Абиотическое опыление (А. Н. Пономарев, Е. И. Демьянова)
Анемофилия. Опыление ветром у покрытосеменных вторично. Анемофильные группы их произошли от энтомофильных предков. Анемофилия характеризуется высокой специализацией. Это представление отнюдь не исключает возможность случайного опыления ветром у архаичных форм. У последних, как говорилось, допускается совмещение разных способов опыления. Существовавшее некогда мнение о первичности ветроопыления и примитивности анемофильных покрытосеменных сейчас полностью оставлено.
Анемофилия представляет особую форму адаптации цветковых растений к неблагоприятным условиям, ограничивающим возможности биотического опыления. В средних и особенно высоких широтах ветер является весьма важным, а иногда даже более надежным агентом опыления, нежели насекомые.
Количественное соотношение энтомофильных и анемофильных родов и видов в мировой флоре цветковых растений не установлено. Ветроопыление указывается рядом исследователей (Куглер, 1970; Фэгри и ван дер Пэйл, 1970) примерно для 21% цветковых растений ФРГ и ГДР. Соответствующие подсчеты недавно были сделаны Эрихом Дауманом и для Чехословакии. Чехословацкая флора характеризуется следующим количественным соотношением родов по способу опыления: энтомофильные роды составляют 74,3%, анемофильные - 17,3%, гидрофильные - 0,5%, переходные типы - 2,8%, сомнительные случаи - 5,1%. Однодольные и двудольные растения весьма различаются в этом отношении. Раздельные подсчеты показали, что пропорция энтомофильных и анемофильных родов у двудольных растений составляет 87 и 4,6%, а у однодольных соответственно 33 и 59%. Высокая пропорция анемофильных родов среди однодольных связана с тем, что во флоре Чехословакии большую роль играют такие крупные анемофильные семейства однодольных, как злаковые и осоковые.
Для правильной оценки относительного значения энтомофилии и анемофилии необходимо учитывать не только количественное соотношение энтомофильных и анемофильных таксонов во флоре, но и их роль в растительных сообществах Земли. По мнению Е. М. Лавренко, большинство видов, доминирующих в растительном покрове внетропической суши, "вверяют перенос своей пыльцы ветру". Господствуя в растительных сообществах на огромных пространствах, они являются подлинными "победителями в борьбе за существование" и выражают наиболее полное приспособление их к современным экологическим условиям внетропических областей, в том числе и к условиям опыления. Таковы, например, злаки в степях, на лугах и в саваннах, различные маревые и полыни в пустынях, карликовые березки в тундре, лесооб-разующие древесные породы (береза, осина, дуб, граб, орешник) в листопадных лесах, многочисленные виды осок и пушиц на болотах и в тундре и т. д. В Арктике, в степях и пустынях, на болотах, на морских побережьях, в листопадных лесах умеренного климата ранней весной анемофильные растения играют ведущую роль, если не по количеству видов, то по количественному участию их особей в растительных сообществах. Вероятно, так именно обстоит дело и в высокогорьях, хотя, по подсчетам уже упомянутого Куглера, количество анемофильных видов с подъемом в горы уменьшается. Однако в данном случае имеет значение не количество анемофильных видов, а их роль в растительном покрове высокогорий.
В тропической зоне, прежде всего в дождевых тропических лесах, изобилующих животными-опылителями, многие виды растений, в том числе некоторые злаки, опыляемые в умеренном климате ветром, представлены здесь энтомофилами. Мощная завеса листьев, фильтрующая пыльцу, и ежедневные ливневые дожди, вымывающие пыльцу из атмосферы, неблагоприятны для опыления ветром. В дождевых тропических лесах существует даже ярусность в способах опыления: деревья верхних ярусов тяготеют к анемофилии, а в более нижних ярусах возрастает значение энтомофилии. Кроме того,, в тропических дождевых лесах необычайно разнообразен состав древесных пород и трудно встретить деревья одного и того же вида, растущие по соседству. Это обстоятельство также неблагоприятно для анемофилии.
