Происхождение голосеменных
Возникновению голосеменных растений предшествовало несколько важных эволюционных событий, которые произошли в течение девонского периода. В области репродуктивной сферы это было возникновение разноспоровости. Разноспоровые формы известны уже начиная со среднего девона (около 400 млн. лет назад). У шалёрии (Chaleuria cirrosa) из среднего девона Нью-Брансуика (Канада) одни спорангии содержат либо преимущественно крупные споры, либо преимущественно маленькие или смесь тех и других. В позднем девоне появляются растения с еще более ясно выраженной разноспоровостью. В области вегетативной сферы это было возникновение камбия и появление древесных форм. Хорошо сохранившаяся вторичная ксилема, свидетельствующая о камбиальной активности, также известна уже из отложений среднего девона (эйфельский век). Следовательно, камбиальная активность возникла немногим менее 400 млн. лет назад. Несколько позднее (живетский век среднего девона) возникают первые известные нам древовидные формы. Одним из первых был аневрофит германский (Aneurophyton germanicum), достигавший более 10 м высоты. К концу среднего девона появляется другое древесное растение - знаменитый археоптерис (Archaeopteris latifolia) - крупное дерево, вероятно, высотой более 30 м, с кроной ветвей, несущих спирально расположенные листья (рис. 94). Таким образом появились необходимые предпосылки для возникновения голосеменных. Это важнейшее в эволюции растительного мира событие произошло уже в девонском периоде. Лучшим доказательством явилась находка в 1968 г. Дж. Петтитом и Ч. Беком в верхнедевонских отложениях Северной Америки хорошо сохранившегося семени, названного археоспермой (Archaeosperma). Это древнейшее известное науке семя, принадлежавшее растению, которое существовало более 370 млн. лет назад, было еще очень примитивным (с интегументом, переходящим наверху в свободные "щупальца", соответствующие свободным концам спорангиев).
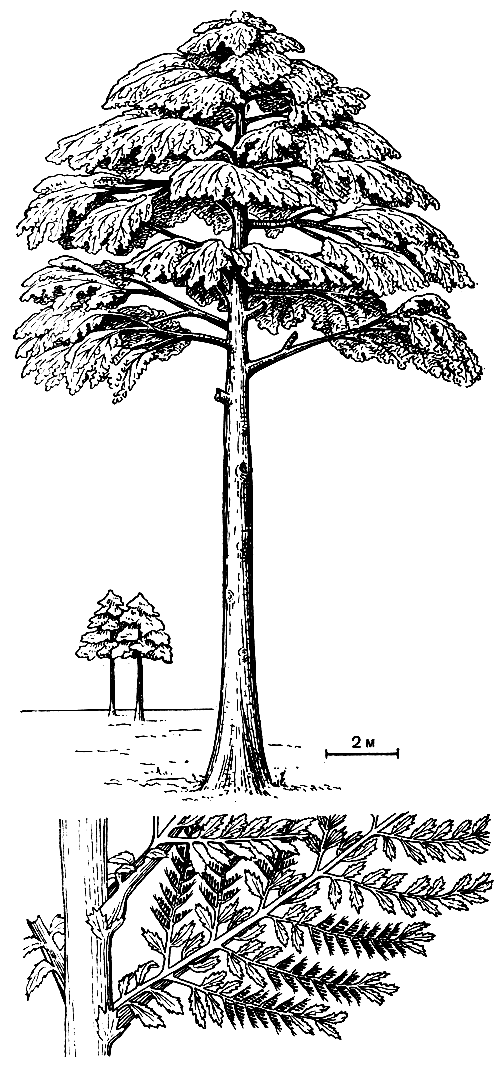
Рис. 94. Археоптерис (Archaeopteris). Внизу - часть облиственного стебля
Из факта нахождения голосеменных в верхнем девоне следует, что предков голосеменных следует искать среди палеозойских растений, возраст которых не моложе позднего девона. Тем самым из числа возможных предков голосеменных выпадают почти все типичные папоротники, от которых нередко пытались вывести голосеменные. В качестве их возможных предков остается рассматривать только древнейших представителей отдела папоротниковидных. Именно среди них мы находим разноспоровые древовидные формы с вторичной древесиной, которые могли дать начало голосеменным.
Нередко высказывается предположение, что голосеменные могли произойти от одного из наиболее высокоорганизованных девонских папоротниковидных, а именно от рода археоптерис или ближайших к нему форм. Но у археоптериса уже имеется сердцевина, вторичная ксилема у него сильно развита и толстая, а трахеиды точечные (с округлыми окаймленными порами). По уровню организации своих вегетативных органов археоптерис мало отличается не только от некоторых пермокарбоновых хвойных, но даже от некоторых современных форм. Можно было бы легко представить себе археоптерис в качестве исходной формы, если бы среди голосеменных не было семейств с гораздо более примитивной организацией. Конечно, можно было бы допустить, что хвойные и родственные им группы имеют самостоятельное происхождение, и тогда их происхождение от растений типа археоптериса не вызвало бы сомнений. Но все говорит о том, что голосеменные представляют собой естественную группу и имеют, следовательно, общее происхождение. Это доказывается не только строением такого сложного органа, как семязачаток, но также строением вегетативных органов. Так, по мнению Ч. Бека (1970), строение стелы семенного папоротника лигиноптериса (Lyginopteris oldhamia) во всех основных чертах, включая явное отсутствие листовых лакун, идентично со строением стелы хвойных. Но если голосеменные действительно имели общее происхождение, то они могли произойти только от предка с протостелической организацией стелы (как у некоторых древнейших семенных папоротников), со слабо развитой вторичной ксилемой и с лестничными трахеидами (сохранившимися даже у некоторых ныне живущих голосеменных). Ни археоптерис и ни аневрофит с их точечными трахеидами не могли быть такой предковой формой. Скорее всего это были какие-то более древние и примитивные протостелические, но в то же время разноспоровые папоротниковидные с лестничными трахеидами во вторичной ксилеме.
Таким образом, мы неизбежно приходим к выводу, что голосеменные произошли не от настоящих (типичных) папоротников, но от одной из боковых разноспоровых ветвей древнейших папоротниковидных растений. Это еще раз подтверждает сформулированное известным палеонтологом Э. Копом (1904) правило, согласно которому в процессе эволюции новые группы организмов обычно происходят не от высших представителей предкового таксона, а от сравнительно наименее специализированных его представителей.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'