Порядок немалиевые (Nemaliales)
Порядок немалиевых объединяет организмы, у которых отсутствуют ауксиллярные клетки. Нити гонимобласта развиваются непосредственно от оплодотворенного карпогона или от нижележащей клетки после того, как в нее перейдет ядро карпогона. Специальных питающих клеток у немалиевых тоже нет. У сложно организованных форм во время оплодотворения или после него происходит слияние клеток карпогонной ветви и несущей клетки в самых разных сочетаниях.
Порядок немалиевых - наиболее примитивная группа красных водорослей. Он объединяет 8 семейств, которые можно разделить на две группы. В первой группе слоевище имеет одноосевое строение, во второй - оно построено по многоосевому типу. Почти все флоридеевые, которые обитают в пресных водах, относятся к немалиевым водорослям.
Семейство акрохетиевые (Acrochaetiaceae)
Эти водоросли устроены наиболее просто. Слоевище состоит из свободно разветвленных однорядных нитей. Они образуют кустики микроскопических размеров (как правило, высотой до 5 мм), и только как исключение встречаются более крупные растения, высотой до 25 мм. Ветвление в этих кустиках попеременное, неправильное (рис. 151, 1; 162).
Часто короткие боковые веточки на вершине заканчиваются волоском, однако имеется немало видов, у которых волоски никогда не образуются.
Лишь немногие акрохетиевые водоросли растут на грунте - на камнях, скалах и т. п., большинство же предпочитает поселяться на других водорослях и животных. Особенно часто их можно встретить на ламинариевых водорослях, на гидроидах, мшанках, моллюсках. При этом они не только прикрепляются к поверхности организмов, но и проникают в ткани хозяина.
Строение базальной части акрохетиевых меняется в зависимости от характера субстрата. На твердом субстрате, будь то камни или водоросли с плотным строением, чаще встречаются псевдопаренхимные подошвы, на рыхлом - свободные стелющиеся нити. В целом для акрохетиевых весьма характерна разнонитчатая структура. Наиболее отчетливо она проявляется у водорослей в молодом состоянии, так как при разнонитчатом строении вначале обычно развивается стелющаяся структура, из клеток которой возникают вертикальные побеги (рис. 162, 2).
Внутриклеточное строение представителей семейства отличается разнообразием. Здесь можно встретить формы с осевым и пристенным хлоропластом, с пиреноидами и без них. Форма хлоропластов служит хорошим признаком для разграничения родов. У одних они звездчатые, у других - лентовидные, спирально закрученные, у третьих - в виде дисков.
Бесполое размножение у акрохетиевых осуществляется преимущественно моноспорами, тетраспоры отмечаются довольно редко. Спорангии сидят на коротких боковых веточках, где располагаются одиночно или по 2-3 вместе (рис. 162,2). Тетраспоры делятся крестообразно. Карпогон - единственная клетка, располагается она сбоку от основных ветвей или на коротких боковых веточках. Изредка встречается двухклеточная карпогонная ветвь. Сперматангии в виде мелких округлых клеток располагаются на вершине боковых веточек (рис. 162, 3).
После оплодотворения и отпадения трихогины карпогон делится поперечными перегородками на несколько клеток, из которых вырастает пучок нитей гонимобласта (рис. 162, 4). Не всем акрохетиевым свойственно половое размножение; некоторые размножаются только бесполым способом, среди них - многие виды родохортона (Bhodochorton).
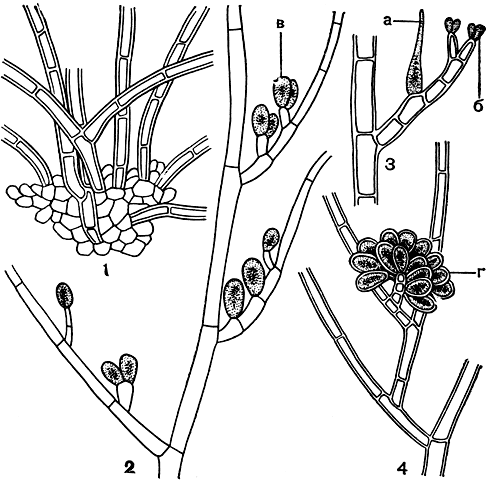
Рис. 162. Строение акрохетиевых: 1 - базальная часть растения Acrochaetium; 2 - часть веточки с моноспорангиями; 3 - часть веточки с карпогоном и спермациями; 4 - веточка с гонимобластом. а - карпогон, б - сперматангий, в - моноспорангий, г - карпоспоры
В природе акрохетиевые распространены чрезвычайно широко. В большинстве своем - это морские организмы, известные во всех морях Мирового океана. Пресноводных видов намного меньше - всего 5, они принадлежат к роду одуинелла (Audouinella).