Опыление ветром у цветковых растений отнюдь не является возвратом к более примитивному способу опыления, свойственному голосеменным. Анемофилия у цветковых возникла на основе обоеполого энтомофильного цветка. Она представляет особое направление их приспособительной эволюции в условиях недостатка насекомых. Это не возврат к прошлому, а дальнейшее развитие процесса опыления цветковых растений. Стоит только вспомнить, что у анемофильных цветковых растений выработались рыльца с огромной воспринимающей поверхностью, улавливающей пыльцу из воздуха, которых вообще нет у голосеменных. Переход от энтомофилии к анемофилии вызвал глубокую структурную перестройку цветка и соцветия.
Анемофилия характеризуется обычно только с морфологической стороны. Такую анемофилию можно назвать структурной. Для нее характерны следующие признаки: отсутствие или значительная редукция околоцветника, образование множества мелких невзрачных цветков, лишенных окраски, запаха и нектара и собранных в соцветия (сережка, кисть, метелка, колосья); цветки нередко однополые в однодомном или двудомном распределении, часто протогиничны; пыльцы много, она легкая, сыпучая, летучая; экзина тонкая и гладкая, лишенная скульптуры; рыльца относительно долговечны и имеют большую поверхность для улавливания пыльцы из воздуха (рис. 39); количество семязачатков в завязи уменьшено до одного.
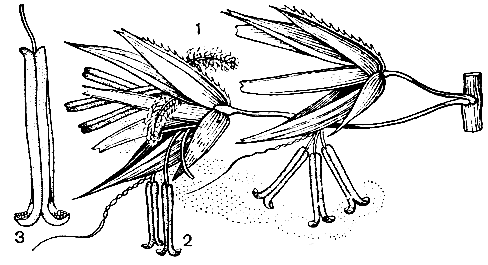
Рис. 39. Ветроопыление у злака райграса высокого (Arrhenatherum elatius): 1 - перистые рыльца; 2 - пыльники на длинных тычиночных нитях; 3 - начало вскрывания пыльников
Перечисленные признаки структурной анемофилии общеизвестны, хотя не все вполне бесспорны. Некоторые из них (обилие, легкость, летучесть пыльцы) встречают серьезные возражения, о чем будет сказано ниже.
У ветроопыляемых растений раздельнополость цветков представляет более частое явление, чем у насекомоопыляемых. В Средней Европе, по данным Куглера, около 99% энтомофильных растений имеют обоеполые цветки, а у анемофильных - 67%. Отсюда следует, что тенденция к разделению полов действительно характеризует анемофилию. Раздельнополость цветков отнюдь не представляет исключительной принадлежности анемофилии. Это явление встречается и у энтомофилов, но гораздо реже. С другой стороны, некоторые типичнейшие анемофилы имеют обоеполые цветки, например злаки. Однако раздельнополость столь часто встречается у анемофильных растений, что ее можно считать характерной чертой анемофилии, для которой она, очевидно, дает какие-то преимущества.
Раздельнополость у анемофилов нередко сочетается с дихогамией, особенно с протогинией. Как уже говорилось, протогинию правильнее рассматривать как приспособление к кратковременному суточному рассеиванию пыльцы, ограниченному немногими часами или даже минутами.
Спорным является вопрос о количестве пыльцы и дальности рассеивания ее у анемофилов. Отнюдь не доказано, что ветроопыляемые растения продуцируют пыльцы (в расчете на одну семяпочку) больше, чем насекомоопыляемые. Нам лишь кажется, что ветроопыляемые растения продуцируют много пыльцы, так как они рассеивают ее вне цветка, в воздухе, в виде облачка. Энтомофилы же не пылят, и мы не видим скопления их пыльцы вне цветка. Раньше было принято считать анемофилию эволюционно более примитивным способом опыления на том основании, что она якобы неэкономна, поскольку много пыльцы (драгоценного материала) рассеивается в воздухе и растрачивается напрасно. Это неверно. Представление об экономичности опыления страдает антропоморфизмом. Пыльца рассеивается в воздухе не напрасно, о чем свидетельствует высокая эффективность перекрестного опыления у злаков. Наконец, у анемофилов имеется своеобразная экономность в рассеивании пыльцы. Она проявляется в суточной ритмике опыления и в наличии взрывчатого и порционного цветения (см. ниже). С другой стороны, у энтомофилов также возможны своя неэкономность и несовершенство в расходовании пыльцы, проявляющиеся как издержки специализации в случае отсутствия соответствующих насекомых-опылителей или несовпадения ареалов растения и его опылителя.
