Биотическое опыление (А. Н. Пономарев, Е. И. Демьянова, И. В. Грушвицкий)
Энтомофилия. Насекомые сыграли выдающуюся роль не только в эволюции цветка, но и в его происхождении. Первичные покрытосеменные, очевидно, имели обоеполые энтомофильные цветки. Обоеполость цветка явилась предпосылкой для автогамии. Раздельнополость и способность к ветроопылению у покрытосеменных имеют вторичное происхождение.
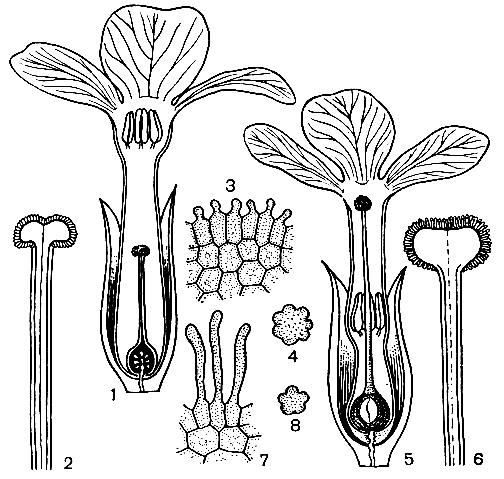
Рис. 34. Гетеростилия у примулы (Primula sp.): 1 - 4 - короткостолбчатая форма (1 - вид цветка в разрезе; 2 - короткий столбик с сосочковидной поверхностью рыльца при малом увеличении; 3 - то же при большом увеличении; 4 - пыльцевое зерно); 5 - 8 - длинностолбчатая форма (5 - вид цветка в разрезе; 6 - длинный столбик с сосочковидной поверхностью рыльца при малом увеличении; 7 - то же при большом увеличении; 8 - пыльцевое зерно)
Первичные цветковые растения, видимо, не были узкоспециализированы по способу опыления, и энтомофилия у них была довольно примитивной. Вполне возможно, что наряду с насекомоопылением некоторую роль играло и ветроопылениеи даже самоопыление. Примитивные цветковые растения еще не обладают способностью к выделению нектара, и приманкой для насекомых служит исключительно пыльца. Перелом в развитии энтомофилии был связан с появлением нектарников и с выделением нектара в цветках. Нектар явился дополнительным к пыльце пищевым ресурсом, привлекавшим насекомых к цветку. Появление нектара вызвало коренное изменение состава опылителей. Если первоначальными опылителями были, вероятно, только жуки, то теперь большую роль в опылении стали играть высшие перепончатокрылые, двукрылые и чешуекрылые. Этот перелом в развитии энтомофилии подготовил быстрое возвышение цветковых растений в середине мелового периода.
Хорошо известно, что насекомые посещают цветки для сбора пищи (пыльцы и нектара). Но иногда играют роль другие причины - поиски убежища, отложение яиц в цветке, сбор строительного материала и даже имитация удовлетворения полового инстинкта.
Пыльца является важнейшим источником пищи, которую насекомые находят в цветке. Она содержит белки, жиры, углеводы, зольные элементы, ферменты и витамины и является высокоценным питательным материалом. Поэтому насекомые собирают пыльцу не только энтомофильных, но и анемофильных растений (орешник, злаки, подорожники и т. д.), однако при этом они не принимают участия в их опылении. Следует, однако, заметить, что в качестве пищи пыльца энтомофильных растений представляет определенные преимущества - она более богата жирами и белками.
Цветки, привлекающие насекомых пыльцой, образуют огромное количество пыльцевых зерен (ветреница, мак, шиповник, зверобой и др.; табл. 1). Мак и пион по размеру пыльцевой продукции превосходят многие анемофильные растения.
У многих растений существует определенный дневной ритм в выделении пыльцы. Вскрытие пыльников и экспонирование пыльцы происходит либо в течение всего дня, либо в определенные сроки (утром, после полудня, в первую половину дня и т. д.).
У энтомофильных растений пыльца относительно долговечна, но вода губительна для пыльцы. Известны разнообразные средства защиты ее от дождя и росы. Из них следует упомянуть ориентацию колокольчатых, бокальчатых и чашевидных цветков открытым зевом венчика вниз. Такая ориентация у некоторых растений бывает постоянной и сохраняется на все время, пока пыльца нуждается в защите (черника, брусника, ландыш, спаржа, наперстянка, одноцветка, колокольчики); у других растений изгибание цветоножек и поникание цветков совершается периодически, перед дождем, в пасмурную погоду или ночью (адонис весенний, ветреницы, звездчатка злаковидная). Такую же функцию защиты пыльцы играет закрывание цветков перед дождем, на ночь и в пасмурную погоду у горечавок, некоторых колокольчиков, кувшинок.
Нектар является важнейшим источником пищи для всех посетителей цветков.Он выделяется особыми экскреторными железами - нектарниками.
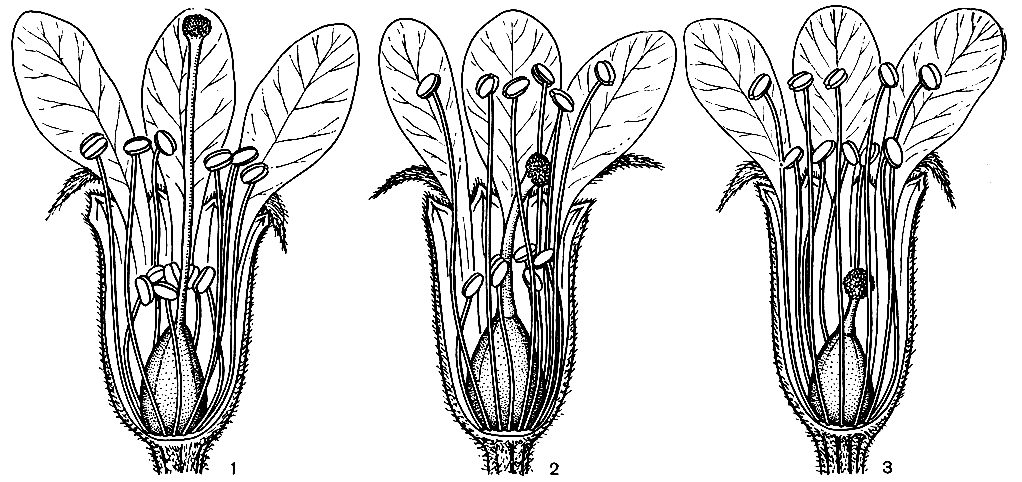
Рис. 35. Гетеростилия у дербенника иволистного (Lythrum salicaria): 1 - длинностолбчатая форма; 2 - среднестолбчатая форма; 3 - короткостолбчатая форма
Нектар представляет собой водный раствор Сахаров - фруктозы, глюкозы, сахарозы, мальтозы и др. Концентрация Сахаров в нектаре зависит от вида растения и условий среды. Она достигает 70% и более (у конского каштана 74,5%, у липы 72%, у душицы обыкновенной 60%, а у софоры японской даже 80%). Обычно содержание Сахаров в нектаре колеблется в пределах 30 - 50%, причем оно меняется в течение дня. Жидкий, а также высококонцентрированный нектар пчелы избегают, тогда как птицы-опылители в тропиках предпочитают водянистый нектар.
