Порядок частуховые (Alismatales) (Н. Н. Цвелев)
Семейство сусаковые (Butomaceae)
Почти на всей территории СССР по берегам водоемов обычно среди другого болотного высокотравья встречается сусак зонтичный (Butomus umbellatus) - единственный представитель семейства сусаковых. Его собранные в крупные зонтиковидные соцветия светло-розовые цветки сразу бросаются в глаза и, наверное, знакомы многим читателям (рис. 1). Сусак широко распространен в Европе и внетропических областях Азии, исключая Арктику, север таежной зоны и высокогорья свыше 1000 м над уровнем моря. Кроме того, он занесен в Северную Америку и вполне натурализовался на юго-востоке Канады и северо-востоке США. В наиболее континентальных районах Азии сусак зонтичный представлен более мелкой и узколистной формой, иногда выделяемой в качестве особого вида - сусака ситникового (В. junceus).

Рис. 1. Сусак зонтичный (Butomus umbellatus): 1 - общий вид; 2 - лист на поперечном разрезе; 3 - соцветие; 4 - цветок в мужской фазе; 5 - цветок в женской фазе; 6 - плодолистик; 7 - он же на продольном разрезе (видны семязачатки); 8 - тычинки во время мужской фазы; 9 - тычинки во время женской фазы; 10 - плод; 11 - семя
Сусак - довольно крупное (высотой 40- 150 см) многолетнее растение с длинным и толстым (обычно 1,5-2 см) горизонтальным моноподиальным корневищем, на нижней стороне которого образуются многочисленные корни, а на верхней стороне двумя рядами расположены трехгранные линейные листья. Из пазух листьев выходят вегетативные почки, дающие начало новым корневищам, и безлистные ножки соцветий. Последние обычно образуются в пазухе каждого девятого листа корневища (включая отмершие листья) на расстоянии 4-7 см друг от друга в числе 1-3 за один летний сезон. Боковые почки корневища, одетые предлистом и чешуевидными низовыми листьями - катафиллами, позднее легко теряют связь с материнским корневищем, давая начало новым особям сусака. У основания листьев имеются хорошо развитые открытые влагалища, а в их пазухах - многочисленные бесцветные внутривлагалищные чешуйки, характерные для многих других водных и болотных однодольных. На них находятся железки, выделяющие вязкую слизь, которая, вероятно, имеет защитное значение. Сосуды у сусака имеются только в корнях.
Расположенные на длинном цилиндрическом цветоносе соцветия сусака имеют вид простых зонтиков с оберткой из прицветников. Однако в действительности этот зонтик является ложным и состоит из одного верхушечного цветка и трех самостоятельных соцветий - извилин, выходящих из пазух прицветников и нередко также разветвленных. Впечатление зонтика создается благодаря сильно укороченным осям извилин и длинным цветоножкам. До цветения соцветия окутаны прицветниками, которые позднее отгибаются вниз.
Околоцветник актиноморфных и обоеполых цветков сусака состоит из 6 расположенных двумя чередующимися кругами светло-розовых или розовато-белых сегментов, из которых наружные - чашелистики - лишь немного мельче внутренних. В отличие от близких семейств лимнохарисовых и частуховых у сусака все сегменты околоцветника сохраняются при плодах. Из 9 тычинок, имеющих лентовидно расширенные нити, 6 наружных расположены по 2 перед наружными сегментами околоцветника, а 3 внутренних - по одной перед внутренними. Пыльцевые зерна сусака однобороздные. Гинецей состоит из 6 лишь близ основания сросшихся друг с другом примитивных кондупликатных и еще не вполне замкнутых плодолистиков, расположенных в 2 не очень четко разграниченных между собой круга. Каждый плодолистик имеет многочисленные анатропные семязачатки и переходит на верхушке в столбик с низбегающим верхушечным рыльцем. Плацентация примитивного ламинально-диффузного типа. Так называемые септальные нектарники находятся в щелях между нижними частями плодолистиков. Выделяемый ими довольно обильный нектар скапливается в виде капелек кнаружи от щелей между плодолистиками. Сахар содержится и в соке, выделяемом рыльцами, которые также отчасти функционируют как дополнительные нектарники.
Цветки сусака энтомофильны и опыляются мелкими перепончатокрылыми, мухами, жуками и другими насекомыми. Самоопылению препятствует протандрия (рис. 1, 4 и 5). Первыми вскрываются 6 наружных тычинок, затем через некоторое время 3 внутренних, причем сначала
они направлены вверх, а затем расходятся в стороны. После освобождения всех пыльников начинается женская фаза цветения, к наступлению которой плодолистики увеличиваются почти вдвое с момента начала цветения. В начале женской фазы все же возможно самоопыление, если в пыльниках сохранилось хотя бы немного пыльцы, перенос которой на рыльца может осуществляться с помощью насекомых или ветра.
В плоде сусака (многолистовке) каждая его часть (листовка) вскрывается по шву плодолистика. Мелкие и легкие короткоцилиндрические семена выпадают из листовок при раскачивании плодоносящих соцветий ветром или крупными животными. Попадая в воду, они довольно быстро тонут, но все же могут распространяться на небольшие расстояния с помощью водных потоков или водоплавающих птиц, а также на ногах животных вместе с комочками почвы. Кроме того, сусак легко размножается вегетативно боковыми почками корневища, которые могут переноситься водой после отделения их от материнского растения на большие расстояния.
Толстые, богатые крахмалом корневища сусака съедобны в печеном виде, а из его листьев можно делать маты, циновки и другие плетеные изделия. Кроме того, семена и корневища используют в Западной Европе в качестве народного лекарственного средства.
Семейство лимнохарисовые (Limnocharitaceae)
Относительно небольшое (4 рода и 14 видов) семейство лимнохарисовых прежде объединялось с семейством сусаковых, от которого оно отличается рядом существенных признаков, в том числе разделенными на пластинку и черешок листьями и двойным околоцветником с опадающими при плодах лепестками. Кроме того, в отличие от сусака, распространенного во внетропических странах, все лимнохарисовые исключительно тропические водные и болотные травы. Род тенагохарис (Tenagocharis) с одним видом тенагохарисом широколистным (Т. latifolia) распространен в тропиках Африки, Азии и Австралии, а остальные роды - лимнохарис (Limnocharis), гидроклейс (Hydrocleis) и остения (Ostenia) - приурочены к тропикам Америки. Лимнохарис желтый (L. flava, рис. 2, 1, табл. 1, 2) интродуцирован и натурализовался в Индии и Юго-Восточной Азии, а гидроклейс кувшинковидный (Н. nymphoides, рис. 2, 7, табл. 1, 3) нередко культивируют в парках и ботанических садах тропических, а отчасти и субтропических стран Старого Света.

Таблица 1. Апоногетоновые и лимнохарисовые: 1 - апоногетон жестколистный (Aponogeton rigidifolius), оранжереи Ленинградского государственного университета; 2 - лимнохарис желтый (Limnocharis flava), оранжереи Ботанического института АН СССР в Ленинграде; 3 - гидроклейс кувшинковидный (Hydrocleys nymphoides), Африка, Гайана
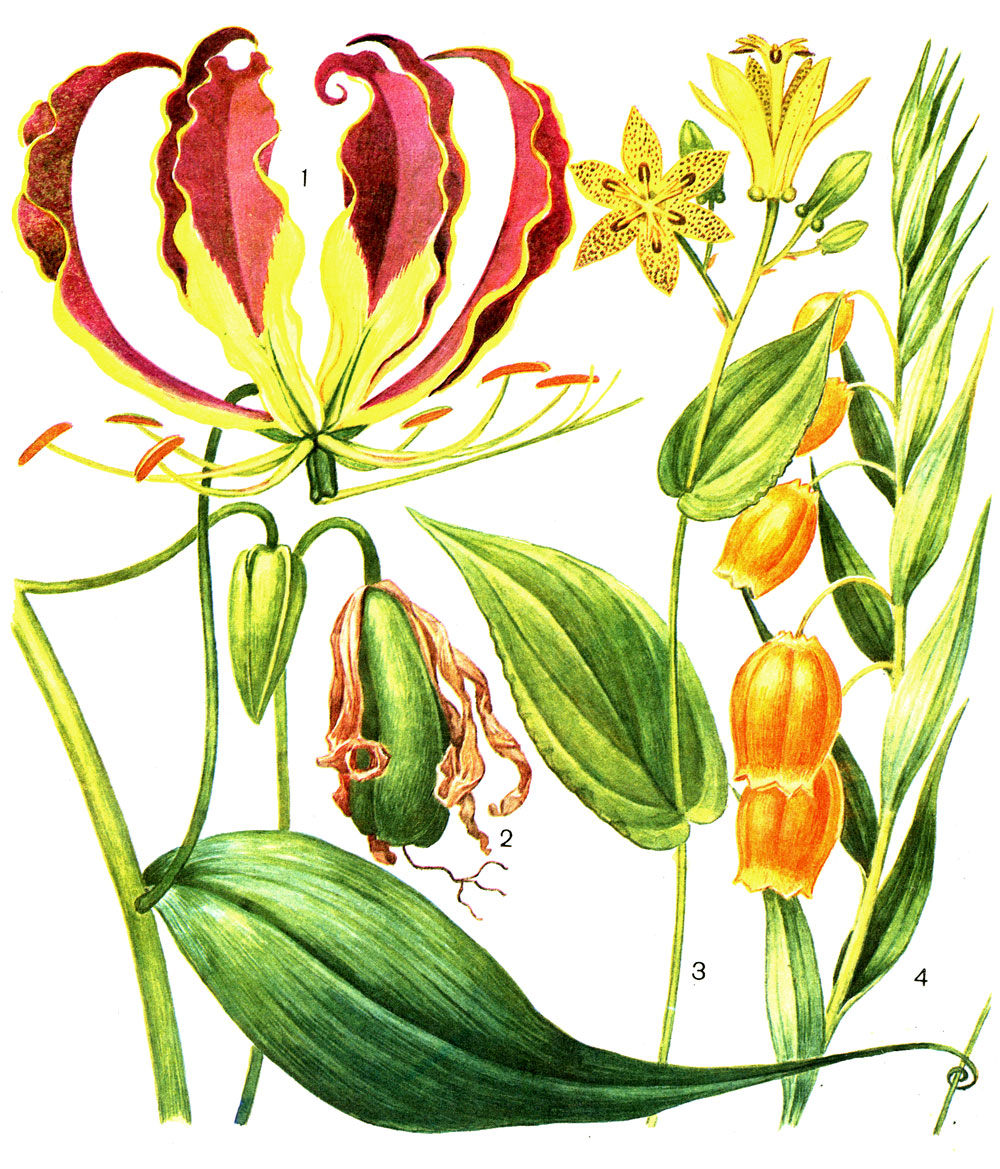
Таблица 2. Мелантиевые. Глориоза великолепная (Gloriosa superba), оранжереи Ботанического института АН СССР в Ленинграде: 1 - часть растения с цветком; 2 - плод. Трициртис крупноногий (Tricyrtis macropoda), там же: 3 - часть растения. Сандерсония оранжевая (Sandersonia aurantiaca): 4 - часть растения
Лимнохарис желтый - многолетнее болотное растение с очень коротким утолщенным корневищем, состоящим из сильно укороченных междоузлий и несущим розетку довольно крупных листьев, имеющих короткое открытое влагалище, черешок и широкоэллиптическую или яйцевидную пластинку с дуговидно-кривобежным жилкованием. Из пазух листьев выходят или безлистные прямостоячие ножки зонтикообразных соцветий, или поникающие и затем ложащиеся на землю ножки вегетативных почек, дающие начало новым особям. Как и у сусаковых, сосуды имеются только в корнях. Но, в отличие от сусаковых, во всех вегетативных частях растения имеются млечные канальцы, а на нижней стороне листовых пластинок под их верхушкой - крупные водяные устьица - гидатоды, выделяющие избыток воды. С лимнохарисом внешне довольно сходен тенагохарис широколистный, который, однако, является однолетником и вегетативно не размножается.
