Классификация и филогения цветковых растений (А. Л. Тахтаджян)
Первые попытки классификации цветковых растений, как и растительного мира вообще, были основаны на немногих, произвольно взятых, легко бросающихся в глаза внешних признаках. Это были чисто искусственные классификации, в которых в одной группе оказывались вместе часто очень далекие в систематическом отношении растения. Начиная с первой искусственной системы итальянского ботаника Андреа Чезальпино (1583), было предложено несколько искусственных классификаций растительного мира, включая цветковые растения. Венцом периода искусственных классификаций была знаменитая "половая система" великого шведского натуралиста Карла Линнея (1735). В основу классификации Линней положил число тычинок, способы их срастания, а также распределение однополых цветков. Он разделил все семенные растения (цветковые и голосеменные) на 23 класса, а к 24-му классу отнес водоросли, грибы, мхи и папоротники. Вследствие крайней искусственности классификации Линнея в один и тот же класс попали роды самых разных семейств и порядков, а, с другой стороны, роды, бесспорно, естественных семейств, например злаков, нередко оказывались в разных классах. Несмотря на эту искусственность, система Линнея была в практическом отношении очень удобна, так как давала возможность быстро определять род и вид растения, благодаря чему оказалась доступной не только специалистам, но и любителям ботаники. Вместе с тем Линней усовершенствовал и утвердил в ботанике и зоологии биноминальную (бинарную) номенклатуру, т. е. обозначение растений и животных двойным именем - по роду и виду. Это сделало систему Линнея еще более удобной для ее использования.
Поворотным пунктом в развитии систематики цветковых растений была книга французского натуралиста Мишеля Адансона "Семейства растений" (1763 - 1764). Он считал необходимым использовать для классификации растений максимально возможное число разных признаков, придавая всем признакам одинаковое значение. Но еще большее значение для систематики цветковых растений имела книга французского ботаника Антуана Лорана Жюсьё (1789), озаглавленная "Роды растений, расположенные согласно естественному порядку". Он разделил растения на 15 классов, в пределах которых различал 100 "естественных порядков". Жюсьё дал им описания и названия, большинство из которых сохранилось до настоящего времени в ранге семейств. Грибы, водоросли, мхи, папоротники, а также наядовые объединялись им под названием бессемядольных (Acotyledones). Семенные растения (без наядовых) он разделил на однодольные (Monocotyledones) и двудольные (Dicotyledones), отнеся к последним также и хвойные.
В XIX в. наибольшее значение имела система швейцарского ботаника Огюстена Пирама де Кандолля (1813, 1819). Он начал издавать обозрение всех известных видов цветковых растений, названное им "Продромус естественной системы царства растений" (от греч. prodromos - предтеча). Это важнейшее в истории систематики растений издание начало выходить в 1824 г. и было закончено его сыном Альфонсом в 1874 г. Многие ботаники продолжали разрабатывать систему де Кандолля, внося в нее более или менее существенные изменения. Логическим завершением всех этих исследований была система английских ботаников Джорджа Бентама и Джозефа Хукера, опубликованная ими в капитальном издании "Роды растений" (Genera plantarum) в 1862 - 1883 гг. Это был значительно улучшенный вариант системы де Кандолля. Хотя система Бентама и Хукера появилась после выхода в свет "Происхождения видов" Чарлза Дарвина и оба они поддерживали идеи Дарвина, сама система была основана на додарвиновских представлениях о виде.
Зачатки эволюционной, или филогенетической, систематики растений существовали еще до переворота, произведенного Дарвином в биологии. Но развитие филогенетической (генеалогической) систематики фактически началось лишь после выхода в свет "Происхождения видов". Дарвин утверждал, что "всякая истинная классификация есть генеалогическая". Он считал, что в зависимости от "размера изменений, пройденных разными группами" в процессе эволюции, они размещаются по разным родам, семействам, порядкам, классам и пр., а сама система представляет собой "генеалогическое распределение существ, как в родословном дереве". Позже в своей книге о происхождении человека (1874) Дарвин писал, что всякая система "должна быть, насколько это возможно, генеалогической по своей классификации, т. е. потомки одной и той же формы должны быть объединены в одну группу, в отличие от потомков любой другой формы; но если родительские формы родственны, также родственны будут и потомки, а две группы при объединении составят более обширную группу". Таким образом, он приравнял "родство" (термин, употреблявшийся в ином смысле авторами "естественных" систем) к эволюционным взаимоотношениям, а систематические группы - к ветвям и веточкам родословного дерева. Систему иерархических взаимоотношений таксонов разных категорий он считал результатом эволюции, что было принципиально новым подходом к систематике и ее задачам.
