Семя (А. П. Меликян, М. Г. Николаева, Г. А. Комар)
Семя цветковых растений по развитию и строению существенно отличается от семени голосеменных (см. с. 258 предыдущего тома "Жизни растений"). Это объясняется различиями в строении семязачатка голосеменных и цветковых растений и в не меньшей степени - в характере оплодотворения и последующих процессов, определяющих превращение семязачатка в зрелое семя.
Прежде всего резко различны происхождение и характер запасающих тканей. Запасающая ткань в семени голосеменных представляет собой вегетативную часть женского гаметофита и имеет, следовательно, гаплоидный характер. Эндосперм цветковых формируется,; как уже отмечалось, в результате слияния одного из спермиев с диплоидным ядром центральной клетки зародышевого мешка и у подавляющего большинства растений имеет триплоидный характер. В семени некоторых цветковых растений сохраняется в большей или меньшей степени ткань нуцеллуса, преобразующаяся в диплоидную запасающую ткань - перисперм.
В отличие от голосеменных число семядолей зародыша цветковых обычно строго определенно: их бывает либо две, либо одна (и очень редко 3 или 4).
Наконец, в формировании семенной кожуры большинства цветковых могут принимать участие два интегумента, в то время как семенная кожура голосеменных всегда образуется из одного интегумента.
Семена цветковых чрезвычайно разнообразны по размерам и форме. Они могут достигать нескольких десятков сантиметров (гигантские семена пальм) и быть почти неразличимыми невооруженным глазом. Например, у орхидных размеры семени составляют лишь несколько микрометров.
В большинстве случаев семена по форме шаровидные, удлиненно-шаровидные или цилиндрические. Такую форму следует считать наиболее отвечающей задаче перенесения неблагоприятных условий, так как в этом случае максимальному объему соответствует минимальная поверхность контакта семени с окружающей средой.
Поверхность семени часто более или менее гладкая. Однако нередко встречаются также семена с крыльями, шипами, ребрами, волосками, сосочками и различными другими выростами, формирующимися в основном из тканей семенной кожуры.
Семена многих цветковых растений снабжены так называемым присемянником или ариллусом (от позднелат. и средневековолат. arillus - сушеный виноград; рис. 46). Это более или менее мясистые образования, имеющие вид выроста в области микропиле, халазы или рубчика или же вид пленки, покрывающей часть семени или все семя целиком. Ариллусы большей частью ярко окрашены. Их окраска может варьировать от розовой, красной всех оттенков (у мускатного ореха, бересклета и др.) до желтой (у бобовых) и даже голубой. Иногда ариллусы бывают белыми, вернее, бесцветными (у молочайных, лилейных). Термин "ариллус" является сборным, он объединяет различные образования. Истинный ариллус - это кольцевидное, мешковидное или оболочкоподобное образование, развивающееся, в отличие от ложного ариллуса, в области рубчика (от семяножки) и не срастающееся в остальных частях семени с семенной кожурой. Напротив, ложный ариллус (ариллоид) представляет собой разрастание наружного интегумента в разных областях семязачатка, в семени же - видоизмененное продолжение семенной кожуры (рис. 47).
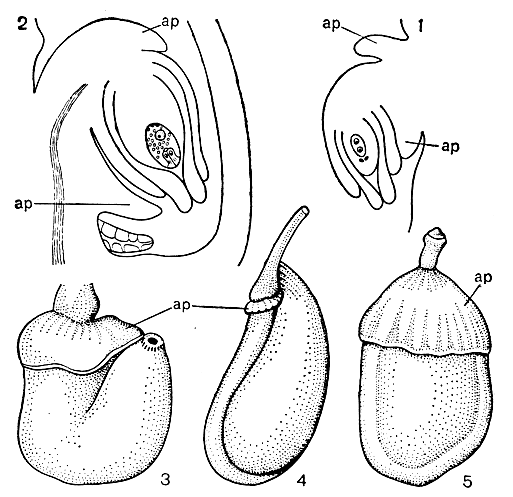
Рис. 46. Настоящие ариллусы (ар): 1 и 2 - заложение и развитие ариллусов у семязачатка булбины однолетней (Bulbina annua); 3 - ариллус в раскрывшемся цветке гиббертии вьющейся (Hibbertia volubilis); 4 - заложение ариллуса у пассифлоры трехлопастной (Passiflora triloba); 5 - зрелый ариллус у того же растения
Все перечисленные выше образования на поверхности семени являются приспособительными, играющими важную роль в процессе распространения семян и тем самым в расселении видов. В частности, ариллусы служат приспособлением растений для привлечения животных и, следовательно, средством для распространения семян с помощью животных, особенно птиц и муравьев. Благодаря некоторым особенностям строения ариллусов они могут также способствовать распространению семян с помощью ветра, воды и т. д. Наконец, они могут служить и средством для раскрывания плода (у мускатного ореха), для отделения семени от плаценты (у молочайных) и т. д.