Остальные два рода лимнохарисовых - гидроклейс и остения - многолетние земноводные растения с длинными, обычно разветвленными стеблями, плавающими в воде или реже стелющимися по илистой почве и укореняющимися в узлах. У обычной водной формы гидроклейса кувшинковидного (рис. 2, 7) листья с длинными черешками и плавающей на поверхности воды широкояйцевидной с сердцевидным основанием пластинкой. Однако погруженные в воду и собранные в розетку листья молодых особей этого вида имеют линейную форму и не разделены на черешок и пластинку. Плавающие ветви гидроклейса обычно заканчиваются пучками укороченных побегов с несколькими листьями и выходящим из пазухи одного из листьев крупным одиночным цветком. У эндемичного для Уругвая рода остении плавающие листья имеют эллиптические пластинки. У гидроклейса и остении стебли и их ветви могут обрываться и продолжать свое развитие в свободно плавающем состоянии.
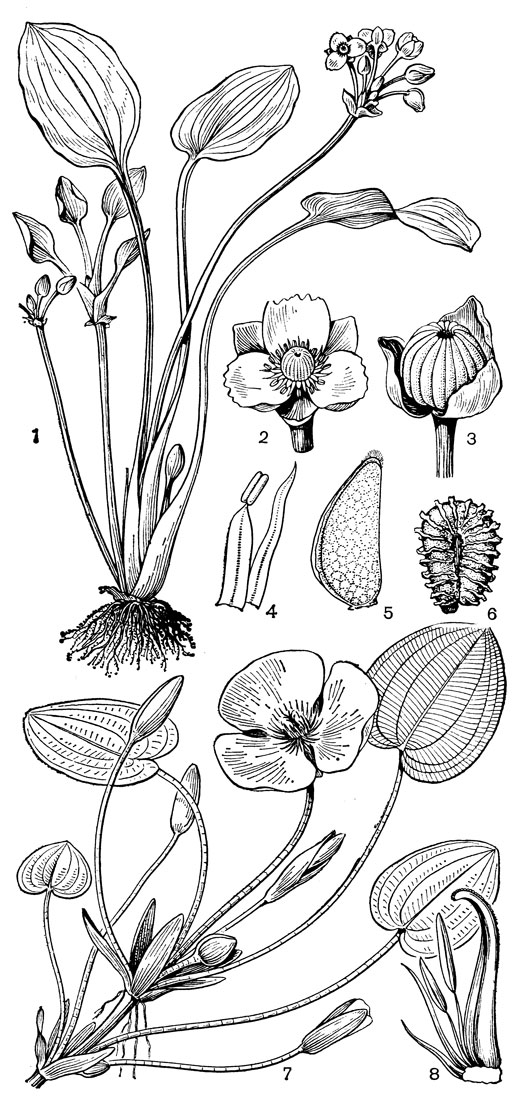
Рис. 2. Лимнохарисовые. Лимнохарис желтый (Limnocharis flava): 1 - общий вид; 2 - цветок; 3 - плод с чашечкой; 4 - тычинка и стаминодий; 5 - плодик; 6 - семя. Гидроклейс кувшинковидный (Hydrocleis nymphoides): 7 - общий вид; 8 - плодолистик с 2 тычинками и 2 стаминодиями
Зонтикообразные соцветия лимнохариса и тенагохариса, подобно соцветиям сусака, являются не зонтиком, а сложным соцветием, состоящим из верхушечного цветка и одного или нескольких соцветий - извилин с сильно укороченной осью. Одиночные цветки гидроклейса и остении всегда возвышаются над поверхностью воды. Все лимнохарисовые имеют обоеполые актиноморфные цветки, околоцветник которых отчетливо разделен на 3 обычно зеленых и остающихся при плодах чашелистика и 3 чередующихся с ними опадающих лепестка желтой, реже белой (у тенагохариса) окраски. Ярко-желтые венчики гидроклейса кувшинковидного достигают в диаматре 4-5 см. Число тычинок в цветке относительно постоянно только у тенагохариса, обычно имеющего 9 тычинок с расширенными ланцетными нитями. У представителей других родов тычинки многочисленные, расположенные в несколько многочленных кругов, причем наружный круг составляют стаминодии - стерильные тычинки без пыльников. В онтогенезе развитие тычинок начинается от центра цветка: сначала развивается самый внутренний круг тычинок, затем расположенный кнаружи от него и наконец круг стаминодиев. Пыльники у всех родов двугнездные с шаровидными безапертурными или 3-4-поровыми пыльцевыми зернами.
Гинецей цветка лимнохарисовых состоит из свободных или немного сросшихся у основания плодолистиков, число и строение которых различно у разных родов, но они всегда имеют многочисленные семязачатки. Плодолистики еще более или менее открытые, кондупликатные, а плацентация ламинально-диффузная. У лимнохариса гинецей наиболее примитивен: многочисленные (15-20) свободные, но приросшие основанием к цветоложу плодолистики имеют сидячие рыльца в виде окруженных сосочками продольных щелевидных отверстий в верхней наружной части плодолистиков. У тенагохариса гинецей представлен обычно 9 плодолистиками, оттянутыми на верхушке в короткий столбик, заканчивающийся покрытым сосочками дисковидным рыльцем. У гидроклейса и остении 3 или 6 (редко 4 или 8) ланцетнолинейных, едва сросшихся у основания плодолистиков, постепенно переходящих в дуговидно согнутый столбик, на верхушке и внутренней стороне которого находится рыльце. Плод у всех лимнохарисовых - многолистовка, части которой - листовки - вскрываются щелью по обращенному внутрь шву плодолистика. Семена имеют гладкую, реже (у лимнохариса) поперечно-морщинистую оболочку и подковообразный (как у частуховых) зародыш.
Цветки лимнохарисовых опыляются различными насекомыми, посещающими их главным образом ради пыльцы, так как они содержат очень мало нектара. У гидроклейса имеются слабо развитые септальные нектарники в щелях между основаниями плодолистиков, а у лимнохариса отмечается небольшое количество нектара на стаминодиях и на поверхности рылец.
Цветки лимнохариса и тенагохариса, по-видимому, способны самоопыляться, если не произошло перекрестного опыления. Так, у тенагохариса уже вскоре после начала цветения завядшие лепестки смыкаются друг с другом, прижимая еще содержащие пыльцу пыльники к рыльцам. В цветках лимнохариса уже через несколько часов после начала цветения чашелистики смыкаются друг с другом, а лепестки и тычинки превращаются в почти однородную слизистую массу. Для гидроклейса, который имеет особенно крупные яркоокрашенные цветки, цветущие не более одного дня, самоопыление менее вероятно.
Семена лимнохарисовых обладают плавучестью и разносятся главным образом водными потоками. Однако они могут распространяться и экзозоохорно: на шерсти животных и перьях водоплавающих птиц, а также на их ногах с комочками почвы. Поперечные морщины с бугорковидными выростами на семенах лимнохариса (рис. 2, 6), вероятно, способствуют такому распространению.