Из морских обитателей самой характерной и распространенной водорослью семейства является акрохетиум (Acrochaetium). В литературе описано около 250 видов этого рода, но в действительности их, по-видимому, гораздо меньше. Когда эти виды начали изучать в культуре, особенно их размножение и циклы развития, то обнаружилось, что многие прежние виды - не что иное, как стадии или формы развития в жизненном цикле других видов.
Другая морская водоросль семейства - родохортон - отличается более грубым и крупным слоевищем, растущим, как правило, на каменистом грунте. На прибойной литорали наших северных морей родохортон часто образует самостоятельный пояс, покрывая затененную поверхность валунов и скал темно-красным налетом.
Филогенетическое положение семейства акрохетиевых не совсем ясно. Хотя его и помещают как наиболее просто организованную группу в начале всей системы флоридеевых, до сих пор неизвестно, говорит ли эта простота о примитивности его представителей или она результат вторичного упрощения.
В последнее время было обнаружено, что некоторые багрянки имеют в своем цикле развития стадии или поколения точно такого же строения, как акрохетиевые. Это наблюдение позволяет предположить, что семейство представляет собой искусственную группу, которая требует более подробного изучения. В этой связи интересно подчеркнуть, что у ряда водорослей семейства неизвестно половое размножение и что они заметно различаются внутриклеточным строением.
Семейство батрахоспермовые (Batrachospermaсеае)
Это семейство интересно тем, что в нем можно проследить первые шаги в усложнении организации слоевища багрянок при переходе от свободно-нитчатого к плотному строению. Характерные черты семейства лучше всего проследить, познакомившись с одной из самых известных пресноводных багрянок - батрахоспермумом (Batrachospermum). Около 50 видов этого рода распространено во всех частях света. Предпочитают они хорошо аэрируемые воды медленно текущих рек и ручьев, но их можно встретить и в прибрежной части озер, там, где отсутствуют сильные колебания температуры. Некоторые проникают даже в торфяные болота, но чаще встречаются вблизи ключей.
Мягкое слизистое слоевище батрахоспермума состоит из центральной однорядной оси, разветвленной моноподиально или псевдодихотомически. Основные ветви густо покрыты пучками коротких веточек ограниченного роста, расположенных мутовками (рис. 151, 2; 163). Строение батрахоспермума легко представить как результат усложнения и упорядочения строения акрохетиевых, и это подтверждается индивидуальным развитием видов батрахоспермума. Вначале образуется стелющаяся по субстрату нить, она ветвится с образованием псевдопаренхимной подошвы, из клеток которой возникает множество вертикальных однорядных побегов, морфологически почти неотличимых от акрохетиума. Эта стадия, которую так и называют стадией акрохетиума, может существовать достаточно долгое время и даже размножаться моноспорами (рис. 163, 2). Немудрено, что ее часто принимают за самостоятельные виды акрохетиума. Развитие этой стадии в большой степени определяется условиями среды. В целом ее можно назвать теневой формой роста, тогда как стадия батрахоспермума - световая форма. В связи с этим в природе они приурочены к разным сезонам. Побеги батрахоспермума развиваются в любом месте стадии акрохетиума. Их строение относится к центральноосевому типу, который в литературе часто называют типом батрахоспермума. Верхушечная клетка центральной оси отчленяет новые сегменты, которые быстро растут в длину и толщину. На каждом из этих сегментов образуются мутовки из 4-6 мелких клеток, которые постепенно вырастают в пучки ветвей ограниченного роста. Конечные клетки боковых веточек нередко преобразуются в волоски. Ветви в пучках связаны большим количеством слизи. Из нижних клеток ветвей ограниченного роста возникают обычно клеточные ризоидные нити, спускающиеся вниз по слоевищу и обволакивающие центральную ось (рис. 151, 2). От этих нитей могут развиваться дополнительные пучки ветвей ограниченного роста, которые вместе с первыми служат ассимиляционными нитями.