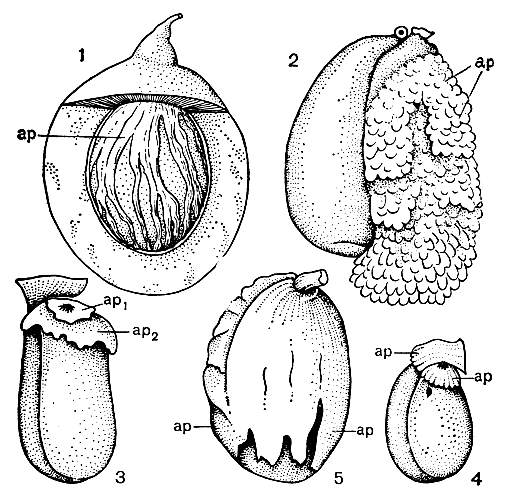
Рис. 47. Некоторые типы ариллоидов (ар): 1 - у зрелого плода миристики душистой, или мускатного ореха (Myristica fragrans); 2 - у семени копытня канадского (Asarum canadense); 3 - у семязачатка клузии желтой (Clusia flava); 4 - у молодого семязачатка бересклета широколистного (Eurnymus latifolia); 5 - у зрелого семязачатка того же растения
Ткань ариллуса богата различными включениями. Жирные масла были обнаружены у представителей мускатниковых, бобовых, страстоцветных, лилейных, амариллисовых; дубильные вещества - у коннаровых и крыжовниковых; рафиды оксалата кальция - у диллениевых; протеин и крахмал - у бобовых, коннаровых, страстоцветных, мускатниковых, лилейных.
До сих пор речь шла о видоизменениях в области семенной кожуры. Что касается лежащего глубже эндосперма, то его поверхность обычно остается гладкой. Однако у представителей некоторых семейств (анноновые, маслиновые, пальмы и др.) она (вместе с семенной кожурой) становится причудливо извилистой, складчатой, вследствие чего в период до созревания семени значительно увеличивается площадь его соприкосновения с окружающими материнскими тканями и возрастает интенсивность обмена веществ между ними. Такой эндосперм, хорошо знакомый нам по семени мускатного ореха, называется морщинистым или руминированным.
Внешне строение зрелых семян отражает некоторые морфологические признаки семязачатков, из которых они сформировались. На семенах обычно заметны микропиле, рубчик и семяшов. Микропиле сохраняется в виде отверстия, которое изнутри часто закупоривается клетками семенной кожуры. Иногда микропилярное отверстие прикрывается специальной крышечкой, формирующейся из клеток интегументов (некоторые нимфейные); реже микропиле на семени не заметно. Рубчик образуется в том месте, где семя отделилось от семяножки. В анатропных семязачатках семяножка оказывается как бы приросшей к семязачатку и на зрелом семени можно различить своеобразный семяшов. При этом рубчик и микропиле располагаются рядом.
Главной частью семени является зародыш. По форме зародыш может быть прямым, согнутым, скрученным, спиральным, кольцевидным, подковообразным и т. д. Различно и положение зародыша в семени. У многих растений он занимает место в центре и со всех сторон окружен запасающей тканью; у других зародыш расположен по периферии семени и тогда в большей или меньшей степени окружает запасающую ткань или примыкает к ней. Несмотря на большое разнообразие формы зародыша и его положения в семени, расположение зародышей в семенах в одном отношении одинаково - кончик корня всегда находится поблизости от микропиле, благодаря чему при прорастании из семени прежде всего выходит корень.
