Классификация и филогения папоротниковидных
За последние годы систематика папоротников достигла больших успехов. Это объясняется прежде всего значительным прогрессом наших знаний в области сравнительной морфологии как ныне живущих, так и вымерших папоротников (особенно древних, палеозойских групп). Сделано очень много, с частности, в изучении устьиц, спор и гаметофитов. Накоплен богатый материал по цитологии папоротников. Все это позволило пересмотреть старые схемы классификации папоротников и подойти значительно ближе к выяснению родственных взаимоотношений между ныне живущими группами папоротников и направлением их эволюции. Синтез всех этих данных позволяет строить классификацию и филогению папоротников на гораздо более надежном фундаменте, чем это было, например, в 1935 г., когда известный английский птеридолог Ф. Бауэр предложил новый вариант своей филогенетической классификации. И хотя среди птеридологов все еще имеются большие разногласия, в частности об объеме высших таксономических единиц папоротников, общая картина эволюции значительно прояснилась. На современном уровне наших знаний отдел папоротниковидных можно подразделить на следующие 7 классов.
Класс 1 - аневрофитопсиды (Aneurophytopsida). Этo очень древняя группа, геологическая история которой известна с конца раннего до середины позднего девона (400-375 млн. лет назад). Наиболее примитивные представители этого класса (особенно среднедевонский род птилофитон - Ptilophyton, известный также под названием Protopteridium) имеют еще много общего с риниофитами и представляют собой, в сущности, промежуточные формы между ними и папоротниками. Предполагается, что они произошли от представителей порядка псилофитовых (Psilophytales). Растения, относящиеся к этому классу, еще не имели настоящих листьев. Лишь у птилофитона конечные веточки были уже уплощенные, листоподобные. Это были своего рода "плосковетки" - первая стадия в возникновении настоящих, плоских, дорсивентральных листьев. Но подобно листьям настоящих папоротников конечные веточки аневрофитопсид были в молодости улиткообразно закручены (рис. 93). Внутреннее строение ветвей всех порядков еще очень примитивное, всегда протостелическое. Первичная ксилема мезархная. Почти у всех представителей класса происходило образование вторичной ксилемы, а у некоторых из них - довольно сложно устроенной вторичной флоэмы. Трахеиды вторичной ксилемы были точечные, т. е. с округлыми окаймленными порами. Спорангии были относительно крупные, верхушечные, одиночные или собранные в пучки. Споры одинаковые, с трехлучевым тетрадным рубцом.