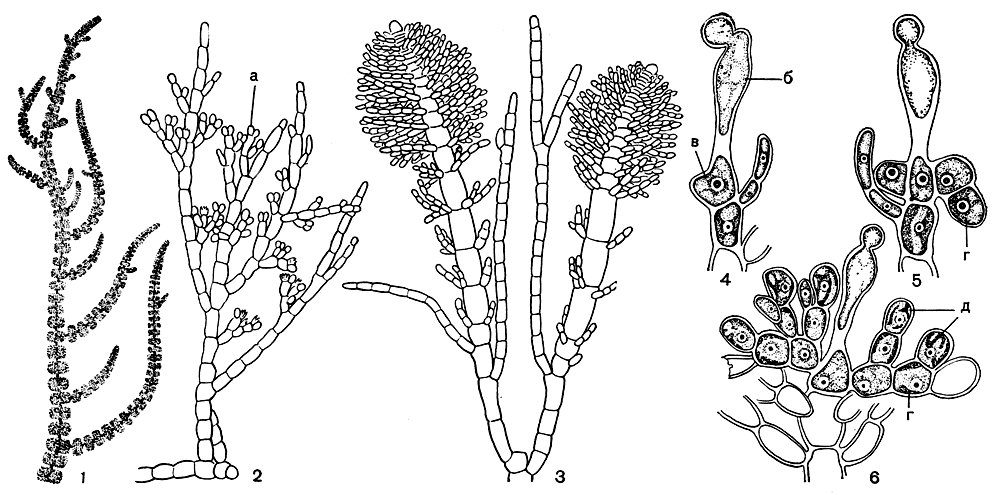
Рис. 163. Строение Batrachospermum: 1 - внешний вид части растения; 2 - ювенильная стадия; 3 - начало образования взрослого слоевища; 4-6 - стадии развития гонимобласта. а - моноспорангии, б - трихогина, в - оплодотворенный карпогон, г - клетки гонимобласта, д - карпоспоры
Органы бесполого размножения - моноспорангии - на растениях батрахоспермума встречаются очень редко, гораздо чаще они образуются на неполовозрелых растениях, подобных акрохетиуму. Половое размножение отличается некоторыми особенностями. Трихогина необычной для красных водорослей кеглевидной или цилиндрической формы. Карпогон образуется на вершине веточек, возникающих от нижних клеток ветвей ограниченного роста (рис. 163, 4). Шаровидные сперматангии развиваются на вершине боковых веточек, по 1-3 на клетках, которые ничем не отличаются от вегетативных. Зрелые гонимобласты - четко отграниченные образования, погруженные в слизь среди ассимиляционных нитей и плотно окруженные ими (рис. 163, 4-6; 151, 2).
Семейство леманеевые (Lemaneaceae)
Пресноводная водоросль леманея (Lemanea) - единственный представитель семейства леманеевых. Разветвленное или простое трубчатое слоевище леманеи (рис. 164, 1) построено по тому же принципу, что и батрахоспермум. Центральная однорядная ось несет мутовки боковых разветвленных ветвей, но, в отличие от батрахоспермума, они располагаются не свободно, а разрастаются и ветвятся так, что смыкаются своими конечными ответвлениями и тем самым образуют вокруг центральной оси, на некотором расстоянии от нее, сплошной коровой слой, который выполняет ассимиляционную функцию. На продольном срезе через слоевище леманеи хорошо видно, что коровой слой образуется из пучков ветвей ограниченного роста, но эти ветви видоизменяются и их клетки дифференцируются. Внешние клетки коры, образованные густо разветвленными веточками последних порядков, самые мелкие и наиболее богатые пигментами, а глубже расположенные клетки крупнее, менее интенсивно окрашенные или совсем бесцветные. Как и у батрахоспермума, у леманеи от нижних клеток боковых веточек образуются ризоидные нити, которые простираются вдоль центральной оси и заполняют полость цилиндрического слоевища (рис. 151, 3).
У леманеи также имеется стадия акрохетиума, но на ней никогда не образуются ни моноспоры, ни тетраспоры. До сих пор у этого рода известно только половое размножение. Сперматангии и карпогон находятся на одном и том же растении. Первые развиваются на внешних клетках коры и собраны в виде отдельных пятен или сплошного кольца на тех участках слоевища, где расположены мутовки ветвей ограниченного роста. Материнская клетка сперматангия вытянута в длину и несет обычно на вершине одноклеточный волосок, который, после того как сперматангий отчленяется, сдвигается в сторону, а потом и совсем отпадает (рис. 164, 2). Карпогонная ветвь возникает под кодовым слоем. Она состоит из 2-7 клеток, самая нижняя клетка нередко несет боковую веточку, на которой также может развиваться карпогонная ветвь (рис. 164, 3). После оплодотворения карпогон дает начало клеточной нити, растущей внутрь слоевища и разветвляющейся с образованием густых пучков нитей (рис. 164, 4). Не только конечные, но и большая часть клеток нитей гонимобласта превращается в карпоспоры, которые выходят после разрыва стенки слоевища.