У различных цветковых растений, опыляемых насекомыми, птицами и летучими мышами, в состав нектара в меньших или в значительно меньших количествах могут входить также другие органические вещества, а именно: аминокислоты, белки, органические кислоты, витамины, алкалоиды, гликозиды и т. д.
В исследованиях последних лет, сообщенных недавно (в 1975 г.) в работе Герберта и Ирен Бейкеров, интересны попытки установить функциональное значение каждой из названных групп веществ нектара и определить их место в ходе сопряженной эволюции цветковых растений и их опылителей.
По составу Сахаров, которые, несомненно, являются основным для опылителей пищевым (углеводным) компонентом нектара, изученные в этом плане энтомофильные растения разделяются на две группы. Преобладающее содержание в нектаре сахарозы по сравнению с глюкозой и фруктозой свойственно цветкам более примитивного строения, обратное соотношение - более специализированным цветкам с глубоко спрятанным нектаром.
Из аминокислот, которые наряду с сахарами могут служить источником питания для опылителей, в этом случае как исходный материал для синтеза белков в нектаре чаще других встречаются аспарагиновая и глютаминовая кислоты, серии, глицин и аланин (у некоторых видов в нектаре присутствует до 13 различных аминокислот). Концентрация аминокислот в целом увеличивается от более примитивных цветков с открытым положением нектара к цветкам растений из подвинутых групп с нектаром, скрытым в глубине цветка, от древесных растений к травянистым, от обладателей актиноморфных цветков к видам с цветками зигоморфными, от раздельнолепестных к спайнолепестным.
Содержание белков в нектаре, достаточно высокое, чтобы служить существенным источником питания для опылителей, отмечено лишь у немногих растений (у вереска обыкновенного, гречихи). Чаще белки присутствуют в нектаре лишь в незначительных количествах и являются ферментами, т. е. веществами, определяющими ход тех или иных химических превращений.
Аскорбиновая кислота (витамин С) - витамин, наиболее часто присутствующий в нектарах, по-видимому, функционирует как антиоксидант, т. е. вещество, препятствующее окислению других органических соединений.
Алкалоиды и гликозиды, токсичные для тех или других групп насекомых, присутствуют в нектаре специализированных растений, приспособленных к опылению лишь определенным опылителем. Их наличие в составе нектара может рассматриваться как выработанное в ходе эволюции средство отпугивания нежелательных посетителей цветков.
Наконец, исследованиями Герберта и Ирен Бейкеров недавно показано, что в нектаре ряда растений (представителей 40 семейств, в том: числе жакаранды из бигнониевых, трихоцереуса из кактусовых, толокнянки обыкновенной, вахты трехлистной, лютика ползучего и многих других) в небольшом количестве содержатся жирные масла, которые в дополнение к сахарам могут служить источником энергии, необходимой для полета опылителей.
Выделение нектара представляет весьма динамичный процесс. Оно зависит от времени дня, возраста и стадии развития цветка, условий освещения, температуры, влажности воздуха и почвы и других экологических условий, причем проявляется эта зависимость у разных видов неоднозначно. Особый интерес представляет зависимость нектаровыделения от стадий развития цветка. Наиболее интенсивное выделение нектара связано, по-видимому, с фазой зрелого рыльца.
Суточная динамика выделения нектара неодинакова у разных видов. Согласно исследованиям Н. Н. Карташовой (1965), наиболее интенсивно нектар выделяется утром (липа, яснотка белая, душица обыкновенная), но у некоторых видов - днем (фацелия), под вечер (синюха голубая, дербенник иволистный) или имеется утренний и вечерний максимумы выделения нектара (клевер луговой, мышиный горошек, чина луговая и др.).
Нектаропродуктивность цветков весьма различна. В тропиках очень богаты нектаром цветки, опыляемые птицами и летучими мышами. У представителей нашей флоры огромное количество нектара обнаружено у княжика сибирского (Atragene sibirica) - до 90 мг и гвоздики пышной (Dianthus super bus) - до 44 мг в одном цветке.
Форма и расположение цветковых нектарников весьма разнообразны. Они возникли независимо в разных филетических линиях покрытосеменных. Нектарники встречаются на разных органах цветка - на чашелистиках, лепестках, цветоложе, тычиночных нитях, на наружной поверхности плодолистиков. Иногда они скрыты в шпорцах, представляющих модифицированные чашелистики (у настурции) или лепестки (у борца, водосбора и живокости). Хорошо известны нектарные ямки (кармашки) у основания лепестков лютиков. Функции нектарников выполняют расширенные основания тычиночных нитей (у гвоздичных). У спайнолепестных двудольных нектарники нередко имеют вид кольца, окаймляющего верхнюю завязь (у норичниковых, бурачниковых, губоцветных). В цветках с нижней завязью они имеют вид диска, окружающего столбик (у зонтичных, колокольчиковых, сложноцветных).
