Порядок гелидиевые (Gelidiales)
Гелидиевые - небольшая, хорошо очерченная группа красных водорослей. В ней выделяют всего одно семейство и 8 родов. Ауксиллярные клетки, как и у нема лиевых, отсутствуют, но еще до оплодотворения рядом с карпогонной ветвью образуются многочисленные мелкоклеточные нити. Это особая питающая ткань, в которой развиваются нити гонимобласта.
Все гелидиевые - исключительно морские обитатели, ограниченные в своем распространении тропическими морями и теплыми морями умеренных широт. Во многих странах они известны не только ученым. Среди багрянок, дающих желирующие вещества, гелидиевые занимают одно из заметных мест, являясь сырьем для добычи самого ценного и высококачественного фикоколлоида - агара. В нашей стране гелидиевые в качестве источника агара не используют, поскольку они встречаются только в южных морях СССР и запасы их здесь невелики.
Центральный и наиболее обширный (около 40 видов) род этой небольшой единообразной группы водорослей - гелидиум. Познакомившись с ним, можно составить представление обо всем порядке в целом. Виды гелидиума - некрупные хрящеватые растения длиной от нескольких сантиметров до нескольких десятков сантиметров. Первоначально цилиндрическое слоевище гелидиума имеет заметно выраженную в эволюции тенденцию уплощаться, поэтому у многих видов конечные веточки или даже целые слоевища становятся сдавленно-цилиндрическими и совсем плоскими, приобретая форму ланцетовидных листочков. Одной из характерных черт рода, как и всего порядка, является двустороннее и перистое ветвление, хотя у ряда просто устроенных видов преобладает еще неправильное расположение ветвей вокруг всего слоевища (рис. 167, 1). Кроме того, примитивным видам свойственно ползучее слоевище, которое нередко приобретает дорсовентральное строение. Главные ветви при таком строении стелются по субстрату, цепляясь за него дополнительными ризоидами - присосками. Склонность к образованию стелющихся ветвей проявляется не только у простоорганизованных видов. Нередко это связано со специфическими условиями обитания. Гелидиевые весьма характерны для прибойной скалистой литорали, где более других способны удержаться стелющиеся, плотно прикрепленные к камням формы водорослей.
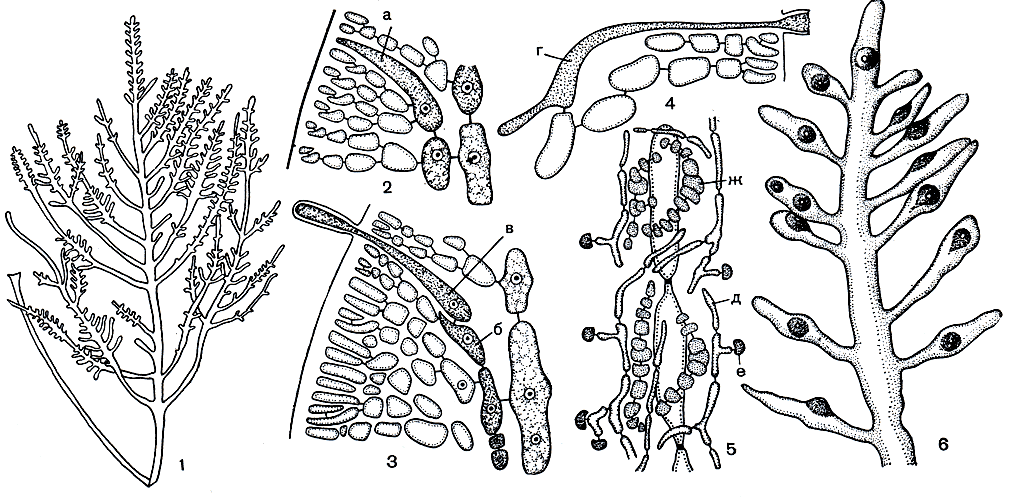
Рис. 167. Строение и размножение Gelidium: 1 - внешний вид растения; 2-5 - развитие карпогонной ветви и гонимобласта; 6 - веточка с цистокарпами. а - инициальная клетка карпогонной ветви, б - несущая клетка, в - карпогон, г - клетка слияния, д - клетки нитей гонимобласта, е - карпоспоры, ж - питающая ткань
Все гелидиевые построены по одноосевому типу. Верхушечная клетка главной оси в результате деления поперечной перегородкой отделяет вниз клетки. Каждая из этих клеток, в свою очередь, отчленяет вбок четыре перицентральные клетки - вначале по одной справа и слева, позднее еще две, при этом все четыре располагаются крестообразно (рис. 152, 1). Из перицентральных клеток возникают разветвленные ветви ограниченного роста, клетки которых, будучи очень тесно расположенными, образуют псевдопаренхимную ткань. Конечные ответвления этих веточек, плотно смыкаясь, формируют внешнюю кору. В процессе формирования слоевища клетки центральной оси вытягиваются в длину и становятся очень тонкими, вследствие этого различить центральную ось в относительно старых частях слоевища почти невозможно. От клеток внутренней части слоевища развивается множество тонких ризоидов. У гелидиума основная масса ризоидов сосредоточена по периферии внутренней части слоевища ближе к коровому слою. Таким образом, особенностью строения гелидиума является образование псевдопаренхимной структуры в результате плотного расположения клеточных нитей, слагающих слоевище, и сильное развитие толстостенных ризоидов, выполняющих, по-видимому, механическую роль. Эти особенности хорошо заметны на поперечном срезе через слоевище (рис. 152, 2). Наличие или отсутствие ризоидов и место, где они образуются, служат хорошим таксономическим признаком для разграничения родов гелидиевых.
