Порядок циатейные (Cyatheales)
Семейство циатейные (Cyatheaceae) (В. Н. Гладкова)
Семейство циатейные объединяет свыше 1000 главным образом тропических видов, более половины которых являются древовидными папоротниками. Представители этой древней группы сочетают в своем морфологическом строении как примитивные черты, так и черты высокой организации, характерные для подвинутых семейств папоротников. Проводящая система варьирует у них от протостелы до сложной диктиостелы, опушение от примитивного, состоящего из волосков, до сложно устроенных чешуй, положение сорусов от маргинального до дорсального на жилках. Спорангии обычно крупные с косым кольцом утолщенных клеток, споры часто с развитой сложно устроенной периной, трилетные тетраэдрические (билатеральные только у части деннштедтиевых). Гаметофиты сердцевидные, зеленые, утолщенные в центре. Наблюдается у циатейных большое разнообразие и в хромосомных числах, составляющих нерегулярные ряды, в которых промежуточные звенья, вероятно, были утрачены в ходе эволюции.
Подсемейство циатейные (Cyatheoideae)
Виды этого подсемейства представляют собой естественную группу, главным признаком которой является поверхностное положение сорусов, сидящих на жилках на приподнятом свободном ложе, а также наличие чешуй в опушении.
Основной род подсемейства циатея (Cyathea) включает около 600 видов, равномерно распределенных между тропическими областями Старого и Нового Света. Особенно обильно виды циатеи представлены в относительно холодных и постоянно влажных горных тропических районах. Здесь среди них часто наблюдается узкий эндемизм: многие виды характерны только для одного района гор или даже растут на одной возвышенности. Некоторые из циатей заходят и в субтропики, на севере они достигают Южной Японии, на юге - Новой Зеландии и даже субантарктических Оклендских островов, Капской области и Огненной Земли.
Циатеи - характерные обитатели влажных горных тропических лесов, где они рассеянно встречаются в нижнем ярусе древостоя. Более многочисленные группы их наблюдаются на осветленных пространствах, по склонам к лесным рекам, на опушках. Заметным элементом растительного покрова они являются в так называемых мшистых лесах высокогорного пояса облаков и туманов - лесах, состоящих из одного яруса заросших мхом и покрытых многочисленными эпифитами невысоких деревьев, среди которых древовидные циатеи иногда даже доминируют. Растут циатеи и на искусственно расчищенных местах, часто присутствуют во вторичных растительных сообществах и играют определенную роль в восстановлении растительного покрова. Многие виды рода устойчивы против пожаров, и на горных склонах, подвергшихся действию огня, они образуют папоротниковые сообщества, которые затем постепенно вытесняются перевершинивающими их деревьями тропического леса.
Стволы циатеи, достигающие в высоту иногда более 20 м, имеют особое, характерное только для древовидных папоротников строение. Они не способны ко вторичному утолщению, и их устойчивость достигается развитием густого сплетения из жестких придаточных корней, составляющих покров ствола, особенно мощный в нижней части, где иногда он превосходит в несколько раз по толщине сам стебель. Опавшие листья часто оставляют на стволе свои жесткие основания, иногда листья опадают полностью. Увядшие листья у некоторых циатей сохраняются на стволе в течение некоторого времени, отгибаясь вниз и образуя своеобразную "юбочку" ниже кроны живых листьев (рис. 125). После опадения листьев на стволе остаются, как шрамы, крупные листовые рубцы - подковообразные, удлиненные или округлые, хорошо заметные только в верхней части ствола, а в нижней вскоре исчезающие под сплетающимися корнями. Описаны случаи ветвления стволов, ветви наблюдались как у основания стволов, так и в верхней части.

Рис. 125. Древовидный папоротник циатея грязноватая (Cyathea contaminans) в горном лесу на острове Ява
Проводящая система стволов циатей представляет собой диктиостелу - полый цилиндр с прорывами, соответствующими листовым основаниям. Каждый ее участок (меристела) окружен склеренхимой и, кроме того, особыми кубическими клетками с кристаллическим содержимым - черта, не встречающаяся в других семействах папоротников. В паренхиме коры стебля и в черешках листьев имеются слизистые мешки.
Листья циатеи могут быть от перистых до четырежды перистых, но обычно они дважды, трижды перистые, очень крупные, длиной до 6 м, на длинных черешках, которые бывают бородавчатыми или шиповатыми. В месте прикрепления первичных сегментов к стержню листа (рахису), а иногда и вдоль рахиса у многих видов имеются участки воздухоносной ткани овальной формы, называемые аэрофорами или пневматодами. Иногда эти участки ткани, служащие для газообмена, выдаются над поверхностью листа.
Необычным образованием у листьев многих видов циатеи являются свойственные им "афлебии" - уменьшенные базальные сегменты, часто почти нитевидно рассеченные, располагающиеся обычно у самого основания черешка. Все вместе они образуют как бы дополнительную кружевную крону на верхушке стебля. Они очень своеобразны и, собранные ботаниками отдельно от растения, иногда принимались за другие роды папоротников. Так, "афлебии" циатеи капской (Cyathea capensis) дважды были описаны как новые виды рода трихоманес (Trichomanes).
С нижней стороны листьев поверхностно на жилках располагаются сорусы, состоящие из перемешанных с волосковидными парафизами спорангиев, имеющих тонкую ножку и сидящих на приподнятом коническом или полушаровидном ложе. Наблюдается большое разнообразие покрывалец. У некоторых видов, прежде искусственно объединяемых в особый род алзофила (Alsophila), покрывальце отсутствует.
Развитие сложного опушения является у циатейных существенным эволюционным приобретением. Древовидная форма роста, дающая этим папоротникам жизненные преимущества в смысле использования света и рассеивания спор, в то же время создала для них некоторые трудности в водоснабжении высоко расположенных листьев. Они вынуждены были поэтому развивать густой покров из чешуй, волосков, шипов, которые все в совокупности, по-разному ориентированные и сложно устроенные, помогают поглощать и задерживать воду, присутствующую в воздухе в виде дождей и туманов.
Существует два главных типа чешуй на черешках листьев циатейных (рис. 126): чешуи структурно однородные, состоящие из одинаковых клеток, и чешуи окаймленные, у которых клетки, расположенные по краю, резко отличаются по форме, окраске и размеру от удлиненных клеток, находящихся в центре. В пределах второго типа чешуй, в свою очередь, имеется большое морфологическое разнообразие, выражающееся в наличии или отсутствии темных щетинок на чешуйках, в форме основания чешуй и характере прикрепления их к черешку.
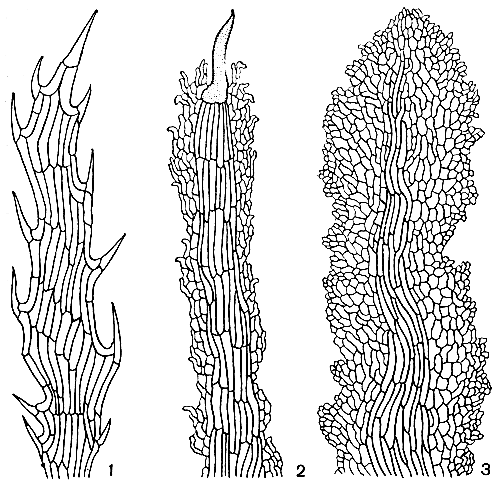
Рис. 126. Типы чешуй в подсемействе циатейных: 1 - циатея Купера (Cyathea cooperi), структурно однородные чешуи; 2 - циатея капская (С. capensis), окаймленные щетинистые чешуи; 3 - кнемидария ощетиненная (Cnemidaria horrida),окаймленные чешуи без щетинок
На основе указанных отличий некоторые птеридологи разделяют род циатея на следующие пять родов: пантропический род сфероптерис (Sphaeropteris, 20 видов) со структурно однородными чешуями, давший начало двум эволюционным линиям со структурно неоднородными окаймленными чешуями; в одной - пантропический род алзофила (Alsophila, 230 видов) и американский род нефелея (Nephelea, 30 видов), обладающие щетинистыми чешуйками, в другой - род циатея в узком смысле (110 видов в Америке) и род трихиптерис (Trichipteris, 90 видов в Америке), не имеющие щетинок на окаймленных чешуях.
Как уже говорилось, большинство циатей являются настоящими древовидными папоротниками. Но, хотя некоторые из них - гиганты в мире папоротников, преобладают среди циатей экземпляры с невысоким или среднего размера стволом, обычно высотой не более 10 м, а часто и гораздо меньшим. Имеются среди них и виды совсем "без ствола", с листьями, отходящими пучком на уровне почвы, как и виды с ползучим стеблем, например новозеландская циатея Коленсо (Cvathea colensoi).
В жизни современного населения тех стран, где растут циатеи, эти папоротники не играют очень большой роли. Являясь неотъемлемой чертой ландшафта, они кажутся здесь банальными растениями. Но для жителей стран с более холодным климатом их экзотический облик, напоминающий о далеком прошлом планеты, весьма привлекателен.
В культуре особенно популярны австралийские и новозеландские виды, в том числе циатея беловатая (С. dealbata), или серебристый древовидный папоротник (ее взрослые листья снизу густо покрыты белым налетом). Она встречается в лесах Новой Зеландии. Широко и успешно культивируются австралийские циатеи - циатея Купера (С. cooperi) и циатея южная (С. australis).
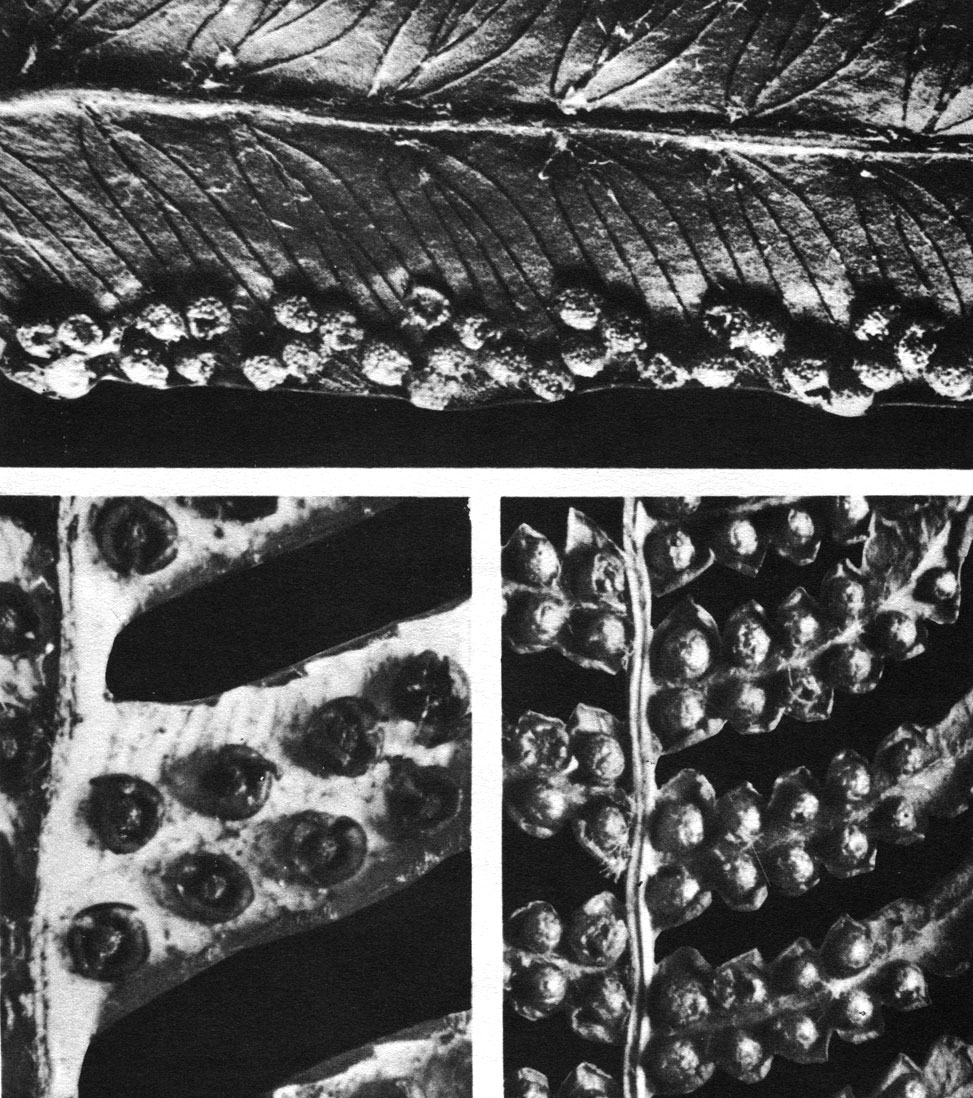
Таблица 29. Типы расположения сорусов у папоротников семейства циатейных: вверху - кнемидария ощетиненная (Cnemidaria horrida); внизу слева - циатея беловатая (Cyathea dealbata), видны чашевидные покрывальца и ложа сорусов; внизу справа - циатея сердцевинная (С. medullaris)
Ценится в культуре быстрорастущая циатея сердцевинная (С. medullaris), называемая на родине (Новая Зеландия, Тасмания и острова Тихого океана) черноствольным или черным папоротником, за темную окраску ствола и чешуй листовых оснований. Сердцевина этой и многих других циатей содержит много крахмала, и раньше местные жители использовали ее в пищу, обычно в печеном виде. По вкусу она напоминает печеные яблоки. Находят применение иногда и твердые части ствола, превращаемые после соответствующей сложной обработки в трости и разнообразные декоративные поделки (рис. 127). Идут стволы циатей и на постройку примитивных сооружений, иногда используют их как столбы, ульи и т. д. Споры и чешуи некоторых видов индейцы употребляют при лечении ран и кровотечений. Сердцевину и молодые листья восточноафриканских видов аборигены используют как средство от ленточных глистов.