Степень развития зародыша в зрелом семени у разных групп цветковых растений различна. Для наиболее примитивных групп характерен так называемый недоразвитый, слабо дифференцированный зародыш. В таких семенах в момент отделения их от материнского растения зародыш иногда очень мелкий, точечный, представлен лишь небольшой группой клеток. Однако ко времени прорастания в этих семенах формируется довольно крупный и хорошо дифференцированный зародыш.
У некоторых паразитных и других высокоспециализированных цветковых растений семена содержат так называемый рудиментарный зародыш. Он мелкий, точечный и к моменту прорастания не дифференцируется. Освобождаясь от оболочки семени, он формирует своеобразную протосому, на которой в дальнейшем возникают почки.

Рис. 48. Поперечный разрез семейной кожуры магнолии крупноцветковой (Magnolia grandiflora): 1 - эпидерма; 2 - слой уплощенных тонкостенных клеток; 3 - мясистая ткань из тонкостенных паренхимных клеток с заключенными между ними маслянистыми клетками; 4 - тонкий слой клеток, отделяющий мясистый слой от расположенного ниже каменистого слоя; 5 - слой каменистых клеток; в - слой тонкостенных клеток; 7 - слой клеток с темно-бурым содержимым; 8 - внутренняя эпидерма; 9 - клетки нуцеллуса
У наиболее подвинутых групп цветковых растений зародыш в семенах хорошо развит, он крупный, а эндосперм и перисперм редуцируются и нередко совсем исчезают, питательные вещества при этом отлагаются в органах зародыша, главным образом в семядолях.
В момент прорастания зародыш питается гетеротрофно, используя заключенные в семени запасы питательных веществ. В зависимости от приуроченности запасающих тканей к тем или иным частям семени у цветковых растений обычно выделяют 4 типа семян. У семян первого типа в качестве запасающей ткани развит перисперм (гвоздичные, маревые). В семенах второго типа имеются и перисперм и эндосперм (нимфейные, перцевые), в семенах третьего типа, характерных для подавляющего большинства цветковых, перисперм отсутствует, но сильно развит эндосперм. Наконец, у семян четвертого типа запасные питательные вещества сосредоточены в семядолях зародыша (бобовые, сложноцветные, тыквенные и др.). У некоторых, особенно примитивных, цветковых часть питательных веществ сохраняется в клетках семенной кожуры.
Семенная кожура (теста, спермодерма) играет важную роль как пограничная зона между зародышем, запасающими тканями и внешней средой. Она обычно достаточно плотная и крепкая и надежно предохраняет зародыш и запасающие ткани от механических, биотических и иных повреждений (рис. 48 и 49). Семенная кожура препятствует иссушению и предотвращает преждевременное насыщение влагой содержимого семени. С другой стороны, она не должна препятствовать проникновению влаги и прорастанию зародыша в нужный момент.
Функции интегументов и их производного - семенной кожуры - меняются в ходе формирования семени из семязачатка. Обычно покровы семязачатка многослойные (особенно у представителей примитивных групп), до оплодотворения все клетки интегументов заполнены питательными веществами. Наличие кутикулярных пленок на поверхности эпидермы интегументов не является преградой для прямого поступления питательных, транзиторных веществ из интегументов в зародышевый мешок. Изменения в гомогенной структуре покровов семязачатка происходят после оплодотворения. В это время начинается усиленный отток питательных веществ из клеток интегументов в клетки эндосперма и зародыша.
Зрелая семенная кожура может складываться из разных типов тканей, главными из которых являются саркотеста (наружная, сочная, мясистая ткань), склеротеста (механическая ткань), паренхотеста (паренхимная ткань) и миксотеста (ослизняющаяся ткань). В семенных покровах различных растений встречаются либо все названные типы тканей, либо некоторые из них (рис. 49).
