Развитие молодого спорофита и формирование коробочки
Оплодотворенная яйцеклетка одевается оболочкой и превращается в зиготу, остающуюся в брюшке архегония. Зигота делится поперечной перегородкой на верхнюю и нижнюю клетки. Нижняя клетка делится и образует нижнюю часть ножки и стопу, которая прорастает сквозь стенку брюшка архегония и внедряется в ткань гаметофита, но не срастается с ней.
Верхняя (апикальная) клетка делится двумя взаимно пересекающимися перегородками, наклоненными друг к другу. Благодаря этому вычленяется двусторонняя верхушечная клетка, отделяющая два ряда сегментов, из которых формируются коробочка и верхняя часть ножки. Рост молодого спорофита на верхнем конце происходит более активно, и деление осуществляется более правильно. В результате развивается стеблеобразное тело с последующей дифференциацией на коробочку и ножку со стопой. Удлинение спорофита приводит к поперечному разрыву окружающего его разросшегося архегония. Верхняя часть архегония приподнимается коробочкой в виде колпачка различной формы, а нижняя часть (влагальце) воротничкообразно окружает основание ножки. Как же формируется коробочка?
Молодой спорофит, как мы отмечали, растет путем отчленения двусторонней верхушечной клеткой двух рядов сегментов. На поперечном срезе в верхней части зародыша наблюдается строгая последовательность деления сегментов, которая не прослеживается при формировании ножки. Вначале сегменты делятся радиально на 4 клетки, образуя квадрант, затем каждая из клеток делится еще раз косой (антиклинальной) перегородкой. Возникают 4 трехсторонние и 4 четырехсторонние клетки. В последних поперечными (периклинальными) стенками обособляются 4 внутренние клетки, образующие эндотеций. Так происходит отделение внешнего слоя из 8 наружных клеток, называемого амфитецием, от внутренних клеток эндотеция.
Дальнейшее развитие амфитеция и эндотеция осуществляется независимо друг от друга, и они дают начало разным тканям коробочки.
На основании того, какие ткани в дальнейшем формируются из амфитеция и эндотеция, различают три основных типа развития спорофита и соответственно три разных типа археспория у сфагновых, андреевых и бриевых мхов.
Клетки наружной и внутренней стенок спорового мешка, а также клетки колонки богаты питательными веществами и доставляют их спорообразующим клеткам.
Материнские клетки спор возникают в результате последовательного деления первичных клеток археспориальной ткани. Число этих делений от 4 до 32. Образование спор осуществляется путем мейоза, в ходе которого внутри оболочки материнской клетки образуется тетрада спор. Как только каждая спора оденется оболочкой, оболочка материнской клетки разрушается и споры освобождаются.
Колпачок мхов чаще всего является пленчатым образованием, частично или целиком покрывающим молодую, а также почти зрелую коробочку, предохраняя ее от внешних воздействий. Колпачок обычно имеет плотную консистенцию и наверху большей частью многослойный. Его суженная буроватая верхушка часто соответствует шейке архегония. Обертка зародыша - эпигон (от греч. epigonos - родившийся после) - первоначально цельная, формируется не только за счет собственна архегония, но и его ножки, а отчасти и стебля.
У большинства листостебельных мхов колпачок достаточно хорошо развит, но у сфагновых и андреевых мхов и у некоторых клейстокарпных эфемеров из бриевых он развит слабо.
Колпачки могут быть очень разнообразны (рис. 49).
