 - общий вид; 5 - плодик
Руппии - многолетние растения, полностью или почти полностью погруженные в воду, с довольно длинными и тонкими ползучими корневищами, от узлов которых отходят неразветвленные корни с очень длинными корневыми волосками. Однако некоторые виды руппии могут расти, цвести и плодоносить также в свободно плавающем состоянии, при соприкосновении с дном водоемов вновь укрепляясь на нем с помощью придаточных корней.
Листья руппии узколинейные, иногда нитевидные, сидячие, обычно лишь с одной слабо заметной жилкой. Приросшие к их нижней части прилистники образуют охватывающее стебель, но незамкнутое влагалище. На стеблях листья обычно располагаются очередно, и только на верхушке имеются одна или две пары супротивных или почти супротивных листьев с немного более вздутыми влагалищами.
Соцветие сначала заключено в расширенное влагалище одного из двух верхних листьев побега. Затем ножка соцветия быстро удлиняется, причем у некоторых видов, например у руппии усиконосной (R. cirrhosa), она достигает очень большой длины и скручивается в виде спирали (рис. 15, 1). Каждое соцветие состоит из 2 сближенных обоеполых цветков, не имеющих прицветников и околоцветника, если не считать двух очень мелких чешуевидных выростов, расположенных у основания тычинок и часто принимаемых за рудименты околоцветника. У руппии очень хорошо выражена протандрия, препятствующая самоопылению, и цветки каждого соцветия проходят как бы две фазы развития: сначала мужскую, потом женскую. В мужской фазе цветок имеет 2 супротивных, почти сидячих пыльника, каждый с 2 крупными почковидными гнездами, разделенными довольно широким связником. Расположенный между пыльниками гинецей из 2-10 свободных плодолистиков в это время еще недоразвит. После вскрытия и освобождения от пыльцы пыльники опадают, но разрастаются и становятся способными к восприятию пыльцы плодолистики с расположенными в верхней части их вентральной (брюшной) стороны сидячими щитовидными рыльцами. Кроме того, во время женской фазы цветения быстро начинают удлиняться основания плодолистиков. В результате этого плодолистики каждого цветка оказываются расположенными на длинных и тонких ножках, выходящих из одного места и производящих впечатление лучей зонтиковидного соцветия. Наблюдая такие "зонтики" при плодах (рис. 15, 1), трудно поверить в то, что каждый отдельный плодик формируется не из целого цветка, а лишь из его части. Невскрывающиеся односемянные плодики руппии обычно имеют обратногрушевидную форму, но более или менее асимметричны. Они принадлежат к числу костянкообразных плодов, так как наружная оболочка у них мясистая и богатая крахмалом, а внутренняя очень твердая.
Отдельные виды руппии заметно отличаются по своей экологии. Наименее галофильный из них - руппия коротконожковая (R. brachypus) растет в дельтах рек и приморских лагунах с очень слабой соленостью воды. К водоемам с большим содержанием солей приурочена широко распространенная руппия морская (R. maritima). Она отсутствует, например, в опресненной Невой части Финского залива (Ленинградская область), но встречается в изобилии в водоемах, образованных солеными источниками, близ курорта Старая Русса (Новгородская область). На юге СССР эта руппия - обычное растение соленых озер, хотя в горько-соленых озерах ее замещает более редкая руппия трапанинская (R. drepanensis). В морских заливах и приморских лагунах с еще более соленой водой встречаются виды с очень длинными, спирально согнутыми ножками соцветий: широко распространенная руппия усиконосная и северотихоокеанская руппия западная (R. occidentalism Кроме того, эти виды имеют более толстые корневища и могут обитать на больших глубинах.
Хотя для руппии отмечалась возможность опыления ветром ее цветков в случаях, когда они возвышаются над поверхностью воды, в настоящее время для всех видов этого рода установлена гидрофилия. Попадая в воду после вскрытия пыльников, эллипсоидальные и немного согнутые, часто почти почковидные пыльцевые зерна свободно плавают в воде, постепенно поднимаясь к ее поверхности, и приходят в соприкосновение с рыльцами цветков, находящихся в женской фазе. Если у менее галофильной руппии морской опыление обычно осуществляется в воде и лишь отчасти на ее поверхности, то у руппии усиконосной и других видов, обитающих в более соленой (и потому более тяжелой) воде, пыльцевые зерна быстро поднимаются на ее поверхность, где в основном и происходит опыление. Расположению соцветий этих видов руппии во время женской фазы цветков на поверхности воды помогают длинные ножки соцветий, спиральные изгибы которых позволяют реагировать на изменения глубины водоемов во время приливов и отливов или во время волнения моря. Кроме того, эти спиралеобразные ножки могут вращать соцветия на поверхности воды, вследствие чего увеличивается возможность контакта их с плавающей пыльцой.
Плодики руппии, вероятно, распространяют рыбы и птицы, использующие в пищу их мясистую оболочку, в то время как твердая косточ ка проходит через пищевод животных без по вреждений (эндозоохория). Руппия легко размножается также вегетативно, с помощью частей корневищ и плавающих побегов, способных укореняться. Зрелые плодики обычно долго сохраняются на материнских побегах и могут вместе с ними перемещаться ветром и морскими тесениями на большие расстояния.