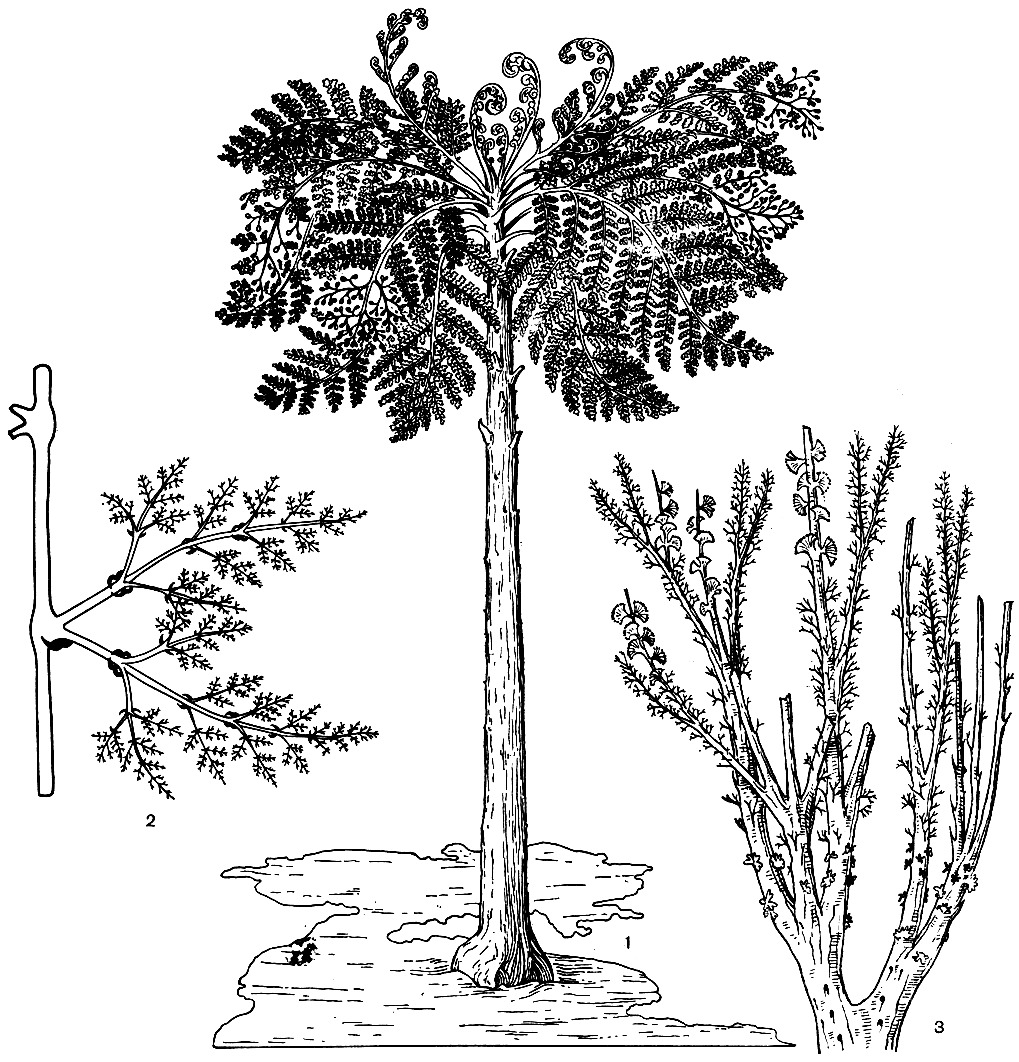
Рис. 93. Вымершие папоротники: 1 - эосперматоптерис (Eospermatopteris), общий вид, реконструкция; 2 - ставроптерис (Stauropteris), часть растения с перисторазветвленными конечными ветвями; 3 - кладоксилон (Cladoxylon), часть растения с пучками спорангиев и разветвленными листоподобными органами
Известный американский ботаник Д. Бирхорст (1971) включает в свой класс аневрофитопсид (название класса было предложено им) также представителей следующего далее класса археоптеридопсид. Однако в настоящее время представляется более правильной точка зрения русского палеоботаника Н. С. Снигиревской, которая предлагает разделить аневрофитопсиды Бирхорста на два самостоятельных класса - аневрофитопсиды в более узком смысле и археоптеридопсиды. В класс аневрофитопсид входит порядок аневрофитовых (Aneurophytales), а также ряд растений, родство которых еще не вполне точно установлено.
Класс 2 - археоптеридопсиды (Archaeopteridopsida). Представители этого класса известны из отложений конца среднего девона - начала карбона (390-360 млн. лет назад). От аневрофитопсид, с которыми их обычно объединяют, они хорошо отличаются наличием настоящих листьев с цельной, вильчато, пальчато или перисторассеченной пластинкой. Но вот что очень интересно и в эволюционном отношении очень важно: эти, казалось бы, столь сходные с листьями папоротников настоящие листья, в молодости не бывали улиткообразно закрученными. Этим они напоминают листья современных офиоглоссопсид (ужовниковых). Строение ветвей сифоностелическое, а не протостелическое, как у аневрофитопсид. Первичная ксилема состояла из кольца мезархных пучков, сливавшихся при образовании вторичной ксилемы в цилиндрический тяж. Взрослые растения имели древовидный облик, отдаленно напоминая современные хвойные (рис. 94). Вторичная ксилема достигала большой мощности, т. е. древесина была пикноксильного типа. Трахеиды вторичной ксилемы были точечные, чем археоптеридопсиды отличаются от всех остальных папоротниковидных, за исключением аневрофитопсид и офиоглоссопсид. Спорангии крупные, расположенные одно- или двурядно на отдельных перисторассеченных спорофиллах, чередующихся со стерильными листьями. Таким образом, по расположению спорангиев археоптеридопсиды резко отличаются от аневрофитопсид, что еще более подтверждает необходимость рассматривать их как два самостоятельных класса. Кроме того, у многих представителей археоптеридопсид известна гетероспория (разноспоровость).