Рис. 127. Барельеф 'Индейцы' из ствола циатеи
Второй род этого подсемейства - кнемидария (Cnemidaria), включающий всего 23 вида, характерен для тропиков Нового Света. По влажным местообитаниям в лесах и на лесных опушках, в горах до высоты 2300 м над уровнем моря, в областях, где нет продолжительных сухих периодов, кнемидарии растут в Мексике и Вест-Индии, в Центральной Америке, на севере и западе Южной Америки до Боливии и на юго-востоке Бразилии. По некоторым признакам кнемидария представляет собой высокую ступень эволюции в подсемействе циатейных. Виды этого рода имеют полудревовидный облик из-за слабо развитого прямостоячего или восходящего стебля, который редко достигает в длину более 1 м и в толщину более 7 см. Листья кнемидарий растут пучком и у некоторых видов бывают внушительных размеров (длиной до 3,5 м). Они всегда просто (однажды) перистые и имеют у большинства видов усложненное жилкование: базальные жилки сегментов у них соединяются, образуя ареолы, или сходятся друг с другом в выемке между сегментами. Округлые сорусы кнемидарии, как и у циатеи, лежат на жилках нижней поверхности листа. Они снабжены обычно чешуевидными покрыв а льдами. Наконец, кнемидария имеет набор в совершенстве развитых и разнообразных чешуй.
Есть у кнемидарии и уникальная черта, по которой ее виды безошибочно отличают от других циатейных: каждая сторона трилетных тетраэдрических спор снабжена в центре крупной (диаметром около 15 мкм) округлой порой, представляющей собой углубление или полость в экзине (рис. 128).
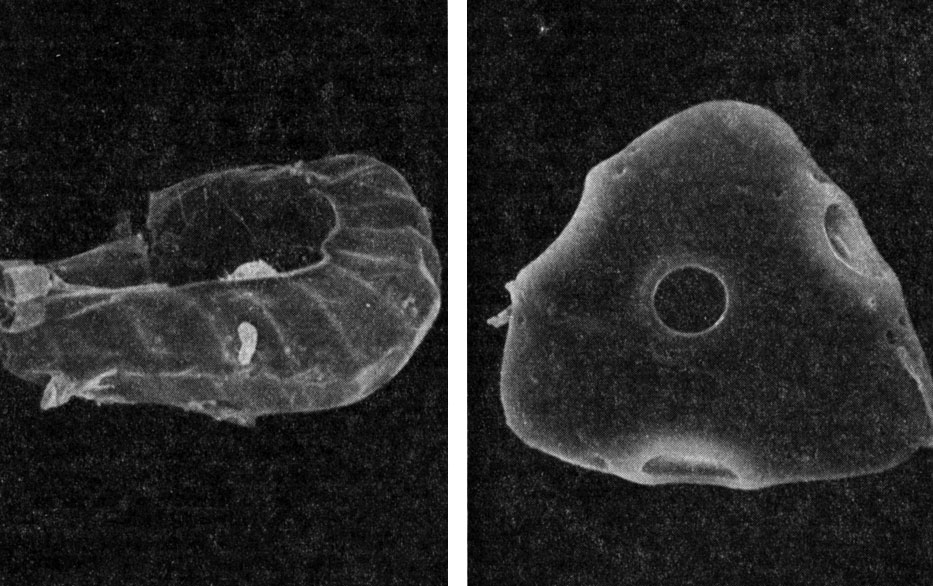
Рис. 128. Кнемидария ощетиненная (Cnemidaria horrida): слева - спорангии (увел, около 250); справа - спора (увел, около 2000)
Наиболее широко распространенной среди кнемидарий, встречающейся в пределах почти всего ареала рода, является кнемидария ощетиненная (С. horrida), которая более других напоминает по облику типичные древовидные папоротники этого подсемейства. Ее стебель достигает в высоту 4 м, а листья с шиповатыми черешками, покрытые снизу по жилкам паутинистым опушением, бывают длиной до 3,5 м.
Подсемейство диксоииевые (Dicksonioideae)
Тесные связи с циатейными имеет подсемейство диксониевые (Dicksonioideae), включающее древовидные (в большинстве случаев) папоротники, относимые к четырем родам: диксония (Dicksonia), цистодиум (Cystodium), кульцита (Culcita) и циботиум (Cibotium). От собственно циатейных роды диксониевых отличаются отсутствием чешуй и своеобразным строением покрывальца, закрывающего округлые сорусы, которые расположены на концах жилок по самому краю листа. Покрывальце диксониевых состоит из двух сомкнутых створок, наружная из которых у большинства видов зеленая и по своему происхождению является несколько видоизмененным краем листа, тогда как внутренняя, более тонкая и почти лишенная хлорофилла, является настоящим покрывальцем. Внутренняя створка прирастает основанием к приподнятому ложу соруса.
Род диксония состоит из 25 видов, обитающих на островах Малайского архипелага, Филиппинах, в Новой Гвинее, в Восточной Австралии, Тасмании и Новой Зеландии, на островах Тихого океана, на острове Св. Елены, а также в тропической Америке. Стволы диксоний по форме и своей внутренней структуре похожи на стволы циатеи. Они достигают в высоту 4-6, иногда 15 м и имеют множество придаточных корней, сплетающихся с основаниями опавших листьев. Крупные их экземпляры, часто обросшие густой массой эпифитов, могут быть у основания диаметром до 2 м, но встречаются среди диксоний и виды, почти лишенные надземного ствола.
Листья диксоний дважды, трижды перистые, сосредоточены на верхушке ствола. Их черешки, как иногда и другие части листа (рахис, жилки), покрыты волосками, которые у основания черешков часто бывают жесткими и длинными. Несущие сорусы сегменты имеют уменьшенного размера пластинку и обычно более глубоколопастные, чем стерильные.
У многих видов диксоний листья более жесткие, чем у циатей. Увядая, они, так же как у циатей, могут на некоторое время оставаться на стволе в виде юбочки в его верхней части.
Обитают диксонии обычно в горных лесах, на высоте до 3000 м над уровнем моря, предпочитая влажные горные ущелья, реже встречаются в сухих местообитаниях. Иногда они образуют крупные рощи, настоящие папоротниковые джунгли, и в этом помогает диксониям способность их стволов образовывать у основания почки, из которых формируются короткие горизонтальные побеги, дающие начало новым папоротникам. Таким образом, при благоприятных условиях диксония может быстро заселять большие территории.
Второй крупный род этого подсемейства - циботиум. Двустворчатое покрывальце у видов этого рода твердое и светлое. Его наружная губа лишена хлорофилла; она не кажется продолжением края листа и по строению полностью подобна внутренней.
В роде циботиум насчитывают 10-15 видов, растущих в лесах тропической Азии (от Ассама до Южного Китая и до западной части Малайского архипелага и Филиппин), а также на Гавайских островах, в Центральной Америке и Мексике. Виды циботиума - большей частью древовидные папоротники с прямым (редко стелющимся) стеблем, верхушка которого защищена толстым покровом из длинных мягких волосков; волоски имеются и на осях дважды, трижды перистых листьев.
В местах своего естественного произрастания виды циботиума прежде играли большую роль в жизни аборигенов. Их молодые листья и крахмалистую сердцевину стволов люди употребляли в пищу, а в годы изобилия применяли как корм для свиней. Обильно растущие на стволах волоски использовали как набивочный материал. Волоски видов циботиума и циатеи, кроме того, издавна применяли в медицине как эффективное кровоостанавливающее средство и перевязочный материал. Этот медикамент экспортировали из азиатских стран, и он был известен под малайским названием "пенгавар Джамби", т. е. "целебное средство из Джамби" (по главному месту вывоза). Волоски пенгавара применяли как гемостатическое средство уже в средние века.
Циботиумы привозили из Азии в виде кусков ствола, которым придавали форму животного, оставляя на стволе 4 или 5 черешков листьев, имитировавших ноги и хвост. Сам же ствол с массой длинных волосков на нем был похож на тело животного. До конца XIX века считали, что циботиум лежит в основе известной легенды о чудесном растении - барашке (Agnus scythicus), прикрепленном к земле с помощью стебля, выходящего из его пупка, и пожирающем траву вокруг в пределах досягаемости этого стебля. Легенда эта нашла отражение и в научном названии одного из видов циботиума - Cibotium barometz ("баромец" - искаженное русское слово "баранец", уменьшительное от баран).
Стволы циботиумов, упругие вследствие большого количества жестких переплетенных корней на них и устойчивые против гниения, иногда использовались для покрытия дорог в болотистых местах. А корни, как и корни других древовидных папоротников, находят применение как среда для выращивания орхидей и бромелиевых. Многие виды циботиума культивируют как декоративные растения. Издавна, например, известен в культуре мексиканский древовидный циботиум Шиде (С. schiedei).
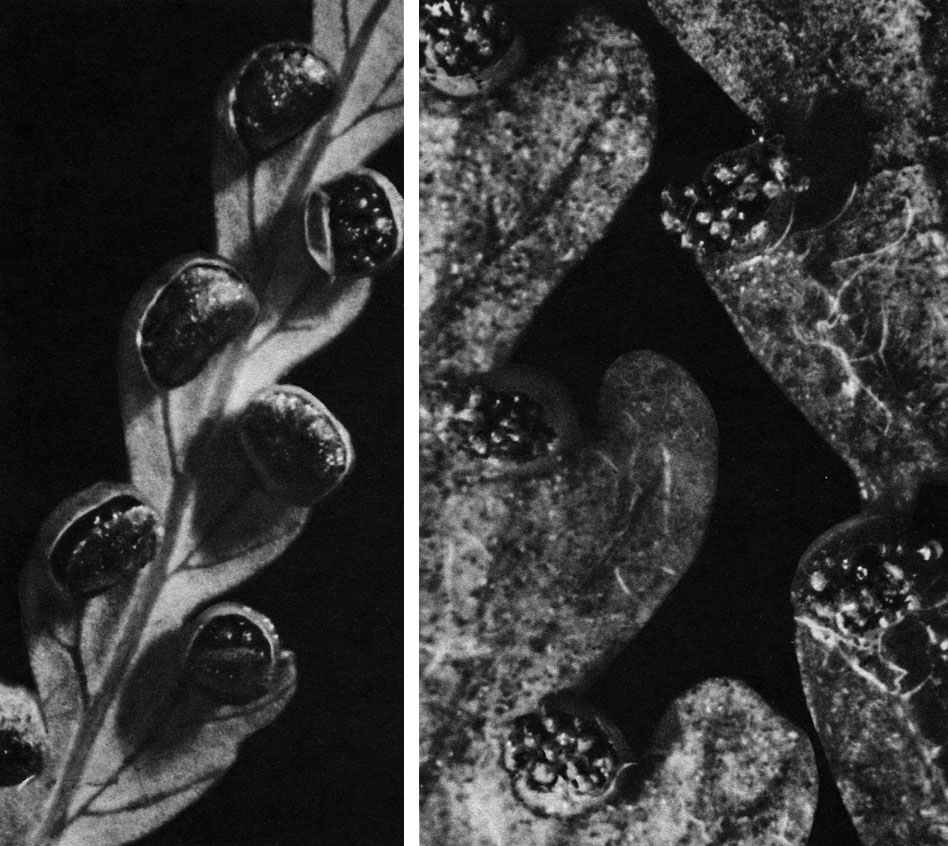
Таблица 30. Типы расположения сорусов у папоротников семейства циатейных: слева - циботиум Шиде (Cibotium schiedei); справа - деннштедтия веховидная (Dennstaedtia cicutaria)
Подсемейство тирсоптерисовые (Thyrsopterldoideae)
Единственный представитель этого подсемейства древовидный папоротник тирсоптерис элегантный (Thyrsopteris elegans, рис. 129) растет только в одном месте на земле - в лесах островов Хуан-Фернандес, в Тихом океане недалеко от побережья Южной Америки. Тирсоптерис достигает 1-1,5 м в высоту, отличаясь ото всех других представителей циатейных резко выраженным диморфизмом стерильной и фертильной части своих многократно перистых листьев. Фертильны обычно 2-3-я пары нижних сегментов первого порядка, на них сегменты последующих порядков совершенно лишены пластинки, и округлые сорусы кажутся сидящими прямо на верхушках осей. Ложе соруса колонновидно приподнято, как у циатеи, а две створки покрывальца, свободные только на ранних стадиях развития, сначала имеют вид шара, а затем приобретают вид неглубокой симметричной чаши. У тирсоптериса отмечена редкая для древовидных папоротников способность давать отпрыски от ствола.
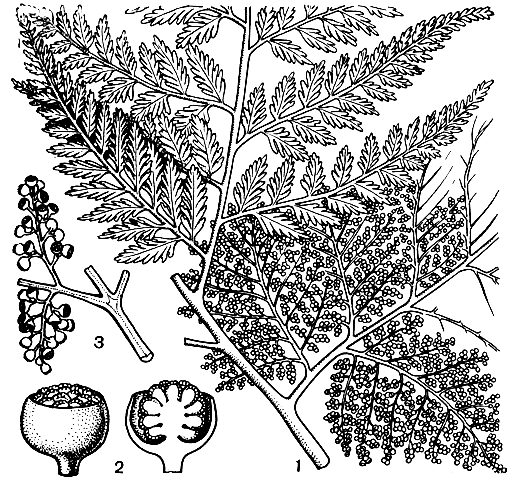
Рис. 129. Тирсоптерис элегантный (Thyrsopteris elegans): 1 - часть листа со стерильным и спороносным (нижним) сегментами; 2 - часть спороносного сегмента; 3 - сорус с покрывальцем
Подсемейства лофосориевые (Lophosorioideae) и метаксиевые (Metaxyoideae)
Два очень древних представителя циатейных роды лофосория (Lophosoria) и метаксия (Metaxya), составляющие два особых подсемейства, отошли от основной линии развития семейства на очень ранней стадии его эволюции. Ископаемые остатки этих папоротников известны в юрских отложениях Европы, Индии, Кореи.
Современные роды - лофосория и метаксия как бы сконцентрировали в себе примитивные признаки, порознь встречающиеся в других подсемействах циатейных. Корневища у этих папоротников соленостелические или с примитивной диктиостелой, опушение состоит только из волосков. Сорусы лишены покрывальца (по-видимому, изначально) и расположены поверхностно на разветвлениях жилок. Крупные спорангии косым, непрерванным кольцом созревают все одновременно.
Род лофосория представлен одним видом, лофосорией четырежды перистой (Lophosoria quadripinnata), произрастающей от Мексики и Вест-Индии до южного Чили. Это невысокое растение с восходящим стеблем и многократно (обычно трижды) перистыми листьями. Метаксия - также тропический американский род (Малые Антильские острова, Центральная Америка и Южная Америка до Боливии) с единственным видом метаксией клювовидной (Metaxya rostrata), имеющей ползучее массивное корневище и однажды перистые листья. Уникальной (среди циатейных) чертой метаксии является расположение сорусов по нескольку на одной жилке. Отличается метаксия от лофосории, кроме того, бороздчатым рахисом, отсутствием склеренхимы и кубических клеток в стебле, строением устьичного аппарата с тремя побочными клетками, строением спор, а также хромосомным числом n = 96 (у лофосории n = 65).
Подсемейство локсомовые (Loxsomoideae)
Не менее древней, чем два предыдущие подсемейства, ветвью циатейных являются роды локсома (Loxsoma) с одним видом в Новой Зеландии и близкий к ней локсомопсис (Loxsomopsis) с тремя видами в Андах. Некоторые птеридологи считают эти роды "живыми ископаемыми", соединяющими в себе черты нескольких современных семейств. По своим вегетативным признакам (ползучее корневище, дважды, трижды перистые листья со свободными жилками) локсома напоминает некоторые деннштедтиевые, по анатомии корневища имеет общие черты с диксониевыми, а по строению сорусов очень близка к семейству гименофилловых. Маргинальный сорус на конце жилки располагается на почти цилиндрическом ложе и заключен в чашевидное покрывальце. Когда в сорусе образуются спорангии, меристема ложа продуцирует ножку, поднимающую сорус над покрывальцем и выносящую его за край листа. Спорангии локсомы крупные, асимметрично-грушевидной формы, с короткой ножкой и неполным почти верхушечным кольцом. Только некоторые клетки в кольце утолщены. Отсутствует и хорошо выраженный стомий, открывание спорангия происходит вертикальной щелью.
Локсома была открыта в 1826 г. ботаником А. Каннингемом и названа в его честь локсомой Каннингема (Loxsoma cunninghamii). В лесах севера Новой Зеландии она привлекает внимание светлой окраской крупных перистых листьев, контрастирующей с темно-зеленым фоном леса.
Род локсомопсис отличается от локсомы, главным образом, своим спорангием, у которого все клетки кольца утолщены и стомий хорошо дифференцирован.
Подсемейство деннштедтиевые (Dennstaedtioideae)
Среди подсемейств семейства циатейных деннштедтиевые занимают несколько обособленное положение и иногда рассматриваются как самостоятельное семейство. Почти все 13 родов этого подсемейства обладают ползучими соленостелическими дорсивентральными корневищами, одетыми волосками. Исключение представляют (имеют чешуйчатые стебли) роды: сакколома (Saccoloma) и ортиоптерис (Orthiopteris). Характерные признаки подсемейства демонстрирует пантропический род деннштедтия (Dennstaedtia), встречающийся также в умеренной части Южной Америки, в Австралии и Новой Зеландии.
Род охватывает около 70 видов крупных лесных папоротников с сильно рассеченными в большинстве случаев листьями. Сорус деннштедтии, как и у диксонии, покрыт двумя покрывальцами: наружным, являющимся модификацией края листа, и внутренним, настоящим покрывальцем. Эти два покрывальца, срастаясь частично или полностью, образуют двустворчатую или чашевидную структуру, внутри которой на приподнятом свободном ложе размещаются спорангии.
Длинное ползучее корневище деннштедтии способно интенсивно ветвиться, благодаря чему эти папоротники часто образуют заросли, а некоторые их виды стали широко распространенными сорными растениями. Ветви на корневище деннштедтии, как и у большинства других родов деннштедтиевых, отходят не непосредственно от корневища, а от листа, точнее, от нижней части его черешка (так называемое эпипетиолярное ветвление). В этом, в частности, проявляется "веточная" природа листьев папоротников.
В пределах подсемейства деннштедтиевых можно проследить эволюционную тенденцию к смещению соруса от края листа на его нижнюю поверхность и связанному с этим изменению в структуре покрывалец. Микролепия (Microlepia) - крупный род подсемейства с 45 преимущественно азиатскими видами (2 ее вида достигают и Приморского края СССР), имеет именно такой смещенный тип соруса, с покрывальцем получашевидной формы, прикрепленным к ложу снизу и частично по сторонам.
Другая эволюционная тенденция в подсемействе выражается в редукции внутреннего покрывальца до полного его исчезновения и в образовании слитного соруса. К этой группе родов принадлежит широко известный орляк (Pteridium) с одним очень полиморфным видом орляком обыкновенным (P. aquilinum), почти космополитно распространенным по всему земному шару (кроме полярных областей и пустынь). Иногда орляк разделяют и на несколько близких видов.
Орляк обладает глубоким подземным ветвящимся корневищем. Боковые восходящие (до самой поверхности земли) ветви его ветвятся повторно и формируют листья. Листья орляка довольно жесткие, с крупной тройчатой пластинкой, чаще всего трижды перистые (рис. 89). Нижняя пара перьев у своего основания имеет нектарники, выделяющие сладкую жидкость, привлекающую муравьев. Край сегментов листьев завернут и закрывает непрерывную линию сорусов, лежащих на сосудистом тяже, соединяющем концы жилок. С внутренней стороны этого тяжа прикреплено слабо развитое внутреннее покрывальце, имеющее вид непрерывной или разорванной пленки, иногда же оно представлено немногочисленными волосками.
Являясь довольно примитивным в своих морфологических чертах (волоски на корневище, эпипетиолярное ветвление, свободное жилкование), орляк в то же время приобрел черты высокой организации в строении проводящей системы корневищ, представляющей собой полициклическую (из двух цилиндров) диктиостелу, которая наряду с трахеидами содержит и настоящие сосуды с лестничными перфорациями.
Орляк распространен на равнинах и в горах (до 3000 м). Наиболее часто он встречается в светлых лесах или на лесных опушках, обычен на песчаной почве в сосновых лесах, на открытых возвышенных местах, в зарослях кустарников.
В естественных местообитаниях орляк редко становится агрессивно расселяющимся растением. Но деятельность человека способствует превращению его в один из самых распространенных папоротников. Глубоко залегающие корневища и способность к бурному вегетативному размножению позволяют орляку осваивать места пожарищ, сведенных земель, заброшенных полей, плантации и пастбища. В некоторых странах орляк стал трудноискоренимым сорняком, требующим специальных мер борьбы.
В жизни человека, однако, орляк играл и играет не только отрицательную, но и большую положительную роль. В Японии, например, его естественных запасов не хватает для удовлетворения потребностей населения в этом папоротнике. Только в Токио ежегодно употребляют в пищу (наподобие спаржи или маслин в европейских странах) более 300 000 кг молодых листьев орляка.
Как пищевое растение орляк известен у многих народов. Маори Новой Зеландии, аборигены Канарских островов, индейцы Америки приготовляли из его высушенных и измельченных корневищ хлеб или употребляли их в пищу в сыром виде. В голодные годы хлеб из орляка пекли и в некоторых европейских странах.
Известно употребление орляка и в медицине как противоглистного средства, для лечения рахита у детей и т. д. Большую популярность этот папоротник имел и в быту населения многих стран. В Англии его листьями крыли крыши домов, использовали как подстилку для скота. Шел орляк и на топливо, на удобрение. Зола этого папоротника издавна служила важным источником промышленного получения поташа (карбоната калия), использовавшегося в производстве тугоплавкого стекла и зеленого мыла. Моющие и отбеливающие свойства этой золы были известны даже до появления мыла. Шарики золы летом заготавливали впрок и использовали для получения щелока для стирки в течение всего года. Эта практика в некоторых районах Британских островов продолжалась вплоть до XIX в.
Орляк является предметом пристального внимания ученых и в наши дни. На состоявшемся в 1974 г. в Англии симпозиуме, посвященном этому папоротнику, широко обсуждали вопросы его истории и таксономии, экологии, генетики, изучения химического состава и канцерогенности некоторых содержащихся в нем веществ, а также возможности его использования.
Орляк прежде относили к птерисовым, сближая его с родом птерис (Pteris), также имеющим слитный сорус, защищенный отогнутым краем листа, Но наличие остатка внутреннего индузия, а также хромосомное число орляка n = 52 говорят о его родстве с деннштедтиевыми, в частности с родом гиполепис и близкими ему родами, у которых преобладает это хромосомное число.
Подсемейства линдсеевые (Lindsaeoideae) и монахосоровые (Monachosoroideae)
Линдсеевые объединяют 6-8 родов и около 250 видов в основном тропических папоротников, близких по своему морфологическому строению к деннштедтиевым. Они также обладают ползучими корневищами (но с протостелой, реже с соленостелой), в опушении которых, в отличие от деннштедтиевых, преобладают узкие чешуйки, а не волоски. Обычно это некрупные папоротники с цельными или перистыми листьями, вдоль края которых на концах жилок лежат сорусы, снабженные внутренним покрывальцем, край листа при этом не загнут. Сорусы часто сливаются друг с другом, образуя ценосорус. Некоторые из линдсеевых, например тропический американский род одонтосория (Odontosoria), имеют колючие лазящие листья с неограниченным ростом и могут подниматься по опоре. Имеются лазящие виды и среди папоротников рода линдсея (Lindsaea). Таковы, например, линдсея гребенчатая (L. pectinata) и лазящая (L. scandens), встречающиеся в изобилии в тенистых лесах тропиков восточного полушария. Они забираются на нижнюю часть стволов деревьев или на скалы. Линдсея, являющаяся самым крупным родом в подсемействе, охватывает свыше 150 видов, растущих в тропических и субтропических областях обоих полушарий, особенно восточного. Папоротники этого рода имеют очень характерный облик. Некрупные листья их с уменьшающимися к верхушке и основанию сегментами, имеющими вид трапеции, полумесяца или параллелограмма, очень напоминают адиантум, но строение сорусов легко отличает их от этого рода.
Большинство линдсей - наземные или наскальные лесные папоротники, растущие часто вблизи рек и болот как на равнинах, так и в горах. Некоторые виды встречаются также в саванне (на увлажненных местах). Изредка линдсеи ведут и эпифитный образ жизни.
Род монахосорум (Monachosorum), представляющий монотипное подсемейство монахосоровых, состоит из 5 видов, обитающих в странах Азии (от Новой Гвинеи до Индии и Японии). Это наземные папоротники с восходящими короткими корневищами, несущими перистые (от однажды до четырежды перистых) листья. Сорусы у рода монахосорум смещены на нижнюю поверхность листа и расположены вдоль терминальной части жилок. Ложе соруса едва приподнято, покрывальце отсутствует. Некоторые птеридологи считают короткие чешуйчатые корневища и дорсальный сорус монахосорума свидетельством его родства с другой линией развития - с семейством асплениевых.
Семейство гименофилловые (Hymenophyllaceae) (Т. Д. Сурова)
Гименофилловые (в переводе на русский язык "пленчатолистные") - одно из самых интересных семейств современных папоротников. Иногда их называют "пленчатыми папоротниками" (по-английски filmy ferns), чем подчеркивается строение их тонких, почти прозрачных листьев (табл. 32, 33), ткани которых подверглись значительной редукции в процессе приспособления к жизни во влажной среде. Все без исключения гименофилловые - сухопутные растения, хотя по строению они скорее напоминают водные, а внешне многие из них очень похожи на мхи и печеночники. Своеобразный облик и строение как спорофита, так и гаметофита этих папоротников дало повод некоторым ботаникам прошлого века считать гименофилловые очень примитивными папоротниками.