Семейство дзанникеллиевые (Zannichelliaceae)
Дзанникеллиевые - относительно небольшое семейство, содержащее всего 4 рода и около 15 видов высокоспециализированных водных растений. Его представители широко, но спорадично распространены в пресных и солоноватых водоемах обоих полушарий, исключая Арктику и значительную часть таежной Евразии и Северной Америки. Почти по всему ареалу семейства (за исключением Австралии), в том числе и на территории СССР, распространен его наиболее крупный род - дзанникеллия (Zannichellia, рис. 16) с 9-10 видами. Род лепилена (Lepilaena) с 4 видами встречается только в Австралии и Новой Зеландии, а монотипный род псевдалътения (Pseudalthenia) - в Южной Африке. Ареал другого монотипного рода - алътении (Althenia) - еще недавно считался ограниченным Западным Средиземноморьем. Лишь в 1975 г. неожиданно выяснилось, что единственный вид этого рода - алътения нитевидная (A. filiformis, рис. 16, 5) - не только встречается в СССР, но и имеет здесь значительную часть своего ареала: от Ростовской области (Манычские озера) до предгорий Алтая. Одновременно этот вид был обнаружен также в Турции.
Такое "расширение" ареала альтении в самое последнее время объясняется невзрачным обликом этого растения, как, впрочем, и всех других дзанникеллиевых. Даже опытные коллекторы нередко принимают их за проростки или вегетативные побеги других водных растений, чаще всего узколистных видов рдеста (Potamogeton).
Дзанникеллиевые - полностью погруженные в воду растения и обычно прикреплены ко дну водоемов с помощью тонких ползучих корневищ, но могут продолжать свое развитие и в свободно плавающем состоянии. Большинство видов - многолетники с длительно существующими или распадающимися на зиму стеблями. Однако в пересыхающих водоемах они могут вести себя как однолетники, образуя большое количество плодов. Капский эндемик псевдалътения Ашерсона (Pseudalthenia aschersonianа) обитает только в пересыхающих водоемах и ежегодно отмирает, являясь уже настоящим однолетником. Корневища дзанникеллии почти не отличаются по внешнему виду от часто также стелющихся по дну стеблей, но у представителей других родов они более обособлены и имеют в узлах опадающие чешуевидные листья.
Нередко сильно разветвленные, тонкие и гибкие стебли дзанникеллиевых несут узколинейные, часто нитевидные сидячие листья с одной не всегда заметной жилкой. Они могут располагаться очередно, супротивно или в ложных мутовках по 3, а на верхушках побегов иногда сближаются пучками. У дзанникеллии листья у основания имеют свободные, охватывающие стебель прилистники, у других родов прилистники срастаются с нижней частью листьев, образуя влагалища. У альтении влагалища в значительной части перепончатые и заканчивающиеся на верхушке язычкоподобным выростом.
Как и у многих других высокоспециализированных водных растений, у дзанникеллиевых всегда однополые цветки сильно редуцированы. Цветки дзанникеллии (рис. 16) расположены в узлах стебля обычно по 2, но принадлежат к разным полам. Мужские цветки представлены всего одной, эфемерно существующей тычинкой с быстро удлиняющейся во время цветения нитью и пыльниками с (2) 4 (8) гнездами и выступающим над гнездами острым выростом связника - надсвязником. Расположенный рядом женский цветок состоит из (1)2-5(9) свободных плодолистиков, окруженных чашевидным или трубчатым покрывалом, которое обычно принимается за сросшиеся друг с другом листочки околоцветника. Казалось бы, расположенные рядом мужской и женский цветки можно принять за один обоеполый цветок, однако эти цветки заканчивают собой самостоятельные, хотя и сильно укороченные побеги: мужской цветок - пазушный боковой побег, а женский цветок - продолжение главного побега над узлом. В этом же узле симподиально образуется вегетативный побег, являющийся продолжением стебля. Каждый плодолистик содержит один ортотропный семязачаток и имеет на верхушке более или менее длинный столбик, заканчивающийся крупным воронковидным рыльцем.
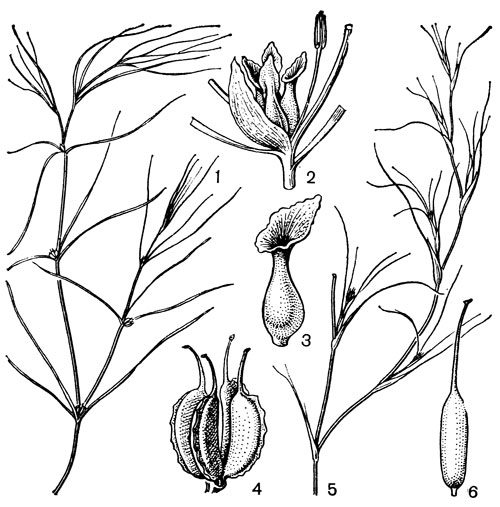
Рис. 16. Дзанникеллиевые. Дзанникеллия болотная (Zannichellia palustris): l - общий вид; 2 - часть растения с мужским и женским цветками; 3 - плодолистик с воронковидным рыльцем; 4 - плод. Альтения нитевидная (Althenia filiformis): 5 - общий вид; 6 - плодик
К дзанникеллии близка по строению цветков псевдальтения, у которой, однако, мужские цветки всегда имеют пыльники с 8 гнездами, а женские цветки состоят только из одного плодолистика, окруженного покрывалообразным околоцветником. У двух остальных родов - альтении (рис. 16, 5) и лепилены - цветки образуются на верхушках облиственных побегов. При этом мужские цветки имеют очень мелкий трехлопастный околоцветник, расположенный непосредственно под пыльником, а обычно трехплодолистиковый гинецей окружен околоцветником из 3 свободных сегментов, расположенных против плодолистиков. У альтении пыльники одногнездные, а рыльце в виде небольшой щитковидной воронки, на верхушке длинного и тонкого столбика. У лепилены пыльники обычно с 4-12 гнездами, а рыльце булавовидное. У очень обособленной новозеландской лепилены двугнездной (Lepilaena bilocularis) пыльники с 2 гнездами, а рыльце языковидное, по краю перистонадрезанное.