Гидроклейс кувшинковидный благодаря крупным ярко-желтым цветкам очень декоративен и довольно часто культивируется в водоемах парков и ботанических садов в тропиках и субтропиках, а севернее - в оранжереях и крупных аквариумах. В СССР этот вид можно встретить в парках Черноморского побережья и в оранжереях ботанических садов, где он известен под названием "водяного мака". Цветки его действительно немного напоминают желтоцветковые полярные маки. Лимнохарис желтый, имеющий сочные и крупные листья, использует в пищу в качестве ценного овощного или салатного растения население как Южной Америки, так и Южной Азии, где этот вид встречается в качестве интродуцированного, но местами вполне натурализовавшегося растения.
Семейство частуховые (Alismataceae)
К семейству частуховых, включающему 13-14 родов и около 100 видов, принадлежат такие широко распространенные в СССР растения берегов водоемов и болот, как стрелолист обыкновенный (Sagittaria sagittifolia, рис. 3, 1) со стреловидными листьями и кистями довольно крупных почти белых цветков и частуха обыкновенная (Alisma plantago-aquatica рис. 4, 7), распространенная повсеместно и часто растущая на влажных местах вдоль дорог и троп подобно широко известному подорожнику большому (Plantago major), за что еще К. Линнеем была названа "водяным подорожником". Частуховые - почти космополитное семейство, виды которого отсутствуют только в значительной части Арктики, на многих островах Тихого океана, в некоторых пустынях и высокогорьях. В северном полушарии они представлены богаче, чем в южном, а виды наиболее крупных родов семейства - стрелолист (Sagittaria) и эхинодорус (Echinodorus) - особенно многочисленны в Америке.
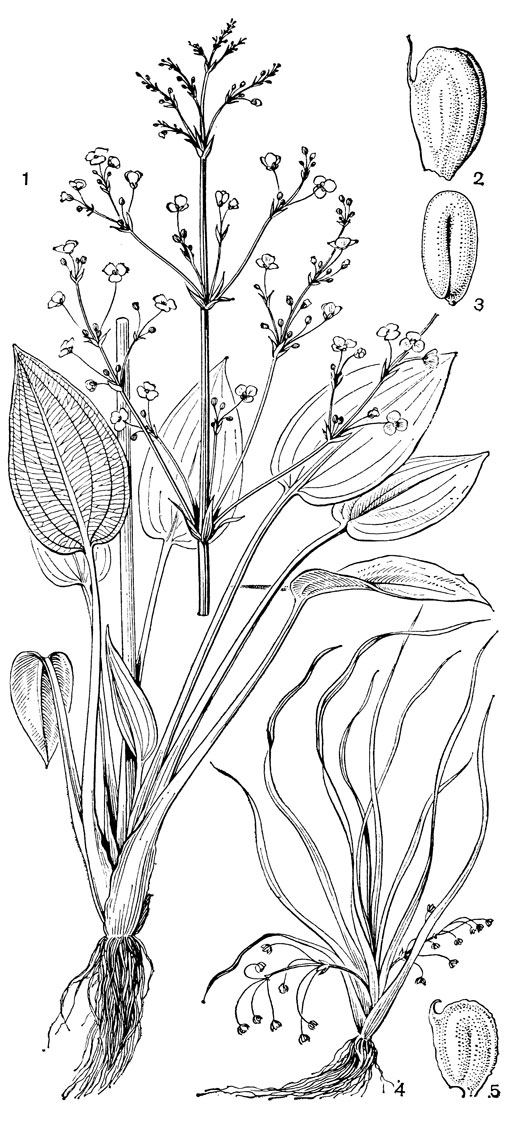
Рис. 4. Виды частухи (Alisma). Частуха обыкновенная (A. plantago-aquatica): 1 - общий вид; 2 - плодик; 3 - семя. Частуха Валенберга (A. wahlenbergii): 4 - общий вид; 5 - плодик
Почти все частуховые - многолетние розеткообразующие травы с коротким и толстым, часто клубнеобразным корневищем, на верхушке которого ежегодно образуются розетка листьев и безлистная ножка, несущая соцветие. Нередко в этот же год образуются еще 1-2 соцветия в определенной последовательности. Корневище в виде толстого, почти шаровидного клубня имеет африканская бурнатия (Burnatia). К немногим частуховым-однолетникам принадлежат африканская раналисма низкая (Ranalisma humile) и североамериканский стрелолист лопатчатый (Sagittaria spathulata). Оба эти растения, достигающие всего 2-7 см в высоту, встречаются на временно затопляемых берегах рек и озер, а также на месте пересохших водоемов. Раналисма низкая - единственное частуховое с всегда одиночными цветками на коротких цветоножках (рис. 5, 5), а у стрелолиста лопатчатого очень короткие ножки соцветий несут всего одну мутовку из 1-3 цветков.
Все частуховые - влаголюбивые растения, и многие из них могут расти как на суше (обычно по берегам водоемов, на болотах и болотистых лугах), так и в воде, хотя соцветия почти всегда подняты над ее поверхностью. Стрелолист обыкновенный может заходить в водоемы до глубины 5 м, но на больших глубинах не образует цветков и имеет только линейные подводные листья. Примером немногих полностью погруженных в воду частуховых может служить частуха Валенберга (Alisma wahlenbergii, рис. 4, 4) - небольшое растение с узколинейными листьями и короткими, дуговидно изогнутыми книзу ножками соцветий, обитающее на песчаном дне лагун и бухт в северной части Балтийского моря. Его соцветия, несущие только клейстогамные цветки, нередко оказываются полностью погруженными в песок, так что этот вид легко принять за молодые вегетативные побеги других водных растений. Близкий вид - частуха злаколистная (A. gramineum) имеет уже две очень отличающиеся друг от друга формы: подводную - с линейными листьями и обычно тоже клейстогамными цветками и наземную - обычно карликовую с хазмогамными цветками и листьями, имеющими ланцетные пластинки. Наземную форму даже описывали в качестве самостоятельного вида - частухи дуговидной (A. arcuatum).
Разнолистность вообще свойственна очень многим земноводным частуховым. Хорошим примером в этом отношении может служить стрелолист обыкновенный, различное строение листьев которого было отмечено и изображено еще в 1703 г. известным ботаником того времени Лёзелем. Развиваясь на дне водоемов, стрелолист обыкновенный сначала образует розетку сидячих широколинейных подводных листьев, затем длинночерешковые, плавающие на поверхности воды листья с эллиптическими или немного стреловидными пластинками и наконец возвышающиеся над водой длинночерешковые листья со стреловидными пластинками (рис. 3, 1). Часто только плавающие листья с сердцевидными пластинками имеет тропический афроазиатский стрелолист цепкоплодный (Sagittaria lappula, рис. 3, 8), а у североамериканского стрелолиста валъковатого (S. teres, рис. 3, 5) все листья подводные, в виде почти цилиндрических, поперечно-перегородчатых черешков без пластинок. Широколинейные подводные листья прикорневой розетки и длинно-черешковые плавающие листья с эллиптическими пластинками имеет европейский эндемик - лурониум плавающий (Luronium natans), однако у него плавающие листья отходят главным образом от узлов сильно удлиненного и плавающего в воде соцветия (рис. 5, 10).
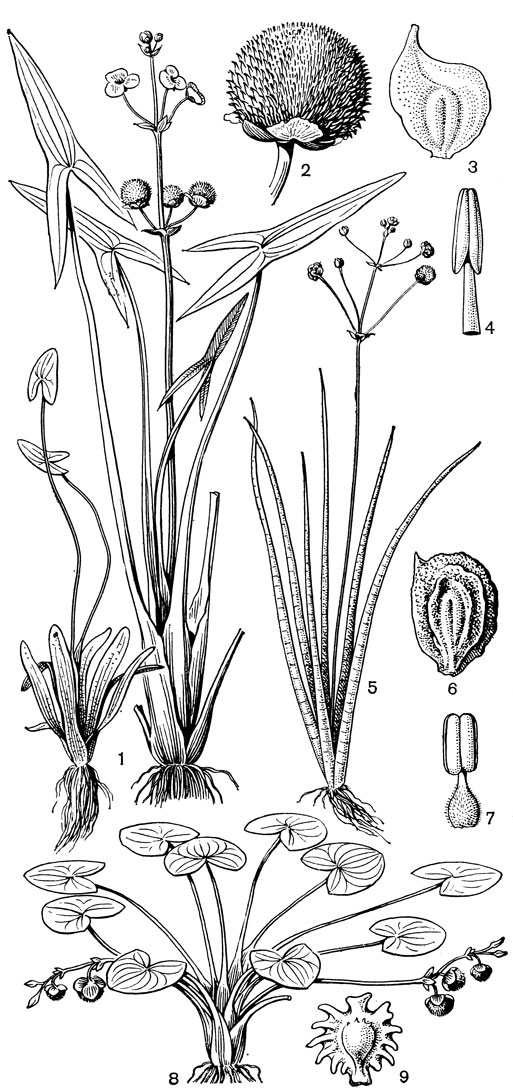
Рис. 3. Виды стрелолиста (Sagittaria). Стрелолист обыкновенный (S. sagittifolia): 1 - общий вид; 2 - плод; 3 - плодик; 4 - тычинка. Стрелолист вальковатый (S. teres): 5 - общий вид; 6 - плодик; 7 - тычинка. Стрелолист цепкоплодный (S. lappula): 8 - общий вид; 9 - плодик
Линейные подводные листья частуховых, представляющие в действительности лишь сильно расширенные листовые черешки без пластинок, имеют параллельное жилкование. Плавающие и возвышающиеся над водой листья, а также листья наземных частуховых ясно дифференцированы на черешок и пластинку различной формы, обычно с дуговидно-кривобежным жилкованием, причем основные жилки соединяются между собой поперечными анастомозами. Иногда, например у раналисмы длинноносиковой (Ranalisma rostratum), основных жилок может быть всего 1-2, не считая средней жилки (рис. 5,2). Основания черешков часто расширены в короткие свободные влагалища, в пазухе которых обычно имеются мелкие внутривлагалищные чешуйки с железками, выделяющими слизистый секрет.