Недавно Стефан Фогель (1974) установил, что цветки некоторых растений тропиков выделяют в качестве приманки для насекомых-опылителей жирные масла. Нектарники в таких цветках отсутствуют. Жирные масла выделяются в них особыми желёзками - элайофорами, образованными подушечками железистых волосков или 1 - 2-слойным железистым эпителием. Опыление производится одиночными пчелами-антофоридами. Самки их собирают масло в чистом виде или в смеси с пыльцой и переносят его на волосках своего тела. Пыльца как добавочный продукт добывается из тех же "масляных" цветков или из особых пыльцевых цветков. Масло, обычно в смеси с пыльцой (в виде кашицы), пчелы откладывают в подземные выводковые ячейки как корм для личинок. Для собственного пропитания пчелы сосут нектар из нормальных нектароносных цветков. В настоящее время известно примерно 1260 видов с масловыделяющими цветками. Они принадлежат к 5 семействам (мальпигиевые, крамериевые, норичниковые, касатиковые, орхидные) и распространены в тропиках Нового Света по обе стороны экватора до 41° северной и южной широты.
В качестве пищи для насекомых могут служить также питательные волоски и ткани (у тропических орхидей, у коровяка).
Для привлечения опылителей служат, помимо пищи, также окраска и запах цветка, форма его имеет лишь небольшое значение.
Заметность цветков определяется их окраской и цветовым контрастом с окружающей зеленью. Окрашены бывают листочки простого околоцветника, чашелистики, лепестки, тычинки, верхушечные листья и прицветники. Окраска цветков представляет полную цветовую гамму от длинноволновых лучей красного цвета до коротковолновых - ультрафиолетовых лучей, не воспринимаемых человеческим глазом, но воспринимаемых пчелами. Она обусловлена различными пигментами - флавоноидами (антоцианами и флавонами), каротиноидами, антофеином, бетацианинами.
Антоцианы растворены в клеточном соке, но иногда могут выпадать в клетках в форме кристаллов. Они очень распространены у растений. Большинство окрасок цветков можно приписать им. В зависимости от реакции клеточного сока антоцианы вызывают красную окраску (в кислом растворе), синюю (в слабо щелочном растворе), фиолетовую (в нейтральном растворе). Желтая окраска цветков зависит от присутствия флавонов, их производных и каротиноидов. Последние определяют, например, желтую окраску цветков лютиков, купальницы, калужницы, чистотела, донника лекарственного, зверобоя продырявленного, подсолнечника и других растений. Чистые кристаллы каротина обусловливают красную окраску привенчика нарцисса. Производные флавонов (лютеолин и др.) вызывают желтую окраску у желтых анютиных глазок, наперстянок и дрока красильного. Коричневый пигмент антофеин вызывает черные пятна на лепестках конских бобов, бурую окраску нектарников у живокости и венчиков некоторых тропических орхидей. Белая окраска цветков не связана с наличием какого-либо особого пигмента, а обусловлена очень рыхлым строением лепестков вследствие наличия в них множества воздухоносных полостей (межклетников).
Кроме огромного разнообразия окрасок, доступных восприятию человека, цветки имеют еще одну "окраску", которая не воспринимается нами, но хорошо различается пчелами. Речь идет об ультрафиолетовом излучении цветков. Сильное ультрафиолетовое отражение обнаружено у цветков многих растений - дербенника иволистного, ослинника двулетнего, переступня двудомного и др. Очень невзрачные для нас цветки переступня хорошо различаются и интенсивно посещаются пчелами благодаря отражению ультрафиолетовых лучей. Отдельные участки венчика в разной степени отражают ультрафиолетовые лучи, подобно тому как и лепестки в разных частях имеют неодинаковую видимую окраску. Действительно, очень часто цветки имеют пеструю окраску, тот или иной узор или рисунок. На лепестках можно видеть точки, штрихи, линии, пятна разной величины и окраски и т. д. Такие знаки или метки рассматривались еще в конце XVIII в. одним из предшественников Ч. Дарвина - Христианом Конрадом Шпренгелем как указатели нектара у нектаровыделяющих цветков или как указатели пыльцы у безнектарных пыльцевых цветков. Примеры этого можно найти почти в любом цветке самой различной окраски, от интенсивно красной до ультрафиолетовой. Эти видимые (цветные) или невидимые (ультрафиолетовые) метки, образующие тот или иной рисунок на цветке, указывают кратчайший путь к нектару. По меткому выражению Чарлза Даддингтона, они являются как бы вывесками ресторана, в который приглашаются опылители. Таковы, например, красный привенчик у нарцисса настоящего, желтое кольцо в голубом цветке незабудки, темно-желтые пятна у основания лепестков примул, фиолетовые штрихи на лепестках герани, темно-желтое пятно на нижней губе льнянки, крапинки (точки) на внутренней стороне нижней губы наперстянки пурпуровой и т. д. Подобные же, но невидимые для нашего глаза, однако видимые насекомыми (пчелиными) ультрафиолетовые метки имеются у очень многих цветков, например у герани луговой, переступня двудомного, турнепса и др. Выяснилось также, что эти указатели нектара различаются не только по цвету, но также по запаху, о чем будет сказано несколько ниже.
У некоторых цветков наблюдается изменение окраски в течение жизни. Наиболее известным примером в этом отношении является медуница (Pulmonaria obscura, P. mollissima), у которой цветки при распускании бывают розово-пурпуровыми, а потом становятся синими. У чины весенней (Orobus vermis) цветки сначала красные, а затем зеленовато-синие. Иногда меняется окраска какой-либо части цветка, как, например, у конского каштана, у которого желтое пятно на лепестках становится сначала оранжевым, а потом карминно-красным, причем нектар выделяется только в начальной (желтой) фазе.
Разнообразная окраска цветков, безусловно, имеет приспособительное значение, привлекая насекомых-опылителей. Насекомые обладают способностью различать цвета, но их цветовое зрение, особенно у пчел, отличается рядом особенностей по сравнению с цветовым зрением человека. Главная особенность цветового зрения пчел заключается в их восприимчивости к коротковолновым ультрафиолетовым лучам.