Плоды дзанникеллиевых состоят из нескольких, реже одного, односемянных невскрывающихся костянковидных плодиков, основание которых разрастается, образуя более или менее длинную, иногда равную по длине расширенной части плода ножку. У дзанникеллии плодики продольно килеватые, немного дуговидно согнутые; у других родов обычно прямые, эллипсоидальные.
Значительная редукция цветков у дзанникеллиевых допускает различные гипотезы, объясняющие их строение. Единственный пыльник мужского цветка дзанникеллии и лепилены нередко принимается за результат срастания 2 или 3 сидячих пыльников, а тычиночная нить - за ножку соцветия, в пользу чего свидетельствует расположение околоцветника не у основания нити, а у основания пыльника у мужских цветков альтении и лепилены. Некоторые авторы принимают плодолистики женских цветков за отдельные цветки, а расположенные супротивно им листочки околоцветника - за прицветники.
Виды дзанникеллиевых приспособлены к водоемам с разной степенью солености. Широко распространенная дзанникеллия болотная (Zannichelia palustris) встречается в пресных или едва солоноватых водоемах, а более галофильная дзанникеллия длинноножковая (Z. pedunculata) - в приморских лагунах и соленых озерах. Альтения нитевидная встречается только в горько-соленых озерах. При этом даже в пределах одного и того же вида наблюдается интересная корреляция между галофильностью и строением плодиков: чем солонее вода в водоеме, тем длиннее ножки плодиков и их столбики. По-видимому, эта особенность связана с какими-то особенностями опыления в соленой воде.
Шаровидные или почти шаровидные пыльцевые зерна дзанникеллиевых тяжелее воды и после вскрытия пыльников медленно опускаются на дно. При этом некоторые из них попадают на воронковидные рыльца женских цветков. Дзанникеллиевые обычно растут большими колониями и образуют множество цветков, так что возможности опыления здесь довольно велики. Самоопылению у однодомных родов - дзанникеллии и псевдальтении в какой-то степени препятствует более раннее развитие мужских цветков, а у родов альтении и лепилены - двудомность: мужские и женские цветки развиваются на разных особях.
Плодики дзанникеллиевых обычно падают на дно, но могут переноситься водными потоками, а также по поверхности воды вместе с побегами материнского растения, на которых зрелые плоды остаются довольно долго. По-видимому, они могут также распространяться поедающими их животными - рыбами и водоплавающими птицами.
Семейство цимодоцеевые (Cymodoceaceae)
Семейство цимодоцеевых, состоящее из 5 родов и около 15 видов, близко к дзанникеллиевым и нередко объединяется с ними. Однако, в отличие от дзанникеллиевых, обитающих в пресных или слабо солоноватых водоемах, цимодоцеевые встречаются только в водах морей и океанов и, наряду с взморниковыми, посидониевыми и некоторыми родами семейства водокрасовых, принадлежат к так называемым "морским травам", напоминающим по облику скорее водоросли, чем цветковые растения. Цимодоцеевые существенно отличаются от дзанникеллиевых и в географическом распространении: если дзанникеллиевые в основном внетропическое семейство, то цимодоцеевые, напротив, распространены преимущественно в тропиках. В северном полушарии лишь цимодоцея узловатая (Суmodocea nodosa, рис. 17) встречается за пределами тропиков: вдоль побережий Средиземного моря - от Пиренейского полуострова до Малой Азии, а вдоль берегов Атлантического океана - от Марокко и Канарских островов до Юго-Западной Франции; в южном полушарии амфиболис антарктический (Аmрhibolis antarctica) заходит в Южную Австралию и Тасманию. Тропики Старого Света заметно богаче цимодоцеевыми, чем неотропики, где встречаются лишь немногие виды этого семейства.
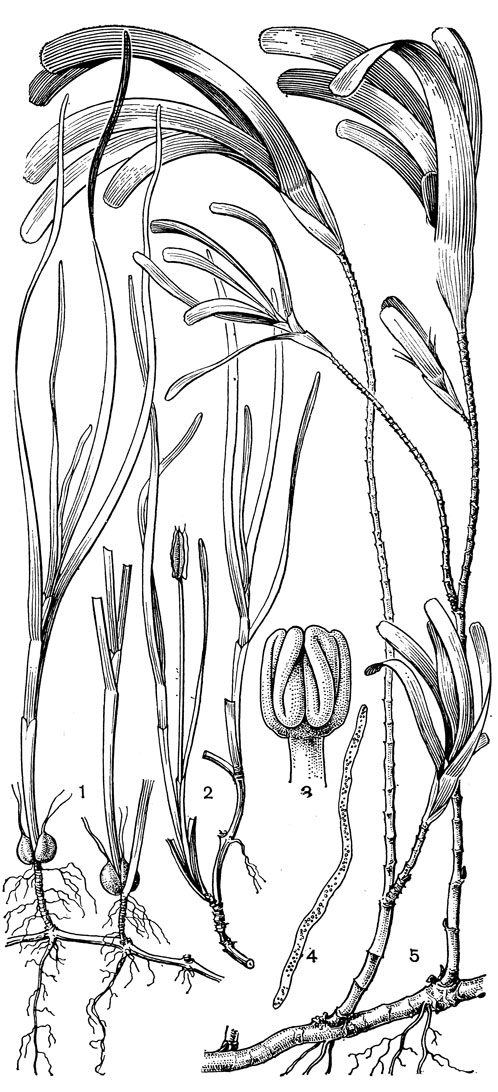
Рис. 17. Цимодоцеевые. Цимодоцея узловатая (Cymodocea nodosa): 1 - общий вид плодоносящего растения; 2 - общий вид растения с мужским цветком; 3 - сросшиеся пыльники; 4 - пыльцевое верно. Талассодендрон реснитчатый (Thalassodendron ciliatum): 5 - общий вид растения с женскими соцветиями
Как и другие "морские травы", цимодоцеевые - полностью погруженные в воду многолетние растения с хорошо развитыми ползучими корневищами, закрепляющимися в грунте морского дна с помощью придаточных корней. Очередные линейные листья обычно сближены на верхушках отходящих от узлов корневища прямостоячих побегов. Наиболее оригинальный представитель семейства - талассодендрон реснитчатый (Thalassodendron ciliatum), обитающий на коралловых рифах Индийского и Тихого океанов, имеет особенно прочные, более или менее деревянистые симподиальные корневища и пучки кожистых, двурядно расположенных листьев шириной около 1 см (рис. 17, 5). У видов других родов корневища травянистые, а листья более узкие (до щетиновидных). Строение листьев, особенно их верхушки, имеет большое значение при разграничении видов этого семейства, так как цветки и плоды встречаются у них очень редко. Листья цимодоцеевых отчетливо разделены на пластинку и свободное (не сросшееся своими краями) влагалище, в пазухе которого имеются внутривлагалищные чешуйки. У основания пластинок обычно присутствуют поперечный перепончатый вырост - язычок - и два более или менее развитых ушка. Жилкование листьев параллельное, не всегда хорошо заметное.