Таблица 32. Гименофнлловые: 1 - трихоманес Манна (Trichomanes mannii) (увел, около 4); 2 - трихоманес кипарисовидный (Т. cupressoides); 3 - трихоманес курчавый (Т. crispiforme) (увел, около 1,5); 4 - трихоманес либерийский (Т. liberiense) (увел, около 7,5)
Гименофилловые - мелкие эпифитные, наскальные или напочвенные папоротники, обитатели тропических дождевых лесов достигают наибольшего разнообразия в горных районах. Большинство гименофилловых приурочено к нижнему ярусу тропических дождевых лесов, поселяются на корнях, стволах, нижних ветвях деревьев, обычно не выше 2 м над поверхностью почвы. Несколько видов, растущих на одном и том же дереве, занимая какую-нибудь его часть, как правило, не смешиваются друг с другом. Имеются факультативные эпифиты: их можно встретить и на стволах деревьев и на почве. Лишь небольшое число видов рода трихоманес (Trichomanes) - типично наземные папоротники. В сумраке нижнего яруса тропического леса эти нежные существа тесно соседствуют с многочисленными мхами и печеночниками, часто превышающими размерами гименофилловые. Зеленый ковер из мхов и гименофилловых задерживает испарение и способствует тем самым накоплению влаги на поверхности субстрата, будь то корни, стволы деревьев и древовидных папоротников или пни, скалы, почва.
Известно около 700 видов гименофилловых, распределяющихся примерно поровну между двумя большими родами - гименофиллом, или тонколистником (Hymenophyllum), и трихоманесом. Некоторые виды или группы видов часто выделяют в самостоятельные роды. Поэтому общее число родов в семействе колеблется (в понимании разных авторов) от 6 до 34. Американский птеридолог К. Мортон (1968) в семейство гименофилловые наряду с широко распространенными трихоманесом и гименофиллом включает еще 4 монотипных рода, встречающихся в южном полушарии. В Новой Зеландии обитает кардиоманес почковидный (Cardiomanes reniforme) - мелкий наземный папоротник, единственный представитель этого рода. В Новой Каледонии исключительно редко встречается розенштокия (Rosenstockia) - крошечное эпифитное растеньице, очень малоизученное, с неясными связями. Южная Америка - местожительство еще двух родов этого семейства. В Чилийских Андах и на острове Хуан-Фернандес можно встретить гименоглоссум окровавленный (Hymenoglossum cruentum), который по строению спорангиев близок к роду гименофилл, но отличается от него, как и от других родов, строением пластинки листа. И наконец, последний род серпиллопсис (Serpyllopsis), представленный видом серпиллопсис дернистый (S. caespitosa), обитает на крайнем юге Южной Америки, на Фолклендских островах и на острове Хуан-Фернандес.
По числу видов гименофилловые составляют примерно 1/14 всех видов папоротников, живущих на земном шаре. Однако их доля значительно возрастает по мере продвижения к югу, составляя на островах Фиджи и Общества 1/10 и достигая максимального значения (1/5) в Новой Зеландии.
Гименофилловые - обитатели тропических и субтропических стран обоих полушарий, но 10 видов этого семейства заходят южнее 50° ю. ш. и прекрасно чувствуют себя в промозглом климате субантарктических островов. Известно также несколько видов, встречающихся в северных умеренных широтах, в том числе в СССР.
Всем видам гименофилловых свойственна в высшей степени необычная специализация, связанная с гигрофильным образом их жизни и позволившая этому, по всей вероятности, достаточно древнему семейству процветать в наши дни в условиях, мало пригодных для большинства других сосудистых растений. Эволюция гименофилловых шла в направлении редукции тканей спорофита и частично гаметофита.
Нежные, почти прозрачные зеленые или красновато-бурые листья большинства этих растений состоят из одного слоя одинаковых клеток (табл. 32, 33). Как исключение, почти во всех родах семейства встречаются виды, в листовой пластинке которых насчитываются 2, 3 или 4 ряда клеток. У гименофилла южного (Н. australe) листья двуслойные. Трехслойные листья свойственны гименофиллам расширенному (Н. dilatatum) и шероховатому (Н. scabrum), а также трихоманесу изящному (Trichomanes elegans). У кардиоманеса почковидного пластинку листа (не менее тонкую, чем у однослойных листьев) составляют четыре ряда клеток. Так или иначе, но устьица и межклетники в листьях гименофилловых отсутствуют.