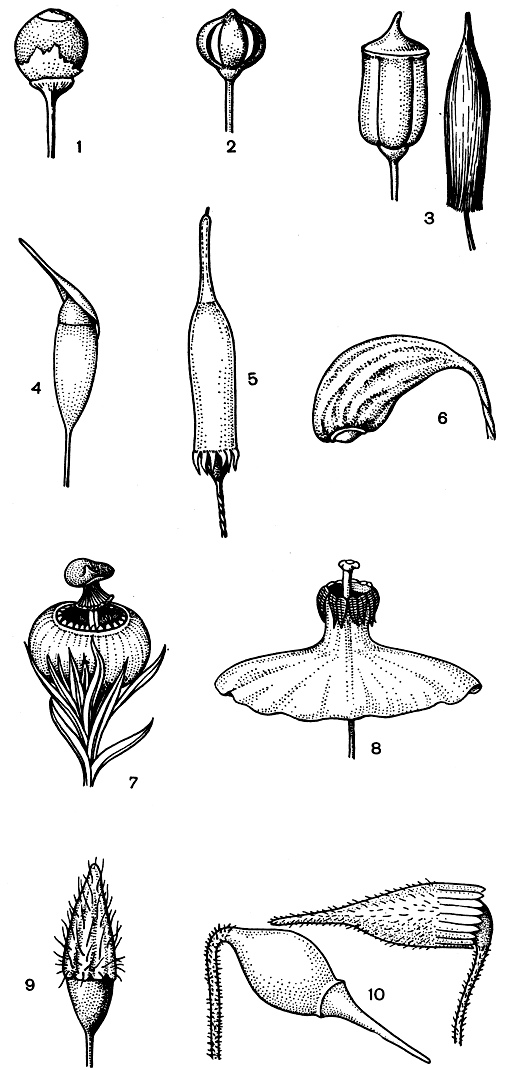
Рис. 49. Различные формы коробочек и колпачков у листостебельных мхов: 1 - сфагнум; 2 - андрея; 3 - политрихум; 4 - гомалия; 5 - энкалипта; 6 - фунария; 7 - скоулерия; 8 - сплахнум; 9 - ортотрихум; 10 - пелекиум
Колпачок опадает еще до полного развития коробочки, реже он долго остается на коробочке и тогда опадает вместе с крышечкой.
У большинства бриевых ножка хорошо развита, хотя длина ее даже у представителей одного семейства может сильно варьировать. Среди бриевых мхов есть роды, у которых ножка отсутствует или не достигает полного развития. Это многие эфемеры с клейстокарпными, погруженными в перихециальные листья коробочками (рис. 50).
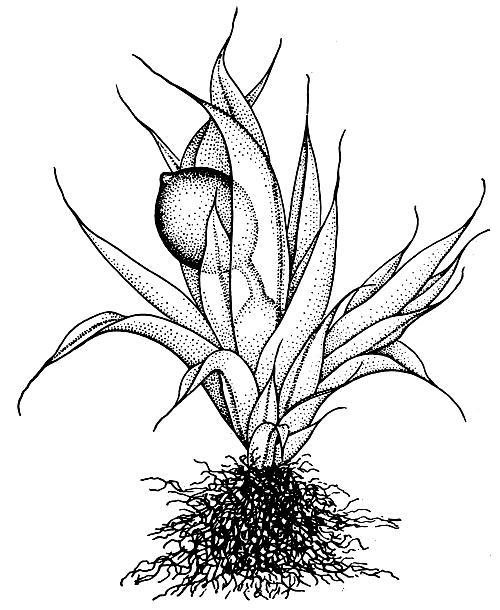
Рис. 50. Фаскум (Phascum piliferum) - общий вид спороносящего растения
У сфагновых и андреевых мхов ножка представлена только коротким шейковидным связующим участком между крупной луковицеобразной стопой и коробочкой и носит название шейки. Функцию ножки у сфагновых мхов и андреи выполняет удлиненная безлистная верхняя часть архегониальной ветви - ложноножка. Вначале короткая, ложноножка после созревания спор быстро удлиняется и поднимает спорогон над перихециальными листьями.
У многих видов в ножке спорогона имеются те же ткани, что и в стебле гаметофита.
У большинства мхов ножка очень чутко реагирует на колебания влажности воздуха. При подсыхании она спирально закручивается, а с ней вместе толчками вращается коробочка, что содействует более активному рассеиванию спор. У тех мхов, ножки которых способны спирально закручиваться (фунария, цинклидиум и др.), наружные стереидные клетки расположены по спирали. А, например, у политрихума обыкновенного (Polytrichum commune) наружные стереиды расположены параллельно оси ножки, поэтому скручивания ножки у него при потере воды не происходит. Впрочем, основной причиной гигроскопического движения ножки является способность наружных стенок стереид необычайно быстро впитывать влагу, разбухать и раскручивать ножку, а при подсыхании соответственно легко терять воду и скручивать ножку.