Цветки цимодоцеевых отличаются от цветков близкого семейства дзанникеллиевых еще более высокой специализацией. Цимодоцеевые - растения двудомные, мужские и женские цветки которых располагаются на разных особях. В отличие от дзанникеллиевых, образующих большое количество плодов, цимодоцеевые размножаются преимущественно вегетативно - частями корневищ - и цветут крайне редко. Цветки образуются по одному или в числе нескольких на сильно укороченных боковых веточках, обычно окруженных листовыми влагалищами, реже (у талассодендрона) 4 листоподобными прицветниками. Мужские цветки состоят из 2, реже 3 (у некоторых видов галодулы - Halodule) более или менее срастающихся друг с другом (у галодулы на разной высоте) пыльников, расположенных на длинной нити (быть может, ножке сильно редуцированного соцветия). У цимодоцеи узловатой (рис. 17, 3) 8-гнездные "двойные" пыльники достигают в длину около 15 мм, окрашены в красный цвет и расположены на нитях длиной 8-10 см. Сидячие или расположенные на очень короткой ножке в числе 1-2 женские цветки состоят из 2 плодолистиков, каждый из которых имеет 1 ортотропный семязачаток и заканчивается коротким столбиком, переходящим в очень длинные (до 10 см), цельные (у галодулы) или 2-4 раза вильчато разветвленные (у других родов) нитевидные рыльцевые ветви.
Плоды цимодоцеевых костянковидные, невскрывающиеся, образованные 1 плодолистиком или 2 сросшимися плодолистиками, но всегда односемянные. У талассодендрона они окружены мясистым внутренним прицветником, который, вероятно, служит пищей для рыб, способствующих таким образом распространению плодов.
Цимодоцеевые обычно образуют большие колонии, нередко вместе с другими "морскими травами", большей частью на песчаном или каменистом дне морских заливов и бухт на глубинах до 3 м и более. Талассодендрон дальше всех других "морских трав" отходит от побережья, заселяя поднимающиеся со дна коралловые рифы.
Биология большинства цимодоцеевых изучена еще недостаточно, так как они редко цветут. У цимодоцеи узловатой цветки образуются в слое ила или песка, из которого обычно выступают лишь расположенные на длинных нитях пыльники мужских цветков и длинные нитевидные рыльцевые ветви женских. Подобно взморниковым и посидониевым, у цимодоцеевых нитевидные пыльцевые зерна, больше похожие на пыльцевые трубочки, чем на "зерна". Плавающие в воде после вскрывания пыльников нитевидные пыльцевые зерна имеют значительно больше шансов зацепиться за рыльцевые ветви, чем пыльцевые зерна другой формы. У талассодендрона, а еще чаще у амфиболиса антарктического отмечается вивипария: женские цветки видоизменяются в молодые растения, позднее отделяющиеся от материнского побега.
У цимодоцеи узловатой зрелый плод довольно крупных размеров (длиной 8-12 мм и шириной 5-7 мм), обычно долго остается на материнском растении, погружаясь в песок и прорастая после разрушения его оболочек. Однако как плоды, так и проростки могут распространяться по воде на большие расстояния во время бурь и штормов. У талассодендрона и амфиболиса проростки вместе с оболочкой плода долго плавают в воде и могут закрепляться на мелководных морских участках, причем плоды амфиболиса имеют специальный гребневидный вырост, способствующий такому закреплению.
Колонии цимодоцеевых служат убежищем и местом икрометания для многих видов рыб, а выброшенные на берег листья и отдельные побеги могут использоваться как хорошее удобрение для полей.
Семейство взморниковые (Zosteraceae)
Небольшое семейство взморниковых (Zosteraceae), представителей которого часто называют "морской травой", принадлежит к очень интересной в биологическом отношении группе цветковых растений, приспособившихся к обитанию в соленой воде морей и океанов. Очень длинные линейные листья взморниковых, полностью погруженные в воду, нетрудно принять за морские водоросли, а их невзрачные цветки, как и плоды, редко удается видеть.