Клетки гелидиевых многоядерные, содержат множество пластинчатых хлоропластов и лишены пиреноидов.
Размножается гелидиум половым и бесполым способом. При этом отличить гаметофиты от спорофитов можно только по органам размножения. Тетраспорангии собраны группами на конечных уплощенных веточках вблизи их вершин (рис. 158, 3). Здесь тетраспорангии располагаются беспорядочно или правильными параллельными рядами. Материнской клеткой тетраспорангии становится поверхностная коровая клетка, которая в процессе созревания спорангия погружается в глубь коры. Тетраспорангии делятся крестообразно с образованием одноядерных тетраспор, или, изредка, двухядерных биспор.
Мужские и женские органы размножения развиваются на разных растениях. Сперматангии образуются на вершине молодых веточек и на основных ветвях. Поверхностные коровые клетки вытягиваются, теряют окраску и превращаются в материнскую клетку сперматангия, которая отчленяет от себя один сперматангий. После его выхода та же клетка может образовать еще один сперматангий. Образование сперматангиев ограничено отдельными участками слоевища, которые имеют форму неправильных пятен. На этих участках все коровые клетки становятся материнскими клетками сперматангиев.
Карпогоны и затем цистокарпы возникают на специальных коротких веточках, которые в зрелом состоянии легко отличить от вегетативных по внешнему строению. Обычно они уплощенные, эллиптической или овальной формы. Клетка, дающая начало карпогону, располагается во внутренней коре, ближе к центру слоевища (рис. 167, 2-5). Эта инициальная клетка сильно вытягивается и достигает поверхности слоевища. Позднее она делится поперек, отчленяя карпогон с длинной трихогиной. При этом нижняя клетка остается в качестве несущей и к карпогонной ветви непосредственно не относится. Таким образом, карпогонная ветвь гелидиума одноклеточная. По мере развития карпогона, еще до оплодотворения, в непосредственной близости от него возникают дополнительные мелкоклеточные нити, образующие рыхлую питающую ткань. После оплодотворения карпогон увеличивается в размерах и постепенно сливается с несущей клеткой. В результате этого слияния, а в ряде случаев и слияния несущей клетки также с соседними вегетативными клетками образуется крупная клетка слияния. Она продолжает увеличиваться в размерах и окружается вегетативными клетками, но нового слияния между ними здесь уже не происходит. Клетка слияния выпускает несколько лопастных отростков. На их вершине образуются клеточные перегородки, отделяющие первичные клетки гонимобласта, из которых после повторных делений развиваются нити гонимобласта. По мере развития они стелются между клетками центральной части слоевища, образуя спутанные массы, а не пучки, как у многих флоридеевых. Процесс оплодотворения стимулирует образование питающих клеток. После него они возникают в еще большем количестве и сливаются с нитями гонимобласта. По мере созревания гонимобласта некоторые клетки его нитей увеличиваются в размерах, вытягиваются и отчленяют в сторону поверхности слоевища одну или несколько крупных карпоспор. В месте развития гонимобласта кора приподнимается и образует крышу цистокарпа.
На ранних стадиях прорастания спор гелидиума (как тетраспор, так и карпоспор) формируется плотный узелок клеток, от которого вниз отходит один или несколько первичных ризоидов. Из этого узелка очень быстро вырастают вертикальные побеги. Характер прорастания гелидиевых весьма своеобразен и не похож на таковой остальных багрянок.
Если сравнить гелидиевые с остальными красными водорослями, то сразу бросается в глаза, что это не только четко очерченная, но и достаточно сильно обособленная группа. До последнего времени считали, что провести какие-либо родственные связи между порядком гелидиевых и другими багрянками невозможно. Это объясняется тем, что не были изучены детали развития женской генеративной системы и гонимобласта гелидиевых. Однако на основе современных знаний все-таки можно говорить о связи гелидиевых с нема лиевыми, с одной стороны, и с криптонемиевыми - с другой. Так же как у некоторых немалиевых, карпогон гелидиевых после оплодотворения сливается с клеткой карпогонной ветви или с несущей клеткой последней. Гонимобласт в обеих группах состоит из разветвленных нитей диффузного роста. Слоевище здесь и там имеет одноосевое строение. В обеих группах имеются специальные питающие нити, которые тесно связаны с карпогонной ветвью. Нити гонимобласта развиваются из карпогона. С другой стороны, существуют филогенетические связи гелидиевых с водорослями семейства дюмонтиевых (порядок криптонемиевых). У последних уже есть ауксиллярные ветви, но морфологически они настолько подобны питающим нитям гелидиевых, что это сходство позволяет считать их гомологичными структурами, т. е. имеющими одинаковое происхождение.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'