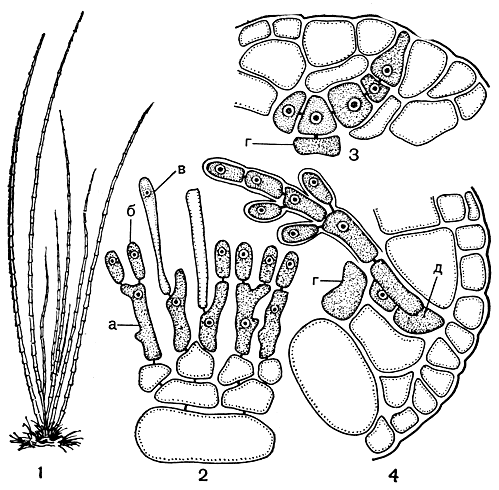
Рис. 164. Строение Lemanea: 1 - внешний вид растений; 2 - образование спарматангиев; 3 - карпогонная ветвь; 4 - начало развития гонимобласта. а - материнская клетка сперматангия, б - сперматангий, в - волосок, г - несущая клетка, д - карпогон
При неблагоприятных условиях они могут довольно долгое время оставаться внутри слоевища. Это является своеобразным приспособлением к меняющимся условиям среды. Так, в засушливый период, когда в мелких водоемах наблюдается недостаток воды, выхода карпоспор не происходит.
Большой интерес ученых вызывает цикл развития леманеи. Вначале он казался очень простым. Были известны только гаметофиты, и поэтому предполагалось, что весь цикл проходит в гаплоидной фазе, диплоидная же ограничена зиготой. Но теперь мы уже знаем, что за внешней простотой этого цикла скрываются сложные процессы. Когда было найдено действительное место редукционного деления, выяснилось, что первоначально в цикле развития присутствовал тетраспорофит, но постепенно он претерпел редукцию и потерял способность образовывать тетраспоры. Карпоспоры леманеи прорастают в растения, которые соответствуют тетраспорофиту. Начинается прорастание с образования стадии акрохетиума, из нее позднее возникают растения леманеи. В определенный период этого роста в клетках леманеи происходит редукционное деление, после чего образуется уже гаплоидная ткань, которая соответствует гаметофиту.
У леманеи очень сильно развита способность к вегетативному размножению и регенерации. Она может выдерживать длительное высыхание, оставаясь жизнеспособной, и, попав вновь в условия увлажнения, давать множество новых побегов. Растет леманея в континентальных водоемах всего мира, предпочитая мелкие холодные быстротекущие воды, где водится форель.
Семейство боннемезониевые (Bonnemaesoniaceae)
Это семейство занимает обособленное место среди немалиевых, построенных по одноосевому типу. Боннемезония (Bonnemaesonia) - водоросль с довольно ограниченным распространением, но она чрезвычайно интересна благодаря особенностям строения и размножения. Слоевище боннемезонии вальковатое или слегка уплощенное, густо покрыто веточками ограниченного роста, которые располагаются в одной плоскости, супротивно, придавая слоевищу перистый вид (рис. 165, 1). Как же растет и формируется слоевище этой водоросли? Апикальная клетка делится косой перегородкой, поэтому возникающие сегменты располагаются под углом друг к другу (рис. 165, 2). От этих центральных клеток отделяются по две друг против друга расположенные перицентральные клетки. Они образуются неодновременно, и та, что отчленяется первой, быстро вырастает в очень короткую неразветвленную ветвь ограниченного роста, которая при созревании слоевища всегда остается стерильной. Другая перицентральная клетка развивается в длинную ветвь неограниченного роста или, оставаясь короткой, несет на себе органы размножения. В ходе дальнейшего развития короткие боковые веточки становятся многорядными. Это происходит следующим образом.