Белые цветки довольно слабо отражают ультрафиолетовые лучи (до 3%) и воспринимаются пчелами как беловато- или сине-зеленые, например у нарцисса, земляники, груши, ветреницы лесной. Белые соцветия маргаритки кажутся пчелам голубовато-зелеными звездочками. Желтые и оранжевые цветки, кажущиеся нам однотонно окрашенными, по-разному отражают ультрафиолетовые лучи, а поэтому окраска их для пчел изменяется от пчелиного желтого до пчелиного пурпурового. Эти цветки отражают ультрафиолет в разной степени - от 2 - 10 до 20 - 40%. Таковы подмаренник настоящий (Galium verum), дрок красильный (Genista tinctoria), зверобой продырявленный (Hypericum perforatum), лапчатка гусиная (Potentilla anserina) и адонис весенний (Adonis vernalis). Пчелы отличаются слепотой к красной окраске, поэтому окраску цветков тюльпана, фасоли и мака они воспринимают как "ультрафиолетовую", поскольку такие цветки отражают ультрафиолетовые лучи.
Дневные бабочки обладают цветовым зрением. Среди них можно наметить две группы. Одни из них - белянки (Pieridae) и парусники (Papilionidae) - летят в поисках пищи на красное, желтое и сине-фиолетовое, тогда как другие - нимфалиды (Nymphalidae) и бархатницы (Satyridae) - оказывают предпочтение желтому и голубому. Среди цветков, опыляемых дневными бабочками с длинным хоботком, надо упомянуть прежде всего пурпурно-красные цветки гвоздик. Дневной бражник (Macroglossum stellatarum из семейства Sphingidae) летает днем при полном солнечном освещении и в вечернее время. Он не воспринимает красную часть спектра, но различает цвета голубой, синий и желтый, в дневные часы посещает также "белые цветки мыльнянки (Saponaria officinalis).
Посещение цветков дневным бражником направляется только оптически, без участия запаха. Специальными опытами, проведенными Фрицем Кноллем, было доказано, что желтые пятна на нижней губе цветка льнянки хорошо различаются чисто оптически. Воздействие запаха было исключено, поскольку цветки прикрывались стеклом. Желтые пятна выступают в роли указателей нектара: они прежде всего (в опытах через стекло) ощупывались хоботком.
Цветки, опыляемые ночными бабочками, имеют обычно светлую окраску - белую, бледно-желтую и светло-пурпуровую. Эта светлая окраска, контрастирующая с общим темным фолом, делает цветки более заметными. Светлая окраска цветков сама по себе, без содействия запаха, может управлять полетом таких крупных бабочек, как бражник вьюнковый (Protoparce convolvuli). Эта ночная бабочка обладает хорошим цветовым зрением. Она отличает не только светлую, но и темно-фиолетовую и темно-пурпуровую окраску от темно-серой или черной даже при такой интенсивности освещения, лри которой человеческий глаз их уже не может воспринимать.
Цветки, опыляемые ночными бабочками, источают к вечеру или ночью сильный аромат. Это дало повод думать, что запах является для ночных бабочек единственным средством ориентации. Привлекающее действие запаха весьма вероятно, однако этот вопрос еще подлежит исследованию. По-видимому, ночные бабочки ориентируются как оптически, так и по запаху.
Двукрылые посещают цветки ради пыльцы и нектара, используемых ими на месте для собственного пропитания. В качестве посетителей и опылителей наиболее важны цветочные мухи (Syrphidae) и жужжала (Bombylius). Посещаемые ими цветки имеют яркую желтую, синюю и фиолетовую окраску, причем и белая окраска не исключена. У жужжал резко выражено цветовое зрение, и они привлекаются к цветку издали оптически.
В географическом распределении окраски цветков обнаруживаются некоторые закономерности. Обращает внимание обилие в тропиках ярких оранжевых и красных цветков, которые почти отсутствуют в средних и высоких широтах. Это связано, видимо, с тем, что в тропиках большую роль в опылении играют птицы, которые различают красный цвет. В высокогорьях и приполярных областях растения с белыми цветками составляют до 38 - 40%, тогда как в Средней Европе таких растений около 23%, я в пустынях и сухих областях их еще меньше. Желтые цветки одинаково часты как в пустынях, так и в приполярных странах (32 и 33%). Что касается растений с синими цветками, то они довольно редки в пустынях (в сухих областях) (только 4%) и в приполярных областях (5%). Связь окраски цветков с комплексом факторов отдельных зон еще очень неясна. По-видимому, не следует видеть в окраске цветка только приспособление к определенным опылителям. Высказываются и другие мнения. Допускается возможность, что пигменты цветка представляют приспособление к поглощению солнечного света, в связи с чем окраска цветков может зависеть от климатических условий.

Таблица 1. Приспособления цветков к опылению: 1 - протандрия у кипрея узколистного, или иванчая (Chamaenerium angustifolium); 2 - пыльценосный цветок у зверобоя продырявленного (Hypericum pertoratum); 3 - взрывчатое цветение у костреца безостого (Bromopsis inermis); 4 - гравитационная автогамия у одноцветки крупноцветковой (Moneses uniflora); 5 - съедобные волоски на тычинках у коровяка медвежьего уха (Verbascum tapsus); 6 - соцветие с наружными бесполыми цветками у калины обыкновенной (Viburnum opulus)
Цветки привлекают насекомых-опылителей не только окраской, но часто также и запахом. Орнитофильные цветки лишены аромата, а у цветков, опыляемых летучими мышами, ощущается кисловатый и затхлый запах. Аромат же энтомофильных цветков очень разнообразен. Различают примерно 500 разных цветочных запахов.