Почти все взморниковые имеют длинные горизонтальные корневища, закрепленные в морском дне с помощью довольно толстых неразветвленных корней и несущие укороченные вегетативные побеги с линейными листьями шириной 0,3-15 мм и длиной до 1,5 м. Корневища обычно моноподиальные, неодревесневающие, за исключением взморника тасманского (Zostera tasmanica), имеющего симподиальные одревесневающие корневища и на этом основании выделенного в последнее время в особый род гетерозостера (Heterozostera). Репродуктивные побеги, у многих взморниковых образующиеся не каждый год, могут быть довольно длинными и разветвленными, с несколькими, иногда многочисленными початковидными соцветиями, называемыми веером или опахалом. У большинства видов рода филлоспадикс (Phyllospadix, рис. 18, 9-12) репродуктивные побеги очень короткие, боковые по отношению к вегетативным побегам и несущие только один веер. Каждый веер состоит из сильно сплюснутой и немного утолщенной оси, на одной стороне которой у однодомного взморника (Zostera, рис. 18, 2) располагаются, чередуясь, мужские и женские цветки, а у двудомного филлоспадикса - только мужские или только женские цветки. У филлоспадикса такие веера выступают из влагалищ кроющих листьев - покрывал и несут по бокам довольно крупные зеленые листочки, называемые ретинакулами. У видов взморника каждый веер заключен во влагалища своего покрывала, причем у видов подрода зостерелла (Zosterella) влагалища открытые и соцветия имеют очень мелкие перепончатые ретинакулы, располагающиеся возле мужских цветков, а у видов подрода зостера (Zostera) влагалища замкнутые (но позднее расщепляющиеся) и соцветия лишены ретинакул. Цветки взморниковых сильно упрощены: мужской состоит из одного сидячего пыльника с гребневидным связником, а женский представлен, по-видимому, двуплодолистиковым гинецеем, состоящим из 1-гнездной завязи с 1 висячим ортотропным семязачатком и короткого столбика с 2 рыльцами. Происхождение ретинакул не вполне ясно: их принимают или за рудимент околоцветника, или за прицветники, или за выросты оси соцветий.
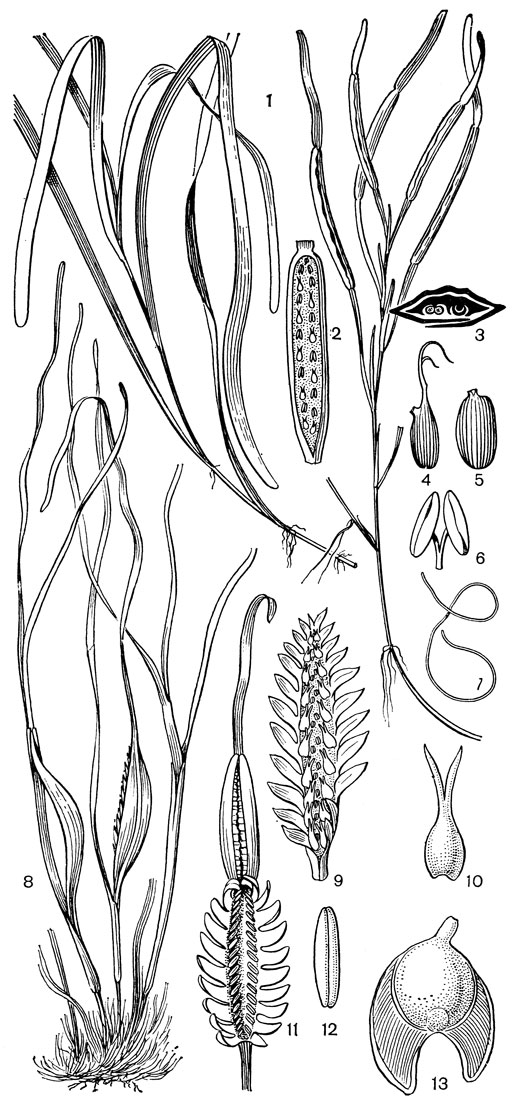
Рис. 18. Взморниковые. Взморник морской (Zostera marina): 1 - общий вид; 2 - соцветие с мужскими и женскими цветками; 3 - оно же на поперечном разрезе (видны сильно сплюснутая ось соцветия, заключенная в листовое влагалище, мужской и женский цветки); 4 - плод; 5 - семя; 6 - мужской цветок; 7 - пыльцевое зерно. Филлоспадикс Скулера (Phyllospadix scouleri): 8 - общий вид; 9 - соцветие с женскими цветками и рудиментами мужских цветков; 10 - женский цветок; 11 - соцветие с мужскими цветками; 12 - мужской цветок; 13 - плод
Пыльцевые зерна лишены экзины и имеют своеобразную нитевидную форму. Плод у видов взморника эллипсоидальный или яйцевидный, с перепончатым околоплодником, у видов филлоспадикса - серповидный или стреловидный с околоплодником, дифференцированным на мягкий экзокарпий и жесткий эндокарпий. После разрушения экзокарпия эндокарпий плода филлоспадикса образует с внутренней стороны боковых выростов как бы щеточки из волокон, которые играют существенную роль в "заякоривании" плодов среди водорослей на камнях и скалах. Семена обоих родов имеют эллипсоидальную форму; эндосперм в них отсутствует.
Семейство взморниковых состоит из 3 родов с 23 видами. Из них наиболее крупный род - взморник с 15 видами делится на 2 очень обособленных подрода: зостера, широко распространенный во внетропической части северного полушария, лишь немного заходящий в Арктику, и зостерелла, распространенный по морским побережьям обоих полушарий, но вне Арктики и преимущественно вне тропиков, хотя некоторые его виды заходят в тропические области Юго-Восточной Азии, Африки и Австралии. Монотипный род гетерозостера обитает главным образом в Австралии (включая Тасманию), но встречается также на небольшом участке южноамериканского побережья в пределах Чили. Ареал рода филлоспадикс (Phyllospadix) с 7 видами ограничен северной частью Тихоокеанского побережья Азии и Северной Америки.