В XIX в. было много попыток построить систему цветковых растений со стороны ботаников, принимавших эволюционное учение. Особенно широкую известность получили системы, созданные рядом немецких ботаников, среди которых особенно широкую известность и признание получила система А. Энглера. Однако существенным недостатком всех этих систем было смешение двух понятий - простоты и примитивности. Не учитывалось то обстоятельство, что простое строение цветка, например строение цветка казуарины, дуба или ивы, может быть не первичным, а вторичным. Игнорировалось значение редукции и вторичного упрощения, что, как мы уже знаем, имело большое значение в эволюции цветка, особенно у анемофильных растений. Поэтому эти системы, в том числе систему Энглера, нельзя назвать филогенетическими.
Еще в 1875 г. известный немецкий ботаник и натурфилософ Александр Браун выдвинул некоторые фундаментальные идеи, предвосхитившие на несколько десятилетий основные принципы филогенетической систематики цветковых растений. Он пришел к выводу о примитивности цветков магнолиевых и родственных семейств и вторичности безлепестных и однополых цветков, которые его современники, а ранее и он сам, считали примитивными. Простоту этих цветков он считал вторичной, результатом упрощения. Брауну принадлежит афоризм: "В природе, как и в искусстве, простое может быть самым совершенным". Таким образом, Браун ясно понимал, что есть два типа простоты строения: простота первичная, какую мы видим у действительно древних, примитивных форм, и простота вторичная, достигнутая в результате упрощения, как в цветке казуарины. Однако Браун вскоре (в 1877 г.) скончался, не осуществив реформы системы цветковых растений на основе сформулированных им принципов. Аналогичные идеи были высказаны также немецким ботаником Карлом Вильгельмом Негели (1884) и французским палеоботаником Гастоном де Сапорта (1885). Но оба эти выдающихся ботаника не были систематиками и не задавались целью построения эволюционной системы цветковых растений. Честь реформы классификации цветковых растений на новых началах принадлежит американскому ботанику Чарлзу Бесси и немецкому ботанику Хансу Халлиру. Их первые работы по системе цветковых растений появились в 1893 г. (Бесси) и 1903 г. (Халлир), но наиболее полный обзор системы Халлира был опубликован в 1912 г., а Бесси - в 1915 г.
В первой половине XX в. появился целый ряд новых систем цветковых растений, построенных на принципах, сформулированных Бесси и Халлиром. Из числа этих систем следует упомянуть систему профессора Петроградского университета Христофора Гоби (1916) и систему английского ботаника Джона Хатчинсона (1926, 1934). Во второй половине XX в. появляются системы А. Л. Тахтаджяна (1966, 1970, 1978), американских ботаников Артура Кронквиста (1968) и Роберта Торна (1968, 1976), датского ботаника Рольфа Дальгрена (1975, 1977) и ряд других систем.
Современная классификация цветковых растений основана на синтезе данных самых различных дисциплин, в первую очередь данных сравнительной морфологии, в том числе морфологии и анатомии репродуктивных и вегетативных органов, эмбриологии, палинологии, органеллографии и цитологии. Наряду с использованием классических методов морфологии растений с каждым днем все шире применяется электронный микроскоп, как сканирующий, так и трансмиссионный, что позволяет взглянуть на ультраструктуру многих тканей и клеток, включая пыльцевые зерна. В результате возможности сравнительно-морфологических исследований бесконечно расширились, что, в свою очередь, обогатило систематику ценным фактическим материалом для построения эволюционной классификации. В частности, большое значение начинает приобретать сравнительное изучение клеточных органелл, например изучение ультраструктуры пластид в протопласте ситовидных элементов (работы X. Д. Бэнке). Возрастающее значение приобретают также методы современной биохимии, особенно химии белков и нуклеиновых кислот. Начинают широко применяться серологические методы. Наконец, расширяется также применение математических методов и особенно компьютерной техники.