Стопа - нижняя часть ножки - довольно глубоко внедряется в ткань гаметофита и таким образом прикрепляет спорогон. Но главная функция стопы - всасывание из гаметофита веществ, необходимых для развития спорофита.
В основании стопы поверхностные клетки сосочкообразно вытянуты, а иногда даже дают разветвленные многоклеточные ризоидообразные выросты. Центральный пучок ножки может продолжаться в стопу и нередко проникает в центральный пучок стебля.
Влагальце служит передатчиком питательных веществ от гаметофита к растущему спорофиту через его стопу. Следует отметить, что между гаметофитом и спорофитом нет прочной связи и спорофит относительно легко может быть отделен от гаметофита.
Коробочка мхов - специализированный орган бесполого размножения, и все особенности ее строения служат выполнению этой функции. Форма коробочек очень разнообразна (рис. 49, табл. 7). Окраска коробочек изменяется от желтой до бурой, но может быть и других оттенков. Поверхность коробочек бывает различной (рис. 49).
В коробочке различают крышечку, колечко, урночку, шейку и апофизу, если последняя развита (рис. 51). У некоторых видов - коробочки с неотделяющейся крышечкой. Такие мхи называют клейстокарпными (закрытоплодными). Мхи с ясно отделяющейся крышечкой называют стегокарпными (крышкоплодными). Крышечка после созревания спор отпадает. В сбрасывании крышечки принимают участие колечко, а также зубцы перистома. Кроме того, ко времени созревания коробочки в кольцевой зоне возникает значительное натяжение тканей, способствующее сбрасыванию крышечки.
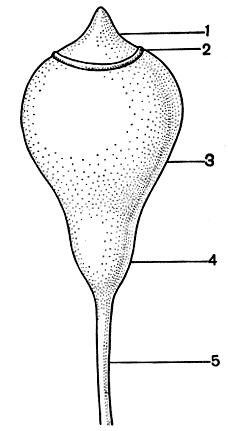
Рис. 51. Строение коробочки бриевых (схема): 1 - крышечка; 2 - колечко; 3 - урночка; 4 - шейка; 5 - ножка
Колечко расположено между краем урночки и крышечкой и состоит из одного или нескольких рядов лежащих друг над другом уплощенных бесцветных, сильно гигроскопичных клеток. Когда эти клетки набухают от воды, колечко расширяется и отделяется как от урночки, так и от крышечки.
Урночкой называют ту часть коробочки, в которой развиваются споры. В верхней части урночки есть широкое или суженное отверстие - устье, закрытое крышечкой. По краю устья часто развиты выросты в виде зубцов разнообразной формы, которые называют перистомом или околоустьем.
Стенки коробочки состоят из двух или нескольких слоев. Наружный слой - экзотеций, являющийся гомологом эпидермы других высших растений, - кутинизирован. Клетки экзотеция бедны плазмой, их наружные оболочки обычно утолщены и плотно прилегают друг к другу. Внутренние слои стенки коробочки крупноклеточные, паренхиматические с тонкими бесцветными клеточными оболочками, составляют так называемую водоносную ткань.
У молодого спорогона клетки стенок богаты хлоропластами, но по мере его созревания число хлоропластов уменьшается и ассимиляционная функция экзотеция сменяется механической. В экзотеции нижней половины урночки и шейки находятся устьица. Различают устьица поверхностные, расположенные на уровне экзотеция, и погруженные, расположенные ниже его уровня. Устьица обычной формы имеют щель и состоят из двух симметричных почковидных замыкающих клеток. Устьица без щели присущи сфагновым. У андреевых, а также у бриевых - архидиума (Archidium), схистостеги (Schistostega), тетрафиса (Tetraphis) - устьица отсутствуют совсем.
Под экзотецием расположена одно- или дву- четырехслойная водоносная ткань. Клетки ее крупнее клеток экзотеция, тонкостенные и заполнены водой. Водоносная ткань снабжает водой расположенную глубже ассимиляционную ткань, а благодаря тонкостенности и прозрачности легко пропускает световые лучи к хлоропластам.
У большинства мхов стенки коробочки и споровый мешок разделены воздушной полостью, пронизанной нитями из хлорофиллоносных клеток. У политриховых коробочка чаще с двумя воздушными полостями, наружной и внутренней, последняя между колонкой и споровым мешком. Нет воздушной полости и ассимиляционной ткани у андреевых и у тех родов, у которых споровый мешок непосредственно примыкает к стенке коробочки, например у тетрафиса, леукодона (Leucodon). Нет ее и у водных мхов.