Все виды взморниковых обычно растут большими колониями, преимущественно в морских заливах, образуя подводные луга. Между родами и подродами имеются существенные различия в экологии. Так, виды филлоспадикса обитают исключительно на подводных камнях и скалах, прикрепляясь к ним толстыми и прочными корнями. В связи с этим их листья также очень прочны за счет обилия механической ткани и, в отличие от листьев взморника, редко встречаются среди береговых выносов. Отмершие листья филлоспадикса оставляют на корневищах у основания побегов обильные волокна, по присутствию которых можно легко отличить виды этого рода от видов взморника.
Виды взморника из подрода зостерелла обитают преимущественно на илистых и песчаных мелководьях, часто в зоне морских отливов, в то время как большинство видов из подрода зостера, имеющих более широкие листья, растут на глубинах свыше 1 м на песчаном или песчано-галечниковом грунте. Самый глубоководный (растущий на глубинах от 3 до 10 м) взморник азиатский (Z. asiatica) имеет наиболее широкие (8-15 мм) листья, что, несомненно, является приспособлением к недостаточному освещению на таких глубинах.
У взморниковых значительно преобладает перекрестное опыление. В роде филлоспадикс оно обеспечивается двудомностью, а в роде взморник - протогинией. Пыльца переносится водой, причем нитевидная форма пыльцевых зерен, похожих скорее на пыльцевые трубочки, чем на обычную пыльцу, способствует более эффективному опылению. Такие пыльцевые зерна отличаются большей гибкостью и, плавая под водой, могут даже закручиваться вокруг выступающих из влагалищ кроющих листьев рылец. У видов с замкнутыми влагалищами кроющих листьев они перед цветением расщепляются. Обычно опыление происходит под водой, но для видов, растущих на мелководьях, отмечается также опыление на поверхности воды: плавающие пыльцевые зерна касаются хотя бы временно выступающих из воды рылец.
Плоды взморниковых также разносятся морскими течениями, причем диаспорами могут служить не только сами плоды, но и легко обламывающиеся у своего основания плодоносящие соцветия с сохранившимися на них плодами. В береговых выносах Японского моря, на советском Дальнем Востоке, нередко можно встретить множество таких соцветий. Очень эффективно и вегетативное размножение с помощью корневищ, вследствие чего взморниковые обычно растут большими группами. Вырванные во время штормов побеги с кусками корневищ могут разноситься на большие расстояния и вновь укореняться, давая начало новым клонам.
Сухие листья взморниковых издавна использовали для набивки матрацев, мягкой мебели и других предметов обихода, а также в качестве упаковочного материала. Выносимые в больших количествах на берег листья и побеги взморника можно использовать как ценное удобрение для полей (в свежем состоянии или после сжигания). Имеются сведения об использовании плодов взморника в пищу (в качестве муки) одним из племен мексиканских индейцев.
Семейство посидониевые (Posidoniaceae)
Название единственного рода семейства по-сидониевых - посидония (Posidonia) - не случайно происходит от имени древнегреческого бога - повелителя морей Посейдона. Подобно взморниковым и цимодоцеевым, посидония принадлежит к "морским травам" - немногим цветковым растениям, обитающим в водах морей и океанов. Из трех видов посидонии один - посидония океанская (P. oceanica, рис. 19) - распространен вдоль побережья Средиземного моря и близлежащих частей Атлантического окена, 2 других, в том числе посидония южная (P. australis) - по берегам Тихого океана во внетропической Австралии и Тасмании.
Посидонии обычно растут большими колониями, образуя вместе с другими "морскими травами" своеобразные подводные луга в морских заливах и бухтах на глубинах до 30, а иногда даже до 50 м. Это полностью погруженные в воду многолетние растения с довольно толстым и прочным моноподиальным ползучим корневищем, от узлов которого отходят придаточные корни и короткие прямостоячие побеги, одетые у своего основания чехлом из волокон - остатков отмерших листьев. По наблюдениям в Южной Франции в колониях посидонии океанской развиваются вегетативные побеги двух типов: в центре колонии прямостоячие, а по ее периферии стелющиеся побеги, которые, образуя в узлах придаточные корни, становятся новыми корневищами. Стелющиеся побеги образуются и в местах изреживания колонии. Наличие побегов двух типов способствует быстрому вегетативному размножению посидонии, захвату ею все новых и новых участков дна.
Сближенные на верхушках прямостоячих и стелющихся побегов, двурядно расположенные очередные широколинейные листья посидонии достигают в длину 10-50 см и в ширину 7 -10мм.
Их длина определяется глубиной произрастания: чем глубже, тем они длиннее. Листья отчетливо разделены на пластинку с 11-17 параллельными жилками и свободное (не сросшееся краями) влагалище, образующее у основания пластинки очень короткие ушки и поперечный перепончатый вырост - язычок. В пазухах влагалищ имеются очень мелкие внутривлагалищные чешуйки.
Цветки посидонии собраны довольно густыми сложными колосьями, которые расположены на относительно коротких (обычно длиной 10- 30см) цветоносах, выходящих из верхушек укороченных ортотропных побегов или из пазух их листьев. Сложный колос обычно состоит из 2-4 колосков, расположенных в пазухах немного вздутых влагалищ сильно уменьшенных листьев. Колоски, в свою очередь, состоят из 2-3, реже 4 двурядно расположенных цветков, из которых 2 нижних всегда обоеполые, а 1-2 верхних - мужские. Околоцветник состоит из 3 быстро опадающих, иногда отсутствующих чешуек. В обоеполых цветках 3, реже 4 тычинки, представленные сидячими 4-гнездными пыльниками с сильно разросшимся связником, который у посидонии океанской имеет на верхушке хвостовидный придаток - надсвязник (рис. 19, 3). Гинецей состоит из одного плодолистика с единственным семязачатком. Плодолистик на верхушке суживается и заканчивается немного лопастным диском, по краю которого располагаются довольно длинные рыльцевые сосочки. В мужских цветках 3 тычинки. Из плодолистика формируется костянковидный плод с мясистым околоплодником, напоминающий недозрелый плод маслины.