Основные различия между двудольными и однодольными
Отдел цветковые растения, или магнолиофиты, подразделяется на два класса: магнолиопсиды, или двудольные (Magnoliopsida, или Dicotyledones), и лилиопсиды, или однодольные (Liliopsida, или Monocotyledones). Основные различия между ними показаны на таблице (с. 109).
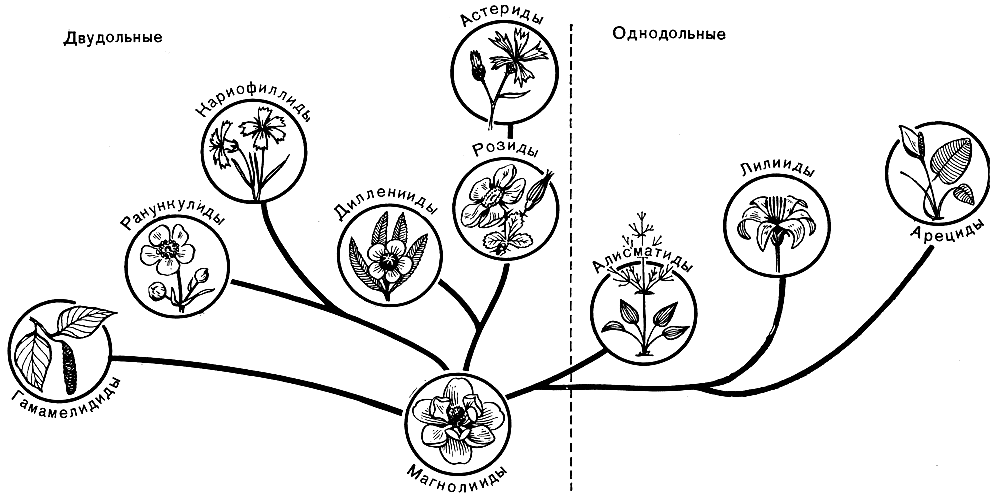
Рис. 50. Родственные связи подклассов двудольных и однодольных цветковых растений
Как хорошо видно из этой таблицы, нет ни одного признака, который служил бы резким различием между двумя классами цветковых растений. Эти классы различаются, в сущности, только комбинацией признаков. Они еще не настолько сильно разошлись в процессе эволюции, чтобы можно было разграничить их по одному какому-нибудь признаку. Тем не менее систематики, как правило, с легкостью устанавливают принадлежность того или иного растения к одному из этих классов. Затруднения вызывают только нимфейные и близкие семейства (объединяемые в порядок Nymphaeales), занимающие в ряде отношений промежуточное положение между двудольными и однодольными.
Однодольные произошли от двудольных и, вероятно, ответвились от них еще на заре эволюции цветковых растений. Наличие среди однодольных ряда семейств с апокарпным гинецеем и однобороздные пыльцевые зерна многих их представителей говорят о том, что однодольные могли произойти только от таких двудольных, которые характеризовались этими признаками. Среди современных двудольных наибольшим числом признаков, общих с признаками однодольных, обладают представители порядка нимфейных. Однако все представители этого порядка являются специализированными во многих отношениях водными растениями и поэтому не могут рассматриваться как вероятные предки однодольных. Но общее их происхождение весьма вероятно. Есть все основания для предположения, что однодольные и порядок нимфейных имеют общее происхождение от каких-то более примитивных наземных травянистых двудольных.