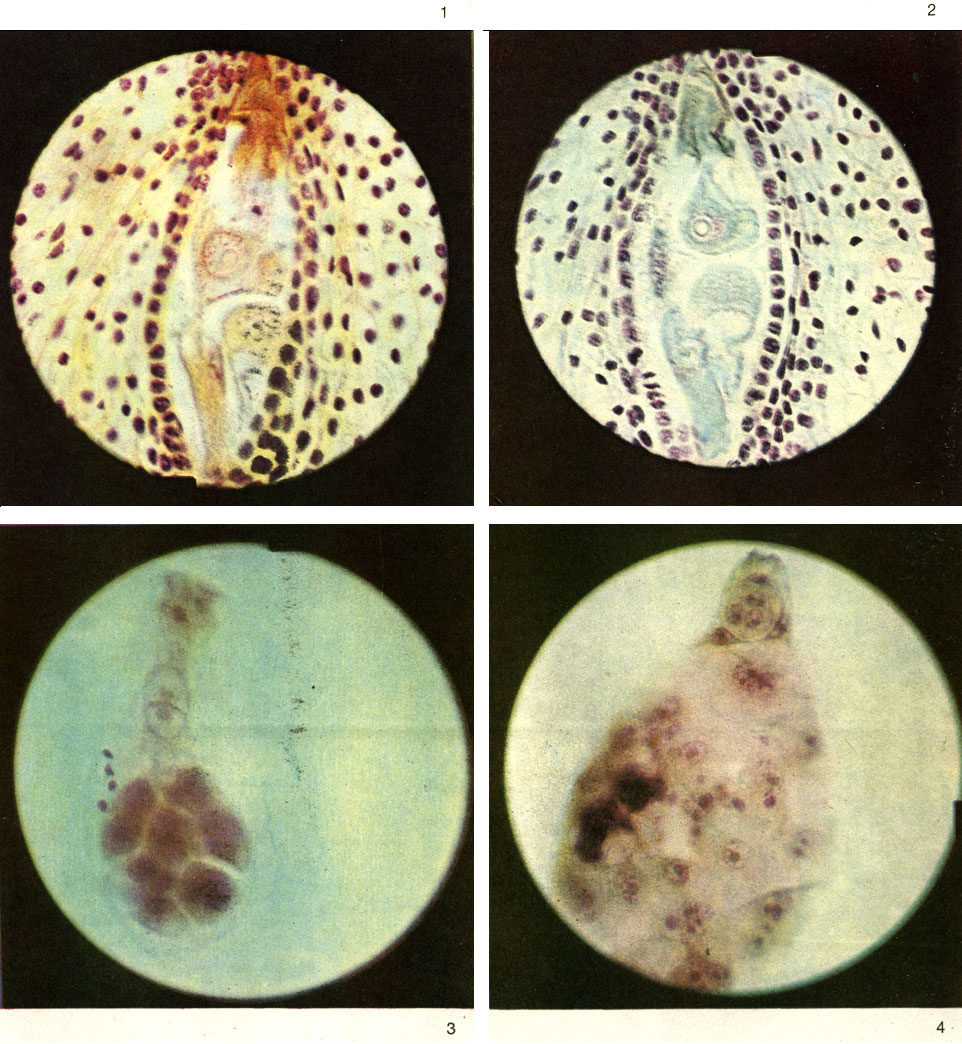
Таблица 2. Оплодотворение у цветковых растений: 1 - продольный срез семязачатка с зародышевым мешком во время оплодотворения у рудбекии рассеченной (Rudbeckia laciniata); синергиды (желтого цвета) разрушаются; недалеко от ядра яйцеклетки красный спермий, в халазальной части 2 антиподы (3-я на другом, срезе), одна из них яйцеклеткоподобная; 2 - продольный срез семязачатка с зародышевым мешком во время оплодотворения у рудбекии рассеченной; в правую синергиду проникла пыльцевая трубка, ее ядро разрушено, зигота еще не приступила к делению, центральное ядро готовится к делению; в халазальной части 2 антиподы, одна из них яйцеклеткоподобная (3-я антипода на другом срезе); 3 - выделенный из семязачатка зародышевый мешок дюпонции Фишера (Dupontia fischeri); хорошо видна зигота, 2 контактирующих ядра и комплекс крупных антипод; слева - мелкие соматические клетки; 4 - выделенный зародышевый мешок с четырехклеточным зародышем, с ядрами эндосперма и дегенерирующими антиподами у фиппсии холодолюбивой (Phippsia algida)
Сравнительно редко цветки и соцветия растений испускают отвратительный трупный запах гниющего мяса или рыбы, разлагающейся мочи и навозной жижи. Такие запахи свойственны початкам аронников (Arum), цветкам южноафриканских стапелий (Stapelia), тропических паразитных раффлезий (Rafflesia), многих видов кирказонов (Aristolochia). Эти цветки опыляются мухами, привлекаемыми запахом падали. К тому же и окраска цветков у раффлезий имитирует разлагающееся мясо.

Таблица 3. Дегенериевые и магнолиевые: 1 - цветок дегенерии фиджийской (Degeneria vitiensis); 2 - бутон магнолии Делавэ (Magnolia delavayi); 3 - цветок магнолии Делавэ
Более распространены у цветков аминоидные запахи, обусловленные наличием различных, аминов (метиламин, диметиламин, триметиламин и т. д.). Такие цветки посещаются настоящими мухами и жуками. Аминоидные запахи свойственны цветкам боярышника, рябины, калины, кизила, барбариса и т. д.

Таблица 4. Ветвь с листьями и цветками эвпоматии Беннетта (Eupomatia bennettii)
Чаще же всего запахи цветков обусловлены эфирными маслами. Последние представляют смесь веществ, различающихся в химическом и физическом отношении. Наиболее важными из этих веществ, входящих в состав эфирных масел и определяющих запахи цветков, являются различные спирты, альдегиды, кетоны, фенолы, простые и сложные эфиры, терпены и т. д. Запах связан с присутствием в цветке какого-либо одного химически чистого вещества, но большей частью определяется их сложной смесью. Так, запах розы вызывается первичными спиртами - гераниолом, неролом, цитронеллоолом, запах ландыша - третичным спиртом линалоолом, запах гиацинта - коричным спиртом, запах фиалки - кетоном пармоном, запах жасмина - сложной смесью веществ - бензилацетатом, линалоолом, индолом, жасмоном и т. д. Ароматические вещества действуют как газы. Они распространяются токами воздуха или благодаря диффузии. Источником запаха являются обычно лепестки, но источают пахучие вещества также тычинки, стаминодии и нектарники. Иногда имеются специальные многоклеточные железки, выделяющие эфирные масла со специфическим запахом. У ясенца (Dictamnus) пурпурные желёзки на тычинках выделяют эфирное масло с лимонным запахом.