В классе один порядок - археоптерисовые (Аrchaeopteridales). Основываясь на наличии ряда общих черт с голосеменными (особенно на наличии вторичной древесины, которая у археоптериса вполне голосеменного типа, а также на разноспоровости ряда представителей), американский палеоботаник Ч. Бек (1960) назвал этот класс (понимаемый им шире, чем здесь) "прогимноспермами" (Progymnospermopsida). Однако Д. Бирхорст (1971) совершенно правильно указывал на то, что название таксона не должно отражать филогенетическую интерпретацию, и поэтому предложил более правильное с точки зрения ботанической номенклатуры название - аневрофитопсиды.
Класс 3 - кладоксилопсиды (Cladoxylopsida). Как и класс аневрофитопсид, восходит к концу раннего девона (позднеэмсское время), но некоторые его представители дожили до раннего карбона. В отличие от аневрофитопсид и археоптеридопсид представители этого класса, по-видимому, не имели камбия и, следовательно, не образовывали вторичной древесины. Хотя представители девонского рода псевдоспорохнус (Pseudosporochnus) достигали, вероятно, в высоту 2 м, среди кладоксилопсид не было настоящих древесных форм, если не считать один вид кладоксилона (Cladoxylon nodosus), который имел, по-видимому, облик маленького деревца. Листья были дихотомически разветвленные. Проводящая система представляла собой довольно своеобразную рассеченную актиностелу, многочисленные меристелы которой составляли вдоль по длине стебля анастомозирующую систему. Ксилема состояла из лестничных и точечных трахеид. Спорангии были верхушечные и лишены каких-либо специальных приспособлений для раскрывания. Кладоксилопсиды были равноспоровыми растениями. Они представляли собой, вероятно, слепую ветвь эволюции.
Класс 4 - зигоптеридопсиды (Zygopteridopsida, или Goenopteridopsida). Более поздняя и относительно более подвинутая группа, чем три предыдущих класса. Геологическая история зигоптеридопсид начинается в позднем девоне и заканчивается в перми, но временем их расцвета был карбон. Как и представители трех предыдущих классов, они еще очень сильно отличались по своему облику от типичных папоротников, хотя некоторыми своими чертами уже несколько больше приближались к ним. Хотя среди них и были древовидные формы, но в большинстве случаев зигоптеридопсиды - небольшие растения. Проводящая система их была обычно протостелической, но в карбоне появились также формы с сифоностелой (анахороптерис - Anachoropteris, апотроптерис - Apotropteris и ботриоптерис - Botryopteris). Камбиальное утолщение стебля отсутствовало или было слабо выражено (зигоптерис - Zygopteris). Трахеиды варьировали от типичных лестничных до точечных (с округлыми окаймленными порами). Ветвление в большинстве случаев было дихотомическое, причем одна ветвь дихотомии становилась "листом", а другая продолжала расти в качестве стебля. У наиболее примитивных представителей не было еще ясно выраженного отличия листа от стебля. Их листоподобные органы представляли собой ограниченные в росте, сильно разветвленные и иногда улиткообразно закрученные в молодости боковые ветви, мало похожие на плоские дорсивентральные листья позднейших, более типичных папоротников.