Таблица 33. Гименофилловые: 1 - трихоманес выгрызенный (Trichomanes erosum) (увел, около 1,5); 2 - трихоманес выгрызенный (увел, около 4); 3 - трихоманес Меттениуса (Т. mettenii) (увел, около 4); 4 - трихоманес Баллара (Т. ballardianum) (увел, около 4); 5 - трихоманес гвинейский (Т. guineense) (увел, около 1,5)
Тип строения листа, характерный для гименофилловых, в других семействах папоротников и наземных цветковых растений не встречается. Исключением являются несколько редких южноамериканских тропических видов гименофиллопсиса (Hymenophyllopsis), единственного рода семейства гименофиллопсисовых (Hymenophyllopsidaceae), очень своеобразных небольших размеров наземных папоротников с прямостоячим, покрытым чешуйками (чего никогда не бывает у гименофилловых) стеблем. Проводящая система стебля у них сифоностелическая (у гименофилловых только протостелическая). Но по строению пластинки листа они удивительно сходны с гименофилловыми. У гименофиллопсиса пластинка листа состоит из 3-4 рядов плотно лежащих клеток и также лишена устьиц.
Спорофит гименофилловых очень варьирует по форме и величине. Как правило, он представлен тонкими ползучими дорсивентральными корневищами с двумя рядами листьев на верхней стороне. Некоторые наземные виды трихоманеса имеют вертикальные корневища со спирально расположенными листьями. Корневище, как уже упоминалось, лишено каких бы то ни было чешуек. От корневища отходят тонкие корни, но у некоторых мелких эпифитных видов корни могут редуцироваться вплоть до полного их исчезновения. Функцию корней в таких случаях берут на себя многочисленные волоски, расположенные на корневище, черешках или на нижней поверхности листа.
Гименофилловые могут достигать значительных размеров. Свисающие, напоминающие тончайшее кружево листья гименофилла прекраснейшего (Hymenophyllum pulcherrimum) достигают длины 60 см, однако наиболее характерными для этого семейства являются листья, размеры которых не превышают 2-3 см, а крошечный трихоманес Гёбеля (Trichomanes goebelianum), длиной 3-4 мм,- настоящий карлик среди сосудистых растений.
Пластинка листа гименофилловых может быть цельная, но обычно она более или менее перисторассеченная. Для некоторых видов трихоманеса, например трихоманеса израстающего (Т. proliferum) из секции гонокормус (Gonocormus), постоянным признаком является деление рахиса на две части - так называемые "пролиферирующие черешки". Эту секцию (как и многие другие) часто выделяют в особый род. У других видов трихоманеса (из секции феея - Feea) ярко выражен диморфизм листьев: вегетативные и спороносные листья отличаются не только по функции, но и по форме. Жилкование большинства видов равно- или неравнодихотомическое, реже перистое. Классический пример примитивнейшего равнодихотомического жилкования - в цельных листьях кардиоманеса почковидного. В листьях некоторых трихоманесов присутствуют так называемые ложные жилки. Предполагают, что в некоторых случаях это обычные жилки, утратившие в результате редукции ксилемы водопроводящую функцию.
Тонкие листья гименофилловых, лишенные устьиц, абсолютно проницаемы для воды. Водный режим этих папоротников похож на режим мхов и печеночников. Процветая в условиях с относительной влажностью, близкой к 100%, гименофилловые по-разному приспособились переносить периодическую засуху. Мелкие наземные и эпифитные папоротнички покрывают субстрат сплошным ковром, что препятствует избыточному испарению и способствует созданию необходимого для них микроклимата. С наступлением засухи (гименофилловые ощущают недостаток влаги при относительной влажности воздуха ниже 90%) некоторые из них быстро теряют воду и высыхают до ломкости, сморщиваясь или скручивая пластинку листа, но легко оживают при насыщении воздуха водяными парами. Специальные опыты показали, что способность пополнять потери воды за счет поглощения паров из воздуха лучше развита у эпифитных гименофилловых.
Несколько видов гименофилловых найдены в необычно сухих для этого семейства местообитаниях. В Новой Зеландии встречается гименофилл крошечный (Hymenophyllum minimum), растущий там не только в дождевых лесах, но также на пропитанных солеными брызгами прибрежных скалах, в горных субальпийских ущельях, в сухих, не защищенных от ветра зарослях лептоспермума (Leptospermum) из миртовых. Ряд видов произрастает даже на каменистых обнажениях в альпийском поясе, выше верхней границы леса. В таких местообитаниях папоротники имеют более мелкие, гуще опушенные листья, по сравнению с представителями, населяющими низинные дождевые леса. Как правило, большую часть жизни их листья находятся в скрученном состоянии и кажутся почти неживыми.
Кардиоманес почковидный приурочен к относительно влажным местам, образует обширные маты, которые в сухую погоду высыхают до такой степени, что похрустывают под ногами, наподобие наших лишайников. Но стоит пройти дождю, и листья вновь становятся зелеными и упругими.
В горных районах Новой Зеландии и Тасмании на стволах и ветвях банксии (Banksia) и атротаксиса (Athrotaxis), часто на мертвых стволах либоцедруса (Libocedrus) образует подушки неожиданного серого цвета удивительное растеньице - гименофилл Малинга (Hymenophyllum malingii), относимый иногда к роду трихоманес и заслуживающий, скорее всего, выделения в особый род аптероптерис (Apteropteris). Отвернув лист, можно заметить красноватую окраску его нижней поверхности. Конечные сегменты листьев свернуты в трубку, и все растение покрыто густой шубой серых волосков. Волоски, как губка, впитывают попадающую на лист воду и препятствуют ее потере. Только после дождя проявляется зеленый цвет фотосинтезирующей ткани, обычно замаскированный серыми и буроокрашенными волосками. Опушение встречается и у других гименофилловых. Расположение волосков и их форма - важный систематический признак. Некоторые исследователи предлагают выделить из рода гименофилл отдельный род сфероциониум (Sphaerocionium), характеризующийся специфическими звездчатыми волосками.
Гименофилловые очень чувствительны к малейшим изменениям привычной им окружающей среды. Пожары, вырубки, выпас животных вызывают изменения освещенности и влажности леса, приводя к полному исчезновению там видов этого семейства. Иногда участки тропических дождевых лесов кажутся нетронутыми, но отсутствие в них гименофилловых указывает на вторичную природу этих лесов. На границах распространения гименофилловые заселяют места, экологические условия которых обеспечивают необходимый им микроклимат с достаточной влажностью, неподвижностью воздуха и значительным затенением. На отвесных скалах, на пнях и корнях деревьев в лесах Великобритании и Ирландии можно увидеть свисающие бледно-зеленые нежные листья гименофилла танбриджского (Hymenophyllum tunbridgense). Его ближайший родственник - гименофилл Вилсона (Н. wilsonii), менее требовательный к свету и влажности, по западным берегам Европы, омываемым Гольфстримом, продвинулся далеко на север, обосновался в Норвегии и достиг 63° с. ш.- Фарерских островов. Несмотря на то что на Британских островах встречаются оба вида, места обитания их четко разделены и никогда не совпадают. Ареал гименофилла танбриджского более разорван, имеются реликтовые местонахождения этого папоротника в Европе и на Кавказе (в районе Батуми), в лесистых малодоступных ущельях, которые, как известно, служат убежищами многих реликтовых растений. Низкие температуры, видимо, не являются фактором, тормозящим распространение гименофилловых на север. Любопытное наблюдение было проведено над гименофиллом танбриджским во Франции в районе Фонтенбло. В период с 1946 по 1954 г. папоротники перенесли в общей сложности 600 дней с морозами. За это время наблюдались довольно длительные периоды без оттепелей, более того, были зарегистрированы 15 дней, в течение которых растения были покрыты льдом, но и это не вызвало их гибели. В еще более суровых условиях растет на нашем Дальнем Востоке гименофилл Райта (Н. wrightii). Замечательной является находка этого теплолюбивого папоротничка на Сахалине с его среднегодовой температурой +0,5 °С и минимальной - 40°С. Только толстый снежный покров спасает его от вымерзания. Малодоступные девственные леса ущелий Аджарии и не тронутая ни пожарами, ни вырубками елово-пихтовая тайга - последние убежища гименофилловых на территории Советского Союза.
Разделение гименофилловых на два основных рода связано с формой покрывальца - двулопастного у гименофилла и трубчатого - у трихоманеса (рис. 130). Сорусы гименофилловых расположены на верхушках жилок по краю листа. Спорангии с косым кольцом. Развитие спорангиев идет в базипетальной последовательности (от верхушки к основанию) на ложе соруса. Сорусы градатные. У гименофилла ложе соруса относительно короткое, имеет ограниченный рост и не выступает, как правило, за пределы покрывальца. У трихоманеса образуется базальная меристематическая ткань и ложе продолжает расти неограниченно долго, свешиваясь в виде длинной, тонкой нити за пределы покрывальца. У гименофилла образуется меньше спорангиев, чем у трихоманеса, но они крупнее и спор в каждом спорангии образуется больше - до 512, в то время как у трихоманеса - до 64. Споры гименофилловых тетраэдрические, по-видимому, без перины, с очень тонкой экзиной. Характерной особенностью их является наличие пигмента. Хлорофилл-содержащие споры встречаются и в некоторых других родах папоротников, ведущих обычно эпифитный образ жизни. Зеленые споры обладают способностью к быстрому прорастанию без выраженного периода покоя, но быстро теряют жизнеспособность. Прорастание спор гименофилловых происходит менее чем за три дня, нередко в еще не раскрывшихся спорангиях. Гаметофиты растут очень медленно. Проходят годы, прежде чем они созревают и начинают продуцировать половые органы. По строению зрелых гаметофитов гименофилловые распадаются на два основных типа. Гаметофит гименофилла и близких родов лентовидный (рис. 131), у трихоманеса - нитчатый (рис. 132). Как отмечалось, нитчатый гаметофит представляет задержанную и несколько видоизмененную ювенильную стадию обычного пластинчатого или лентовидного гаметофита. Гаметофиты гименофилловых зеленые, обычно обоеполые, но в культуре у гименофилла игловидного (Н. acanthoides) отмечены и раздельнополые.
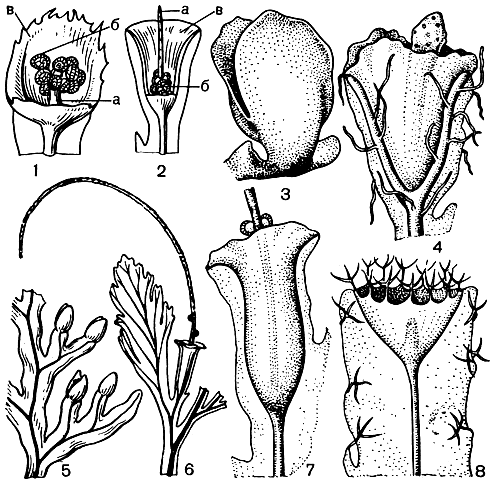
Рис. 130. Сорусы гименофилловых: 1 - строение соруса гименофилла бородчатого (Hymenophyllum barbatum) - удалена передняя лопасть покрывальца; 2 - строение соруса трихоманеса маленького (Trichomanes minutum) - удалена часть покрывальца; а - ложе соруса; б - спорангии; в - покрывальце; 3 - гименофилл щитовидный (Hymenophyllum peltatum); 4 - серпиллопсис дернистый (Serpyllopsis caespitosa); 5 - гименофилл осмундовидный (Hymenophyllum osmundoides); 6 - трихоманес зубчатый (Trichomanes dentatum); 7 - трихоманес жилковатый (Т. venosum); 8 - трихоманес Лайелла (Т. lyallii)
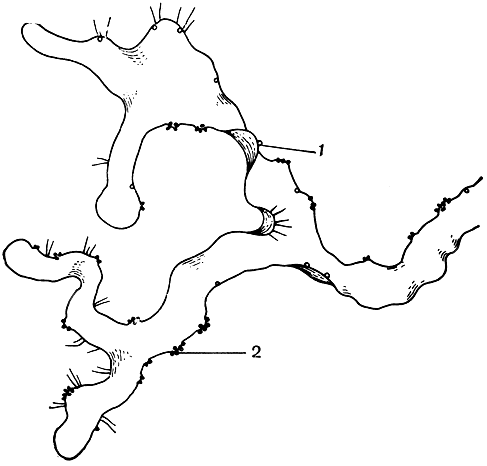
Рис. 131. Ленювидный гаметофит гименофилла Курца (Hymenohyllum kurzii) - брюшная сторона: 1 - антеридии; 2 - архегонии
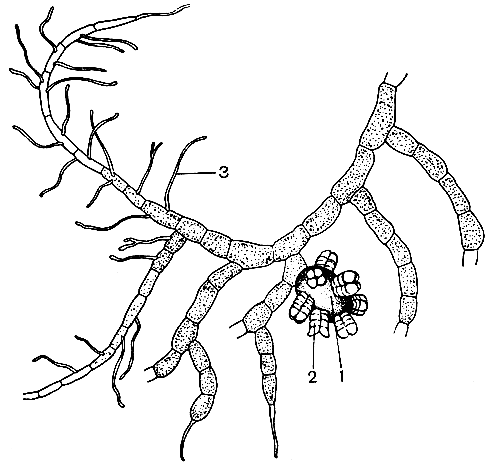
Рис. 132. Нитчатый гаметофит трихомацеса двугубого (Trichomanes bilabiatum): 1 - архегониофор; 2 - архегоний; 3 - ризоиды
Антеридии у лентовидных гаметофитов расположены по краям, а у нитчатых - на концах ветвей. Более крупные и сложные по строению антеридии у гименофилла, а у трихоманеса они в значительной мере вторично упрощены. Архегонии у лентовидных гаметофитов сидят близ точек роста на брюшной стороне таллома, у нитчатых - на специальных ветвях. В гаметофитах гименофилловых, найденных в природных условиях, обнаружена микориза, однако присутствие гриба не является, вероятно, обязательным условием для нормального развития, как, например, у схизейных. Гаметофиты обоих типов строения обильно ветвятся и способны к вегетативному размножению при помощи гемм (почек) (рис. 133), возникающих на геммиферах - клетках-подставках бутылковидной формы. После образования геммы клетка-подставка теряет хлорофилл и гемма отламывается, прорастая в новую нить. Геммы образуются как на молодых, так и на вполне зрелых гаметофитах. У гаметофитов гименофилловых, попавших в неблагоприятные условия, было отмечено усиленное геммообразование.
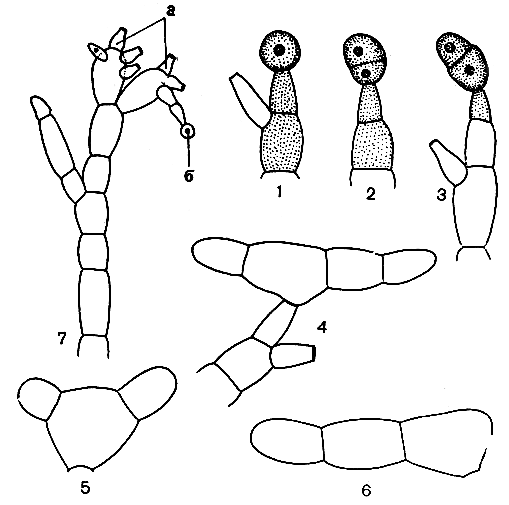
Рис. 133. Размножение гаметофита трихоманеса с помощью гемм: 1, 2, 3 - стадии формирования геммы; 4, 5, 6 - стадии развития из геммы молодой нити гаметофита; 7 - участок нити гаметофита со старыми стеригмами и геммой: а - старые стеригмы; б - развивающаяся гемма
Родственные связи гименофилловых с другими группами папоротников не установлены. Большинство исследователей согласны с тем, что представители этого семейства прошли длительный период независимого развития, в результате чего образовались их столь уникальные морфологические структуры. Кажется вполне вероятным близость гименофилловых к циатейным, а именно - к локсоме (Loxsoma) и локсомопсису (Loxsomopsis) из подсемейства локсомовых.
Семейство асплениевые (Aspleniaceae) (В. Н. Гладкова)
В семейство асплениевых входят около 4000 видов наземных и эпифитных папоротников, населяющих преимущественно влажные местообитания. Их характерными признаками являются: диктиостелические корневища, покрытые чешуями, дорсальные сорусы, снабженные настоящими покрывальцами (индузиями), и билатеральные споры с развитой периной.
Подсемейство асплеииевые (Asplenioideae)
По числу видов в семействе асплениевых доминирует одноименное подсемейство, объединяющее 9-12 родов, среди которых центральным является род асплениум, или костенец (Asplenium), включающий около 700 видов. Представители этого рода распространены практически во всех областях земного шара, но наибольшее их разнообразие наблюдается в тропиках. Род представляют растения очень разного облика, от маленьких скальных папоротничков умеренной зоны, надземная часть которых часто едва достигает 10-15 см высоты, до крупных жестколистных папоротников тропических лесов, с листьями длиной около 2 м. И тем не менее это один из самых четких и естественных родов папоротников, что показывает, в частности, однотипное строение их сорусов и проводящей системы черешков листьев. Сорусы у них удлиненные, линейные или линейно-продолговатые, расположены обычно с одной стороны боковых жилок косо по отношению к средней жилке и прикрыты узким покрывальцем, повторяющим форму соруса (табл. 34). В основании черешков листьев всегда имеются два проводящих пучка, выше сливающиеся в один Х-образный пучок.
Таблица 34. Типы расположения сорусов у папоротников семейства асплениевых: вверху слева - листовик обыкновенный (Phyllitis scolopendrium); вверху справа - гимнокарпиум трехраздельный (Gymnocarpium dryopteris); внизу слева - вудвардия укореняющаяся (Woodwardia radicans); внизу справа - блехнум восточный (Blechnum orientale)
В горных, реже равнинных лесах тропиков виды рода асплениум часто поселяются на стволах деревьев, т. е. являются эпифитами. Как и другие эпифиты они вынуждены развивать различные приспособления, способствующие аккумуляции гумуса, абсорбции воды и защищающие их от избыточного испарения. Среди тропических эпифитов рода асплениум имеется целая группа видов, так называемых папоротников-гнезд, достигших высокого совершенства на пути развития этих приспособлений (рис. 134).

Рис. 134. Асплениум гнездовой, или птичье гнездо (Asplenium слева - папоротник на стволе дерева в тропическом лесу; внизу вверху справа - оранжерейный экземпляр
Из группы папоротников-гнезд наиболее обычен и широко распространен в тропических лесах Старого Света папоротник асплениум гнездовой, или птичье гнездо (Asplenium nidus). Это светолюбивый эпифит, растущий на стволах и ветвях тропических деревьев, но изредка встречающийся также на почве. Поселившись, на дереве, он обычно живет на нем в течение многих лет, пока под его тяжестью не обломится ветвь или пока не погибнет само, дерево. Птичье гнездо имеет толстое прямое корневище, одетое коричневыми чешуями и покрытое; массой: спутанных, сильно опушенных корней. Его кожистые цельные листья длиной до 2 м и шириной до 20, а иногда и до 60 см растут в виде плотной розетки на верхушке корневища и все вместе составляют своеобразную вместительную корзину, в которую падают сверху листья, кусочки коры, пыль. Накапливающаяся гниющая масса органических остатков образует покров над верхушкой корневища, а отходящие от него корни растут сквозь эту массу, получая из нее необходимые питательные вещества. Время от времени верхушка корневища продуцирует новые листья, растущие сначала вертикально, а затем грациозно искривляющиеся. Пронизанные корнями органические остатки при этом прочно удерживаются между основаниями старых и новых листьев. Таким способом растение иногда накапливает столь большое количество гумуса, что в нем поселяются даже дождевые черви. Вся эта масса материала представляет собой эффективную губку, поглощающую дождевую воду в столь больших количествах, что она стекает по стволу долго после того, как дождь кончился, и используется другими растениями (мхами, другими видами папоротников), поселяющимися ниже по стволу на том же дереве, а также папоротниками и орхидеями, часто растущими прямо на массе старых корней этого замечательного эпифита. По данным известного английского птеридолога P. Xолтума, этот папоротник обычно растет там, где суточные колебания температуры незначительны, и наиболее обилен в областях с коротким сухим сезоном.
Папоротники-гнезда представляют собой весьма эффектное зрелище в природе. Их часто выращивают как декоративные растения в тропических странах, а в странах с умеренным климатом они являются весьма обычным компонентом папоротниковых оранжерей.
Среди тропических эпифитов из рода асплениум встречаются и мелкие формы, едва достигающие общей высоты 20-25 см. К ним относится, например, широко распространенный в тропической Африке эпифит влажных лесов асплениум Манна (A. mannii) с перистыми листьями длиной около 10-12 см. Кроме нормальных листьев, этот папоротник продуцирует особые столоновидные, лишенные пластинки листья, на которых на расстоянии 2-6 см друг от друга появляются новые, дочерние растеньица. Многие другие тропические виды рода асплениум растут на скалах или на влажной почве под пологом леса. Их можно встретить в горах и на равнинах, во влажных горных долинах, в затененных и умеренно освещенных местах. Некоторые виды растут на приморских скалах, где они омываются солеными волнами.
В умеренных и холодных областях виды рода асплениум - в большинстве случаев невысокие растения с перистыми или дихотомически раздельными скученными листьями, растущие на валунах, в расщелинах скал и трещинах стен, по каменистым горным склонам, на туфах, серпентинах, кислых и щелочных горных породах, иногда на песках. Горным и скальным видам асплениумов умеренной флоры свойственно вертикальное или короткое ползучее, часто ветвящееся корневище с густой массой корней, уходящих в расщелины скал и камней и прочно удерживающих растение на субстрате. Эти небольшие скальные папоротнички обладают необыкновенным изяществом (рис. 135).