Обоеполые, реже однополые, всегда актиноморфные цветки частуховых обычно собраны в кистевидные или метелкообразные соцветия, расположенные на безлистных стеблях. В узлах соцветий имеются лишь видоизмененные, нередко чешуевидные листья и прицветники. Листья с хорошо развитыми пластинками отсутствуют в соцветиях всех частуховых, кроме лурониума, у которого длинные плавающие в воде соцветия несут в узлах нормально развитые плавающие листья, поддерживающие соцветие у поверхности воды. Цветки лурониума возвышаются над водой, производя впечатление одиночных, а не собранных в соцветие (рис. 5, 10). Сильно разветвленные соцветия некоторых видов частухи и калъдезии (Caldesia) могут достигать метра в высоту и нести весьма многочисленные цветки. Многие другие виды из разных родов семейства, например стрелолист обыкновенный (рис. 3, 1) и звездоплодник многосемянный (Damasonium polyspermum, рис. 5, 6), имеют кистевидные или зонтиковидные соцветия, у раналисмы низкой (рис. 5, 5) редуцированные до одного цветка на короткой цветоножке. Цветки и веточки в соцветиях частуховых почти всегда располагаются мутовками, чаще всего по 3. У представителей афроазиатского тропического рода виснерия (Wisneria) прицветники у основания каждой мутовки соцветия срастаются своими боковыми сторонами в чашевидную или колокольчатую обертку.
Околоцветник цветков частуховых отчетливо разделен на чашечку и венчик. Чашечка состоит из 3 зеленых чашелистиков, обычно остающихся, нередко даже разрастающихся при плодах, а венчик - из 3 белых, реже розовато-белых или розовых, обычно опадающих при плодах лепестков. В клейстогамных цветках, в том числе у упомянутой выше частухи Валенберга, а также у видов бурнатии и виснерии лепестки очень слабо развиты или вообще отсутствуют. Тычинок в цветке обычно 6, реже 9 или более, со свободными, нитевидными или расширенными в нижней части нитями и двугнездными пыльниками. Только в цветках виснерии всего 3 тычинки. У многих родов с 6 тычинками, в том числе у частухи, они располагаются парами перед лепестками, в других случаях 6, 9 или 12 тычинок расположены чередующимися кругами по 3, причем тычинки наружного круга противостоят чашелистикам. У стрелолиста, эхинодоруса и близких родов многочисленные тычинки расположены по спирали.
Гинецей состоит из свободных, редко (у звездоплодника) сросшихся у основания друг с другом плодолистиков, число которых варьирует от 3 и 6 до многочисленных в неопределенном количестве, причем в последнем случае они могут располагаться мутовчато в один круг (у частухи) или по спирали на сильно выпуклом цветоложе (у стрелолиста). Интересно, что у частухи края плодолистиков во время цветения не замкнуты, затем тесно смыкаются, но даже у зрелого плода не срастаются друг с другом. Более или менее длинный столбик, переходящий в покрытое сосочками рыльце, отходит или от верхушки плодолистика, или от его внутренней стороны ниже верхушки. Почти у всех родов семейства каждый плодолистик имеет только один базальный или почти базальный семязачаток. Лишь звездоплодник является в этом отношении исключением: 3 его вида имеют от 2 до 7 семязачатков в плодолистике, а звездоплодник многосемянный (рис. 5, 8) даже 7-20, напоминая представителей близкого семейства лимнохарисовых.

Рис. 5. Частуховые. Раналисма длиннонос и кован (Ranalisma rosratum): l - общий вид; 2 - цветок; 3 - плод в продольном разрезе; 4 - плодик. Раналисма низкая (R. humilе): 5 - общий вид. Звездоплодник многосемянный (Damasonium polyspermum): 6 - общий вид; 7 - цветок; 8 - незрелый плод на продольном разрезе; 9 - семя. Лурониум плавающий (Luronium natans): 10 - общий вид; 11 - плодик
Формирующийся из гинецея плод частуховых обычно легко распадается на более или менее многочисленные орешкообразные или мешочкообразные, редко (у кальдезии) костянкообразные части - плодики, содержащие по одному семени. Лишь у звездоплодника плодики, становящиеся звездчато распростертыми за счет разрастания цветоложа, долго остаются в соединении друг с другом и часто содержат больше одного семени" Состоящие из орешкообразных спирально расположенных плодиков плоды некоторых частуховых, например раналисмы или балъделлии (Balcleliia), очень похожи на плоды некоторых лютиков (Ranunculus). Семена частуховых лишены эндосперма и имеют гладкую или поперечно бугорчато-морщинистую оболочку, сквозь которую часто просвечивает зародыш очень характерной для семейства подковообразной формы.
Как уже отмечалось выше, все частуховые - обитатели более или менее переувлажненных местообитаний: водоемов, болот и болотистых лугов, нередко заходящие довольно глубоко в воду. Не только лурониум плавающий, но и многие виды стрелолиста вообще, как правило, растут в воде, имея только погруженные и плавающие на поверхности воды листья. Некоторые эфемерно вегетирующие виды, например виды звездоплодника, могут в изобилии разрастаться на месте быстро пересыхающих водоемов.
Все частуховые, кроме немногих видов с исключительно клейстогамными цветками, опыляются разнообразными насекомыми, а отчасти также улитками, хотя у нередко растущей большими зарослями частухи обыкновенной и других видов рода не исключена возможность опыления цветков с помощью ветра. Отчасти ветроопыляемой может быть и африканская бурнатия девятитычинковая (Burnatia enneandra) - единственный представитель семейства с двудомными однополыми цветками, имеющими очень мелкие лепестки и собранными в крупные метелкообразные соцветия. В мужском цветке этого растения 9 тычинок и около 12 рудиментарных плодолистиков, а в женском цветке - около 12 плодолистиков и часто 1-2 рудиментарные тычинки. У многих частуховых с обоеполыми цветками, по-видимому, нередко происходит самоопыление. Обычно самоопыляющейся, вероятно, является наземная форма частухи злаколистной, имеющая более мелкие лепестки и более короткие столбики, чем у других, преимущественно наземных, видов частух. Ее подводная форма с клейстогамными цветками составляет переход к облигатно клейстогамной частухе Валенберга.
Многочисленные виды стрелолиста, а также виды виснерии обычно имеют однополые, но однодомные цветки, располагающиеся в разных частях соцветия: мужские - в верхней, а женские - в нижней его части. Уже у стрелолиста обыкновенного нередко встречаются обоеполые цветки, а у ряда тропических видов этого рода, в том числе у стрелолиста цепкоплодного, нижние цветки обычно имеют по одному кругу вполне развитых тычинок, а верхние мужские цветки - довольно крупные рудименты гинецея. У видов стрелолиста верхние цветки обычно зацветают позднее нижних, что отчасти препятствует самоопылению.
В цветках частуховых приспособлениями к энтомофилии служат обычно яркая окраска венчиков и присутствие нектара в цветках. У частухи, кальдезии и многих других частуховых имеются только характерные для многих однодольных септальные нектарники в щелях между плодолистиками. У видов эхинодоруса они отсутствуют, но имеются слабо функционирующие нектарники у основания листочков околоцветника или вокруг основания гинецея. В цветках стрелолиста слабо развитые нектарники располагаются у основания всех тычинок и плодолистиков, но особенно сильно развиты они у основания стаминодиев и рудиментарных плодолистиков.
Плодики большинства частуховых имеют подэпидермальную воздухоносную ткань и способны даже в течение нескольких месяцев плавать по поверхности воды. Лишь после разрушения этой ткани семена падают на дно водоема и прорастают. Гидрохорный способ распространения дополняется другими. Так, очень легкие и обладающие большой "парусностью" за счет присутствия крыловидной каймы плодики стрелолиста могут распространяться и с помощью ветра. Еще большее значение имеет экзозоохорный способ распространения: плодики многих частуховых, в особенности видов частухи, могут переноситься с комочками почвы на ногах животных и человека. У некоторых видов на плодиках имеются различные выросты, способствующие экзозоохории, например у стрелолиста цепкоплодного (рис. 3, 9) или у кальдезии шипоплодной (Caldesia acanthocarpa). Долго сохраняющиеся на растении твердые и острые плодики звездоплодника (рис. 5) могут распространяться как животными, так и ветром по типу перекатиполя. Последний способ распространения, вероятно, встречается и у частуховых с крупными, растопырено разветвленными соцветиями, в том числе у частухи обыкновенной. Плодики частуховых нередко находят в желудках рыб и других животных, что свидетельствует о возможности эндозоохории. Такой способ распространения особенно вероятен для костянкообразных плодиков кальдезии.