Среди представителей этого класса наблюдаются все переходы от радиально-симметричных листовых органов к более типичным дорсивентральным листьям, которые уже начинают преобладать в позднем карбоне и в перми. Процесс уплощения листоподобных органов начинался с их конечных долек, и у радиально-симметричных листьев их конечные сегменты были уже иногда плоскими. Спорангии были верхушечные, довольно крупные и раскрывались верхушечной порой или чаще продольной трещиной. Продольное раскрывание происходило посредством примитивного кольца, представлявшего обычно широкую полоску из толстостенных клеток. Кольцо было приурочено к одной только стороне спорангия или протягивалось по обе его стороны. Спорангии сидели одиночно либо более или менее срастались ножками в пучки; своего рода примитивные верхушечные сорусы, которые у некоторых форм, в свою очередь, превращались в синангии. Растения были обычно равноспоровые, реже разноспоровые (ставроптерис - Stauropteris, рис. 93). Зигоптеридопсиды, как и три предыдущих класса, произошли от порядка псилофитовых (отдел риниофиты). По мнению многих исследователей, от зигоптеридопсид берут начало многие типичные папоротники (в частности, осмундовые, схизейные и глейхениевые). В этот класс входит порядок зигоптерисовые (Zygopteridales), содержащий несколько семейств (в том числе зигоптерисовые - Zygopteridaceae, ботриоптерисовые - Botryopteridaceae и ставроптерисовые - Stauropteridaceae). Вполне возможно, что зигоптеридопсиды представляют собой не вполне естественную группу.
Класс 5 - офиоглоссопсиды, или ужовниковые (Ophioglossopsida). Современные папоротники. Происходят они, вероятно, непосредственно от равноспоровых форм палеозойских аневрофитопсид. Небольшие или средних размеров многолетние растения, обычно наземные, редко эпифиты (некоторые тропические виды ужовника - Ophioglossum). Спорофиты более или менее мясистые, лишенные механических тканей. Корневище чаще короткое, простое. В редких случаях наблюдается дихотомическое ветвление. Чешуи на корневищах отсутствуют. Проводящая система корневища представляет собой эктофлойную сифоностелу или диктиостелу, но у некоторых представителей (гроздовник - Botrychium, гельминтостахис - Helminthostachys, и ряд видов ужовника) молодое корневище имеет протостелическое строение. Ксилема гельминтостахиса экзархная, но у остальных родов она эндархная, что несколько необычно для папоротников.
У более крупных гроздовников имеется активно функционирующий камбий, причем у гроздовника вирджинского (Botrychium virginianum) вторичная ксилема хорошо развита и имеет лучи. Эта особенность отличает офиоглоссопсиды почти от всех остальных современных папоротников и сближает их с палеозойскими аневрофитопсидами. Более того, у гельминтостахиса и гроздовника окаймленные поры трахеид округлые или овальные и с явственным торусом, как у многих голосеменных. Листья отличаются от листьев других современных папоротников отсутствием улиткообразного закручивания в молодости, наличием особых влагалищных прилистникоподобных образований при основании, а также обычно мягкостью. Подобно корневищам они обнаруживают иногда признаки дихотомического ветвления. Характерна диморфность листьев.
Растения равноспоровые. Спорангии довольно крупные, содержат большое число спор (от 1500 до 15 000), имеют массивные многослойные стенки и снабжены устьицами. Они лишены кольца и раскрываются морфологически верхушечной щелью. Споры трилетные, вначале тетраэдрические, но затем становятся почти шаровидными. У некоторых видов в спорах наблюдаются бледные хлоропласты. Гаметофиты крупные, мясистые, микоризные, дихотомически или неправильно ветвистые. В классе один порядок - ужовниковые (Ophioglossales), содержащий одно семейство Ophioglossaceae с тремя подсемействами - Botrychioideae, Helminthostachydoideae и Ophioglossoideae.