Колонка обычно проходит в коробочке от шейки до крышечки. У очень немногих видов она вытягивается и выступает наружу из урночки и поднимает оставшуюся прикрепленной к ней крышечку (рис. 49, 52). После отпадения крышечки у многих мхов колонка сморщивается и остается на дне урночки.
Перистом состоит из одно- или двурядных зубцов, из ресничек или нитей разнообразной формы, характерных для таксонов разных рангов (рис. 52). У некоторых мхов перистом совершенно отсутствует; у других бывает недоразвитый, рудиментарный.
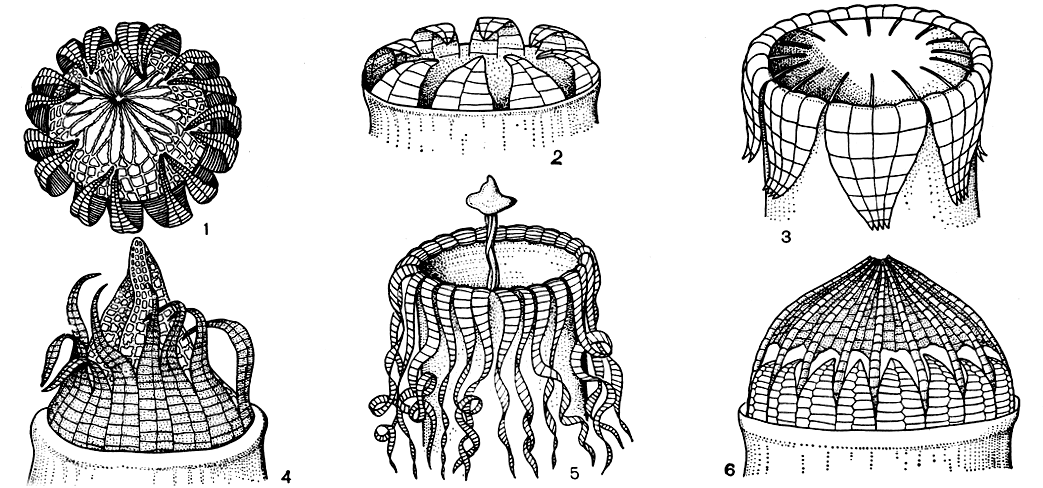
Рис. 52. Тины перистомов у бриевых мхов: 1 - тиммия (Timmia); 2 - октоблефарум (Octoblepharum); 3 - улота (Ulota); 4 - фонтиналис (Fontinalis) 5 - тейлория (Тауloria); 6 - цинклидиум (Cinclidium)
Зубцы простого (однорядного) перистома и экзостома (наружного ряда двойного перистома) обладают способностью к гигроскопическим движениям, поскольку состоят из двух пластиночек разной анатомической структуры и соответственно имеют различную степень гигроскопичности. Гигроскопические движения зубцов активно содействуют постепенному высеиванию спор из урночки и их распространению, а также обусловливают замыкание устья урночки во влажной атмосфере, предохраняя споры от намокания и прорастания внутри коробочки.
Нижняя часть коробочки, не содержащая спор,- шейка (рис. 51) - постепенно или внезапно переходит в ножку. Нередко на шейке наблюдается одностороннее разрастание ткани, так называемый зобик. Зобики характерны для коробочек некоторых мхов из дикрановых (Dicranaceae).
Чрезвычайно интересным приспособительным образованием спорогона является апофиза (от греч. аро - из и physa - вздутие), представляющая собой вздутие нижней части коробочки весьма разнообразной формы. Особенно типичное развитие апофизы у видов семейства сплахновых (Splachnaceae). У родов тетраплодон (Tetraplodon) и сплахнум (Splachnum) она бывает значительно шире коробочки и более ярко окрашенной в желтый и различные тона красного цвета (табл. 7, рис. 49). Яркая окраска апофизы, а также выделяемая через устьица жидкость с неприятным запахом служат средством привлечения мух, которые являются распространителями клейких спор этих мхов.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'