Многие частуховые размножаются также вегетативно с помощью стелющихся и укореняющихся в узлах надземных побегов (например, у раналисмы длинноносиковой, рис. 5, 1) или ползучих подземных побегов, заканчивающихся клубенькообразными зимующими почками (у многих видов стрелолиста). У некоторых видов кальдезии и эхинодоруса в соцветиях вместо цветков образуются вегетативные почки, дающие начало молодым растениям после того, как соцветия ложатся на влажную почву. Европейская калъдезия белозоролистная (С. parnassifolia) размножается преимущественно таким способом, так как вполне развитые плоды образуются у нее довольно редко.
Еще в 1827 г. бельгийский ботаник Дюмортье установил в пределах семейства частуховых 2 основные трибы: собственно частуховых (Alismeae) с мутовчато расположенными плодиками и стрелолистовых (Sagittarieae) со спирально расположенными плодиками на сильно выпуклом цветоложе. К первой трибе принадлежит большинство родов семейства, из которых в умеренно теплых областях северного полушария наиболее распространен род частуха с 12 видами. Из других родов этой трибы наиболее обособлены роды звездоплодник со срастающимися при плодах у своего основания длиннозаостренными плодиками, содержащими более чем один семязачаток, и лурониум с плавающими в воде облиственными соцветиями.
К трибе стрелолистовых принадлежат наиболее крупные (каждый с 25-30 видами) роды семейства: эхинодорус и стрелолист. Первый из них распространен в тропической, а отчасти и в субтропической Америке, замещаясь в Старом Свете прежде присоединявшимися к нему родами бальделлия (в Европе и Северной Африке) с 2 видами и раналисма (в Южной Азии и Африке) тоже с 2 видами. Большинство видов стрелолиста встречается в Северной Америке, но 3 вида - стрелолист обыкновенный (S. sagittifolia), стрелолист плавающий (S. natans) и стрелолист трехлистный (S. trifolia) - широко распространены в Евразии, в том числе и на территории СССР. Еще несколько видов стрелолиста, образующих особый подрод лофотокарпус (Lophotocarpus), распространены в тропических странах обоих полушарий и нередко выделяются в самостоятельный род (сюда принадлежит стрелолист цепкоплодный, рис. 3, 8).
Проведенное в последнее время исследование пыльцы всех родов семейства частуховых в основном подтвердило деление его на 2 трибы, однако бальделлия по строению пыльцевых зерен оказалась более близкой к родам трибы частуховых, а кальдезия по строению пыльцы (а также плодиков) заслуживает выделения в самостоятельную, монотипную трибу.
Один из видов стрелолиста - стрелолист трехлистный - довольно широко культивируется в Китае, Японии и некоторых других азиатских странах в качестве овощного растения ради съедобных клубней и добываемого из них крахмала. Культивируемые разновидности этого вида имеют особенно крупные клубни и более широкие листья по сравнению с дикорастущими. Следует отметить, что многие частуховые, как внетропические, так и тропические, принадлежат к числу очень редких и, по-видимому, быстро вымирающих растений. Таковы европейские виды кальдезия белозоролистная и лурониум плавающий, сохранившиеся лишь в немногих изолированных местонахождениях.
Семейство водокрасовые (Hydrocharitaceae)
К семейству водокрасовых принадлежат такие широко распространенные и, наверное, известные многим читателям водные растения, как водокрас обыкновенный (Hydrocharis morsus-ranае, рис. 6, 1) с розетками плавающих листьев, похожих на листья мелкой кувшинки, и телорез обыкновенный (Stratiotes aloides, рис. 7, 1) с розетками наполовину погруженных в воду жестких линейно-ланцетных листьев с пильчатозубчатым краем, о который легко порезаться (отсюда происходит название этого растения). Оба эти вида имеют относительно крупные цветки с 3 белыми лепестками. Еще одно водокрасовое стало обычным растением водоемов почти во всей Европе - это североамериканская элодея канадская (Elodea canadensis, рис. 7, 2). Ее прозвали "водяной чумой" за способность, размножаясь вегетативно (в Европу были завезены только женские особи этого вида, не приносящие плодов), быстро заполнять водоемы, вытесняя из них другие водные растения. В Европе элодея впервые отмечена в 1836 г. в Ирландии, затем она заселила водоемы Англии, а к 1860 г. распространилась не только в другие страны Западной Европы, но и в страны южного полушария - Австралию и Новую Зеландию.
Семейство водокрасовых включает 16 родов и около 120 видов, распространенных почти повсюду, где есть водоемы, исключая Арктику, север таежной зоны Евразии и высокогорья. Если не считать занесенные на другие континенты и натурализовавшиеся там виды, подобные элодее канадской, только 2 рода - оттелия (Ottelia) и валлиснерия (Vallisneria) - встречаются как в восточном, так и в западном полушарии. Четыре рода - элодея, эгерия (Egeria), лимнобиум (Limnobium) и гидромистрия (Hydromystria) - содержат только американские виды, а остальные пресноводные роды семейства принадлежат Старому Свету. Три рода водокрасовых - энгалус (Enhalus), талассия (Thalassia) и солелюбка (Halophila) - относятся к так называемым "морским травам" - цветковым растениям, приспособившимся к жизни в соленой воде морей и океанов. Морские водокрасовые распространены преимущественно в тропических областях обоих полушарий, где температура воды обычно не опускается ниже + 20 °С.
За пределы тропиков выходит только солелюбка овалънолистная (Н. ovalis, рис. 8, 1), которая в Японии распространена до изотермы в + 10 °С, но не дает там цветков и плодов. Этот последний вид принадлежит к числу немногих "морских трав", которые могут расти не только близ побережий морей и океанов, но и вдали от них. Так, во время 24-го рейса научно-исследовательского судна "Академик Вернадский" в августе 1981 г. цветущие и плодоносящие особи солелюбки вместе с корневищами и листовыми розетками талассодендрона реснитчатого (из семейства цимодоцеевых) были подняты драгой в большом количестве с глубины 10-15 м на банке Сая-де-Малья в районе подводного Маскаренского хребта среди Индийского океана.
Водокрасовые - многолетние, реже однолетние (некоторые виды оттелии и бликсы - Blyxa) растения, частично или полностью погруженные в воду. По внешнему облику они так разнообразны, что трудно поверить в принадлежность их к одному и тому же семейству. Относительно примитивными в отношении жизненной формы можно считать многолетние виды оттелии с очень коротким утолщенным вертикальным корневищем, закрепленным на дне водоема и несущим розетку прикорневых листьев. Некоторые виды этого рода имеют только погруженные в. воду, обычно линейные или ланцетные листья; другим свойственна разнолистность: первыег подводные листья у них ланцетные или линейные, не дифференцированные на черешок и пластинку, а следующие за ними надводные или плавающие в воде листья разделены на довольна длинный черешок и эллиптическую или яйцевидную пластинку (например, у оттелии частуховидной - Ottelia alismoides, рис. 6, 4). У также розеткообразующей валлиснерии имеются только подводные линейные листья. Такие же листья у "морских трав" - энгалуса и талассии, но у них они расположены пучками на верхушках коротких вертикальных побегов, отходящих от длинного ползучего корневища (рис. 8, 6). Третий род морских водокрасовых - солелюбка также имеет длинные ползучие корневища, но листья у видов этого рода эллиптические и расположенные на укороченных побегах различным образом. У солелюбки овальнолистной (рис. 8, 1) длинночерешковые листья расположены по 2 на каждом из сильно укороченных боковых побегов корневища; у солелюбки мелкоколючей (Halophila spinnlosa) многочисленные сидячие листья расположены двумя рядами на относительно длинных побегах; у солелюбки Энгелъманна (Н. engelmannii) сидячие листья: расположены пальчато по 5-7 на верхушках также довольно длинных прямостоячих побегов. Жилкование листьев у всех видов солелюбки производит впечатление перистого, так как 2 дуговидные прикраевые жилки соединены со срединной жилкой сильно развитыми анастомозами.