Рис. 135. Папоротники подсемейства асплениевых: 1 - асплениум постенный (Asplenium rutamuraria); 2 - асплениум волосовидный (A. trichomanes); 3 - асплениум северный (A. septentrionale); 4 - цетерах аптечный (Ceterach officinarum); 5 - камптосорус корнелюбивый (Gamptosorus rhizophyllus)
Почти повсюду в Европе, в умеренной Азии и Северной Америке на известняковых скалах и стенах может быть встречен асплениум постенный (A. rutamuraria) - папоротник с коротким ползучим корневищем и дважды (у основания трижды) перистыми черешчатыми листьями длиной 3-15 см. Столь же широко распространен, предпочитая селиться на известняковых породах в горных областях, асплениум зеленый (A. viride). Его перистые листья длиной 5-15 см образуют густую дерновинку на верхушке косого корневища, прочно закрепленного в субстрате. На затененных скалах, на известняковых и кислых горных породах в Европе, Азии, Африке и Северной Америке растет асплениум волосовидный (A. trichomanes) с изящными, длинными (до 30 см), суженными к верхушке, однажды перистыми листьями.
Некоторые виды рода асплениум обнаруживают очень строгую приуроченность к определенным субстратам. Так, асплениум помесный (A. adulterinum) встречается в горах Восточной и Центральной Европы и Скандинавии почти исключительно на серпентинах.
Всего в Европе известно 20 видов асплениума. Около 20 распространены и в СССР. Многие из них способны скрещиваться между собой.
В природе довольно часто встречаются гибридные формы и даже гибридогенные виды.
Сравнительно немногие виды асплениума имеют практическую ценность. Некоторых из них используют в народной медицине как ранозаживляющие, противолихорадочные, болеутоляющие, тонизирующие, вяжущие, при цинге, желтухе и т. д. Однако основное практическое применение асплениум находит как декоративное растение. Особую ценность представляют собой папоротники, обладающие красивыми вечнозелеными кожистыми листьями, долго не увядающими в букетах.
Интересной и практически важной особенностью многих экзотических видов асплениума является способность их листьев к образованию выводковых почек. Широко известен этим свойством растущий в Новой Зеландии и выращиваемый в оранжереях и в комнатах асплениум луковиценосный (A. bulbiferum). На верхней стороне его трижды перистых листьев можно видеть выводковые почки, которые прорастают, еще будучи прикрепленными к материнскому растению. Отделившись от него и попав на влажную почву, они укореняются и дают начало новым растениям. Асплениум луковиценосный относительно быстро растет, не требует большого уяода и весьма популярен в культуре. Реже культивируется асплениум живородящий (A. viviparum) с более тонко рассеченными листьями (рис. 136).

Рис. 136. Асилениум живородящий (Asplenium viviparum): лист с молодыми растеньицами, развившимися из выводковых почек
Вегетативное размножение свойственно не только асплениуму, но и многим другим асплениевым, например камптосорусу, или кривокучнику (Camptosorus). В этот род входят два вида: камптосорусы сибирский (С. sibiricus), растущий на покрытых мхом скалах в Восточной Сибири, на Дальнем Востоке, в Японии, Китае и Корее, и корнелюбивый (С. rhizophyllus) - североамериканский скальный вид, называемый также "странствующим папоротником" (рис. 135). Оба вида вечнозеленые, имеют цельные листья с сильно оттянутой верхушкой, переходящей в длинный жгут. Жгут заканчивается почкой, которая при соприкосновении с субстратом развивается в новое растение. Странствуя таким способом, виды завоевывают себе жизненное пространство.
В систематическом отношении очень близок к роду асплениум род листовик (Phyllitis). Из четырех его видов, распространенных в северном полушарии (Европа, Кавказ, Восточная и Юго-Восточная Азия, Северная Америка), наиболее известен листовик обыкновенный (P. scolopendrium), или папоротник олений язык, названный так за языковидную форму его крупных (длиной до 60 см), ярко-зеленых, глянцевых, цельных листьев (табл. 34, 35). Их нижняя поверхность исчерчена линейными сорусами различной длины. Расположение сорусов у них весьма необычно; они лежат супротивно и плотно друг к другу на двух соседних жилках листа, образуя пары (сдвоенные сорусы). Покрывальца в молодом состоянии слегка налегают друг на друга свободными краями, и вся структура производит впечатление единого соруса. Листовик обыкновенный теневынослив, растет на влажных, затененных скалах, на сырых почвах в укрытых местах и западинах в лесах, иногда встречается на известняках. Он весьма декоративен, и его нередко выращивают в садах. В культуре особенно ценятся уродливые формы этого папоротника с многократно вильчато раздельной верхушкой листа, с сильно волнистым краем листа, а также формы, образующие почки на листьях. Листья листовика используют в народной медицине и в гомеопатии.

Таблица 35. Папоротники: вверху слева - листовик обыкновенный (Phyllitis scolopendrium); вверхусправа - вудсия эльбская (Woodsia) слева - щитовник мужской (Dryopteris filix-mas); внизу справа - полистихум копьевидный (Polystichum lonchitis)
К числу ксерофильных представителей семейства асплениевых принадлежит род цетерах, или скребница (Ceterach), распространенный в горных областях Европы, Азии, Африки, Мадагаскара. Из 2 или 3 видов этого рода наиболее широко известен цетерах аптечный (С. officinarum). Необычны кожистые перистонадрезные листья этого вида с округлояйцевидными или продолговатыми чередующимися долями. Сверху они голые, а снизу сплошь покрыты бурыми черепитчато налегающими, ланцетными, пленчатыми чешуями. Во время длительной сухой погоды листья свертываются таким образом, что снаружи оказываются их нижние, защищенные чешуями поверхности.
В этом подсемействе наблюдается интенсивная гибридизация, сочетающаяся с полиплоидией: здесь нередки 12-плоидные виды, существуют даже 16-плоидные с соматическим числом хромосом 576. Более того, как предполагают японские ученые С. Татуно и С. Каваками, характерное для всего подсемейства основное хромосомное число х=36 само является результатом древней полиплоидии и возникло из первичного числа х=12. Отсюда следует, что самый низкий современный уровень полиплоидии в подсемействе - гексаплоидный. Интересно, что процент полиплоидных видов в этом подсемействе значительно выше в тропических и южных умеренных зонах по сравнению с северными умеренными. Но главным центром видообразования этой группы папоротников являются тропики.
Подсемейство щитовнитовые (Dryopteridoideae)
Это подсемейство охватывает около 500 видов, большая часть которых относится к роду щитовник (Dryopteris) и роду полистихум (Роlystichum).
Род щитовник включает около 150 наземных, главным образом лесных видов. Хотя щитовник широко распространен по всему земному шару, от холодных областей Евразии и Северной Америки до тропиков Азии, Африки и Южной Америки, род этот замечателен тем, что в противоположность многим другим папоротникам большинство его видов растет в умеренной зоне. Центр видового разнообразия рода находится в Гималаях и Восточной Азии (Китай, Япония), где сосредоточено около 100 его видов. Ископаемые остатки щитовника находили уже в меловых отложениях, но его развитие происходило в основном в третичном периоде.
Одним из наиболее красивых и широко известных лесных папоротников является щитовник мужской (Dryopteris filixmas, табл. 35). Его толстое, косо поднимающееся корневище, одетое широкими мягкими чешуями и остатками листовых черешков, несет на верхушке пучок крупных листьев с дважды перистой пластинкой. На сегментах пластинки с нижней стороны видно по 5-8 сорусов, расположенных на разветвлениях жилок (рис. 137) и прикрытых почковидными покрывальцами.
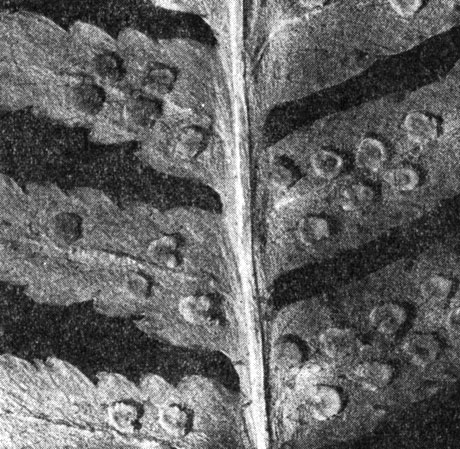
Рис. 137. Щитовник мужской (Dryopteris filix-mas): часть листа с сорусами
Листья щитовника мужского достигают длины 1 и даже 1,5 м, но растут они медленно. Появившись в виде листовых зачатков вокруг точки роста на верхушке корневища, они на второе лето принимают характерную для папоротников улиткообразную форму, располагаясь в виде небольших густо покрытых бурыми чешуями спиралек в самой внутренней части листового пучка. Густой покров из чешуй и свернутое положение листьев защищают меристематическую ткань растущей верхушки листа от повреждений и высыхания. Только весной третьего года молодые листья развертываются и достигают полного развития. Осенью листья увядают. К этому времени щитовник успевает рассеять споры, из которых вырастает сердцевидный гаметофит, покрытый железистыми волосками.
Щитовник мужской освоил обширные территории от суровой Гренландии и Скандинавии до Мексики и Средиземноморья, от Кольского полуострова до горно-лесных районов Кавказа, Средней Азии и юга Сибири. В горах он поселяется в буковых, еловых, пихтовых и арчовых лесах, поднимаясь до альпийского и горно-тундрового пояса. В Арктике растет по защищенным от ветра южным склонам, покрытым зимой мощным снежным покровом. Но основная часть его ареала лежит в лесной зоне, где он встречается в хвойных, смешанных и широколиственных лесах.
Щитовник мужской широко известен как лекарственное растение. Лекарственные свойства этого растения были известны уже врачам античной эпохи и средневековья. О нем упоминается, в частности, в сочинениях Диоскорида и Плиния.
Щитовник мужской как глистогонное средство входит в Государственную Фармакопею СССР. Лекарственное сырье (корневища) заготавливают осенью, и из свежесобранных корневищ получают препарат филиксан. Действующими началами в нем являются производные флороглюцина: филицин, флаваспидиновая кислота, аспидинол и др. Эти вещества вызывают паралич мускулатуры ленточных глистов, которые затем с помощью солевого слабительного выводятся из организма.
Использование этого папоротника в народной медицине значительно шире. Необходимо помнить, однако, что корневище щитовника мужского и извлечения из него ядовиты и при самолечении могут вызывать острое отравление, выражающееся в судорогах, помрачнении сознания, сердцебиении и рвоте (применять эти препараты следует только по назначению врача).
Лекарственные свойства были обнаружены и у других видов щитовника, например у щитовника шартрского, или игольчатого (D. carthusiana), корневища которого также используют как противоглистное средство. Этот вид широко распространен в Евразии и Северной Америке по сырым мшистым и травяным хвойным и смешанным лесам, в горах и в долинах рек, в зарослях кустарников, с которыми он продвигается даже в тундру. Щитовник шартрский отличается меньшими размерами, его трижды, четырежды перистые листья с остроконечными зубчиками на сегментах не превышают в длину 50 см.
Среди немногих видов щитовника, заходящих в арктическую зону, наиболее характерен щитовник пахучий (D. fragrans), встречающийся даже в высокоширотных арктических районах. За пределами Арктики растет в лесной зоне Евразии и Северной Америки. Щитовник пахучий поселяется в трещинах скал различных горных пород, по каменистым россыпям, в кустарниках, предпочитая хорошо прогреваемые южные склоны, на которых иногда образует заросли. Листья сохраняются на растении в течение нескольких лет; они зимуют под снегом в зеленом состоянии. Летом по мере развития молодых листьев самые старые отмирают. Они-то и защищают зимой живые части растения от холода и ветра. Свое название "пахучий" этот вид получил за сильный приятный запах, который он издает в теплую солнечную погоду. Запах продуцируют многочисленные железки на листьях, выделяющие смолистые вещества.
Листья этого вида могут быть использованы в парфюмерии. Кроме того, их широко применяют в народной медицине в виде отваров и настоек как ранозаживляющее, противовоспалительное, болеутоляющее и общеукрепляющее средство.
Сильный запах, напоминающий запах бальзама, свойствен и другим видам щитовника, например европейскому щитовнику Виллара (D. villarii), дважды перистые кожистые листья которого также густо покрыты короткостебельчатыми желтоватыми железками. Лишь небольшое количество видов щитовника встречается в горах тропиков - в лесах и на лесных опушках, вокруг камней и в углублениях скал.
Среди тропических щитовников имеются крупные экземпляры, массивные корневища которых образуют прямой стволик высотой до 15-20 см и диаметром до 8-10 см с торчащими вверх кожистыми листьями на верхушке. Такой облик свойствен щитовнику Валлиха (D. wallichiana), произрастающему в тропической Азии, на Мадагаскаре и в Африке. Он напоминает карликовый древовидный папоротник, так же как и щитовник почти древовидный (D. subarborea) - очень красивое растение с коротким и толстым, поднимающимся над землей корневищем и трижды, четырежды перистыми листьями, пластинка которых достигает длины 150 см. Он растет в Юго-Восточной Азии (Малакка, Калимантан, Суматра).
Некоторые декоративные щитовники введены в культуру; многие из них - прекрасные садовые растения. В европейских садах и оранжереях довольно часто культивируется щитовник Зиболъда (D. sieboldii), родом из Китая и Японии, непарноперистые листья которого заканчиваются крупным, вытянутым сегментом.
Второй род этого подсемейства - полистихум, или многорядник (Polystichum), включает свыше 200 видов, богато представленных, как и виды щитовника, во флоре областей умеренной зоны, особенно в Китае, а также встречающихся и в тропиках. Этот род включает и типично альпийские виды, проникающие в горы до границы вечных снегов.
Виды полистихума - наземные папоротники с короткими восходящими или ползучими корневищами, густо покрытыми разнообразного размера и формы чешуями. Листья их обычно грубые, узкие, кожистые, по краю пильчатые или зубчатые (зубцы заканчиваются щетинкой). Черешки листьев обычно с диморфными чешуями - широкими зубчатыми или бахромчатыми и узкими волосовидными. Сорусы, прикрытые щитовидными покрывальцами, расположены на середине или конце разветвлений боковых жилок и образуют правильные ряды, один или два с обеих сторон срединной жилки сегмента (отсюда название рода - многорядник). В умеренных областях Евразии и Северной Америки распространен полистихум копьевидный (P. lonchitis), скальный горно-лесной вид с толстым коротким корневищем и плотными перистыми листьями, пильчатые сегменты которых сближены и серповидно искривлены вверх. Листья его зимуют под снегом в зеленом состоянии и функционируют в течение нескольких лет.
Широко распространен в Европе полистихум щетинконосный (P. setiferum) с толстым твердым корневищем и дважды перистыми листьями. У нас этот вид встречается в Крыму и в тенистых буковых лесах на Кавказе. Полистихум одетый (P. vestitum), встречающийся в лесах Новой Зеландии, Тасмании и на субантарктических островах к югу от Новой Зеландии, образует стволик высотой до 1 м и напоминает древовидный папоротник.
Несомненное родство щитовника и полистихума подтверждается, в частности, тем, что виды того и другого имеют одно и то же основное хромосомное число: х = 41.
Некоторые тропические роды, относимые к этому подсемейству, обладают диморфными листьями и акростихоидными спорангиями (спорангии равномерно распределены по поверхности фертильных листьев, отличающихся по внешнему облику от стерильных). Это, на пример, род полиботрия (Polybotria) из Южной Америки. Для других родов характерны сорусы без покрывальца (виды рода стигматоптериса - Stigmatopteris). Все это свидетельствует о разных эволюционных направлениях в подсемействе. Сходные тенденции в эволюции, приводящие к диморфизму листьев, а также ко вторичной потере покрывалец, свойственны и другим подсемействам семейства асплениевых.
Подсемейство кочедыжниковые (Athyrioideae)
Близкородственно щитовниковым подсемейство кочедыжниковых, или атириевых, объединяющее свыше 20 родов. В центральный род этого подсемейства кочедыжник (Athyrium) входят около 200 видов наземных папоротников, распространенных главным образом в умеренной зоне северного полушария (лишь немногие виды растут в тропиках). Это довольно крупные, преимущественно лесные растения с дважды, трижды перистыми тонкими листьями, с продолговатыми или искривленными сорусами на них. Корневища их толстые короткие или длинные ползучие, часто ветвистые, одетые непрозрачными чешуями. В основании листовых черешков у кочедыжника, как и у других представителей этого подсемейства, два проводящих пучка, которые выше объединяются, образуя подковообразную структуру.
Типичным представителем рода является кочедыжник женский (A. filix-femina, рис. 138) - характерный папоротник лесной зоны Евразии и Северной Америки. Он населяет влажные леса, луга, берега рек, встречается на болотистых местах. По ивнякам и березовым рощам лесотундры кочедыжник женский заходит в пределы Арктики. Собранные в раскидистый пучок крупные листья с короткими, покрытыми редкими чешуями черешками и дважды, трижды перистой тонкой пластинкой придают декоративный облик этому растению. Изящество его тонко рассеченных светло-зеленых листьев особенно бросается в глаза в сравнении с щитовником мужским, с которым он иногда растет рядом. Это обстоятельство и послужило в средние века причиной появления названий: более груболистный - папоротник мужской, более нежный и изящный - папоротник женский. Названия эти были сохранены, хотя виды эти, по современным представлениям, относятся к разным родам, а понятия "мужской" и "женский" по отношению к растениям, размножающимся спорами, может быть употреблено только в переносном смысле.