Таблица 5. Магнолиевые Батумского ботанического сада: 1 - магнолия обратноовальная (Magnolia obovata); 2 - магнолия вирджинская (М. virginiana); 3 - манглиетия тонковатая (Manglietia tenuipes); 4 - тюльпанное дерево (Liriodendron tulipifera)
Упоминавшиеся выше метки или знаки на лепестках в виде пятен, точек, штрихов разной окраски, образующие тот или иной рисунок,; воспринимаются насекомыми не только оптически, но и по запаху. Как было сравнительно недавно (1954) установлено немецким ученым Терезой Леке, они отличаются от основной части лепестков не только по окраске, но и по качеству и интенсивности запаха. Так, у нарцисса настоящего красный привенчик источает более интенсивный и своеобразный запах, чем прочие части цветка, причем запах этот усиливается к основанию привенчика и листочков околоцветника. Медоносные пчелы отличают запах привенчика от запаха околоцветника. Подобное же явление обнаружено у видов первоцвета, настурции, ириса германского, живокости высокой, фиалки трехцветной. У нашего обычного сорного растения пикульника красивого светло-желтые цветки имеют на нижней губе венчика темно-фиолетовое пятно, которое обнаруживает качественно иной, но более слабый запах, чем остальной венчик. У конского каштана (Aesculus hippocastanum), подснежника белоснежного (Galanthus nivalis) цветные метки на лепестках пахнут интенсивнее, чем другие части венчика. У конского каштана одновременно с изменением окраски цветного пятна на лепестках с желтой на красную изменяется и запах. Однако в ряде случаев различие в качестве и интенсивности запаха отдельных частей венчика не сопровождается соответствующими особенностями в их окраске. У вьюнка полевого белые полоски, не заметные на белом венчике, пахнут сильнее и иначе, чем последний. У нектарников запах иной и более интенсивный, чем у других частей цветка.

Таблица 6. Магнолиевые. Магнолии Батумского ботанического сада: 1 - магнолия трехлепестная (Magnolia tripetala); 2 - магнолия китайская (М. sinensis); 3 - магнолия Уотсона (М. watsonii). Магнолии США: 4 - магнолия крупнолистная (М. macrophylla)
Запах пыльцы отличается от запаха цветка. Медоносные пчелы их хорошо различают. У протандричных цветков горца змеиного, или раковых шеек (Polygonum bistorta), запах появляется при созревании и вскрытии пыльников в тычиночную фазу и исчезает после ее окончания. Довольно неприятные запахи цветков калины и дерена белого обязаны своим происхождением аминам, содержащимся в пыльце. У омелы мужские цветки пахнут сильнее, чем женские. Источником запаха является выстилающий слой (тапетум) пыльников, реже стаминодии. У ветроопыляемых и орнитофильных цветков запах пыльцы очень слабый. По мнению Отто Порша, запах пыльцы является наиболее древним запахом цветков.

Таблица 7. Магнолиевые. Магнолиевые Батумского ботанического сада: 1 - магнолия лилиецветковая (Magnolia liliflora); 2 - микелия буроватая (Michelia fuscata). Магнолиевые в оранжерее Ботанического института АН СССР в Ленинграде: 3 - магнолия кобус (Magnolia kobus); 4 - микелия фиго (Michelia figo)
Прежде приписывали большое значение медовому запаху в привлечении насекомых к цветку. Однако чистый нектар лишен собственного запаха, но может поглощать запах других частей цветка. Насекомые до посещения цветка не могут установить, имеется в них нектар или нет. Об этом свидетельствуют безуспешные попытки их высасывать нектар из пыльцевых цветков, лишенных нектара (мак, зверобой, люпин).

Таблица 8. Магнолия крупноцветковая (Magnolia grandiflora), США: 1 - цветок; 2 - тычинки и плодолистики (околоцветник удален); 3 - плодолистики (околоцветник и тычинки опали)
В испускании запаха, как и в выделении нектара, наблюдается известная суточная периодичность. Наиболее известным примером в этом отношении являются мыльнянка, смолевки, ослинник, вечерница, петуния, любка двулистная, лилия кудреватая и т. д. У многих из этих растений цветки распускаются к вечеру и открыты ночью - они опыляются ночными бабочками. Наоборот, цветки, которые распускаются днем и опыляются дневными бабочками, пчелами и шмелями, наиболее ароматны в дневное время (клевер, люцерна, яблоня и др.).
Какова сравнительная роль окраски и запаха в привлечении насекомых к цветкам?
Эти факторы действуют совместно, но относительное значение их неодинаково на дальнем и близком расстоянии. Дальнее действие вызывает прилет насекомого-опылителя к цветку,, а действие с близкого расстояния влечет за собой посещение цветка. Прилет к цветку необязательно сопровождается его посещением. Иногда насекомое, подлетев к цветку, не посещает его, а как бы "отворачивается" от него.
Окраска цветков является в большинстве случаев фактором дальнего действия. Насекомые-опылители в прилете к цветку ориентируются чисто оптически. Таковы пчелиные, дневной бражник, бражник вьюнковый, бражник олеандровый (Deilephila nerii), дневные бабочки, жужжала и т. д. Такое оптическое дальнее действие основано преимущественно на цветовом контрасте цветка с окружающей обстановкой.
Запах как фактор дальнего действия, за некоторыми исключениями, о которых будет сказано позднее, не играет существенной роли в прилете насекомых к цветку. Наоборот, он выступает как решающий химический фактор близкого действия, определяющий посещение цветков. Без воздействия запаха на близком расстоянии, в сущности, не происходит никакого или почти никакого посещения цветков. Опыты показали, что запах цветка на близком расстоянии действует сильнее, чем окраска его, как бы перебивая или заглушая действие последней. Таким образом, прилет насекомых к цветку и последующее посещение его определяются взаимодействием (игрой) оптического (окраска) и химического (запах) факторов на дальнем и близком расстоянии.