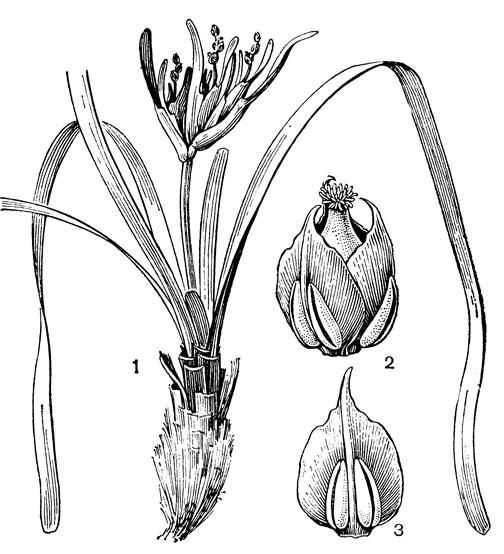
Рис. 19. Посидония океанская (Posidonia oceanica): 1 - общий вид; 2 - цветок; 3 - тычинка с нижней (наружной) стороны
Опыление цветков происходит в воде. Подобно многим другим морским травам, посидонии имеют нитевидные пыльцевые зерна, что существенно облегчает возможность контакта их с рыльцами. Самоопылению, по-видимому, препятствует протандрия. Плоды посидонии довольно долго плавают в воде и лишь после разрушения околоплодника падают на дно и прорастают. Вероятно, они распространяются также эндозоохорно, так как их мясистый околоплодник может служить пищей для рыб и водоплавающих птиц. Однако оба вида посидонии размножаются главным образом вегетативно: с помощью разносимых водой частей корневищ и отдельных побегов, которые способны легко укореняться. Цветущие экземпляры посидонии встречаются очень редко.
Листья и побеги посидонии, подобно листьям взморниковых, в большом количестве выбрасываются морем на берег. Их используют в качестве упаковочного материала, а в Северной Африке - для покрытия крыш. В Австралии из листьев посидонии южной получают волокно для изготовления грубых тканей.
Семейство наядовые (Najadaceae)
В древнегреческой мифологии наядами назывались нимфы, населяющие реки, ручьи и озера. Отсюда и происходит название единственного рода семейства наядовых - наяда (Najas, рис. 20). Около 50 видов наяд принадлежат к числу полностью погруженных в воду растений. Наяды широко распространены как в тропических, так и во внетропических областях обоих полушарий, но встречаются далеко не повсеместно, отсутствуя, например, в Арктике и значительной части таежной зоны Евразии, а также в некоторых пустынях и высокогорьях.
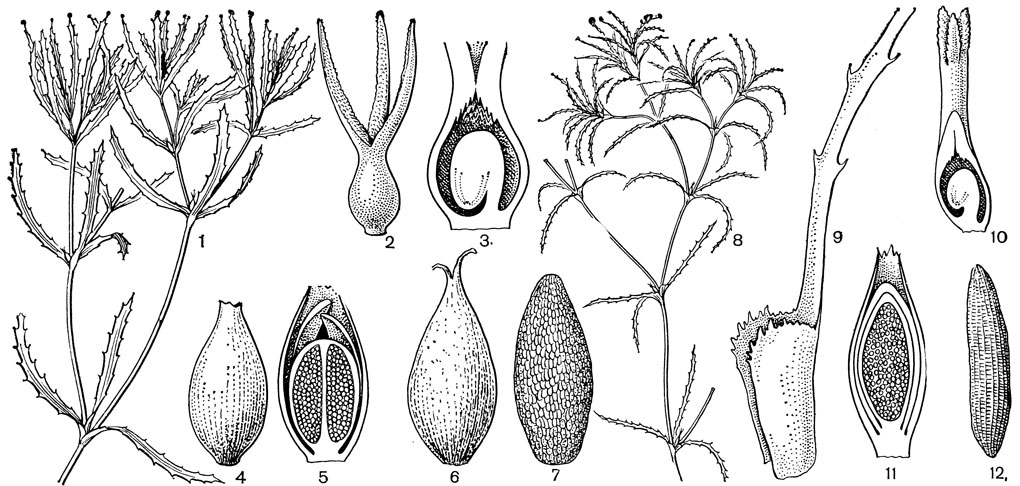
Рис. 20. Наядовые. Наяда большая (Najas major): 1 - общий вид; 2 - женский цветок; 3 - завязь с семязачатком в продольном разрезе; 4 - мужской цветок; 5 - он же в продольном разрезе; 6 - плод; 7 - семя. Наяда малая (N. minor): 8 - общий вид; 9 - нижняя часть листа с влагалищем; 10 - женский цветок в продольном разрезе; 11 - мужской цветок в продольном разрезе; 12 - семя
Из-за подводного образа жизни и невзрачного облика наяд даже опытные коллекторы нередко их не замечают. В отличие от водных растений родственных семейств - рдестовых, рупЦиевых и дзанникеллиевых - все наяды - однолетние растения, ежегодно образующие большое количество плодов и полностью отмирающие. Большинство видов имеет сильно разветвленные, но очень хрупкие стебли, легко разламывающиеся на отдельные части, которые плавают у поверхности воды, продолжая цветение и плодоношение. Из 8 наяд флоры СССР лишь у наяды гибкой (N. flexilis) гибкие стебли, похожие на стебли узколистных рдестов. Стебли представителей подрода собственно наяд (Najas) часто имеют рассеянные шипы, а у североафриканской наяды Делилё (N. delilei) они почти по всей длине (особенно в верхней части) густо покрыты шипами.