Вопрос о пыльцевой продукции анемофильных и энтомофильных растений исследован немецким ученым Францем Полем. Оказалось, что не существует принципиального различия между ветроопыляемыми и насекомоопыляемыми растениями по количеству продуцируемой пыльцы. Разные виды продуцировали то большее, то меньшее количество пыльцы на одну семяпочку, независимо от того, были ли они анемофилами или энтомофилами. Например, анемофильная рожь образует пыльцы на один семязачаток меньше (52 310), чем энтомофильные конский каштан, клен явор, груша (соответственно 451 543, 94 078 и 60 778), но больше, чем энтомофильная липа мелколистная (43 500). В свою очередь, последняя продуцирует больше пыльцы, чем такие типичнейшие анемофилы, как кукуруза (14 636) и береза (6734). Таким образом, представление о более высокой пыльцевой продукции анемофилов по сравнению с энтомофилами не нашло поддержки в полученных фактических данных. Оно крайне сомнительно и, видимо, должно быть оставлено.
Принято думать, что пыльца у анемофилов легкая, летучая и разносится ветром на большие расстояния. С этим нельзя согласиться. В действительности дело обстоит совсем наоборот. Пыльца у ветроопыляемых растений тяжелая, так как она богата крахмалом. Поэтому такая пыльца разносится воздушными течениями на небольшие расстояния. Прямые наблюдения показали, что пыльца злаков, пустынных маревых, многих ветроопыляемых древесных пород (береза, дуб, граб) оседает неподалеку от продуцирующих ее растений. Изучение рассеивания пыльцы дикорастущих злаков в штате Небраска, проведенное Джонсом и Ньюэлом, показало, что содержание злаковой пыльцы в приземном слое воздуха составляло на дистанции 25 м от источника ее 31 % , 75 м - 10% , 125 м - 4,4%, 200 м - 1,2%, 300 м - 0,8% от того количества ее, которое имелось в центре поля. У такого строго перекрестноопыляемого растения, как кострец безостый, на 16 мм2 предметного стекла на расстоянии 300 м от поля было уловлено только 3 пыльцевых зерна, или 0,5% от количества пыльцы в пределах поля. Подобные же данные получены и для многих других ветроопыляемых растений.
Вопрос о сфере эффективного ветроопыления имеет важное значение в селекции при определении дальности изоляции чистосортных посевов во избежание "загрязнения" их пыльцой других сортов. Изоляция в 300 - 500 м считается достаточной для этой цели. Для иллюстрации возможности ветроопыления на дальней дистанции обычно приводится факт заноса большого количества пыльцы дуба на остров Гельголанд с соседнего материка на расстояние примерно 60 - 70 км от ближайших дубовых насаждений. Известны случаи заноса пыльцы древесных пород в Арктику на многие сотни километров. Факты эти не подлежат сомнению. Однако биологическое значение дальнего воздушного транспорта пыльцы очень сомнительно для целей опыления. Пыльца, находясь в воздухе, быстро утрачивает жизнеспособность, как это было установлено для злаков. Эффективное перекрестное опыление при помощи ветра осуществляется на небольшом расстоянии, между ближайшими особями, удаленными друг от друга самое большее на несколько десятков или сотен метров. Иначе говоря, популяции ветроопыляемых видов более замкнуты, чем это до сих пор казалось, поскольку сфера эффективного ветроопыления у них ограничена небольшими дистанциями.
Мы склонны думать, вопреки обычному мнению, что именно тяжелая пыльца и слабая ее летучесть у анемофильных растений способствуют перекрестному опылению при помощи ветра и являются важными приспособлениями к анемофилии.
Сыпучесть пыльцы характерна для анемофильных растений. Она связана с отсутствием или малым количеством клеящих веществ и жирных масел на поверхности зерен. Последние благодаря этому сухие, обособлены, не склеены друг с другом. Остатки небольшого количества масел на поверхности пыльцевых зерен многих анемофильных растений (например, у подорожников, злаков) свидетельствуют об их происхождении от энтомофильных предков, но у некоторых ветроопыляемых видов (береза и лещина) пыльца полностью лишена следов жирных масел.