Ближайшие предки однодольных были скорее всего наземными растениями, приспособленными к постоянной или временной влажности. По мнению Дж. Бьюса (1927), ранние однодольные были болотными растениями или растениями лесной опушки. Дж. Л. Стеббинс (1974) высказывает предположение, что первые однодольные появились во влажной среде, по берегам рек и озер. Первичные однодольные были, вероятно, многолетними корневищными травами с цельными эллиптическими листьями, имеющими дуговидное жилкование, и разбросанными по поперечному разрезу стебля бессосудистыми проводящими пучками с остаточным внутрипучковым камбием. Цветки были в верхушечных соцветиях, 3-членные, с околоцветником в двух кругах, с андроцеем из примитивных лентовидных тычинок и с апокарпным гинецеем из примитивных кондупликатных плодолистиков. Пыльцевые зерна были однобороздные и в зрелом состоянии двухклеточные. Семена имели обильный эндосперм.
По числу видов, а также родов и семейств однодольные сильно уступают двудольным. Тем не менее роль однодольных в природе чрезвычайно велика, особенно в травянистых сообществах. Многие важнейшие культурные растения, в том числе хлебные злаки и сахарный тростник, относятся к однодольным.
Классы двудольных и однодольных, в свою очередь, подразделяются на подклассы, которые делятся на порядки (иногда объединяемые в надпорядки), семейства, роды и виды со всеми промежуточными категориями (рис. 50).
Класс двудольных, включающий около 325 семейств, около 10 000 родов и до 180 000 видов, подразделяется на 7 подклассов.
Подкласс 1. Магнолииды (Magnoliidae). Большей частью древесные растения, реже наземные или водные травы, иногда паразиты. Сосуды у некоторых форм отсутствуют. В паренхимных тканях часто имеются секреторные клетки. Устьица обычно с 2 побочными клетками. Цветки, как правило, обоеполые, часто спиральные или спироциклические. Зрелая пыльца 2-клеточная или реже 3-клеточная. Оболочка пыльцевых зерен однобороздная, 3 - 6-бороздная, ругатная, поровая или безапертурная. Гинецей в большинстве случаев апокарпный. Семязачатки чаще всего битегмальные (с двойным интегументом), крассинуцеллятные или, редко, тенуинуцеллятные. Эндосперм обычно целлюлярный, реже нуклеарный, редко гелобиальный. Семена большей частью с маленьким или очень маленьким зародышем и обильным эндоспермом, иногда также с периспермом.
В подкласс магнолиид входят наиболее примитивные порядки двудольных, в том числе магнолиевые, бадьяновые, лавровые и нимфейные. Хотя среди представителей подкласса нет ни одной ныне живущей формы, сочетающей в себе все примитивные признаки, магнолииды в целом представляют собой группу, которая стоит наиболее близко к гипотетической исходной группе, давшей начало ныне живущим цветковым растениям.
Подкласс 2. Ранункулиды (Ranunculidae). Близок к подклассу магнолиид, но более подвинут. Большей частью травы. Все представители обладают сосудами. Секреторные клетки в паренхимных тканях обычно отсутствуют (за исключением луносемянниковых). Устьица разных типов, в большинстве случаев без побочных клеток. Цветки обоеполые или однополые, часто спиральные или спироциклические. Зрелая пыльца большей частью 2-клеточная. Оболочка пыльцевых зерен трехбороздная или производная от трехбороздного типа, но никогда не бывает однобороздной. Семязачатки обычно битегмальные и крассинуцеллятные или реже тенуинуцеллятные. Семена чаще всего с маленьким зародышем и большей частью с обильным эндоспермом, редко без эндосперма.
В подкласс ранункулид входит порядок лютиковых и близкие к нему порядки. По всей вероятности, ранункулиды происходят непосредственно от магнолиид, скорее всего от предков типа бадьяновых.
Подкласс 3. Гамамелидиды (Hamamelididae). Большей частью древесные растения с сосудами (за исключением порядка троходендровых). Устьица с 2 или большим числом побочных клеток или побочные клетки отсутствуют. Цветки в большинстве случаев анемофильные, более или менее редуцированные, большей частью однополые; околоцветник обычно слабо развит, и цветки, как правило, безлепестные и часто также без чашечки. Зрелая пыльца большей частью 2-клеточная, трехбороздная или производная от трехбороздного типа. Гинецей обычно ценокарпный. Семязачатки часто битегмальные и в большинстве случаев крассинуцеллятные. Плоды большей частью односемянные. Семена с обильным или скудным эндоспермом или вовсе без эндосперма.