Класс 6 - мараттиопсиды (Marattiopsida). Современные папоротники, геологическая история которых восходит к карбону (пермо-карбоновый род псарониус - Psaronius - и др.). Происходят, вероятно, непосредственно от зигоптеридопсид. Многолетние растения, варьирующие от мелких форм до очень крупных. Стебли представляют собой дорсивентральные корневища или толстые клубневидные стволы. Стебли отличаются мясистостью. В стеблях, как и в других вегетативных органах, имеются большие лизигенные слизевые ходы, являющиеся одной из особенностей мараттиопсид. Молодой стебель протостелический, но на уровне третьего или четвертого листа протостела сменяется амфифлойной сифоностелой, которая в корневищах сохраняется до конца. У крупных форм образуется диктиостела очень сложного строения (наиболее сложная у рода ангиоптерис - Angiopteris). Трахеиды лестничные. У рода ангиоптерис наблюдается очень слабое развитие вторичной ксилемы. Для корней мараттиопсид, в отличие от остальных папоротников, характерно полиархное актиностелическое строение проводящей системы. Корни несут своеобразные многоклеточные корневые волоски. Первые образующиеся корни обычно содержат в коре микоризный фикомицетный гриб. Листья большинства мараттиопсид перистые (обычно сложноперистые), но у рода кристенсения (Christensenia) - пальчатые, а у одного вида - данеи простолистной (Danaea simplicifolia) - цельные. Молодые листья всегда спирально закрученные. Очень характерно наличие у основания листьев двух толстых прилистниковидных образований, соединенных вместе особой поперечной перемычкой.
Равноспоровые растения. Спорангии довольно крупные, толстостенные и снабжены устьицами. Расположенные на нижней (абаксиальной) стороне зеленых листьев вдоль жилок спорангии образуют сорусы или срослись в различного рода синангии. Покрывальце отсутствует. Зрелые спорангии раскрываются посредством щелей или пор, причем у некоторых родов (ангиоптерис, макроглоссум - Macroglossum, архангиоптерис - Archangiopteris) имеются примитивные кольца, напоминающие по своему строению кольца некоторых зигоптеридопсид. Споры трилетные или монолетные. Число их в каждом спорангии очень велико (от 1450 до 7500).. Гаметофиты надземные, темно-зеленые, мясистые, дорсивентральные, сердцевидные или продолговатые, долговечные. Антеридии большие и погруженные, как у офиоглоссопсид, и развиваются сходным образом. Архегонии сходны с архегониями офиоглоссопсид, но шейки их более короткие и широкие. В класс входит один порядок - мараттиевые (Marattiales), который состоит из одного современного семейства Marattiaceae, иногда разделяемого на три семейства - Marattiaceae, Christenseniaceae и Danaeaceae. В двух последних семействах по одному роду. Другие авторы признают только два семейства, а некоторые, наоборот, разделяют этот порядок на четыре семейства.
Класс 7 - полиподиопсиды (Polypodiopsida). Современные папоротники, геологическая история которых восходит к карбону. Происходят, вероятно, непосредственно от зигоптеридопсид. Многолетние или очень редко однолетние растения, варьирующие от мелких форм до очень крупных. Стебли или представляют собой прямостоячие стволы, у древовидных форм достигающие иногда значительной высоты, или превращены в корневища. Проводящая система от протостелы до очень специализированных типов диктиостелы. Трахеиды лестничные. Редко имеются сосуды. Листья очень различных типов как по размерам, так и по форме, жилкованию и плотности. Молодые листья спирально закрученные. Равноспоровые или редко разноспоровые растения. Спорангии в большинстве случаев собраны в сорусы. Покрывальце может быть или отсутствовать. Спорангии имеют однослойные стенки и лишены устьиц. Имеется кольцо, которое в пределах класса отличается большим разнообразием и достигает высокой специализации. По мере специализации кольца число спор в каждом спорангии постепенно уменьшается. Споры трилетные или монолетные. Гаметофиты у большинства представителей класса зеленые, наземные и обоеполые, в некоторых случаях - со слабо выраженной тенденцией к разделению полов, но у разноспоровых форм совершенно раздельнополые и сильно редуцированные. Антеридии довольно сильно отличаются от антеридиев офиоглоссопсид и мараттиопсид: они более или менее выступающие над поверхностью гаметофита, более мелкие и производят значительно меньшее число сперматозоидов (у высших форм до 32 в каждом антеридии). Архегонии сравнительно мало отличаются от архегониев офиоглоссопсид и мараттиопсид.
Полиподиопсиды подразделяются на 3 подкласса: Polypodiidae (порядки Osmundales, Schizaeales, Polypodiales и Cyatheales), Marsileidae (порядок Marsileales) и Salviniidae (порядок Salviniales).
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'