Таким образом, цветковые растения характеризуются большим разнообразием в строении семенной кожуры. Особенно разнообразна ее анатомическая структура (рис. 49). По меткому выражению Н. В. Цингер, "ни один растительный орган или ткань не может сравниться с семенной кожурой по богатству и разнообразию анатомических структур". Роды и виды растений хорошо различаются по строению семенной кожуры, поэтому изучение ее структуры очень важно для диагностики и систематики цветковых растений.
Рост и развитие семени протекают на фоне активных биохимических процессов. Из материнского растения к нему интенсивно притекают питательные вещества (сахара, аминокислоты, витамины и многие другие), в семени идут энергичные процессы синтеза. В этот период в семенах содержится большое количество таких гормонов, как ауксины, гиббереллины и цитокинины, осуществляющие регулирование процессов, связанных с ростом.
Интересным, но до сих пор не разгаданным с точки зрения физиологии является тот факт, что у многих растений зародыш останавливается в росте задолго до наступления зрелости семени, не достигнув полного развития. У одних видов, например у женьшеня, лимонника китайского, эта остановка в росте, как показал И. В. Грушвицкий, происходит очень рано; в этих случаях почти не дифференцированный зародыш с трудом можно обнаружить в зрелом семени. У ясеня обыкновенного, видов борщевика остановка роста семени происходит позже, когда зародыш имеет довольно большие размеры и у него можно различить семядоли и осевую часть, однако он занимает по длине лишь часть семени. Известны случаи, когда зародыш, казалось бы, уже вполне развит и по длине почти равен длине семени (виды бересклета), однако анатомические исследования показали, что у него процесс формирования завершен еще не полностью.
Содержание воды в семенах на начальных фазах их формирования очень высокое, но по мере роста и созревания происходит их обезвоживание. Снижение влажности имеет место даже у семян, заключенных в сочные плоды, очень богатые влагой, в такие, как лимон, вишня, арбуз.
Наконец, семя достигает окончательных размеров и процессы роста останавливаются. Однако еще некоторое время продолжается приток веществ в семена из материнского растения. Теперь они начинают откладываться в запасной форме - в виде труднорастворимых, малоподвижных соединений, таких, как крахмал, жиры, запасные белки. В зависимости от того, какие запасные вещества преобладают, семена разделяют на крахмалистые (пшеница, кукуруза и другие злаки), масличные (подсолнечник, лен, арахис) или белковые (большинство бобовых). Благодаря накоплению этих и других веществ семена ряда растений являются для человека неоценимым источником пищевых (мука, крупы, растительные масла) и многих полезных лекарственных и технически ценных продуктов. Что же касается самих растений, то запасные вещества их семян необходимы для питания прорастающего зародыша.
Завершение роста плода и семени сопровождается снижением активности таких гормонов, как гиббереллины и цитокинины. В то же время вместе с отложением запасных веществ в семенах происходит накопление индолилуксусной и абсцизовой кислот. Содержание их возрастает до такого уровня, при котором они не стимулируют, а тормозят рост, что является важным приспособительным свойством, так как предупреждает возможность несвоевременного прорастания семян.
По мере наступления зрелости затухает активность ферментов, влажность семян снижается до минимума (5 - 10%), поступление веществ из растения прекращается. Покровы семени претерпевают существенные изменения: их ткани отмирают, уплотняются, одревесневают.
Строение зрелых семян, в особенности их покровов, биохимические и физиологические свойства зародышей, необычайная способность семян высыхать, не теряя жизнеспособности, определяют их свойство находиться в покое. При этом семена многих цветковых растений, переживая различные неблагоприятные воздействия, сохраняют длительное время способность прорасти и дать жизнь новому растению. Далеко не все семена обладают способностью прорастать сразу после созревания. Легко прорастают семена многих растений, живущих в жарких странах с влажным климатом, например растения мангровых зарослей. Семена видов ризофоры прорастают уже на дереве, проросток падает в насыщенный водой ил и легко укореняется. Растения, образующие легко и быстро прорастающие семена, известны и в нашем умеренном поясе. В большинстве случаев это растения, цветущие ранней весной, например многие виды ивы, серебристый клен. При благоприятных условиях их семена, созревающие в начале лета, сразу прорастают и к осени образуются уже окрепшие растеньица, способные пережить зиму. Не успевшие прорасти семена таких растений быстро теряют всхожесть.