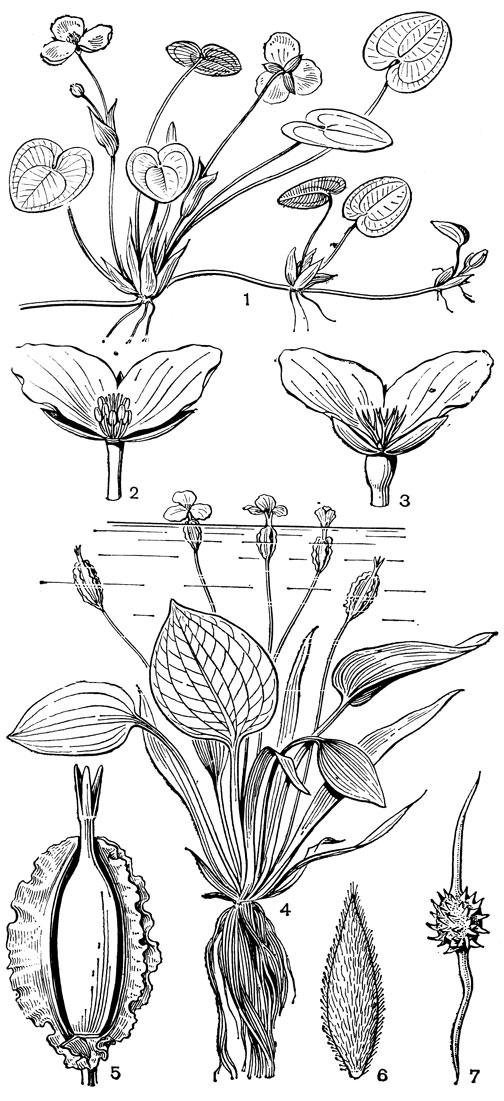
Рис. 6. Водокрасовые. Водокрас обыкновенный (Hydrocharis morsusranae): 1 - общий вид; 2 - мужской цветок с удаленным спереди лепестком; 3 - женский цветок с удаленным спереди лепестком. Оттелия частуховидная (Ottelia alismoides): 4 - общий вид; 5 - плод со вскрытым спереди покрывалом; 6 - семя. Бликса колючесемянная (Blyxa echinosperma): 7 - семя
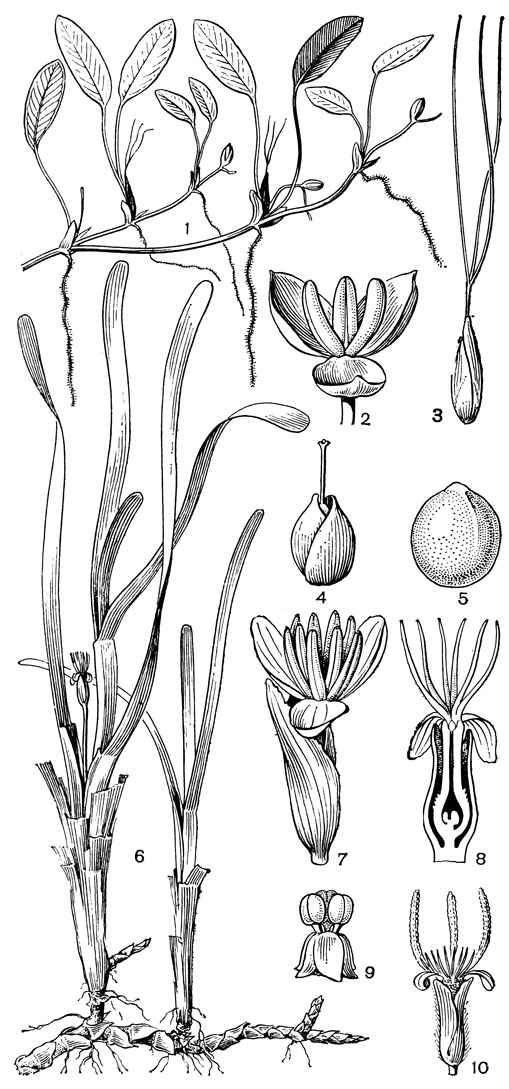
Рис. 8. Морские водокрасовые. Солелюбка овальнолистная (Halophila ovalis): 1 - общий вид; 2 - мужской цветок; 3 - женский цветок с покрывалом; 4 - плод с покрывалом; 5 - семя. Талассия черепаховая (Thalassia testudinum): 6 - общий вид; 7 - мужской цветок с покрывалом; 8 - женский цветок с покрывалом. Энгалус аировидный (Enhalus асоroides): 9 - свободноплавающий мужской цветок; 10 - женский цветок с покрывалом
У упомянутых выше водокраса и телореза розеткообразующие особи свободно плавают у поверхности воды, причем сердцевидные пластинки длинночерешковых листьев водокраса расположены на ее поверхности, а у телореза не дифференцированные на черешок и пластинку листья наполовину погружены в воду. Совершенно другой облик имеют водокрасовые с длинными, разветвленными и густо облиственными стеблями, которые могут быть закрепленными на дне водоемов, но легко обрываются или обламываются, продолжая свое развитие в свободно плавающем состоянии. На мелководьях плавающие побеги могут снова прикрепляться к дну водоема с помощью придаточных корней. Примерами таких водокрасовых служат элодея и гидрилла (Нуdrilla), мелкие сидячие линейно-ланцетные листья которых имеют всего одну жилку и расположены на стебле очередно, супротивно или, чаще, ложными мутовками по 3-5.
У водокрасовых с плавающими на поверхности воды листовыми пластинками, например у водокраса, они обычно цельнокрайние. У родов с мелкими расставленными листьями листовые пластинки более или менее зубчатые; зубчики часто имеются и в верхней части подводных лентовидных листьев валлиснерии и энгалуса. Выше уже говорилось об остропильчатых по краю листьях телореза. Напоминают в этом отношении телорез некоторые виды оттелии с погруженными в воду листьями, а у оттелии колючей (Ottelia muricata) шиповидные зубцы имеются не только по краю листа, но и на его поверхности вдоль жилок. У многих водокрасовых листья близ основания влагалищеобразно расширены, а в пазухах их имеются мелкие внутривлагалищные чешуйки с выделяющими слизь железками. Водокрасовые полностью лишены сосудов, их нет даже в корнях.
Строение цветков водокрасовых соответствует двум основным направлениям их эволюции в пределах семейства: от обоеполых цветков к однополым и двудомным и от энтомофильных к анемофильным и гидрофильным. Цветоносы, несущие одиночные цветки или верхоцветные соцветия полузонтики, возникают как боковые побеги из пазух расставленных или собранных в розетку листьев. У представителей всех родов семейства на этих цветоносах имеется так называемое покрывало, образованное двумя (редко одним) свободными или сросшимися друг с другом прицветниками и служащее защитой для развивающихся в них соцветий и одиночных цветков, а у некоторых родов и для плодов. Покрывало чаще бывает перепончатым, но может быть и травянистым, а у некоторых видов оттелии, в том числе оттелии частуховидной (рис. 6, 5), оно несет до 10 продольных крыловидных выростов.
Цветки водокрасовых почти всегда актиноморфные и трехчленные, обычно с околоцветником из 6 сегментов, которые часто дифференцированы на 3 наружных, более мелких и травянистых сегмента, образующих чашечку, и 3 внутренних, более крупных и окрашенных в белый, реже розовый или желтый цвет сегмента, образующих венчик. Лишь у валлиснерии и бликсы цветки могут быть зигоморфными за счет редукции их частей (например, одной из трех тычинок в мужских цветках валлиснерии). Большинство родов семейства имеет однополые и часто двудомные цветки. Обоеполые цветки встречаются у всех видов оттелии, а также у многих видов бликсы и некоторых видов элодеи, в том числе у элодеи гренадской (Elodea granatensis, рис. 7, 6). Почти одинаковую величину и строение членов околоцветника имеют цветки гидриллыг элодеи и лагаросифона (Lagarosiphon), обычно мелкие и малозаметные, что связано с переходом к гидроанемофилии. У представителей другого гидроанемофильного рода - валлиснерии - лепестки редуцированы до очень мелких чешуек, а у солелюбки, цветки которой опыляются под водой, лепестки редуцируются полностью.
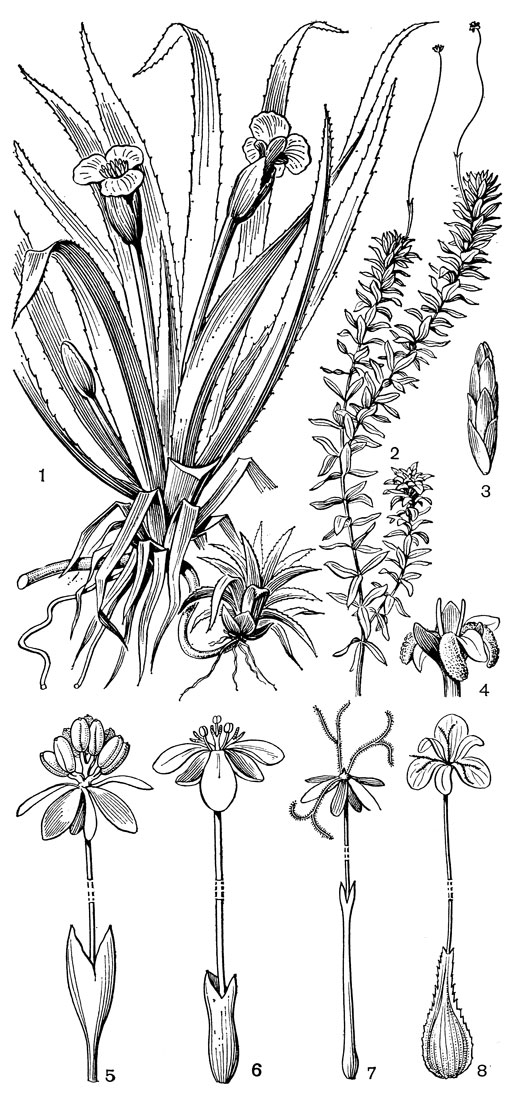
Рис. 7. Водокрасовые. Телорез обыкновенный (Stratiotes aloides): 1 - общий вид. Элодея канадская (Elodea canadensis): 2 - часть побега с женским цветком; 3 - зимующая почка; 4 - женский цветок; 5 - мужской цветок с покрывалом. Элодея гренадская (Е. granatensis): 6 - обоеполый цветок с покрывалом. Элодея болотниковидная (Е. callitrichoides): 7 - женский цветок с покрывалом. Лагаросифон большой (Lagarosiphon major): 8 - женский цветок с покрывалом
Андроцей в цветках водокрасовых обычно представлен 1-5, редко (у телореза) более чем 5, не всегда отчетливыми трехчленными кругами тычинок. В мужских и обоеполых цветках представителей энтомофильных родов один или несколько кругов тычинок видоизменены в стаминодии, несущие выделяющие нектар железки и, таким образом, функционирующие как нектарники. У телореза 15-30 стаминодиев находятся между лепестками и фертильными тычинками, а у водокраса и близких родов в стаминодии видоизменены 1-2 круга самых внутренних тычинок. Стаминодии сохраняются и в женских цветках тех водокрасовых, которые имеют их в мужских цветках, причем у водокраса они прирастают к плодолистикам и производят впечатление выростов гинецея. У гидроанемофильных и гидрофильных водокрасовых как стаминодии, так и фертильные тычинки имеют тенденцию к редукции. Так, у солелюбки только 3 фертильные тычинки, а в очень мелких мужских цветках валлиснерии число их уменьшается до двух (третья превращается в стаминодий). Наконец, у близкого к валлиснерии рода майдения (Maidenia) в мужских цветках всего только одна фертильная тычинка. У большинства водокрасовых пыльники расположены на довольно длинных тычиночных нитях, но у представителей морских родов с гидрофильным способом опыления они сидячие или почти сидячие. Строение пыльников также различно у разных родов. Обычно они имеют 2 двугнездные теки, но у телореза и оттелии - 2 одногнездные теки, а у водокраса - только одну одногнездную теку. У лагаросифона и валлиснерии пыльники очень мелкие с относительно крупными и немногочисленными пыльцевыми зернами. У всех пресноводных водокрасовых и энгалуса пыльцевые зерна шаровидные, обычно с покрытой шипиками наружной оболочкой - экзиной, а у морских родов талассия и солелюбка пыльцевые зерна имеют гладкую оболочку и соединены клейкой массой в цепочкообразные нити. Кроме того, пыльцевые зерна солелюбки имеют почковидную форму и полностью лишены экзины.
