Класс семенные папоротники, или лигиноптеридопсиды (Lyginopteridopsida, или pteridospermae) (Н. С. Снигиревская)
Изучая палеозойские и раннемезозойские растения, находимые в земных слоях в виде отпечатков и окаменелостей, палеоботаники нередко встречают ископаемые листья, по строению неотличимые от перистых листьев настоящих папоротников, но несущие семена на своих верхушках или по бокам жилок.
Такое удивительное сочетание признаков семенных растений и папоротников, впервые установленное в 1904 г. выдающимися английскими палеоботаниками Ф. Оливером и Д. Скоттом, и дало повод назвать эту группу растений семенными папоротниками (Pteridospermae).
Это была обширная группа растений, из которой к настоящему времени описано несколько сот видов, объединяемых в четыре основных порядка - лигиноптерисовые (Lyginopteridales), медуллозовые (Medullosales), кейтониевые (Caytoniales) и глоссоптерисовые (Glossopteridales) и около 10 семейств.
Внешний вид этих вымерших растений наиболее удачно передает реконструкция медуллозы Ноэ (Medullosa noei), выполненная американскими палеоботаниками В. Стюартом и Т. Делевориасом по отпечаткам листьев и срезам окаменелых стеблевых частей (рис. 159).
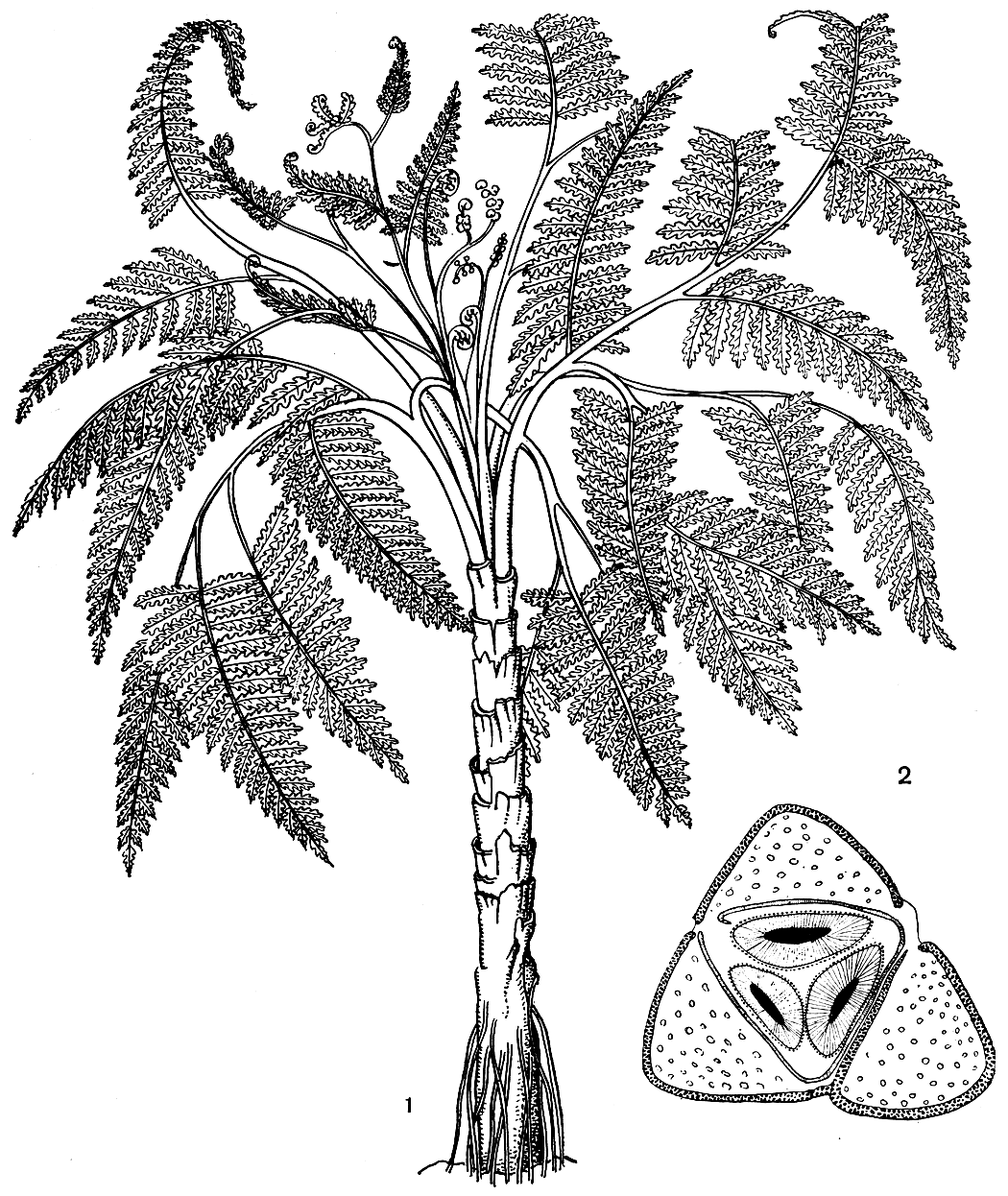
Рис. 159. Медуллоза Ноэ (Medullosa noei): 1 - общий вид, реконструкция; 2 - схема поперечного среза через стебель с тремя меристелами
Семенные папоротники - полностью вымершие растения. Они существовали с позднего девона до раннего мела. Наиболее часто их остатки встречаются в отложениях карбона и перми. Одни из них были распространены по всему земному шару, другие, как, например, ангароптеридиум (Angaropteridium), были свойственны только ангарским флорам, а глоссоптерисовые и користоспермовые, напротив, приурочены главным образом к районам, соответствующим древнему материку Гондване, который существовал с середины карбона до конца триаса.