У подавляющего большинства цветковых растений созревшие семена не способны сразу прорастать, находясь в состоянии так называемого покоя. Покой может быть вынужденным, связанным с отсутствием необходимых для прорастания условий (температурных, влажности и т. д.). Наиболее обычным случаем вынужденного покоя являются сухие семена. Наряду с этим семена растений, обитающих большей частью в областях с сезонными колебаниями температуры и влажности (в умеренном и субтропическом поясах, а также в некоторых тропических областях, характеризующихся сменой засушливых и дождливых периодов), обладают способностью находиться в состоянии органического покоя; он определяется особыми свойствами самого семени. Термин "покой" при этом имеет условное значение. В большинстве случаев в таких семенах протекают достаточно активные метаболические процессы (дыхание, а иногда и рост зародыша), но прорастание заторможено. Семена, находящиеся в органическом покое, даже в условиях, благоприятных для прорастания, не прорастают совсем или прорастают медленно и дают лишь небольшой процент всходов. Для того чтобы приобрести способность прорастать, они должны подвергнуться действию особых условий, как правило, не совпадающих с условиями их прорастания.
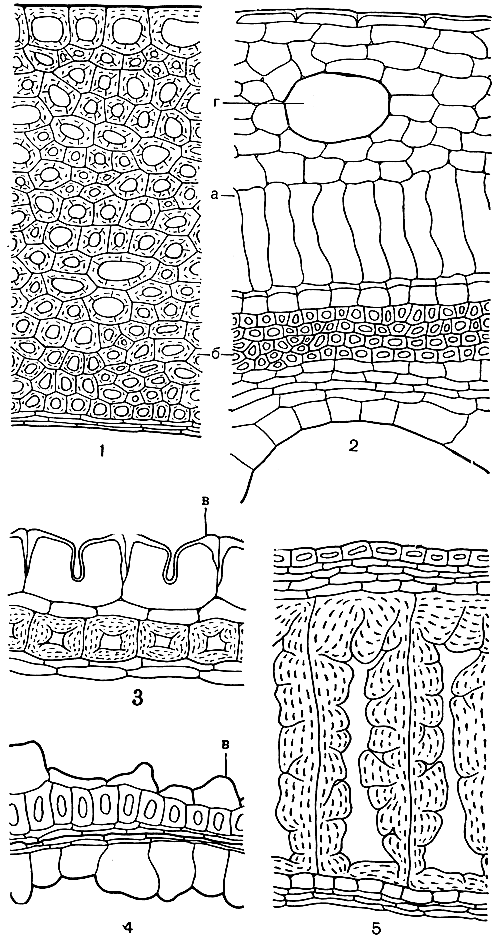
Рис. 49. Некоторые типы семенной кожуры: 1 - гамамелис вирджинский (Hamamelis virginiana, увел. 200); 2 - эвпоматия лавровая (Eupomatia laurina, увел. 200); з - актинидия мозолистая (Actinidia callosa, увел. 400); 4 - платан восточный (Platanus orientalis, увел. 400); 5 - конопля посевная (Cannabis sativa, увел. 300); а - саркотеста; б - склеро-теста; в - ослизняющиеся клетки; г - эфирномасличный канал
Способность семян находиться в вынужденном или органическом покое выработалась у растений в ходе эволюции как средство пережить неблагоприятное для роста проростка время года. Таким путем создается запас семян в почве. Если в силу каких-либо неожиданных неблагоприятных погодных колебаний, например поздневесенних заморозков или периодов засухи, появившиеся проростки погибнут, в следующие годы смогут прорасти семена, сохранившиеся в почве в покоящемся состоянии, и таким образом будут воспроизведены растения данного вида. Покоящиеся семена иногда могут лежать в почве в набухшем состоянии, не теряя жизнеспособности, в течение многих лет.