Рис. 138. Кочедыжник женский (Athyrium filix-feminа): часть листа с сорусами
На сегментах листьев кочедыжника женского по обе стороны от средней жилки располагаются сорусы, вытянутые вдоль разветвлений жилок или охватывающие их своим изгибом в виде подковы или крючка (рис. 138). Сорусы закрыты такой же формы покрывальцем с бахромчатым краем. На примере кочедыжника женского можно проследить, как почковидный сорус щитовниковых превратился в удлиненный сорус, характерный для некоторых родов кочедыжниковых. Иногда разнообразные по форме сорусы наблюдаются у этого папоротника даже в пределах одного листа.
Большое количество морфологических разновидностей в природе послужило источником для получения множества культурных форм кочедыжника женского, выращиваемых в садах. В некоторых птеридологических справочниках прошлого века зарегистрировано до 300 разновидностей женского папоротника, встречающихся в культуре. Корневища этого папоротника, как и папоротника мужского, широко использовали в народной медицине. Жареные корневища и молодые листья употребляют в пищу западноамериканские индейцы.
Род кочедыжник связан переходными формами с другим, иногда соединяемым с ним представителем этого подсемейства - родом диплазиум (Diplazium), насчитывающим до 400 видов. Виды диплазиума распространены преимущественно в тропических лесах, где они занимают такое же место, как виды кочедыжника в лесах умеренной зоны, т. е. Являются обычнейшими компонентами лесных сообществ. В отличие от кочедыжника, типичные представители которого имеют довольно короткие сорусы, видам диплазиума свойственны длинные, как у асплениумов, расположенные вдоль боковых жилок сорусы с покрывальцем, прикрепленным со стороны жилки. В сравнении с родом кочедыжник, типичные виды диплазиума характеризуются более плотными и менее рассеченными листьями.
К роду диплазиум принадлежит один из наиболее важных в пищевом отношении папоротников тропиков - диплазиум съедобный. (D. esculentum). Он растет в тропиках от Индии до Филиппин и островов Фиджи на влажных открытых местах. Молодые сочные верхушки листьев этого вида употребляют в пищу как зелень или салат.
В умеренных зонах немногочисленные диплазиумы - обычно невысокие (15-25 см) лесные растения, с длинными, тонкими, ползучими корневищами, покрытыми чешуями. По мшистым сырым лесам, каменистым россыпям, речным берегам некоторые из них широко расселились на территории Евразии и Северной Америки.
В роде пузырник, или цистоптерис (Cystopteris), насчитывают немногим более 10 видов, но их малочисленность как бы компенсируется очень широким распространением некоторых из них. Так, пузырник ломкий (С. fragilis) населяет арктические и лесные районы Евразии, Северной Америки, субантарктические районы Южной Америки, встречается в Гренландии, Исландии, в горах Западной Африки, на Тасмании, в Новой Зеландии. В пределах своего обширного ареала он всегда приурочен к районам с умеренно теплым или прохладным климатом, с достаточной влажностью. Растет пузырник ломкий обычно по выходам коренных горных пород (известняки, мергели, сланцы), по расщелинам скал, по речным берегам, в тенистых хвойных лесах. Довольно обычен он и на альпийских высотах в высокогорьях умеренной зоны. Пузырник ломкий имеет тонкое ползучее корневище, покрытое черноватыми остатками старых черешков. Некрупные (длиной 10-15 см) голые листья с дважды, трижды перистой пластинкой расставлены по корневищу. Черешки листьев очень тонкие и ломкие (отсюда название вида). Сорусы округлые, прикрыты мешковидным, слегка вздутым (отсюда название "пузырник") покрывальцем, отгибающимся и засыхающим у зрелых сорусов (рис. 139).

Рис. 139. Кочедыжниковые. Пузырник ломкий (Cystopteris fragilis): 1 - общий вид; 2 - сегмент листа с сорусами. Вудсия эльбская (Woodsia ilvensis): 3 - общий вид; 4 - фрагмент сегмента с сорусом
Широкому распространению некоторых видов пузырника, несомненно, содействует их способность к вегетативному размножению. У пузырника луковиценосного (С. bulbifera), растущего в Северной Америке во влажных лесах, по оврагам и на известняковых скалах, на нижней стороне листьев образуются почки сферической формы, напоминающие луковички. Почки отваливаются и прорастают в новые спорофиты, позволяя этому папоротнику не только образовывать большие заросли на одном месте, но и завоевывать новые территории.
К роду вудсия (Woodsia) также принадлежат небольшие горные и скальные папоротнички с горизонтальными короткими корневищами, закрепляющимися в трещинах скал и камней. Зрелые листья видов вудсии варьируют длиной от 3 до 60 см. Они обычно узкие, по консистенции довольно плотные, пластинка у разных видов от однажды до трижды перистой (табл. 35). Черешки листьев многих видов имеют в разной степени выраженное сочленение в виде кольцеобразного рубчика, что является приспособлением к листопадности. Все листья скучены на конце корневища, на которому ряда видов сохраняется большое количество оснований черешков отмерших листьев, образующих у них плотную "щеточку". Сорусы сидят на концах жилок и прикрыты нижним (прикрепленным под сорусом) покрывальцем, форма которого варьирует от группы волосовидных долек, окружающих почти плоское ложе соруса, до крупной блюдцевидной структуры, прикрывающей сорус сверху. В зрелом состоянии сорусы полностью закрывают нижнюю поверхность листьев, напоминая буроватый войлок.
В роде вудсия насчитывается около 25 видов, произрастающих по всему земному шару, кроме Австралии и Антарктики. Около 15 видов этого рода, в том числе многие примитивные, растут в Азии (Гималаи, Тибет). Эта область, очевидно, является центром происхождения рода.
К видам, широко распространенным в северной умеренной зоне и заходящим в Арктику, относится вудсия эльбская (W. ilvensis) - невысокий папоротник, густо покрытый бурыми пленками и волосками. Ее можно встретить по замшелым расщелинам скал, на выходах различных горных пород (табл. 35), по коренным берегам рек. Корневища этого папоротника иногда образуют плотные сплетения, и тогда колонии растения полностью закрывают верхушки сухих скал и валунов. Вудсия эльбская встречается также по каменистым склонам гор среди гольцов, где является одним из характернейших папоротников, по крутым обрывам и россыпям, по гребням и склонам тундровых гряд и холмов.
Хорошо знаком жителям умеренных областей род гимнокарпиум, или голокучник (Gymnocarpium), занимающий несколько обособленное положение в этом подсемействе. В отличие от большинства других видов кочедыжниковых виды гимнокарпиума не имеют покрывальца. Отличает его от других родов, кроме того, отсутствие заметной перины на спорах, но, как и многие другие кочедыжниковые, он имеет два свободных проводящих пучка в основании черешков листьев и хромосомное число n = 40.
К роду гимнокарпиум принадлежат 5-8 видов, распространенных в северном полушарии. Это небольшие высотой (15-30 см) лесные и скальные растения с одиночными, расставленными по ветвящемуся корневищу листьями. Листья гимнокарпиума очень нежные, небольшие, на зиму опадают.
Одним из распространенных лесных папоротников Евразии и Северной Америки является гимнокарпиум трехраздельный (G. dryopteris, рис. 140), известный также под названием папоротника Линнея. Его всегда легко узнать по своеобразной форме листьев: пластинка их разделена на 3 почти равные части, сидящие на длинных черешочках, которые имеют заметные сочленения в местах соединения с главной осью. Лист в целом имеет форму равностороннего треугольника. Корневище блестящее, черное, иногда разрастается на значительной площади, благодаря чему этот папоротник встречается большими группами в хвойных и широколиственных лесах. В ельниках это нередко даже фоновое растение травянистого яруса.
Рис. 140. Кочедыжниковые: слева - гимнокарпиум трехраздельный (Gymnocarpium dryopteris); справа - фегоптерис связывающий (Phegopteris connectilis)
У большинства папоротников умеренной зоны листья, продуцирующие споры, не отличаются или мало отличаются по внешнему облику от стерильных листьев. Роды же страусник (Matteuccia) и оноклея (Onoclea), также относимые к этому подсемейству или иногда рассматриваемые как самостоятельное подсемейство и даже семейство, принадлежат к тому меньшинству папоротников, спороносные листья которых обнаруживают значительную редукцию размеров пластинки и резко отличаются от стерильных.
Род страусник состоит из 2-3 видов, произрастающих в северном полушарии. Относящийся к этому роду страусник обыкновенный (Matteuccia struthiopteris) - один из наиболее красивых и часто культивируемых в открытом грунте видов папоротников. Стерильные листья его, перистые с перистонадрезными сегментами, длиной до 1,7 м, образуют правильной формы воронку, которая окружает пучок появляющихся позднее и более коротких спороносных листьев. По форме однажды перистые спороносные листья напоминают страусовое перо, за что и дано название рода. Края сегментов спороносных листьев свернуты до средней жилки, что обеспечивает защиту расположенным на концах разветвлений жилок округлым сорусам. Спорофиллы страусника сначала светло-зеленые, затем становятся темно-коричневыми, контрастируя своей окраской со светло-зелеными стерильными листьями. Осенью стерильные листья увядают, а спороносные остаются зимовать. Их темно-коричневые верхушки иногда можно видеть торчащими над снежной поверхностью. Весной освобождаются споры, края листьев при этом разворачиваются.
Страусник широко распространен в умеренной зоне северного полушария по болотам и влажным, покрытым лесом склонам, по тенистым берегам рек и ручьев, в пойменных лесах. Молодые листья этого папоротника в некоторых странах употребляют в пищу как овощи.
Единственный вид рода оноклея (Onoclea), оноклея чувствительная (О. sensibilis) имеет диморфные листья, стерильные опадающие, с длинными черешками и перисторассеченной пластинкой и фертильные зимующие, пластинка которых много короче пластинки стерильных листьев, дважды перистая с очень характерными четковидными сегментами второго порядка (рис. 141). Шаровидная форма этих сегментов достигается путем заворачивания внутрь их краев. Стерильные листья оноклеи чернеют с наступлением холодов (отсюда название вида "чувствительная"). В отличие от страусника, характеризующегося прямым мощным корневищем, оноклея имеет длинное и тонкое ползучее корневище, и фертильные листья на нем располагаются рядом со стерильными. Современный ареал этого рода охватывает восток Северной Америки и Восточную Азию, включая советский Дальний Восток. Оноклея растет по сырым лугам, болотам, в приречных лесах. Ископаемые остатки оноклеи, найденные в третичных флорах Западной Европы, Шотландии, Восточного Казахстана, Туркмении, Сибири, Северной Америки, говорят о значительно более широком распространении этого рода в прошлом. Еще в конце третичного периода (в плиоцене) или в начале четвертичного оноклея обитала в бассейне Северной Двины.