В подкласс гамамелидид входят порядки троходендровых, гамамелис овых, крапивных,, буковых и близкие к ним порядки. Гамамелидиды произошли, вероятно, непосредственно от магнолиид.
Подкласс 4. Кариофиллиды (Caryophyllidae). Обычно травянистые растения, полукустарники или низкие кустарники, редко небольшие деревья. Листья цельные. Сосуды всегда имеются, членики сосудов с простой перфорацией. Устьица с 2 или 3 (редко 4) побочными клетками или побочные клетки отсутствуют. Цветки обоеполые или редко однополые, большей частью безлепестные. Зрелая пыльца обычно 3-клеточная. Оболочка пыльцевых зерен трехбороздная или производная от трехбороздного типа. Гинецей апокарпный или чаще ценокарпный. Семязачатки обычно битегмальные, крассинуцеллятные. Семена большей частью с согнутым периферическим зародышем, обычно с периспермом.
В подкласс кариофиллид входят порядки гвоздичных, гречишных и плюмбаговых. Кариофиллиды произошли, вероятно, от примитивных представителей ранункулид.
Подкласс 5. Дилленииды (Dilleniidae). Деревья, кустарники или травы. Листья цельные или различным образом расчлененные. Устьица разных типов, большей частью без побочных клеток. Сосуды всегда имеются; членики сосудов с лестничной или простой перфорацией. Цветки обоеполые или однополые, с двойным околоцветником или реже безлепестные; у более примитивных семейств околоцветник часто спиральный или спироциклический. Андроцей, когда он состоит из многих тычинок, развивается в центробежной последовательности. Зрелая пыльца 2-клеточная или реже 3-клеточная. Оболочка пыльцевых зерен трехбороздная или производная от трехбороздного типа. Гинецей апокарпный или чаще ценокарпный. Семязачатки обычно битегмальные и большей частью крассинуцеллятные. Семена, как правило, с эндоспермом.
В подкласс входят порядки диллениевых, чайных, фиалковых, мальвовых, вересковых, первоцветных, молочайных и др. По всей вероятности, дилленииды произошли от каких-то древних магнолиид.
Подкласс 6. Розиды (Rosidae). Деревья, кустарники или травы. Листья цельные или различным образом расчлененные. Устьица разных типов, чаще всего без побочных клеток или с 2 побочными клетками. Сосуды всегда имеются, членики сосудов с лестничной или чаще с простой перфорацией. Цветки большей частью обоеполые, с двойным околоцветником или безлепестные. Андроцей, когда он состоит из многих тычинок, развивается в центростремительной последовательности. Зрелая пыльца обычно 2-клеточная. Оболочка пыльцевых зерен трехбороздная или производная от трехбороздного типа. Гинецей апокарпный или ценокарпный. Семязачатки, как правило, битегмальные и крассинуцеллятные. Семена с эндоспермом или без него.
В подкласс розид входят порядки камнеломковых, розовых, бобовых, протейных, миртовых, рутовых, сапиндовых, гераниевых, кизиловых, аралиевых, крушиновых, санталовых и др. Розиды произошли, вероятно, от ближайших предков диллениид.
Подкласс 7. Астериды (Asteridae). Деревья, кустарники или чаще травы. Листья цельные или различным образом расчлененные. Устьица большей частью с 2, 4 (часто) или 6 (редко) побочными клетками. Сосуды всегда имеются, членики сосудов с лестничной или чаще с простой перфорацией. Цветки обычно обоеполые, почти всегда сростнолепестные. Тычинки, как правило, в одинаковом или в меньшем числе с долями венчика. Зрелая пыльца 3-клеточная или 2-клеточная. Оболочка пыльцевых зерен трехбороздная или производная от трехбороздного типа. Гинецей всегда ценокарпный, по-видимому, морфологически всегда паракарпный, обычно из 2 - 5, редко 6 - 14 плодолистиков. Семязачатки всегда унитегмальные, тенуинуцеллятные или редко крассинуцеллятные. Семена с эндоспермом или без него.