У многих видов препятствием для прорастания семян является водонепроницаемость кожуры, что обусловлено присутствием в ней так называемого палисадного слоя, состоящего из сильно удлиненных толстостенных клеток. Кроме того, кожура покрыта водонепроницаемой воскообразной пленкой - кутикулой. Согласно наблюдениям А. В. Попцова, явление "твердосемянности" представляет собой истинный покой: до тех пор пока к зародышу не проникнет вода, все процессы жизнедеятельности в семени подавлены почти полностью. Твердосемянность широко распространена у растений из семейств бобовых и некоторых других и особенно у растений, произрастающих в засушливых областях. В природе "твердые" семена приобретают способность набухать обычно под влиянием температурных воздействий (прогревание, промораживание или резкие колебания температуры), приводящих к нарушению целостности кожуры. В практике сельского хозяйства их подвергают перед посевом специальной обработке (перетирание с песком, ошпаривание кипятком и т. д.).
Известны виды, у которых торможение прорастания связано с присутствием в околоплоднике нераскрывающихся плодов веществ, ингибирующих прорастание. Такие семена прорастают после промывания плодов, разрушения или удаления околоплодника. Наиболее ярким примером такого "химического" покоя являются соплодия свеклы.
У различных видов из многих семейств семена находятся в так называемом морфологическом покое, связанном с недоразвитием зародыша. До тех пор, пока такие семена не попадут в условия, при которых зародыш закончит развитие, они не смогут прорасти. Процесс доразвития обычно протекает в набухших семенах в тепле (+10...+ 25° С).
Особенно широко распространен покой семян, вызываемый физиологическими причинами. Причины торможения прорастания таких семян представляют предмет исследования многих ученых. Полученные данные показывают,, что в этом случае действует так называемый физиологический механизм торможения прорастания, в котором соединяются пониженная способность зародыша к росту и пониженная газопроницаемость окружающих его покровов. Состояние зародышей покоящихся семян зависит от неблагоприятного для процессов роста баланса гормонов и низкой активности ферментов.
Физиологический покой семян различается по глубине. Свежесобранные семена хлебных злаков, салата, подсолнечника, многих сорных и других растений находятся в неглубоком покое. Достаточно повредить или удалить покровы, как они прорастают. В природе такие семена постепенно приобретают способность прорастать за время сухого хранения или под влиянием хотя бы кратковременного охлаждения в набухшем состоянии. Многие из них являются светочувствительными, т. е. сухое хранение или охлаждение у них может быть заменено облучением.
Изучение причин светочувствительности семян представляет собой особую проблему, которой посвящено огромное количество работ. Немецкий ученый Вильгельм Кинцель исследовал семена более 900 видов растений и нашел, что у 672 из них семена в той или иной степени реагируют на свет. Классическими объектами изучения светочувствительности являются семена салата, сельдерея, видов березы и ряда других растений. Установлено, что наиболее сильное действие оказывают лучи красной части спектра. Реакция на свет в большой степени зависит от температурных условий, в которых находятся набухшие семена. Например, у салата она отсутствует при температуре + 15° С и ниже и сильно проявляется при ее повышении. Выдерживание таких семян в темноте при повышенной температуре вызывает у них термопокой, гораздо более глубокий, чем покой первичный. Семена некоторых видов, например видов чернушки, наоборот, на свету не способны прорастать; их называют темновсхожими. В основе реакции семян на свет лежит присутствие в них особого пигмента - фитохрома, переходящего под влиянием красного света в активную, а под влиянием дальнего красного - в неактивную форму.
Прорастание семян, характеризующихся неглубоким физиологическим покоем, можно стимулировать, намачивая их в растворах гиббереллина, а в некоторых случаях цитокининов и ряда других веществ.
Значительно труднее вызвать прорастание семян, находящихся в глубоком физиологическом покое, характерном для многих древесных (виды яблони, клена и др.) и некоторых травянистых растений. У таких семян удаление покровов хотя и вызывает некоторый рост зародышей, но он замедлен и зачастую ненормален: растут главным образом семядоли, а рост осевых частей зародыша очень затруднен. После высадки таких проростков в землю нередко образуются розеточные растения, так как у них заторможен и рост побега.