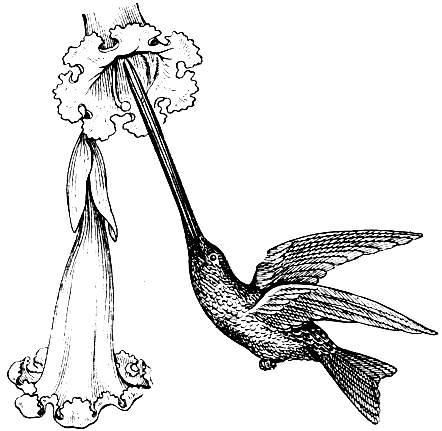
Рис. 36. Орнитофилия. Перекрестное опыление с помощью колибри
Однако в отдельных случаях запах является важнейшим фактором дальнего действия, вызывая прилет насекомых к цветку. К числу насекомых, ориентирующихся по запаху с дальнего расстояния, принадлежат прежде всего такие, которые разыскивают пищу на экскрементах, падали и навозе и откладывают на них яйца. Сюда относятся многие двукрылые (мухи). Они ориентируются в прилете и посещении цветков трупным запахом (у аронников, раффлезий, стапелий). Бабочки также могут лететь к цветку исключительно по причине воздействия запаха. Так, например, ориентируется бабочка ленточник Камиллы (Limenitis Camilla) при посещении цветков крушины и липы. Ночные бабочки также в основном ориентируются по запаху.
Несколько слов следует сказать о привлечении с дальнего расстояния насекомых-опылителей половыми аттрактантами цветков. При этом цветки издают аромат, напоминающий запах самок, привлекающий самцов. Самцы одиночных пчел (Eucera, Andrena, Corytes и др.) посещают цветки разных видов рода офрис (Ophrys) из семейства орхидных и производят на них движения совокупления, как бы пытаясь спариться с "самкой". Запах и форма цветков офрисов, имитируя самок-опылителей и действуя на органы чувств самцов, побуждают их к перелету с цветка на цветок для удовлетворения полового инстинкта. При этом происходит перенос поллиниев этих орхидей и перекрестное опыление. Посещение и опыление цветков офрисов самцами прекращается, как только обнаруживаются самки соответствующего вида, появляющиеся позднее самцов.
По мнению шведского специалиста в области биологии опыления Бертила Кулленберга, цветки офриса источают запахи, имитирующие запахи самок. Эти половые вещества выделены в чистом виде. Будучи помещены под сеткой (в трубочке), они привлекали в опытах Кулленберга самцов андрены с дальнего расстояния.
Подобное же явление сексуальной обусловленности опыления половыми аттрактантами обнаружено у ряда других орхидных (Oncidium, Brassia, Calochilus, Cryptostylis).
Другие виды биотического опыления. Мы уже видели, какую огромную роль в опылении цветковых растений играет многообразный мир насекомых. Другие наземные беспозвоночные подобных биотических связей не выработали. Ранее допускали возможность участия моллюсков (улиток) в опылении некоторых растений. Однако сейчас даже наиболее вероятные случаи так называемой малакофилии (опыления моллюсками), например, у белокрыльника считаются сомнительными. Напротив, из числа позвоночных в опылении растений, несомненно, широкое участие принимают в первую очередь птицы, затем летучие мыши и, очевидно, некоторые нелетающие млекопитающие. Много интересных сведений об этих экзотических способах опыления приводят английские ученые Майкл Проктор и Питер Ио в своей монографии "Опыление цветков" (1973).
Опыление птицами (орнитофилия) особенно характерно для тропиков, но представлено также во внетропических областях южного полушария - в Австралии, Южной Африке и Южной Америке. В северном полушарии только в Северной Америке посещение цветков птицами (несколько видов колибри) установлено в далеко отстоящих от тропиков районах, до Аляски включительно.
Орнитофильными являются такие известные растения, как виды эвкалиптов (Eucalyptus), канн (Саппа), акаций (Acacia), фуксий (Fuchsia), алоэ (Aloe), некоторые виды кактусов и многие другие.
Цветки, опыляемые птицами (рис. 36), характеризуются отсутствием запаха, что связано со слабо развитым у птиц обонянием. Зато птицы весьма чувствительны к цвету и, в отличие от насекомых, хорошо различают красную окраску. Птицеопыляемые цветки отличаются ярким привлекающим цветом околоцветника или прицветных листьев, чаще всего красным или оранжевым, реже с синеватым или фиолетовым оттенком. Нередко имеет место контрастирующая пестрота; такая "попугайная" окраска (сочетание зеленой, желтой и алой расцветки) характерна, например, для многих птицеопыляемых бромелиевых.
Нектар у орнитофильных растений водянистый (в нем всего около 5% сахара) и иногда слизистый. При этом он продуцируется в больших количествах. Так, виды рода банксия (Banksia, семейство протейных) образуют столь много нектара, что местные жители австралийцы используют его в пищу.
В разных областях опылителями растений являются птицы, относящиеся к разным семействам, - колибри (Trochilidae) и цветочницы (Coerebidae) в Америке, нектарницы (Nectariniidae), медососы (Meliphagidae) и белоглазки (Zosteropidae) в тропиках Старого Света, цветососовые (Dicaeidae) и попугаи лори (Loriinae) в Австралии и тропической Азии, гавайские цветочницы (Drepanididae) на Гавайских островах.
В строении тела и в повадках всех птиц-опылителей отражена многовековая приспособленность к опыляемым растениям. Мелкие птицы обычно порхают в воздухе (рис. 36), посасывая нектар из цветков, не садясь на них. Благодаря высокой скорости и точности полета и приспособленности многих представителей к посещению всего одного вида растения такие птицы являются весьма эффективными опылителями; некоторые из них могут за день посетить многие тысячи цветков. Другие виды птиц, обычно более крупные, садятся на землю, добывая нектар из цветков и соцветий, расположенных у самой почвы.
Взаимная приспособленность птиц и орнитофильных растений развивалась независимо и параллельно в разных их группах. Так, среди цветковых орнитофильные виды относятся как к классу двудольных (лютиковые, гамамелисовые, миртовые, бобовые, протейные, вересковые, жимолостные, мареновые, геснериевые, норичниковые, губоцветные и др.), так и к классу однодольных (лилейные, амариллисовые, бромелиевые, банановые).
То же можно сказать и об опылении летучими мышами. Хироптерофилия, как называют эту систему опыления, распространена в тропиках, особенно в тропиках Азии и Америки, в меньшей степени Африки, главным образом Западной Африки. Хироптерофильные растения - большей частью деревья (притом нередко высокие) и лианы, реже это кустарники и даже травы. Они относятся ко многим семействам из разных подклассов двудольных, более всего из подклассов Dilleniidae (баобабовые, маркгравиевые, сапотовые и др.), Rosidae (миртовые, бобовые, соннератиевые) и Asteridae (бигнониевые, геснериевые, синюховые), а также к однодольным (виды банана, агавы). "Услугами" летучих мышей пользуются, в частности, цветки таких широко известных представителей семейства баобабовых, как обитатель африканских саванн баобаб (Adansonia digitata), южноамериканские "шерстяное дерево" (Ceiba pentandra) и бальса (Ochroma lagopus), наконец, азиатский дуриан (Durio zibethinus).