Экзина пыльцевых зерен анемофильных растений тонкая, вследствие чего пыльца очень чувствительна к сухости и быстро утрачивает жизнеспособность. Особенно замечательны в этом отношении злаки, у которых жизнеспособность пыльцы ограничена немногими минутами (у цветущих после полудня) или часами (у цветущих рано утром). Рыльца же, напротив долговечны, сохраняются живыми в течение нескольких суток, особенно в случае протогинии (у подорожников, осок, маревых, некоторых злаков и т. д ).
Мощная листовая поверхность представляет собой как бы завесу, препятствующую распространению пыльцы. Поэтому у ряда древесных растений цветение происходит еще до развертывания листьев или соцветия располагаются на концах ветвей, а не в глубине кроны (лещина, береза и др.). У травянистых ветроопыляемых растений соцветия возвышаются над листовой массой (злаки, осоки, подорожники).
Охарактеризованная выше совокупность признаков структурной анемофилии оставляет в забвении динамику самого процесса. Между тем именно в этой динамике проявляются наиболее важные и удивительные приспособления к опылению ветром. Они не уступают в своей эффективности соответствующим приспособлениям у наиболее специализированных энтомофилов, но крайне своеобразны. Мы имеем здесь в виду суточную ритмику (суточную периодичность) цветения и опыления. Этот замечательный феномен составляет суть динамической анемофилии. Наиболее отчетливо она проявляется у злаков.
Суточная ритмика цветения и опыления злаков открыта более 100 лет назад, но оставалась малоизвестной и не была по достоинству оценена. Лишь в последние десятилетия выяснилось приспособительное значение динамической анемофилии, возможность использования ее в таксономических целях и ее роль в процессе видообразования у злаков.
Динамическая анемофилия проявляется в приуроченности цветения и опыления каждого вида злаков к определенному периоду суток. Совмещение рассеивания пыльцы у особей каждого вида злаков в пределах ограниченного времени, иногда очень краткого, весьма повышает шансы опыления - попадания на рыльце пыльцы, переносимой воздушными течениями. При таком одновременном и кратком цветении в течение суток у каждого вида создается в соответствующие сроки высокая насыщенность приземного слоя воздуха его пыльцой, несравненно более значительная, чем она была бы в том случае, когда то же самое количество ее рассеивалось непрерывно в течение многих часов или суток подряд. В этом и проявляется своеобразная экономность анемофилии. Суточная периодичность опыления представляет поэтому важнейшее приспособление злаков к опылению ветром.
По характеру суточной периодичности цветения и опыления можно выделить ночные, утренние, околополуденные, послеполуденные, вечерние, с двухразовым цветением (утро и вечер) и круглосуточно цветущие злаки. В средних широтах большинство злаков цветет утром и многие после полудня. В тропиках дело обстоит точно так же, но там, кроме того, некоторые группы злаков (сорговые) цветут преимущественно ночью.
Ведущим экологическим фактором цветения злаков является температура. Для каждого вида имеются свои кардинальные температурные точки цветения (максимальная, минимальная, оптимальная температура). Колебания температуры стимулируют начало цветения. Относительная влажность воздуха не регулирует суточных сроков цветения, но является важным фактором экологии опыления, поскольку пыльца злаков очень чувствительна к сухости воздуха. Для злаков, цветущих в светлое время суток, интенсивность света не играет роли, тогда как для ночных злаков свет (точнее, отсутствие света) играет решающую роль, например у видов сорго. Механическое сотрясение соцветий при ветре способствует цветению злаков, усиливая или ускоряя его. Рожь при ветре отцветает быстрее, чем при тихой погоде.
Динамическая анемофилия утренних, ночных и послеполуденных злаков протекает по-разному, в чем проявляется адаптация ее к резко различным условиям этих периодов суток.