Наиболее распространено мнение, что семенные папоротники карбона были обитателями более возвышенных участков, обрамлявших приморские заболоченные леса. Однако все больше данных за то, что, например, заключительные этапы существования формации карбонового низинного заболоченного леса, как правило, сопровождались пышным развитием семенных папоротников или кордаитовых.
Семенные папоротники были похожи на настоящие папоротники не только по строению перистых листьев, но и по внешнему облику растений: древовидных, лиановидных и травянистых.
Стебли древовидных семенных папоротников были или прямыми, или дихо- и моноподиально ветвящимися. У одних листья сидели в очередном порядке на радиально-симметричном стебле, у других были сближены на верхушке и располагались в виде кроны, как у многих современных древовидных папоротников. Поверхность стебля многих растений была покрыта волосками, а иногда особыми железками, как, например, железками булавовидной формы у калиматотеки (Calymmatotheca), которые выполняли, по-видимому, секреторную функцию. У одних растений стволы были более или менее гладкие, с рубцами от опавших листьев; у других покрыты основаниями опавших листьев, как у медуллозы. У лигиноптерисовых (Lyginopteridaceae) стебли были диаметром 2-4 см, а у медуллозовых (Medullosaceae) их диаметр достигал 20 см. У некоторых семенных папоротников на стебле между листьями и в узлах располагались воздушные корни. Стебли пельтаспермовых (Peltaspermaceae) и кейтониевых (Caytoniaceae) до сих пор не найдены, и обе группы известны только по находкам стерильных и фертильных листьев и семян.
Листорасположение у семенных папоротников было спиральным. Как у лигиноптерисовых, так и у медуллозовых листья были довольно крупными и в основании вильчато делились на две симметричные половины, каждая из которых была перисто рассечена. В почкосложении листья были улиткообразно закручены, как у современных и многих ископаемых папоротников.
У растений из порядка глоссоптерисовых листья сидели спирально или мутовчато (глоссоптерис) в утолщенных узлах тонких стеблей. Они были довольно крупными, но очень различными по форме - цельные, цельнокрайние, линейные, ланцетные, лопатчатые, яйцевидные, сидячие и на коротких черешках. Если у более древнего рода гангамоптерис (Gangamopteris) не было ясно выраженной средней жилки, то у глоссоптериса (Glossopteris) она была развита хорошо.
Размножение семенных папоротников происходило с помощью семян, в которых до сих пор не найдены зародыши. Отсутствие зародыша, констатированное при изучении многих сотен ископаемых семян этих растений, до сих пор представляется загадкой. Существует точка зрения, согласно которой семенные папоротники не имели настоящего семени, хотя у них был семязачаток. В связи с этим их, а также современные саговниковые и гинкговые причисляли не к семенным, а к так называемым семязачатковым растениям. Однако едва ли это решает вопрос. Несомненно, что зародыш должен был развиться до прорастания этого семязачатка. По-видимому, развитие зародыша у семенных папоротников происходило в семязачатках, уже опавших с материнского растения на почву, как это происходит нередко у ныне живущего грлосеменного гинкго двулопастного.