Летучие мыши, в отличие от деятельных днем птиц, посещают и опыляют цветки в темное время суток - в сумерках и ночью. С этим связаны такие особенности цветков хироптерофильных растений, как тусклый зеленовато-желтый, коричневый или фиолетовый цвет и раскрывание цветков вечером, так что их пыльца и нектар становятся доступными для посетителей только с наступлением темноты. Известный голландский ботаник ван дер Пэйл характеризует и другие особенности цветков, опыляемых летучими мышами. Цветки или соцветия здесь обычно крупные, с прочным околоцветником и прочными "посадочными площадками" для опылителей в виде толстых цветоножек, или цветоносов, или, наконец, прилегающих безлистных участков ветвей. В цветках образуется много слизистого нектара и пыльцы. Наконец, они имеют неприятный, затхлый запах, имитирующий запах секреции желез самих летучих мышей, позволяющий последним, по предположению ван дер Пэйла, ориентироваться в стае. При этом неприятный для человека запах цветков привлекает летучих мышей.
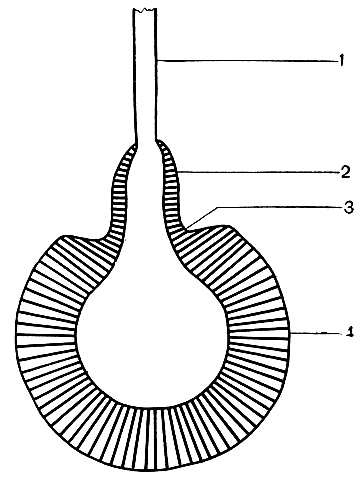
Рис. 37. Схематическое изображение строения соцветия паркий Клаппертона (Parkia clappertoniana): 1 - цветонос; 2 - стерильные цветки, выделяющие нектар; 3 - кольцевой желобок; 4 - плодущие цветки
В качестве примера хироптерофильного растения можно привести паркию (Parkia clappertoniana) из подсемейства мимозовых семейства бобовых. У этого дерева саванн Западной Африки соцветия представляют собой плотные шаровидные скопления из двух тысяч и более цветков, расположенных на длинных (более 30 см), прочных и гибких цветоносах, обращенных книзу. Как и у других растений, опыляемых летучими мышами (например, у баобаба), соцветия висят далеко от листвы и бывают хорошо заметны для опылителей. Цветки испускают слабый фруктовый запах. Соцветие состоит из фертильных (плодущих) протандричных и мужских (стерильных) цветков. Главная функция стерильных цветков - выделение нектара, который, стекая вниз по соцветию, скапливается в небольшом углублении - кольцевом желобке (рис. 37). Последний располагается на границе между стерильными и плодущими цветками. Каждое соцветие паркий за одну ночь выделяет около 5 мл нектара.
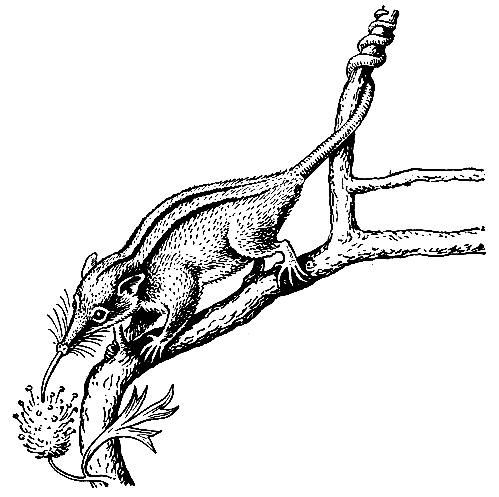
Рис. 38. Опыление с помощью нелетающих млекопитающих. Хоботноголовый кускус (Tarsipes spencerae) на ветви опыляемого растения.
Известны также случаи участия некоторых нелетающих млекопитающих в опылении цветков ряда тропических растений. Недавно, в 1978 г., накопленные в этой области данные обстоятельно проанализированы американскими учеными - зоологом Робертом Зюсманом и ботаником Питером Рейвеном - в интересной обзорной статье "Опыление лемурами и сумчатыми: архаичная коэволюционная система". В качестве постоянных посетителей и вероятных опылителей ряда древесных растений из миртовых, протейных, сарколеновых и некоторых других семейств авторы приводят представителей таких групп, как мелкие сумчатые в Австралии, грызуны в Австралии и Южной Африке, лемуры на Мадагаскаре.
Хорошим примером такой связи является австралийский хоботноголовый кускус (Tarsipes spencerae), описание и изображение (рис. 38) которого приводятся в VI томе издания "Жизнь животных" (М., 1968). Это мелкое ночное животное из сумчатых замечательно приспособлено к древесному образу жизни и питанию пыльцой и нектаром цветков. Отмечается удивительная аналогия образа жизни этого кускуса и маленьких австралийских же опыляющих растения птиц-медососов. Язык этого проворного зверька, легко перебирающегося с ветки на ветку с помощью цепкого хвоста, представляет собой своеобразную кисточку для сбора пыльцы, а его вытянутая в виде хоботка мордочка приспособлена к втягиванию нектара. Интересно, что, как и птицы-медососы, хоботноголовый кускус совершает миграции, связанные с ходом зацветания древесных растений, на которых он проводит жизнь и получает пищу.
Что касается самих растений, в опылении которых принимают участие нелетающие млекопитающие, то им, по данным указанных выше авторов, свойственны крупные и прочные цветки (или соцветия), относительно небольшое число цветков (или соцветий) на одном растении,, сильный аромат и выделение обильного нектара.
Зюсман и Рейвен обосновывают мысль о том, что подобные системы (нелетающие млекопитающие - опыляемые растения) были более широко распространены в третичное время и в наши дни сохранились лишь как реликтовые.
На дому почистить домашний диван в Москве. Качественная чистка диванов
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'