Рис. 141. Кочедыжниковые. Оноклея чувствительная (Onoclea sensibilis): 1 - спороносный (слева) и стерильный листья; 2 - фрагмент сегмента листа с сорусами. Страусник обыкновенный (Matteuccia struthiopteris): 3 - спороносный и стерильный листья; 4 - фрагмент спороносного листа; 5 - часть сегмента с сорусами
Роды страусник и оноклея приобрели специализированные черты в строении спор. Крупные, с периной споры этих папоротников зеленые, фотосинтезирующие, имеющие заметные хлоропласты. Наличие сформированных хлоропластов позволяет спорам прорастать почти немедленно после рассеивания. Споры рассеиваются всегда весной, когда имеется достаточная влажность для их прорастания, хотя закладываются и формируются они в предыдущий сезон.
Подсемейство элафоглоссовые (Elaphoglossoideae)
В состав трех родов, составляющих это подсемейство, входят обитатели тропических и субтропических областей. Среди преимущественно наземных растений семейства асплениевых виды элафоглоссовых являются исключением: они в большинстве своем эпифиты.
Большая часть видов этого подсемейства составляет один крупный род элафоглоссум (Elaphoglossum), широко распространенный в тропических и субтропических районах земного шара, но особенно обильно представленный в тропиках Нового Света. Из 400 его видов около 350 произрастает в тропической Америке. Особенно много своеобразных, часто эндемичных видов элафоглоссума в Андах, где они поднимаются в горы до высоты 3400 м.
Виды элафоглоссума имеют восходящие или короткие ползучие корневища, способные также взбираться по скалам. Их листья, большей частью цельные, имеют сочленение с особыми выростами корневища, на которых они образуются. У многих видов, растущих в крайних условиях существования, листья толстые и кожистые, часто покрытые чешуями и волосками, что уменьшает интенсивность транспирации. Фертильные листья, обычно имеющие редуцированную пластинку и отличающиеся от стерильных, с нижней стороны густо покрыты спорангиями. На молодых листьях сначала образуются округлые сорусы, но по мере роста листа сорусы сливаются вместе и возникает акростихоидный тип спорангиев.
Характерной чертой многих видов элафоглоссума, как и других представителей подсемейства, является наличие парафиз - стерильных образований в сорусе, перемешанных со спорангиями и играющих, по-видимому, защитную роль. Установлено, что эпифитные виды, более подверженные воздействию изменяющихся условий среды, имеют парафизы чаще, чем наземные.
Споры элафоглоссума билатеральные, с периной сложной и разнообразной структуры. Вырастающие из споры гаметофиты относятся к числу медленно развивающихся, но долго живущих. На ранних стадиях они имеют сердцевидную форму, а затем в течение нескольких месяцев или даже лет сильно удлиняются, приобретая лентовидную форму, часто с волнистыми или кудрявыми краями, усаженными многочисленными ризоидами. Антеридии и архегонии появляются на гаметофите только тогда, когда он достигает возраста 4-8 месяцев, а в некоторых случаях даже одного года. Такое позднее появление половых органов связано, вероятно, с замедленным развитием гаметофита, а длительный рост последнего является следствием существования в ровном климате тропической зоны.
Другие роды этого подсемейства, насчитывающие всего по нескольку видов,- эпифитный папоротник пельтаптерис (Peltapteris) и наземный микростафилла (Microstaphylla) - отличаются от элафоглоссума главным образом рассеченными в разной степени листьями. Род пельтаптерис имеет, кроме того, и небольшие отличия в строении гаметофита, который лишен волосков, свойственных гаметофитам элафоглоссума и большинству других родов асплениевых, и в его развитии нет сердцевидной стадии.
Подсемейство ломариопсисовые (Lomariopsidoideae)
Подсемейство ломариопсисовых по своим морфологическим свойствам и основному хромосомному числу (х = 41) близко к предыдущему и иногда с ним объединяется. Шесть-семь родов и 165 видов этого подсемейства распространены в тропических районах земного шара. Виды ломариопсисовых характеризуются дорсивентральными корневищами с двумя или несколькими рядами листьев на их дорсальной стороне и акростихоидными спорангиями.
Близкородственные роды болбитис (Bolbitis), включающий 86 видов, и эгенолфия (Egenolfia), состоящая из 11 видов, произрастают главным образом в тропической Азии в пределах Индо-Малайской области. Они заселяют скалы вблизи лесных рек и ручьев, иногда могут забираться на небольшую высоту по стволам деревьев или другой опоре. Это маленькие или среднего размера папоротники с диморфными, обычно перистыми листьями. Фертильные листья рода болбитис имеют длинные черешки и узкие сегменты. У стерильных листьев сегменты более широкие, а терминаль-ный сегмент часто ремневидно или жгутовидно вытянут и образует на верхушке выводковые почки. Иногда они образуются и на боковых сегментах.
Один из видов болбитиса - болбитис Хедело (Bolbitis heudelotii) - известен как аквариумное растение. Он был ввезен в Европу впервые в 1959 г. По свидетельству известного аквариумиста М. Д. Махлина, этот папоротник с красивыми перистыми листьями и ползучим корневищем, покрытым золотисто-желтыми или темно-коричневыми чешуйками, является великолепным, хотя пока еще и редким, украшением аквариумов. На родине, в тропической Западной Африке он поселяется на гниющих стволах деревьев в прибрежных лесах, и, кроме того, растет в ручьях и реках на глубине не более 1 м.
Виды эгенолфии - обитатели главным образом горных областей и, как и виды рода болбитис, приурочены к скалистым и каменистым субстратам вблизи водных потоков в густых лесах.
У ломариопсисовых наблюдается большая вариабельность листьев. Так, у эгенолфии, кроме фертильных и стерильных листьев, могут возникать листья смешанного типа, с чередующимися спороносными и стерильными сегментами. Тенденция к гетерофиллии в подсемействе наиболее выражена в роде тератофиллум (Teratophyllum), виды которого, кроме нормальных перистых стерильных и фертильных листьев (акрофиллов), всегда образуют у основания корневищ особые, обычно дважды перистые листья - батифиллы. К роду тератофиллум принадлежат 12 видов лазящих папоротников, произрастающих в Юго-Восточной Азии, Новой Каледонии, Австралии (Квинсленд) и Полинезии. Начинают свою жизнь виды тератофиллума на почве или у самого основания стволов деревьев в очень влажных и тенистых лесах. Сначала это небольшие папоротники с тонкими корневищами, несущими очень маленькие листья. По мере роста корневище начинает забираться по стволу дерева, цепляясь за его кору с помощью корней и образуя обычно прижатые к стволу батифиллы. Достигнув более значительной высоты, корневище продуцирует листья совсем иного облика - распростертые горизонтально, перистые, более жесткие и крупные. Еще выше, на высоте 10-15 м, могут появиться фертильные (с очень узкими сегментами) листья, образованию которых особенно способствует сухая погода. Расположение спороносных листьев на большой высоте благоприятно для рассеивания спор, которое будет тем успешнее, чем выше находятся спорофиллы.
Другой характерной особенностью листьев тератофиллума является наличие сочленения у сегментов листа со стержнем, позволяющее растению при дефиците влаги с легкостью сбрасывать часть листьев для уменьшения испаряющей поверхности.
Сходны с тератофиллумом по характеру роста два других рода этого подсемейства - ломаграмма (Lomagramma), 15 видов которой произрастают в тропиках Старого Света, и пантропический род ломариопсис (Lomariopsis), включающий около 40 видов. Эти папоротники также укореняются в почве, а затем поднимаются по деревьям на высоту до 15 м и более. Гаметофиты ломаграммы вырастают обычно на влажных скалах, и молодые растения с прямыми длинночерешковыми листьями-батифиллами могут долго ползти по скалистому субстрату, пока не встретят дерево, ствол которого смогут использовать как опору для подъема вверх.
Подсемейство телиптерисовые (Thelypteridoideae)
Своеобразную группу, иногда рассматриваемую как самостоятельное семейство, составляют телиптерисовые, широко представленные в тропических и субтропических странах обоих полушарий, где сосредоточено около 900 видов. Только 1% видового состава подсемейства свойствен умеренной зоне, а во флоре Европы насчитывается всего 5 видов.
Телиптерисовые хорошо отличаются от других папоротников наличием одноклеточных игловидных волосков на верхней стороне рахиса и двух проводящих пучков в черешке, которые сливаются у основания пластинки. Разделение этой группы папоротников на роды представляет, напротив, значительные трудности для птеридологов из-за большого количества признаков, разным образом сочетающихся у этих папоротников. Современные птеридологи приходят к диаметрально противоположным решениям. Одни объединяют почти все виды телиптерисовых в один крупный род телиптерис (Thelypteris), другие разбивают его на большое число мелких родов, характеризующихся общим основным хромосомным числом (в подсемействе в целом оно варьирует от 27 до 36) или одинаковым типом строения спор, опушения, жилкования, наличием или отсутствием редуцированных сегментов у основания листа и т. д.
Телиптерисовые - наземные растения с ползучими или вертикальными корневищами и перистыми (до трижды перистых), обычно кожистыми листьями с округлыми или удлиненными сорусами на простых или вильчатых боковых жилках. Характерной особенностью листьев многих тропических видов телиптерисовых является наличие у них по обеим сторонам рахиса и черешка аэрофоров - полосок особой тонкостенной ткани. Участок такой ткани имеется, кроме того, у основания каждого листового сегмента. Аэрофоры выполняют дыхательную функцию, на их поверхности расположены устьица, а в подстилающей их ткани имеются хорошо развитые межклетники. Аэрофоры на молодых листьях хорошо заметны, так как они светлые, вследствие наличия в них воздуха. У некоторых тропических видов, обитающих в очень влажных условиях, аэрофоры у основания сегментов выпуклые. Листья этих видов в молодом состоянии покрыты слизью. Прорастающие сквозь слизь аэрофоры, видимо, уменьшают трудности газообмена.
Важнейшими тропическими родами телиптерисовых (при дробной классификации подсемейства) являются гониоптерис (Goniopteris) в тропической Америке, сферостефанос (Sphaerostephanos), мезофлебион (Mesophlebion) и корифоптерис (Coryphopteris) в Азии и на островах Тихого океана, а также циклосорус (Cyclosorus) с небольшим количеством видов в тропиках обоих полушарий.
В умеренных флорах северного полушария наиболее часто встречается фегоптерис связывающий (Phegopteris connectilis, или Thelypteris phegopteris). Это изящный лесной папоротник с тонким ползучим корневищем, снабженным светло-коричневыми ланцетными чешуями. По корневищу расставлены заостренные листья с длинными рассеянно-волосистыми черешками и треугольной пластинкой, с отклоненной нижней парой крупных перисторассеченных сегментов. Округлые сорусы, лишенные покрывальца, располагаются близко к краю сегментов. Растет фегоптерис связывающий часто в хвойных, смешанных и широколиственных лесах, встречается в долинах рек, среди обломков скал, на горных луговинах и на гольцах.
Другие местообитания занимает телиптерис болотный (Т. palustris), растущий по окраинам торфяных и осоковых болот, по заросшим лесом берегам рек, на сырых лугах и в заболоченных лесах. Иногда он принимает участие даже в образовании сплавин. Среди болотных папоротников это наиболее широко распространенный вид. Он отличается от предыдущего узколанцетной, суженной к основанию пластинкой листа, сорусы на которой в молодом состоянии закрыты покрывальцем и расположены примерно посредине между средней жилкой и краем сегмента.
Подсемейство блехновые (Blechnoideae)
Подсемейство блехновые, или дербянковые, объединяет 6-11 родов наземных, иногда лазящих по скалам и деревьям, или очень редко эпифитных папоротников, большинство которых обитает в южном полушарии. Виды из этого подсемейства имеют линейные сорусы, которые располагаются в 1-3 ряда параллельно и близко к средней жилке сегмента листа, иногда сливаясь и образуя непрерывный ценосорус. Пленчатые покрывальца сорусов также линейные и открываются со стороны жилки.
Род блехнум, или дербянка (Blechnum), с большим разнообразием видов (свыше 200), распространен преимущественно в тропических и субтропических, реже в умеренных областях южного полушария. Только несколько его видов заходят в северное полушарие, и из них один достигает северной умеренной зоны. У большинства видов блехнума обычно толстое, восходящее или вертикальное, иногда древеснеющее корневище и перистые, изоморфные или диморфные листья с ценосорусами вдоль средних жилок. Корневище часто образует надземный стволик высотой до 1,5 м, и листья тогда собраны на нем в плотную розетку. Такие папоротники с блестящими листьями напоминают по внешнему облику виды саговников. К числу древовидных блехнумов принадлежат блехнум бразильский (В. brasiliense), растущий в Перу и Бразилии. Вместе со своими глубоко-перистораздельными листьями он может достигать высоты 4 м (табл. 36).