Утро представляет наиболее обычное и благоприятное время для цветения злаков. Относительная влажность воздуха утром бывает наибольшей (свыше 70 - 80%), а температура воздуха пониженной (менее +16... +18 °С). Утренние (а также ночные) злаки как бы убегают от дневного зноя и засухи и уберегают пыльцу от их губительного действия. Цветение и опыление у них длятся около 4 - 6 ч, нарастают и затухают постепенно, характеризуются одновершинной кривой. Такое цветение можно назвать постепенным. Пыльца утренних злаков сохраняет жизнеспособность в течение нескольких часов, а при пасмурной и прохладной погоде даже нескольких суток. Опыление осуществляется при пониженной температуре. К числу утренних злаков принадлежит овсяница луговая, райграс высокий, ежа сборная и др.
Совсем иначе протекают цветение и опыление послеполуденных злаков. Они цветут и пылят при очень высокой температуре и низкой относительной влажности воздуха. Пыльца их в этих условиях недолговечна и утрачивает способность к прорастанию уже через 3 - 5 мин после рассеивания, если температура воздуха очень высокая. Казалось бы, что в этих ксеротермных условиях послеполуденного времени перекрестное опыление злаков с помощью ветра должно быть обречено на неудачу. Однако это не так. У послеполуденных злаков обнаружено замечательное приспособление к этим крайним условиям среды в форме взрывчатого и порционного цветения.
Взрывчатое цветение - это внезапное, одновременное, массовое и стремительное раскрывание цветков у локальной популяции вида, осуществляющееся во мгновение ока как бы по мановению волшебной палочки. Порционное цветение представляет особую форму взрывчатого. В этом случае в пределах суточного периода цветения бывает не один, а два-три последовательных взрыва (порции) цветения. Последнее протекает как бы отдельными толчками (порциями). Взрывы цветения длятся всего 3 - 5 мин, а межпорционные паузы, когда раскрывания цветков совсем не происходит, продолжаются 30 - 60 мин, а иногда растягиваются до 1,5 - 3 ч.
Одиночные и порционные взрывы цветения вызываются колебаниями температуры (0,5 - 2 ), обычно ступенчатыми, преимущественно в сторону ее понижения. Поэтому-то эти злаки и цветут после полудня, когда уже начинается понижение температуры после дневного максимума. Таков механизм экологической регуляции последовательных взрывов цветения, осуществляющихся у локальной популяции вида строго одновременно на значительной площади.
Взрывы цветения можно вызвать искусственно. Для этого нужно внести в более прохладное помещение лаборатории или комнаты срезанные соцветия злака или затенить плотной тканью участок популяции; вследствие понижения температуры в тени через 5 - 7 мин произойдет взрыв цветения. Опыление у таких послеполуденных злаков осуществляется в течение нескольких минут, так как пыльца их теряет жизнеспособность примерно через 3 - 5 мин (при температуре около и больше +30 °С и относительной влажности около и меньше 20%).
В качестве примеров послеполуденных злаков с порционным цветением можно назвать кострец безостый (Bromopsis inermis, табл. 1), кострец береговой (В. riparia), житняк гребенчатый (Agropyron cristatum), житняк сибирский (A. sibiricum), житняк пустынный (A. desertorum), пырей ползучий (Elytrigia repens), пырей волосоносный (Е. trichophora), пырей средний (Е. intermedia), овсец Шелля (Helictotrichon schellianum), ячмень короткоостый (Hordeum brevisubulatum) и др.
Взрывчатое и порционное цветение - своеобразное, тонкое, гибкое и важное приспособление злаков к опылению ветром в крайних ксеротермных условиях послеполуденного времени. Опыление осуществляется в краткие мгновения. Тончайшая экологическая регуляция одиночных и порционных взрывов цветения обеспечивает точную синхронизацию их, что повышает эффективность перекрестного опыления при помощи ветра.
Динамическая анемофилия, правда в менее дифференцированной форме, обнаружена и у других ветроопыляемых растений - у ситниковых, осоковых, подорожниковых, полыней, маревых.
Суточная периодичность опыления является если и не всеобщей, то все же широко распространенной у ветроопыляемых растений. Это важное приспособление к опылению ветром. Поэтому вполне правомерно говорить не только о структурной, но и о динамической анемофилии у растений.