Как в обоеполых, так и в женских цветках водокрасовых гинецей состоит из (2)3-6, редко до 15 плодолистиков с многочисленными, очень варьирующими по расположению у разных родов семязачатками. Интересно, что, несмотря на всегда нижнюю и одногнездную завязь с неполными перегородками, гинецей водокрасовых сохраняет апокарпность, так как плодолистики у большинства родов не срастаются друг с другом. Особенно близок к апокарпному гинецей телореза и энгалуса, в то время как у лимнобиума и некоторых других родов он скорее синкарпный, чем апокарпный. Наименьшее число плодолистиков - 3 - у валлиснерии, бликсы, гидриллы и других мелколистных родов; у солелюбки число плодолистиков может варьировать от 2 до 5. Наибольшее число (9-15) плодолистиков известно только у нескольких видов оттелии, нередко выделяемых в самостоятельный род буттия (Boottia). Столбик на верхушке завязи развит только у валлиснерии и бликсы; у представителей других родов ветви рыльца сидячие или почти сидячие, обычно двураздельные и покрытые сосочками. Цельные рыльцевые ветви имеют только бликса и солелюбка, причем у солелюбки они совсем не похожи на рыльцевые ветви других водокрасовых: длинные, нитевидные, лишенные сосочков, но бороздчатые на всей внутренней стороне, подобно рыльцевым ветвям некоторых "морских трав" из других семейств. Следует отметить, что женские цветки однодомных и двудомных водокрасовых почти всегда бывают одиночными, но у одних родов (например, у оттелии) они сидячие и плод развивается внутри покрывала, а у других родов они во время цветения выступают на более или менее длинных цветоножках из покрывала.
Многосемянные плоды водокрасовых, имеющие мясистый околоплодник, обычно не вскрываются, и семена освобождаются в результате постепенного разрушения и ослизнения околоплодника. Однако в литературе отмечается, что плоды телореза и водокраса разрываются под давлением разбухающей желатинообразной массы, выделяемой оболочками семян. Относительно немногосемянные плоды гидриллы по форме напоминают стручки некоторых крестоцветных. В зрелом состоянии они обычно имеют перетяжки между семенами, расположенными здесь в один ряд. Наименьшее количество семян (нередко только по одному в каждом из трех плодолистиков) развивается у солелюбки, плоды которой отличаются от плодов других водокрасовых также своей тонкостенностью.
Выше уже отмечалось большое разнообразие жизненных форм водокрасовых, хотя все они принадлежат к настоящим водным растениям. Пресноводные водокрасовые прежде всего делятся на виды с длинными стеблями и очень мелкими расставленными листьями (например, элодея канадская, рис. 7, 2) и виды розеткообразующие с безлистными ножками часто одноцветковых соцветий. Последние можно разделить еще на закрепленные на дне водоемов (например, оттелия частуховидная, рис. 6, 4) и свободно плавающие у поверхности воды (например, водокрас обыкновенный, рис. 6, 1, телорез обыкновенный, рис. 7, 1). Морские водокрасовые отличаются мощно развитой системой корневищ, которые у талассии черепаховой (Thalassia testndinum) составляют 75-90% всей массы растения, и часто лентовидными листьями, собранными пучками на коротких прямостоячих побегах, отходящих от узлов корневища. Для обитающего в тропических морях Старого Света энгалуса аировидного (Enchalus acoroides) отмечается очень быстрый рост основания листьев - 1,1-2,2 см в сутки. Это рассматривается как приспособление против обрастания листьев поселяющимися на их поверхности организмами, значительно снижающими их фотосинтетическую активность. Длина листьев этого растения достигает 60 см при ширине 1-1,5 см.
По характеру приспособления к разным способам опыления водокрасовые также могут быть разделены на ряд групп. К первой из них принадлежат наиболее примитивные по строению цветка энтомофильные виды водокраса, телореза, оттелии, лимнобиума, эгерии, венчики которых имеют относительно крупные (диаметром 1 - 5 см) размеры и ярко окрашены. Самоопылению у них часто препятствует двудомность, наиболее выраженная у телореза и эгерии. У водокраса и лимнобиума мужские и женские цветки нередко встречаются на одной и той же особи, но почти всегда на разных розетках. Следует отметить, что у двудомных водокрасовых особи разного пола часто распространены очень неравномерно, следствием чего является отсутствие плодоносящих растений в значительных по площади частях ареала этих видов. Так, элодея канадская представлена в Европе только женскими особями, а у телореза обыкновенного в северной части ареала встречаются также только женские экземпляры, не приносящие плодов. У эгерии густолистной (Egeria densa), напротив, женские экземпляры встречаются крайне редко. Опылителями энтомофильных водокрасовых, по-видимому, являются мелкие перепончатокрылые, мухи, жуки и другие насекомые со жвалами и короткими хоботками. У некоторых видов оттелии наряду с энтомофильными, вполне развитыми цветками отмечено присутствие клейстогамных цветков. Так, у культивировавшейся в Швейцарии оттелии овалънолистной (Ottelia ovalis) из Новой Каледонии летом регулярно образовывались энтомофильные цветки, а весной и осенью - мелкие, погруженные в воду клейстогамные цветки в пазухах листовых влагалищ. Такие клейстогамные цветки имели карликовые, но нормально функционирующие тычинки и рыльца внутри сомкнутых листочков околоцветника. Пыльники в этих цветках не вскрывались обычным способом, а их стенка разрушалась там, где она соприкасалась с рыльцевой ветвью.
Анемофильная группа водокрасовых представлена лишь американским тропическим родом гидромистрия, иногда присоединяемым к роду лимнобиум. Женские цветки гидромистрии имеют длинные и покрытые длинными сосочками рыльцевые ветви, возвышающиеся над поверхностью воды, как и мужские цветки. Имеется указание, что опыление может произойти и на поверхности воды: завядшие рыльцевые ветви опускаются на поверхность воды, где может находиться и пыльца.
Большая группа водокрасовых имеет очень оригинальный способ опыления, переходный от анемофилии к гидрофилии,- гидроанемофилию, когда опыление происходит с помощью ветра, но на поверхности воды или близ нее. Гидроанемофилия более простого типа известна у относительно богатого видами рода элодея. Как мужские, так и женские цветки, а у 3 видов подрода апаланта (Apalanthe) - обоеполые цветки образуются здесь в пазухах листьев по одному в урновидном или трубчатом покрывале. На поверхность воды они выносятся быстро и сильно удлиняющейся верхушкой завязи - гипантием, а у мужских цветков - разрастающейся цветоножкой. У видов с обоеполыми цветками (например, у элодеи гренадской, рис. 7, 6) гипантии достигают в длину 1,5-6 см, у большинства видов с однополыми цветками - 15 см, а у некоторых видов, например у элодеи болотниковидной (Elodea callitrichoides, рис. 7, 7),- 25-30 см. Женские цветки элодеи обычно погружены в воду, на поверхности которой остаются рыльцевые ветви с водоотталкивающей поверхностью. Пыльники мужских цветков возвышаются над водой и вскрываются взрывчато, рассеивая на поверхность воды пыльцевые зерна. Последние, как и рыльца, не смачиваются водой, благодаря тому что между шипиками экзины задерживается воздух. Плавая по поверхности воды, пыльцевые зерна приходят в соприкосновение с рыльцевыми ветвями женских цветков.