Семязачатки развивались на расчлененных папоротниковидных листьях, располагаясь или по краю, или на верхушке листа (рис. 160). У наиболее примитивных родов семязачатки сидели на концах боковых или верхушечных перышек (сегментов), у более подвинутых родов (например, у сфеноптериса тонкого - Sphenopteris tenuis) - на поверхности перышек. Иногда положение семязачатков было вторичноверхушечным за счет редукции и видоизменения перышек в результате их превращения в семяножку.
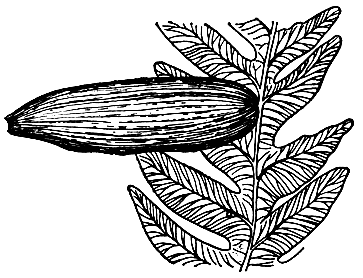
Рис. 160. Семенной папоротник алетоптерис Норина(Аlеthopteris norinii). Семязачаток, сидящий на листе папоротникового типа
У большинства семенных папоротников семязачатки были окружены так называемой купулой, т. е. особым покровом, имеющим у различных растений очень разное строение и форму, иногда чашевидную, конусовидную или дисковидную. Считается, что купула возникла из сегментов (долек) листа, окружающих семязачаток. Листовое происхождение купулы подтверждается ее строением у наиболее древних представителей группы.
Семязачатки были одиночными, как у лигиноптерисовых, или многочисленными, как у глоссоптерисовых. У лигиноптерисовых они достигали в длину 5,5 мм, у пельтаспермовых - 7 мм, а у користоспермовых - 3,5-7 мм.
Рассмотрим строение семязачатка на примере калиматотеки Хёнингхауса (Calymmatotheca hoeninghausii, рис. 161). Снаружи семязачаток окружен купулой, несущей железки. Из ножки купулы проводящий пучок переходит в ножку семязачатка, имеющего овальную форму. По строению проводящий пучок ножки сходен с пучком листового черешка. Наружный покров семязачатка (интегумент) возникает, по мнению ученых, в результате длительной эволюции из кольцеобразно расположенных спорангиев, которые, потеряв фертильность и срастаясь между собой, обеспечили защиту центральному, сохранившему фертильность мегаспорангию. Таким образом, семязачаток - это синангий. Подтверждение этому - большое количество примитивно устроенных семязачатков, у которых сохранились признаки синангиальной структуры. Нуцеллус, соответствующий мегаспорангию, обычно тесно спаян с интегументом, но иногда, оставаясь свободным в верхушечной области семязачатка, он образует так называемую колонку пыльцевой камеры, в которой нередко находят пыльцу, причем иногда сохранившуюся в проросшем состоянии. На этом основании сделано предположение, что пыльцевая камера была заполнена сахаристой жидкостью, которая являлась благоприятной средой для прорастания пыльцы. Центральная часть семязачатка занята большой функционирующей мегаспорой, от которой у семенных папоротников чаще сохраняется только оболочка, но нередко также ткань гаметофита с архегониями, в которых даже сохраняются яйцеклетки.
Мужские спороношения у растений этой группы - обычного папоротникового типа. Они представлены или краевыми микроспорангиями, как у лигиноптерисовых, или микросинангиями, иногда сложно устроенными, как у медуллозовых и кейтониевых. Пожалуй, наиболее интересны сложно устроенные и очень разнообразные по форме мужские спороношения медуллозовых.
Микроспоры семенных папоротников были разными по величине и строению. Например, у пельтаспермовых они были очень мелкими, диаметром всего 30 мкм, однобороздными, а у лигиноптерисовых и медуллозовых - диаметром до 400-500 мкм, и имели одну дистальную борозду наряду с трехлучевым тетрадным рубцом. Проросшие микроспоры этого типа были найдены в пыльцевых камерах некоторых семян, в частности, у пахитесты (Pachytesta). Предполагают, что гигантские микроспоры переносились на семязачатки, видимо, еще до полного развития мужского гаметофита, который завершал свое развитие в питательной жидко-сти пыльцевой камеры семязачатка.
Семенные папоротники - группа наиболее примитивная среди голосеменных растений. Одни ученые приходят к выводу, что они занимают промежуточное положение между настоящими папоротниками и голосеменными, в то время как другие считают эти группы возникшими и эволюционировавшими параллельно. Как уже сказано, листья семенных папоротников часто очень похожи на листья настоящих папоротников или даже неотличимы от них. Это представляет яркий пример параллельного развития гомологичных органов. Но наличие семян, анатомическое строение стебля и корня и способность семенных папоротников к вторичному росту достаточно резко отличают их от настоящих папоротников.