Гидрофилия. У большинства водных растений цветки и соцветия возвышаются над поверхностью воды. Опыление у них осуществляется, как и у наземных сородичей, в воздушной среде при помощи насекомых (белая водяная лилия, частуха, стрелолист, турча, пузырчатка и др.) или ветра (рдесты - виды Polamogeton, уруть и др.).
Собственно гидрофилия, т. е. перенос пыльцы в воде или по водной поверхности, встречается у относительно немногих водных растений. Это особый случай абиотического опыления, обусловленный эдафическими условиями.
Опыление в толще воды свойственно взморникам (Zostera), роголистникам (Ceratophyllum), наядам (Najas), дзаннике ллиевым(Zannichelliaсеае), видам болотника (Callitriche) и немногим другим водным растениям. Имеется много общего в приспособлениях этих растений к опылению в воде. Цветки распускаются в воде, не выступая над ее поверхностью. В пыльниках нет утолщений в стенках клеток эндотеция, а нередко последний совсем утрачен. Пыльцевое зерно имеет лишь одну тонкую внутреннюю оболочку (интину), экзина же (наружная оболочка) крайне редуцирована, ее плотность близка к плотности воды. У многих видов пыльцевые зерна часто нитевидны (у Zostera 2000 X 8 мкм) или сферические пыльцевые зерна соединены посредством студневидной массы в цепочковидные образования (у Thalassia hemprichii), а иногда они сразу же после вскрытия пыльника прорастают в трубочку и переносятся водой уже в таком состоянии. Нитевидная форма и "червеобразное" состояние пыльцевых зерен повышают их плавучесть и шансы на опыление. Нитевидные пыльцевые зерна взморников оплетаются вокруг их рылец. Однако шансы на опыление у описываемых водных растений невелики. Вероятность повторного опыления совсем ничтожна. Эти гидрофильные растения имеют, подобно анемофилам, только один семязачаток в завязи. Размножение у них преимущественно вегетативное.
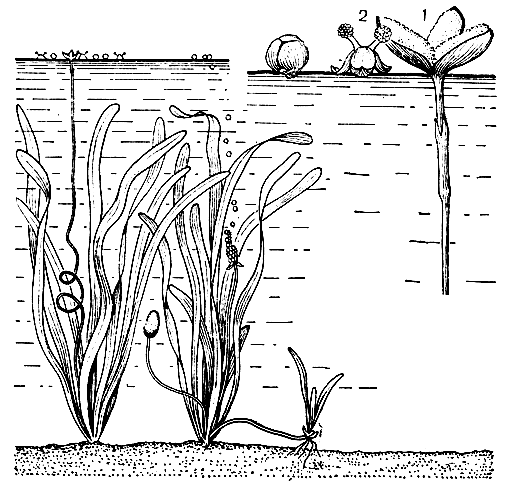
Рис. 40. Гидрофилия у валлиснерии (Vallisneria spiralis): слева - общий вид двудомной валлиснерии (женское и мужское растение); справа - опыление у валлиснерии, совершающееся на поверхности воды (1 - женский цветок; 2 - мужской цветок)
Иначе происходит опыление на поверхности воды. В этом случае цветки располагаются и распускаются на водной поверхности или непосредственно над ней. Шансов на опыление здесь больше,; чем под водой. В связи с этим в завязях находится не один, а несколько семязачатков. Опыление на поверхности воды свойственно валлиснерии, элодее, руппии и другим растениям..
Известнейший пример опыления на поверхности воды представляет валлиснерия - двудомное растение, произрастающее в водоемах, а также выращиваемое в аквариумах. Мелкие мужские цветки с двумя тычинками свободно плавают, как на лодочках, на откинутых книзу трех листочках околоцветника. Подгоняемые ветром, они передвигаются по водной поверхности, приближаются к плавающим женским цветкам. При этом может произойти случайное соприкосновение пыльцевой массы вскрывшихся пыльников мужских цветков с лопастями рыльца женских цветков (рис. 40). Сходно происходит опыление у элодеи. Возможно также опыление рылец пыльцой, плавающей на поверхности воды. При опылении в воде и на ее поверхности пыльца не погибает при погружении в воду.
Развитие гидрофилии происходило, видимо, двумя путями - непосредственно от энтомофильных предков или от них же, но через промежуточный этап анемофилии.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'