В обширный подкласс астерид входят порядки ворсянковых, горечавковых, норичниковых, губоцветных, колокольчиковых, сложноцветных и др. По всей вероятности, астериды происходят от примитивных розид, скорее всего от каких-то древних форм, близких к современным древесным представителям порядка камнеломковых.
Класс однодольных, содержащий около 65 семейств, около 3000 родов и не менее 60 000 видов, подразделяется на 3 подкласса.
Подкласс 1. Алисматиды (Alismatidae). Водные или болотные травы. Устьица с 2 или реже 4 побочными клетками. Сосуды отсутствуют или имеются только в корнях. Цветки обоеполые или однополые. Околоцветник развит или редуцирован, часто отсутствует. Зрелая пыльца обычно 3-клеточная. Оболочка пыльцевых зерен однобороздная, дву-, многопоровая или безапертурная. Гинецей большей частью апокарпный, реже ценокарпный. Семязачатки битегмальные, крассинуцеллятные или реже тенуинуцеллятные. Эндосперм нуклеарный или гелобиальный. Семена без эндосперма.
В подкласс алисматид входят порядки частуховых, водокрасовых, наядовых и др. Алисматиды произошли, вероятно, от какой-то вымершей группы травянистых магнолиид, стоявших близко к предкам современных нимфейных.
Подкласс 2. Лилииды (Liliidae). Травы или вторичные древовидные формы. Устьица аномоцитные или с побочными клетками, обычно с 2 побочными клетками (парацитные). Сосуды только в корнях или во всех вегетативных органах, очень редко отсутствуют. Цветки обоеполые или редко однополые. Околоцветник хорошо развит и состоит из сходных между собой (обычно лепестковидных) или ясно различающихся чашелистиков и лепестков, или же околоцветник редуцирован. Зрелая пыльца обычно 2-клеточная, реже 3-клеточная. Оболочка пыльцевых зерен однобороздная, однопоровая (иногда 1 - 4-поровая) или реже безапертурная. Гинецей, как правило, ценокарпный, редко (у примитивных триурисовых и некоторых примитивных лилейных) более или менее апокарпный. Семязачатки обычно битегмальные или очень редко унитегмальные, крассинуцеллятные или реже тенуинуцеллятные. Эндосперм нуклеарный или реже гелобиальный. Семена обычно с обильным эндоспермом, но у порядка имбирных с периспермом и остатком эндосперма или только с периспермом.
В подкласс лилиид входят порядки лилейных, имбирных, орхидных, бромелиевых, ситниковых, осоковых, коммелиновых, эриокауловых, рестиевых, злаковых и др. Происхождение, вероятно, общее с алисматидами.
Подкласс 3. Арециды (Arecidae). Травы или вторичные древовидные формы. Устьица с 2, 4, 6 (чаще всего с 4) побочными клетками. Сосуды во всех вегетативных органах или только в корнях (аронниковые). Цветки обоеполые или чаще однополые. Околоцветник развит и состоит из очень схожих между собой чашелистиков и лепестков, или он более или менее редуцирован, иногда отсутствует. Цветки собраны в метельчатые или шаровидные соцветия либо в початки, которые большей частью снабжены покрывалом. Зрелая пыльца обычно 2-клеточная. Оболочка пыльцевых зерен разных типов, главным образом однобороздная. Гинецей апокарпный (некоторые пальмы) или чаще ценокарпный. Семязачатки битегмальные и крассинуцеллятные, редко тенуинуцеллятные. Эндосперм обычно нуклеарный. Семена с эндоспермом, как правило, обильным.
В подкласс арецид входят порядки пальмовых, циклантовых, аронниковых, пандановых, рогозовых. Наиболее вероятно, что арециды имеют общее происхождение с лилиидами.
Магнолиопсилы, или двулольные (magnoliopsida, или dicotyledones) класс магнолиопсиды, или двудольные (magnoliopsida, или dicotyledones)

|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'