Уже у одного из видов элодеи - элодеи Нутталла (Е. nuttallii) - мужские цветки в фазе бутонов обрывают относительно короткие цветоножки, всплывают на поверхность воды и здесь расцветают, высыпая пыльцу. Результатом дальнейшей специализации к гидроанемофилии является замечательный способ опыления, свойственный таким водокрасовым, как валлиснерия, гидрилла, лагаросифон, майдения, нехамандра (Nechamandra) и энгалус, и уже описанный на примере валлиснерии в первой части пятого тома "Жизни растений" (рис. 40). Обычно образующиеся в большом количестве очень мелкие мужские цветки в фазе бутонов отделяются от материнского растения и всплывают на поверхность воды. Затем сегменты околоцветника отгибаются в стороны и цветки, подобно крошечным корабликам, свободно плавают на поверхности воды. Важно отметить, что, в отличие от элодеи Нуттала, пыльца здесь не выпадает из пыльников, оставаясь в виде соединенного с ними комочка, а нити тычинок располагаются под острым углом к поверхности воды. Опыление происходит путем случайного соприкосновения комочка пыльцы с возвышающимися над водой рыльцевыми ветвями женских цветков, т. е. не на поверхности воды, как у элодеи, а в воздухе. Попавшая в воду пыльца быстро погибает. Женские цветки выносятся на поверхность воды различными способами. У валлиснерии сильно удлиняется цветоножка (она растет со скоростью до 2 см в час), после цветения она спирально скручивается и погружает развивающийся плод в воду. У гидриллы, подобно элодее, удлиняется гипантий, который внешне напоминает цветоножку, но в действительности находится между околоцветником и расположенной в пазухе листа сидячей завязью. У энгалуса, напротив, удлиняется ножка одноцветкового соцветия и цветок выносится на поверхность воды, одетый в покрывало. Различное строение имеют и мужские цветки у разных родов. Так, у лагаросифона все 6 сегментов околоцветника отгибаются книзу, образуя "кораблик", а роль паруса играют 3 крупных, направленных вверх стаминодия. У валлиснерии 2 сегмента околоцветника образуют "кораблик", а третий, более маленький сегмент приподнят над водой и служит парусом.
Наиболее высокоспециализированный способ опыления - настоящую гидрофилию - имеют 2 морских рода водокрасовых - талассия и солелюбка. Важным приспособлением к этому способу опыления являются здесь склеенные в цепочкообразные нити пыльцевые зерна, которые в таком виде, естественно, имеют больше шансов зацепиться в воде за длинные рыльцевые ветви.
У большинства водокрасовых покрытые клейкой массой семена могут распространяться как потоками воды, так и экзозоохорно - с помощью водных животных. У талассии и солелюбки семена обычно падают на дно сразу после освобождения из плода, но все же могут распространяться на более далекие расстояния морскими течениями или во время штормов. У энгалуса семена плавают по поверхности воды до разрушения семенной оболочки, после чего тяжелый зародыш падает на дно и прорастает. У бликсы колючесемянной (Blyxa echinosperma) семена имеют специальные шипообразные выросты для заякоривания в шерсти или перьях животных (рис. 6, 7).
У большинства водокрасовых возможности полового размножения очень ограничены. Многие виды этого семейства редко цветут и еще реже приносят плоды. Даже водокрас обыкновенный, несмотря на обильное ежегодное цветение, очень редко плодоносит. У широко, но рассеянно распространенной в СССР гидриллы мутовчатой (Hydrilla verticillata) только недавно Н. С. Пробатовой были обнаружены цветки и плоды. Отсюда понятно, что большинство водокрасовых распространяются главным образом вегетативно. У элодеи, гидриллы и других родов с длинными ветвистыми стеблями каждая часть растения, несущая почку или хотя бы один узел, способна регенерировать в новую особь. Кроме того, внетропические виды этих родов осенью образуют большое количество зимующих почек, также дающих начало новым особям. Именно за счет такого интенсивного вегетативного размножения занесенная в Европу элодея канадская стала "водяной чумой". Водокрас, телорез, валлиснерия и многие другие розеткообразующие водокрасовые образуют столонообразные пазушные побеги, несущие на верхушке вегетативную почку, которая, еще не теряя связи с материнским растением, быстро развивается в молодую особь. Морские водокрасовые легко размножаются путем переноса частей корневищ или облиственных побегов с обломками корневищ водой, особенно во время штормов.
Водокрасовые делят на 4 неравных по объему подсемейства: собственно водокрасовые (Hydrocharitoideae), валлиснериевые (Vallisnerioideae), талассиевые (Thalassioideae) и солелюбковые (Halophiloideae). К первому из них принадлежат наиболее примитивные по строению цветков, часто энтомофильные роды, обычно с разделенным на чашечку и венчик околоцветником, с покрывалом из 1-2 свободных прицветников и гинецеем из 6 и больше плодолистиков. Подсемейство собственно водокрасовых, в свою очередь, делится на 4 трибы: оттелиевые (Ottelieае), водокрасовые (Hydrochariteae), телорезовые (Stratioteae) и энгалусовые (Enhaleae). В трибу оттелиевых входит наиболее крупный род семейства - оттелия, 40 видов которого распространены главным образом в тропических и субтропических областях обоих полушарий, но один вид - оттелия частуховидная (рис. 6, 4) доходит в Восточной Азии до южной части советского Дальнего Востока. Среди представителей этого рода, для которого очень характерно травянистое, плотно охватывающее плод покрывало, имеются виды с обоеполыми, однодомными и двудомными цветками, с плавающими на поверхности воды и полностью погруженными в воду листьями, многолетники и однолетники. К трибе водокрасовых, кроме широко распространенного водокраса (рис. 6, 2), принадлежат еще небольшие американские роды лимнобиум и гидромистрия, из которых последний имеет опыляющиеся ветром цветки. Все представители этой трибы - плавающие у поверхности воды розеткообразующие растения. Единственный вид трибы телорезовых - телорез обыкновенный (рис. 7, 1) довольно широко распространен во внетропических областях Евразии. Быстро разрастаясь, его розетки часто полностью занимают поверхность водоемов, которые производят в этом случае обманчивое впечатление зеленых "лужаек", через которые не только нельзя пройти, но и трудно проплыть на лодке. Триба энгалусовых тоже состоит из одного вида - энгалуса аировидного (Е. acoroides), распространенного в тропической части Индийского океана. По строению вегетативных органов энгалус сходен с "морскими травами" из других семейства по способу опыления - с валлиснерией, занимая промежуточное положение между подсемействами.
Подсемейство валлиснериевых, для которых характерно присутствие только 3 плодолистиков и покрывала из 2 сросшихся друг с другом прицветников, а также некоторая редукция околоцветника в связи с приспособлением к гидроанемофилии, представлено 3 трибами: гидрилловые (Hydrilleae), валлиснериевые (Vallisnerieae) и бликсовые (Blyxeae). К трибе гидрилловых принадлежат мелколистные водокрасовые с длинными разветвленными стеблями.
Наиболее крупный род - элодея включает 18 американских видов, из которых элодея канадская (рис. 7, 2) в настоящее время распространена также в Евразии и Австралии. Внешне очень похожие на элодею представители рода эгерия имеют, однако, энтомофильные цветки. Широко известным аквариумным растением является валлиснерия спиральная (Vallisneria spiralis), обитающая в южной половине Евразии и в Северной Африке. В СССР особенно большие заросли она образует в мелководьях северной части Каспийского моря близ устья Волги. Кроме валлиснерии, к трибе валлиснериевых принадлежит еще западноавстралийский род майдения, замечательный еще более мелкими, чем у валлиснерии, свободноплавающими мужскими цветками с одной вполне развитой тычинкой. Трибу бликсовых составляет всего один, но очень полиморфный род бликса с 10 видами, распространенными в Южной и Юго-Восточной Азии, Африке и Австралии. В его пределах есть виды с расставленными и собранными в розетку листьями, с обоеполыми и однополыми цветками.
Подсемейство талассиевых представлено лишь одним родом талассия с 2 видами: талассией черепаховой (рис. 8, 6), распространенной в Карибском море, и талассией Хемприха (Таlassia hemprichii), распространенной в Индийском океане и западной части Тихого океана. Это морские длиннокорневищные травы, имеющие двудомные цветки со слабо развитым околоцветником из 6 почти одинаковых сегментов и гинецеем из 6-12 плодолистиков. Цветки опыляются под водой, и пыльцевые зерна склеены в цепочковидные нити.
Наиболее высокоспециализированное подсемейство солелюбковых включает только один пантропический род солелюбка с 5 видами. Подобно другим "морским травам", это длиннокорневищные многолетники, однако листья у них не линейные, а обычно эллиптические, у солелюбки овальнолистной (рис. 8, 1) с довольно длинным черешком. Мужские цветки солелюбки расположены на коротких цветоножках и состоят из 3 сегментов околоцветника и 3 тычинок с сидячими пыльниками. Женские цветки сидячие и окруженные покрывалом из 2 перепончатых прицветников. Яйцевидная завязь несет на верхушке рудименты 3 сегментов околоцветника и 2-5 (чаще 3) длинных нитевидных рыльцевых ветви. Пыльцевые зерна солелюбки также склеены в цепочки.
Многие водокрасовые используются или могут быть использованы в качестве аквариумных растений (виды элодеи, валлиснерии, эгерии, лимнобиума и др.). Листья оттелии частуховидной идут на салат, а плоды этого растения, как и плоды энгалуса аировидного, употребляют в пищу в печеном или вареном виде. Некоторые виды оттелии применяют в народной медицине. Из волокон листьев энгалуса можно изготовлять грубую ткань. Некоторые водокрасовые, особенно элодея канадская и телорез обыкновенный, способны полностью заполнять водоемы, мешая речному судоходству, однако они могут быть использованы в качестве хорошего удобрения для полей.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'