Анатомическое строение стеблей и корней семенных папоротников очень специфично. Так, стебель молодого растения калиматотеки был протостелическим, но с возрастом он становился сифоностелическим с хорошо развитой сердцевиной и гнездами склеренхимных клеток (рис. 161). Вокруг сердцевины располагались многочисленные мезархные пучки, их метаксилема состояла из крупных (в сечении) лестничных, сетчатых и точечных трахеид. Последние несли многорядные окаймленные поры на всех стенках. Как у большинства семенных папоротников, у калиматотеки была довольно мощная вторичная ксилема, состоящая также из точечных трахеид, но с порами только на радиальных стенках. Довольно широкие древесинные лучи, состоящие из паренхимных клеток, разбивали вторичную ксилему на секторы. Интересной особенностью внутреннего строения стебля этого семенного папоротника является структура наружной коры, которая состояла из сложного сетчатого переплетения тяжей склеренхимных клеток. На тангентальном срезе они имели вид ячеистой стенки, а на поперечном - вид радиально вытянутых полос, имеющих форму трабекул. Петли сетки были заполнены паренхимной тканью. Листовые следы имели двухпучковое строение. В основании листа эти два пучка срастались в F-образный (на поперечном срезе) пучок.
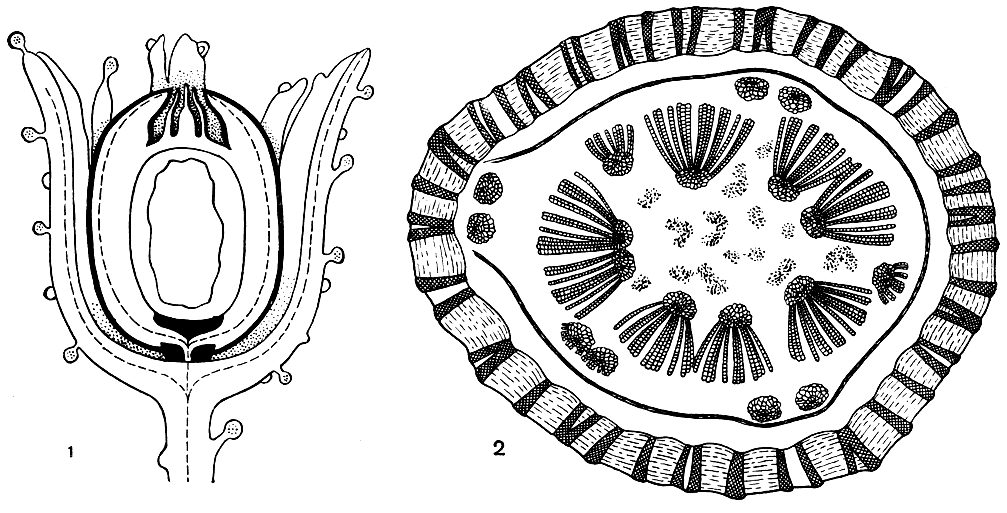
Рис. 161. Калиматотека Хёнингхауса (Calymmatotheca hoeninghausii): 1 - строение семязачатка; 2 - схема поперечного среза через стебель
У медуллозовых строение как стебля, так и листовых черешков было принципиально иным. Их стела рассечена на ряд элементарных стел (меристел), каждая из которых развивала свои вторичные проводящие ткани - ксилему и флоэму (рис. 159). Число меристел у видов медуллозы было различным. Хотя меристелы имеют вид самостоятельных проводящих систем, они анастомозируют друг с другом, соединяются в узлах, затем снова расходятся. Наружная кора у медуллозовых была обычно более просто устроена, чем у лигиноптерисовых, и состояла из массивной паренхимной ткани, в которой были рассеяны одиночные секреторные каналы, как у мараттиевых и саговниковых. Меристелы были погружены в основную ткань стебля, представленную паренхимой, в наружной части которой проходили многочисленные листовые следы. Листовые следы в своей нижней части близ стелы стебля имели концентрическое строение, а в наружной зоне коры и в листовом черешке были представлены многочисленными коллатеральными пучками. Этим они очень напоминают стебли однодольных растений.
Остатки семенных папоротников играют большую роль в сложении каменных углей СССР, Западной Европы и Северной Америки. Почти на каждом срезе через угольные почки, составляющие окаменевшие участки древнего торфяника, превращенного в уголь, можно найти стебли, корни, семена и пыльцу семенных папоротников.
Группа семенных папоротников взята на вооружение биостратиграфами, занимающимися вопросами расчленения палеозойских и мезозойских отложений. Например, по появлению отпечатков листьев растения каллиптерис (Callipteris) проводится нижняя граница пермского периода.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'