Явление ненормального роста зародышей, извлеченных из семян, находящихся в глубоком покое, получило название физиологической карликовости. Американский ученый Флоренс Флемион в течение 10 лет выращивала персик в теплой оранжерее. Все это время он сохранял черты карликовости.
Преодоление глубокого покоя происходит при посеве семян осенью или в ходе так называемой искусственной холодной стратификации под влиянием длительного выдерживания семян при пониженной положительной температуре (0... + 70 С), во влажной среде (в песке, торфе, опилках), в условиях хорошей аэрации. Длительность и оптимальная температура стратификации различна для разных видов*: семенам клена татарского необходимо 2 - 4-месячное пребывание при 0... +3°C. У некоторых растений семена наиболее успешно прорастают после стратификации при +5... + 7° С. Семена сорного растения недотроги мелкоцветковой (Impatiens parviflora) прорастают только после 5,5 - 6 месяцев стратификации при температуре не выше +3° С. После необходимого периода охлаждения семена приобретают способность прорастать в тепле; оптимальной температурой прорастания у большинства видов этой категории является температура +9... 4 - 10° С. Если же прервать холодную стратификацию даже кратковременным действием повышенной температуры (в природе это нередко происходит благодаря ранним оттепелям), изменения, происшедшие под влиянием пониженной температуры, исчезают и семена впадают в состояние вторичного покоя. Чем выше температура во время перерыва стратификации, тем быстрее возникает вторичный покой. Например, у семян клена татарского это происходит при +15° С за 10, а при +25° С за 5 суток. Для проращивания таких семян стратификацию необходимо начинать сначала. Это свойство семян имеет большое приспособительное значение, предотвращая провокационное прорастание.
Торможение прорастания семян нередко вызывается сочетанием недоразвития зародыша с действием физиологического механизма торможения прорастания различной силы. Семена,, находящиеся в таком морфофизиологическом покое, должны подвергаться сначала теплой стратификации, для того чтобы прошло доразвитие зародыша, а затем действию пониженной температуры.
До последнего времени холодная стратификация считалась единственным способом преодоления глубокого покоя. Однако в последние годы работами, проведенными в Ботаническом институте им. В. Л. Комарова под руководством М. Г. Николаевой, было установлено, что у некоторых растений, в частности у разных видов клена, можно вызвать прорастание значительной части семян без всякой стратификации путем замачивания их в течение двух суток в растворе разных цитокининов, удалив предварительно околоплодник. Оказалось, что у бересклета и женьшеня также можно вызвать прорастание семян без указанной выше длительной и сложной стратификации. После двухдневного намачивания в растворе гиббереллина, а затем в течение такого же времени в растворе кинетина значительная часть семян прорастает при +9... +10° С в течение 30 - 40 суток.
У некоторых растений покой семян имеет особый характер. Так, семена калины обыкновенной прорастают без стратификации и у них развивается хорошая корневая система. Однако побег не будет образовываться до тех пор, пока семена не подвергнутся длительной холодной стратификации, так как в глубоком физиологическом покое находится эпикотиль - точка роста побега. Еще более сложен покой у семян ландыша. Семена этого растения должны подвергнуться длительной холодной стратификации, после чего у них появится корешок. Однако побег начнет развиваться только после того, как проросшие семена пробудут лето в тепле, а затем подвергнутся действию второго периода пониженной температуры. У таких семян двойной покой: покой гипокотиля и зародышевого корня и покой эпикотиля. В естественных условиях они прорастают только на вторую весну.
У очень многих растений торможение прорастания семян обусловлено несколькими причинами (комбинированный покой). Известны пустынные виды рода ферула (Ferula), в околоплоднике которых содержатся сильные ингибиторы прорастания; сами семена содержат недоразвитый зародыш и находятся, кроме того, под контролем физиологического механизма торможения прорастания. Не менее многообразны причины, вызывающие торможение прорастания у семян известных декоративных кустарников - снежной ягоды и падуба. Они обладают твердым околоплодником и очень маленьким недоразвитым зародышем. Для того чтобы вызвать прорастание, такие семена приходится подвергать сложной предпосевной обработке. Без этого в питомниках семена могут лежать в земле в течение нескольких лет, образуя так называемые мертвые посевы и давая лишь единичные всходы.