Таблица 36. Папоротники: вверху слева - блехнум бразильский (Blechnum brasiliense); вверху справа - адиантум трапециевидный (Adiantumtrapeziforme); внизу слева - страусник обыкновенный (Matteucciastruthiopteris); внизу справа - орляк обыкновенный (Pteridium aquilinum)
Стволик высотой от 50 см до 1 м имеет блехнум горбатый (В. gibbum), растущий в Новой Каледонии и на Новых Гебридах, и многие другие виды этого рода. Древовидные блехнумы часто культивируются в оранжереях, а также в открытом грунте как фоновые и бордюрные растения. Привлекательность их усиливается благодаря свойственной многим блехновым розовой окраске молодых листьев, а также способности продуцировать на них выводковые почки.
Совсем иной облик имеет некрупный травянистый папоротник блехнум колосистый (В. spicant), произрастающий в Северной Америке (от Аляски до Калифорнии) и в Евразии, главным образом в темнохвойных лесах и на лугах. В СССР он встречается на Кавказе, в Карпатах и в некоторых районах Прибалтики. Блехнум колосистый называют иногда оленьим папоротником, так как его вечнозеленые листья служат зимой кормом оленям, лосям, карибу. Косое короткое корневище блехнума колосистого несет диморфные листья. Стерильные листья кожистые, вечнозеленые, перистые, длиной 8-70 см, образуют раскидистую, прижатую к земле розетку, в центре которой располагаются прямостоячие длинно-черешковые и более длинные фертильные листья, сегменты которых редуцированы до ширины 1-2 мм и практически полностью заняты линейными сливающимися сорусами. В отличие от стерильных фертильные листья недолговечны, они отмирают к осени. Блехнум колосистый в некоторых странах Европы относится к числу редких и охраняемых растений.
Некоторые другие виды блехнума также являются травянистыми растениями. Но более характерны для этого подсемейства древовидные папоротники. Древовидный облик имеют почти все виды небольшого рода садлерия (Sadleria). Из четырех видов этого рода, встречающегося только на Гавайских островах в лесах и по крутым прибрежным склонам, один вид - садлерия циатеевидная (S. cyatheoides) - замечателен своей способностью поселяться на отвердевших потоках лавы. Появившись единичными экземплярами, выросшими из спор, растения со временем заселяют на лавах большие пространства. Аборигенному населению Гавайских островов древовидные садлерии и другие древовидные папоротники служили источником пулу - шерстистой массы, состоящей из мягких волосовидных чешуек, которые в изобилии образуются вокруг растущей верхушки и у основания черешков листьев этих папоротников. Пулу использовали при бальзамировании для заполнения внутренних полостей тела. В XIX в. пулу даже экспортировали с Гавайских островов как материал для набивания подушек и матрацев.
Среди блехновых имеются и настоящие лианы. К ним относится близкий к блехнуму род салпихлена (Salpichlaena), с единственным видом салпихленой вьющейся (S. volubilis), растущей в Южной Америке и Вест-Индии. Она забирается на значительную высоту на деревья в тропических лесах с помощью своих дважды перистых, длиной до 1 м листьев, черешок и рахис которых обвиваются вокруг опоры.
Другим способом достигает той же цели стенохлена болотная (Stenochlaena palustris) - представитель небольшого рода стенохлена (Stenochlaena), пять видов которого распространены в тропиках Старого Света. Стенохлена болотная начинает свою жизнь на светлых или умеренно затененных, но достаточно влажных местах и часто образует заросли на почве. Обладая длинным ползучим корневищем, она в конце концов забирается на деревья (часто до самой их вершины), укрепляясь на них с помощью придаточных корней. Сочные молодые листья стенохлены иногда употребляют в пищу как овощ, а ее стебли использовали в прежние времена для изготовления корабельных канатов.
Преимущественно в северном полушарии лежит ареал еще одного блехноидного папоротника - рода вудвардия (Woodwardia) с 12 видами в Гималаях, Китае, Японии, на Филиппинах, Яве, в Новой Гвинее, в Северной Америке. Один вид ее встречается в Южной Европе и на Азорских островах. В геологическом прошлом этот род был распространен еще шире. Ископаемые остатки его представителей найдены во многих областях Европы, Северной Азии, в Арктике.
Виды вудвардии - крупные наземные папоротники с короткими восходящими или длинными ползучими корневищами. Листья у них однажды или дважды перистые или перистораздельные, диморфные или изоморфные. Отличительной особенностью их являются анастомозирующие жилки, образующие ареолы вдоль средней жилки, а по направлению к краю листа свободные. По этим ареолам в погружениях лежат ровные ряды сорусов. Характерным представителем рода является вудвардия укореняющаяся (W. radicans), названная так за способность ее молодых листьев продуцировать вегетативно новые растения. Этот папоротник с перистыми листьями длиной до 2,5 м имеет разорванный ареал: растет в тихоокеанской части Северной Америки до Гватемалы, в Южной Европе и Восточной Азии. Внетропические виды вудвардии часто обладают длинными ползучими корневищами и являются растениями влажных местообитаний, а некоторые из них, как североамериканский вид вудвардия вирджинская (W. virginica), стали настоящими болотными растениями. Вудвардия вирджинская часто поселяется в сфагновых болотах, образуя обширные сплетения своими корневищами и становясь вместе с другими болотными растениями торфообразователем.
Семейство даваллиевые (Davalliaceae) (Т. Д. Сурова)
Среди тропических эпифитных папоротников, особенно в Старом Свете, обычны представители даваллиевых. Любители комнатных и оранжерейных растений, вероятно, знакомы с такими широко распространенными в культуре папоротниками, как даваллия (Davallia), популярное название которой - папоротник "заячьи (беличьи) лапки", и нефролеписом, или меч-папоротником (Nephrolepis), многочисленные садовые формы которого выращивают в оранжереях всего мира. В естественных условиях виды этого семейства населяют леса преимущественно тропических стран. Незначительное число видов встречается в умеренной зоне. Виды рода хумата (Humata) и некоторые нефролеписы достигают севера Японии, а даваллия канарская (Davallia canadensis) обитает на Канарских островах, в Макаронезии, Марокко и на Пиренейском полуострове.
В состав семейства входят 12 родов, насчитывающих в общей сложности около 230 видов. По морфологическим признакам даваллиевые четко подразделяются на две большие группы, которые рассматриваются либо как подсемейства, либо как самостоятельные семейства.
Подсемейство даваллиевые (Davallioideae) объединяет 7-8 родов, близкородственных роду даваллия: гимнограммитис (Gymnogrammitis), даваллодес (Davallodes), хумата, араиостегия (Araiostegia), леукостегия (Leucostegia), трогостолон (Trogostolon) и сцифулярия (Scyphularia). Все они, за исключением леукостегии, типичные эпифиты средних (около полуметра) размеров. Самый маленький папоротник в подсемействе - хумата крошечная (Humata parvula) - не превышает в длину 5 см, а наиболее крупный - леукостегия погруженная (Leucostegia immersa) - достигает высоты 1,5 м.
Папоротники подсемейства даваллиевых имеют длинные ползучие дорсивентральные корневища. На их верхней стороне в два ряда на значительном расстоянии друг от друга располагаются листья. Мясистые, толщиной с палец, густо покрытые рыжеватыми чешуйками, корневища способны запасать в своих тканях некоторое количество воды. Проводящая система всех даваллиевых диктиостелическая, но встречается ряд типов стел, уклоняющихся от обычной диктиостелы. Они возникли в связи с дорсивентральностью корневищ.
Листья видов папоротников в подсемействе перисторассеченные, но у некоторых видов хуматы пластинка листа цельная. Опушение, как правило, хорошо заметно на молодых листьях, с возрастом оно может сохраниться, но чаще исчезает. Черешки листьев сочленяются с особыми выростами корневища - филлоподиями. Они или мало выражены и сравнительно короткие у даваллий, или достигают значительной длины (от 2 до 9 см) у олеандры волнистой (Oleandra undulata, рис. 142), принадлежащей к подсемейству олеандровых (Oleandroideae). Наличие сочленения между листом и корневищем играет большую роль в жизни даваллиевых. Благодаря этому свойству даваллиевые являются листопадными. Смена старых листьев на новые происходит путем поочередного, а в некоторых случаях одновременного сбрасывания всех листьев. Последнее особенно важно для растений, обитающих в странах с сезонным климатом. Некоторые даваллии сбрасывают свои листья с наступлением сухого периода и в таком виде ожидают начала новых дождей.
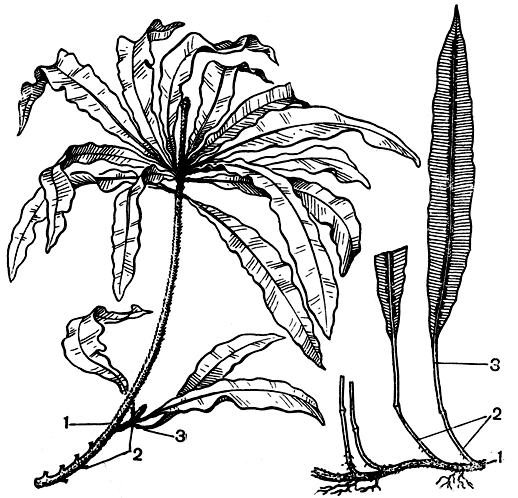
Рис. 142. Олеандры: слева - олеандра пестиковая (Oleandra pistillaris); справа - олеандра волнистая (О. undulata): 1 - корневище; 2 - филлоподии; 3 - черешок
Листья с сочленением - явление довольно редкое, но не уникальное среди папоротников; они имеются у ряда видов из полиподиевых и асплениевых. Однако анатомическое строение черешка на границе сочленения и способ формирования разделительного слоя у даваллиевых и полиподиевых различны.
Благодаря листопадности и мясистым, запасающим воду корневищам даваллиевые легко переносят значительную засуху. Почти все виды этого семейства - эпифиты верхнего яруса леса. Даваллия крыночковидная (D. руxidata, рис. 143) иногда живет на других светолюбивых эпифитных папоротниках, в частности на некоторых видах платицериума.
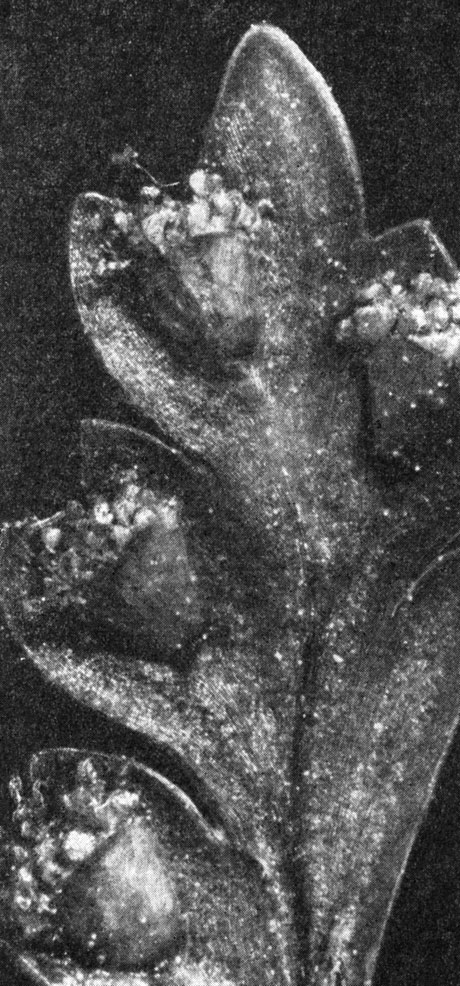
Рис. 143. Даваллия крыночковидная (Davallia pyxidata): часть листа с сорусами (сильно увел.)
Как правило, эпифитные папоротники имеют более или менее ксерофитный облик (небольших размеров плотные кожистые листья, менее рассеченные, нежели листья папоротников, обитающих в местах с достаточным увлажнением). Наиболее ксерофильные папоротники в подсемействе даваллиевых - виды рода хумата.
Род хумата объединяет 50 видов, распространенных на Мадагаскаре, в тропической Азии, Океании. Это небольшие, часто наскальные папоротники с чешуйчатыми ползучими корневищами и сочлененными с ними простыми или слаборассеченными кожистыми листьями. Часто спороносные листья рассечены значительно сильнее, чем вегетативные. Листья некоторых видов хуматы скручиваются в сухую погоду и раскручиваются при повышении влажности. Хумата крошечная и ряд других эпифитных видов поселяются на старых мангровых деревьях, некоторые виды встречаются преимущественно на открытых скалах по берегу моря. У хуматы, как и у большинства представителей подсемейства даваллиевых, сорусы располагаются на концах жилок и защищены кармашковидным покрывальцем (рис. 144), которое прикреплено основанием к пластинке листа; боковые его стороны и верхняя часть свободны. У леукостегии оно прикреплено основанием и сбоку примерно до своей половины, а у даваллии остается свободным только верхний край, и из образовавшегося "кармашка" высовываются созревшие спорангии на длинных ножках.
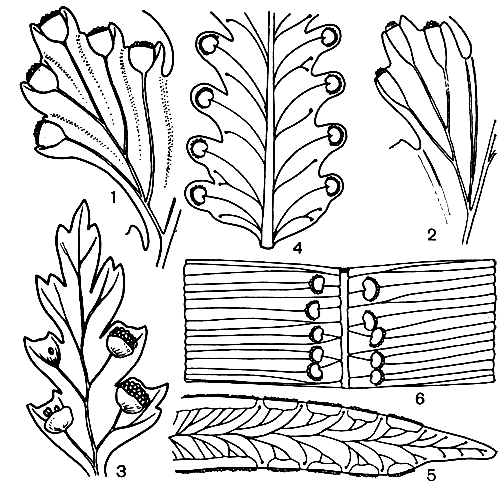
Рис. 144. Схема расположения сорусов у папоротников семейства даваллиевых: 1 - даваллия мелкозубчатая (Davallia denticulata); 2 - даваллия плотная (D. solida); 3 - хумата ассамская (Humata assamica); 4 - нефролепис остроконечный (Nephrolepis acuminata); 5 - нефролепис остролистный (N. acutifolia); 6 - олеандра волнистая (Oleandra undulata)
В род даваллия включают 40 или 50 видов, широко распространенных в тропиках. Даваллия, как и все роды подсемейства, обитает только в странах Старого Света. Все виды даваллии эпифиты, и те немногие, которые можно встретить на почве, имеют, по-видимому, эпифитное происхождение. Кроме способа прикрепления покрывальца, даваллия отличается от хуматы только большими размерами и более рассеченной пластинкой листа. Но если учесть, что хумата покрытая (Humata vestita) имеет такие же размеры, как и самая маленькая из давалдий - даваллия трихоманесовая (Davallia trichomanoides), а степень рассеченности листа очень варьирует, то единственным более или менее надежным признаком, разделяющим эти роды, является форма покрывальца.
В оранжереях часто выращивают даваллии мелкозубчатую (D. denticulata), распростертую (D. divaricata), канарскую, плотную (D. solida) и др. Много лет известна в культуре даваллия Мариса (D. mariesii), в диком виде встречающаяся в Японии. Корневища этого папоротника в больших количествах Япония экспортирует в другие страны мира. Мясистые корневища, густо покрытые бурыми чешуями, связывают проволокой, придавая им форму тела обезьянки. Затем их снабжают маленькими фарфоровыми обезьяньими головками, и в таком безлистном состоянии транспортируют для продажи. Прибывших "обезьянок" подвешивают в оранжереях или в зимних садах и начинают обильный полив, после чего на них вскоре появляются листья.
Но основным способом является размножение при помощи спор. Спорангии даваллии смешанные, на длинных (4-5 клеток в длину) ножках, без парафиз. Они снабжены кольцом из 11-16 толстостенных клеток. Споры всех даваллиевых бобовидной формы, билатеральные, монолетные. У папоротников подсемейства даваллиевых перина не обнаружена. По мере созревания спорангии открываются, освобождая споры, которые для успешного прорастания должны попасть на соответствующий субстрат, обеспечивающий им надежную защиту и достаточную влажность. Прекрасным местом для развития спор и гаметофитов многих папоротников являются стволы масличной и кебонговой пальм, медленно гниющие остатки листьев которых обеспечивают всем необходимым и гаметофиты и молодые спорофиты. Чаще других видов поселяется на пальмовых стволах даваллия мелкозубчатая, которая является своеобразным эпифитным сорняком на плантациях масличной пальмы в Африке и Индонезии.
Попав на подходящий субстрат, спустя 6-7 дней споры даваллии мелкозубчатой прорастают. Зрелые гаметофиты тонкие, вытянутые, сердцевидные (рис. 145). На брюшной стороне имеются одноклеточные ризоиды, мягкие, поначалу бесцветные, с возрастом коричневеющие. Антеридии появляются по краю гаметофита или недалеко от него. Они становятся заметными в трехмесячном возрасте. Архегонии возникают на брюшной стороне спустя две недели после появления антеридиев. Через 3-5 дней после оплодотворения начинается развитие зародыша.

Рис. 145. Развитие гаметофита даваллии мелкозубчатой (Davallia denticulata): 1, 2, 3 - стадии развития нитчатой формы гаметофита; 4, 5 - формирование лопатчатого гаметофита; 6 - зрелый гаметофит с несколькими архегониями и антеридиями; 7, 8 - стадии развития архегония; 9, 10, 11, 12, 13 - стадии развития антеридия; 14 - молодой спорофит на гаметофите
Кроме перечисленных родов, к подсемейству даваллиевых некоторые ботаники относят род румора (Rumohra), распространенный в Австралии и Новой Зеландии, на Новой Гвинее, в Китае, Южной Африке, Центральной и Южной Америке.
Также широко распространены роды олеандра и нефролепис - представители другого подсемейства - олеандровых. Кроме олеандры и нефролеписа, в его состав входят артроптерис (Arthropteris) и псаммиосорус (Psammiosorus) - два небольших рода, распространение которых ограничено Старым Светом. Папоротники этого подсемейства по морфологии корневищ (листья на радиальных корневищах тесно сближены), форме покрывальца (если оно присутствует, имеет обычно округло-почковидную форму), наличию перины у спор хорошо отличаются от собственно даваллиевых.
Наименее специализированным родом в подсемействе олеандровых считают нефролепис (около 30 видов) - род преимущественно тропического распространения. За пределами тропиков он встречается лишь в Японии и Новой Зеландии.
Нефролеписы - обитатели открытых мест. Нефролепис возвышенный (N. exaltata) - одно из первых высших растений, обычно поселяющихся после извержения (уже спустя 4-5 месяцев) на склонах вулканов. Но этот нефролепис не переносит затенения и по мере формирования древесного покрова исчезает.
Нефролеписы - крупные папоротники. Листья нефролеписа остроконечного (Nephrolepis acuminata) достигают длины 2,5 м, а нефролеписа диксониевидного (N. dicksonioides) - 3,5 м. Они однажды перистые, не сочлененные с корневищем, но перья всегда сочленены со стержнем (рис. 146). При старении листа перья желтеют и опадают, оставляя торчащими голые стержни. Корневище, как правило, короткое, вертикальное, на верхушке несет пучок листьев.

Рис. 146. Гетерофиллия у садовых форм нефролеписа возвышенного (Nephrolepis exaltata): листья формы 'Норвуд'

Рис. 146. Гетерофиллия у садовых форм нефролеписа возвышенного (Nephrolepis exaltata): листья формы 'Верона'
Сорусы у нефролеписов располагаются на концах жилок. Они либо округлые, либо вытянутые вдоль края, как у нефролеписа остроконечного. Покрывальце округлое или продолговатое, фиксированное в одной точке или прикрепленное вдоль основания (рис. 147). Спорангии на ножках, разновозрастные в пределах одного соруса. Споры билатеральные, монолетные, мелкие, значительно мельче, чем у папоротников подсемейства даваллиевых, с более или менее хорошо различимой периной. Гаметофит сердцевидной формы. Кроме обычного размножения при помощи спор, нефролеписы легко размножаются вегетативно. На их корневищах образуются наземные безлистные, покрытые чешуйками укореняющиеся побеги, подобные усам земляники. Это очень эффективное средство размножения. В течение одного года одно растение может образовывать свыше ста новых. Некоторые виды этого рода размножаются при помощи клубней, в изобилии образующихся на подземных побегах - столонах. Старые экземпляры нефролеписа сердцевиднолистного (N. cordifolia), выращиваемого в теплицах, могут производить свыше двухсот клубней в год. Самые крупные из них достигают длины 2-2,5 м. Молодые клубни белые или серебристые из-за многочисленных чешуек, покрывающих их поверхность. В тканях клубня содержится большое количество воды, Сахаров, белков и жиров. При отделении клубни могут прорастать немедленно без всякого периода покоя. Обычно из одного клубня вырастает одно растение. Оно всегда имеет нормальные листья, такие же, как и листья материнского растения. Выращенные из усов особи могут иметь листья, сильно отличающиеся от обычных однажды перистых листьев нефролеписа. В начале XX в. в Бостоне (США) была выведена форма нефролеписа возвышенного с многократно тонко рассеченными листьями, получившие название "бостонский (или кружевной) папоротник". В настоящее время получено большое количество причудливых садовых форм нефролеписа возвышенного, многие из которых легко переносят затенение и сухость воздуха, и поэтому их с успехом выращивают в обычных городских квартирах.
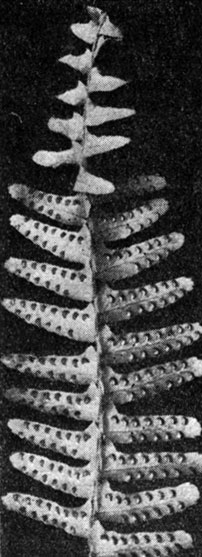
Рис. 147. Нефролепис сердцевиднолистный (Nephrolepis cordifolia) - нижняя сторона листа с сорусами
Очень похожи на нефролепис папоротники рода артроптерис. Их листья сочленены с корневищем, а листочки, как и у нефролеписа, сочленяются с рахисом. Виды артроптериса - эпифиты с длинными лазящими корневищами, покрытыми пельтатными чешуями. Сорусы поверхностные, на концах жилок, без покрывальца у артроптериса нежного (Arthropteris tenella) или с почковидно-округлым покрывальцем. В роде 20 видов, населяющих, в отличие от нефролеписа, исключительно районы Старого Света (тропическая Африка, Азия, Австралия, Новая Зеландия).
Несколько обособленно в подсемействе стоит род олеандра. В роде всего 40 видов, из них три - в Америке, столько же в Африке, остальные в Юго-Восточной Азии и Полинезии. Это типичные эпифиты с длинными корневищами. Листья причленяются к филлоподиям, которые иногда достигают значительной длины. Олеандра обладает необычным для папоротников характером роста. На корневище очень скученно располагаются листья, как у нефролеписа, но корневище продолжает расти и на некотором расстоянии формирует пучок новых листьев и т. д. Часть корневища в промежутках между соседними пучками листьев обвивается вокруг ствола дерева. Листья олеандры простые, цельные, гладкие и блестящие или иногда опушенные, очень жесткие, иногда с хрящеватым краем. Форма и консистенция листа этих папоротников напоминают всем хорошо известный олеандр (Nerium oleander), в честь которого этот род и получил свое название. Листья одинаковые или реже вегетативные и спороносные отличаются по форме. Своеобразно устроена корневая система у папоротников этого рода. Корни у олеандры образуются не на корневищах, а на отходящих от них корнеподобных образованиях - ризофорах.
Строение покрытых пельтатными чешуями корневищ, особенности спор даваллиевых указывают на то, что это семейство является филогенетически подвинутым. Возможных предков этой группы папоротников, по мнению многих специалистов, следует искать среди предков папоротников из подсемейства аспидиевых (семейство асплениевые).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'