У всех наяд линейные, часто почти нитевидные, сидячие листья с одной не всегда заметной жилкой, обычно легко отличающиеся от листьев других водных растений относительной жесткостью и хрупкостью. Кроме того, у собственно наяд по краю листьев, а иногда и вдоль средней жилки расположены колючие зубцы. У видов другого подрода - каулинии (Caulinia), нередко выделяемого в самостоятельный род, значительно более узкие листья также имеют по краю зубцы, но обычно очень короткие и малозаметные. У некоторых видов этого подрода, например у наяды гибкой и наяды злаковидной (N. graminea), зубчики по краю листьев заметны лишь при увеличении. Чаще всего листья наяд располагаются на стебле ложными мутовками по 3, реже по 1-2. У них короткие, но хорошо развитые влагалища,, обычно по верхнему краю зубчатые и нередко образующие тупые или острые ушки по бокам основания листовой пластинки. В пазухах влагалищ обычно расположены 2 очень мелкие внутривлагалищные чешуйки, вероятно предлистовой природы.
Как и у многих других полностью погруженных в воду растений, цветки наяд сильно упрощены. У всех наяд однополые цветки, причем цветки разных полов могут находиться на одном и том же растении (у видов подрода каулиния) или на разных (у собственно наяд). И мужские и женские цветки располагаются в узлах стебля по одному или по 2-4. Они кажутся сидячими, но в действительности заканчивают собою сильно редуцированные боковые веточки. Мужской цветок состоит из одного сидячего пыльника с 1 или 4, реже 2 гнездами, расположенного на очень короткой ножке и одетого трубчатым или бутылковидным, на верхушке как бы двугубым внутренним покрывалом. Это покрывало обычно принимают за околоцветник, хотя некоторые авторы считают, что оно образовано прицветниками. Женские цветки всех наяд не имеют такого околоцветника и состоят из одного почти сидячего плодолистика с одним базальным анатропным семязачатком. Верхняя, суженная часть плодолистика заканчивается 2, реже 3-4 рыльцевыми ветвями, что позволяет некоторым авторам принимать гинецей наяд за псевдомономерный, образованный более чем одним плодолистиком. У многих видов наяд есть еще одно, внешнее покрывало цветков обычно бутылковидной формы. У собственно наяд, например у наяды большой (N. major, рис. 20, 4), такое покрывало имеется только при мужских цветках, а у женских оно отсутствует или прирастает к завязи. В подроде каулиния внешнее покрывало может вообще отсутствовать, а если имеется, то у цветков обоих полов, например у наяды малой (N. minor, рис. 20, 10,11). Это покрывало считают гомологичным прицветнику или влагалищу обычного листа, что подтверждается наличием на его верхушке зубчиков, почти всегда имеющихся по краю верхушки листовых влагалищ.
Плоды наяд нераскрывающиеся, односемянные, с тонким, но мясистым околоплодником. Их форма варьирует от широкояйцевидной (у казахстанской наяды короткоплодной - N. marina subsp. brachycarpa) до очень узкоэллипсоидальной (у многих видов подрода каулиния). Скульптура семенной оболочки, обычно представленная ячейками разной величины и формы, служит важным систематическим признаком как у современных, так и у ископаемых наяд.
Хотя большинство наяд встречается в пресных водоемах, имеются немногие виды, например широко распространенная в Евразии наяда морская (N. marina), обитающие в солоноватых водоемах, обычно в приморских лагунах и соленых озерах. Некоторые виды могут расти в небольших, быстро пересыхающих водоемах, а широко распространенная в тропиках и субтропиках Евразии наяда злаковидная нередко в изобилии разрастается на рисовых полях, являясь их сорняком. В Бирме этот вид наблюдали в образованных теплыми источниками водоемах с температурой +35 °С, а другой вид - наяда тонколистная (N. tenuifolia) - найден на острове Ява даже в вулканических озерах с температурой до +60 °С. Многие наяды принадлежат к числу редчайших растений, например известная лишь из немногих местонахождений на территории Евразии (в том числе на северном побережье Финского залива) наяда тончайшая (N. tenuissima).
Цветки всех наяд опыляются под водой. После созревания пыльцы ножка мужского цветка заметно удлиняется и выносит пыльник вместе с внутренним покрывалом - околоцветником - из внешнего покрывала. Губы околоцветника расходятся в стороны, пыльник открывается отверстиями на его верхушке, и пыльца через расширившийся канал между губами выходит в воду. Пыльцевые зерна наяд шаровидные или широкоэллипсоидальные, богатые крахмалом и почти лишенные наружной оболочки - экзины, но пыльцевые зерна наяды большой дистально-однобороздные и покрыты тонкой сетчатой экзиной (З. В. Волховских, 1982). Уже во время выхода в воду из пыльника они начинают прорастать, образуя пыльцевую трубочку, что существенно облегчает возможности контактов между ними и покрытыми очень короткими сосочками рыльцевыми ветвями. Прорастающие до попадания на рыльца пыльцевые зерна наяд можно считать переходными к еще более высокоспециализированным, нитевидным пыльцевым зернам, характерным для "морских трав" из семейств взморниковых, цимодоцеевых и посидониевых.
Плоды наяд обычно распространяются водными потоками. Ломкость стеблей большинства видов, по-видимому, также способствует распространению плодов вместе с частями материнского растения. Вероятно, плоды наяд могут распространяться и эндозоохорно: с помощью поедающих их рыб и водоплавающих птиц.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'