Набухшие семена, находящиеся в органическом покое, в большинстве случаев особенно устойчивы к неблагоприятным условиям среды. Длительность жизни зрелых сухих семян различна. Обычно семена, которые легко прорастают, теряют всхожесть в течение нескольких месяцев, недель и даже дней. К числу таких семян относятся семена большинства видов ивы, серебристого клена и некоторых других растений. У других видов семена сохраняют жизнеспособность в течение нескольких лет, а иногда в течение десятков и даже сотен лет. Если подсушенные семена хранить в герметических условиях при температуре, близкой к абсолютному нулю (-273° С), они сохраняют жизнеспособность практически бесконечно.
Часто упоминаемый пример долговечности семян пшеницы, найденных в египетских пирамидах, оказался недостоверным. В действительности всхожесть семян этого растения сохраняется лишь в течение нескольких лет. Наибольшая долговечность свойственна "твердым" семенам. Так, удалось прорастить семена лотоса, пролежавшие погребенными в торфянике в Маньчжурии по первоначальной оценке более 200 лет. Однако, согласно современным данным радиоизотопного анализа, возраст этих семян превышает 1000 лет. Еще большую долговечность обнаружили семена люпина, извлеченные из погребенных льдов Аляски: их возраст, как показал также радиоизотопный анализ, достигает 10 000 лет.
Для того чтобы семена могли прорасти, необходимы благоприятные условия увлажнения, аэрации и температуры. Диапазон температур, при которых семена способны прорастать, зависит от географического происхождения растений. Температура, при которой происходит прорастание семян, у северных растений ниже, чем у южных, особенно тропических видов. Семена пшеницы, например, могут прорастать при 0... + 10 С, между тем для прорастания семян кукурузы необходима температура не ниже +12° С. Минимальная температура, необходимая для прорастания семян дыни и особенно семян многих тропических растений, например пальм, составляет +20 и даже + 25° С.
Прорастание семян сопровождается сложными биохимическими и анатомо-физиологическими процессами. Как только в семена начинает поступать вода, в них прежде всего резко усиливается дыхание и одновременно активизируются различные ферменты, образовавшиеся еще в период созревания. Под их влиянием запасные питательные вещества гидролизуются, превращаясь в подвижную легко усвояемую форму. Жиры и крахмал превращаются в органические кислоты и сахара, белки - в аминокислоты. Перемещаясь в зародыш из запасающих органов, питательные вещества становятся субстратом для начинающихся в нем процессов синтеза, в первую очередь новых нуклеиновых кислот и ферментативных белков, необходимых для начала роста.
Рост зародыша обычно начинается с прорыва покровов энергично удлиняющимся гипокотилем и зародышевым корнем. Это удлинение происходит за счет растяжения образующих их клеток. Позднее начинается деление клеток. После появления корня начинают расти и зеленеть семядоли, наконец, появляется побег и образуются настоящие листья. У одних растений (тыква, подсолнечник, клен) семядоли в процессе роста освобождаются от окружающих покровов, выносятся гипокотилем над поверхностью почвы и становятся первыми ассимилирующими органами нового растения. Такой тип прорастания носит название надземного. У других растений (дуб, каштан) семядоли остаются в семени под землей и являются источником питания проростка (подземное прорастание). После появления над поверхностью почвы побега с листьями эти проростки также переходят к автотрофному типу питания.
Таким образом, благодаря различным необычным свойствам удивительного образования, каким являются семена, они могут длительное время сохранять в себе живой зародыш и в состоянии покоя благополучно переживать различные, иногда весьма неблагоприятные воздействия. Приспособления, способствующие переживанию неблагоприятных условий, и факторы, выводящие семена из покоя, определяются характером этих условий. Семена, вышедшие из состояния покоя, при благоприятных условиях успешно прорастают, давая начало новым растениям.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'