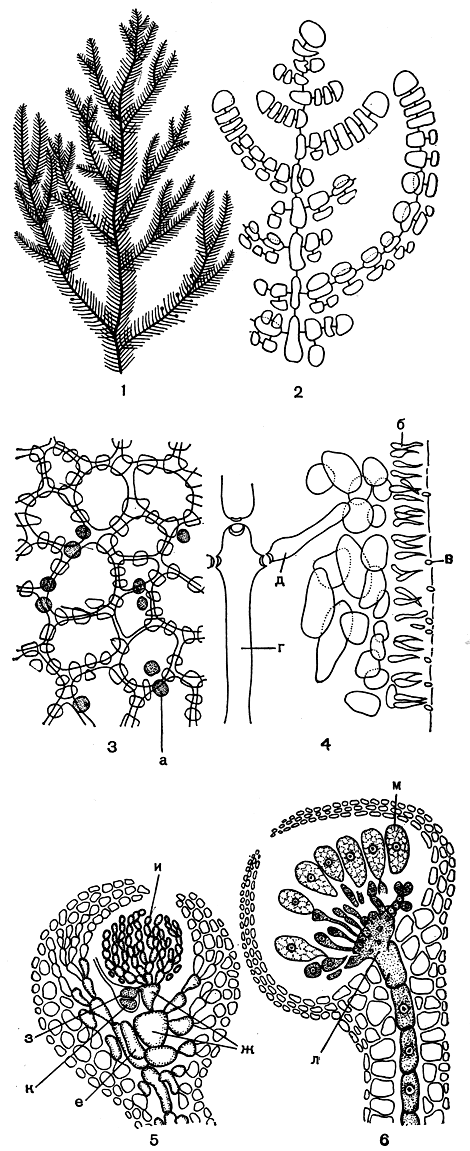
Рис. 165. Строение боннемезониевых: 1 - внешний вид веточки Bonnemaesonia; 2 - вершина веточки; 3 - строение коры с поверхности слоевища; 4 - часть продольного среза слоевища со сперматангиями; 5, 6- развитие цистокарпа. а - железистые клетки, б - материнская клетка сперматангия, в - сперматангий, г - центральная ось, д - перицентральная клетка, е - несущая клетка, ж - клетки кар- погонной ветви, з- карпогон, и - питающие нити, к - первая клетка гонимобласта, л - клетка слияния, м - карпоспоры
Первичные клетки веточек делятся на одну центральную и три расположенные вокруг нее перицентральные клетки. После этого каждая из трех перицентральных клеток образует по четыре клетки, которые составляют первичную кору боннемезониевых. По мере того как центральные клетки растут в длину, коровые образуют вокруг них плотный слой довольно крупных клеток. Постепенно снаружи от первичной коры образуется мелкоклеточный внешний коровой слой. Внешняя кора не образует сплошного покрова, клетки ее имеют сетчатое расположение, и, если смотреть на слоевище с поверхности, можно заметить, что они сосредоточены у клеточных стенок первичной коры. В коре разбросано много железистых клеток (рис. 165, 3). Формирование ветвей неограниченного роста происходит подобным, но более сложным способом. На поперечном срезе через слоевище можно различить ограниченную коровым слоем полость, в центре которой проходит основная ось. Ризоиды, развивающиеся у других немалиевых вокруг центральной оси, здесь отсутствуют.
Плодоносные веточки боннемезонии гомологичны ветвям неограниченного роста, поэтому они располагаются напротив коротких веточек и обычно чередуются с ними на оси. Мужские веточки, за исключением вершины и основания, как муфтой, покрыты сперматангиями. При образовании последних клетки внешней коры располагаются сплошным слоем и все превращаются в материнские клетки сперматангиев. Одна клетка дает начало двум или трем сперматангиям (рис. 165, 4).
В каждой женской плодоносной веточке образуется, как правило, по одной карпогонной ветви; при этом несущей клеткой служит одна из трех перицентральных клеток. Карпогонная ветвь трехклеточная с длинной, спирально закрученной трихогиной. Интересно, что у боннемезонии оболочка цистокарпа начинает формироваться еще до оплодотворения. От всех трех перицентральных клеток, в том числе и от несущей клетки карпогонной ветви, образуются клеточные нити, которые участвуют в построении стенок цистокарпа. Также до оплодотворения подкарпогонная клетка делится, и от нее образуется пучок мелкоклеточных разветвленных нитей, которые можно рассматривать как питающие. Уже на первых порах развития оплодотворенного карпогона питающие клетки сливаются друг с другом и с подкарпогонной клеткой, обеспечивая питанием развивающийся гонимобласт. Первая клетка гонимобласта отчленяется от карпогона так, что направлена внутрь молодого цистокарпа, ближе к его дну, на котором расположена группа крупных, богатых содержимым клеток. Первая клетка быстро делится, в результате образуется клеточный диск из нескольких слоев клеток. По мере образования этого первичного гонимобласта питательные клетки, соединенные с подкарпогонной, уменьшаются в размерах и теряют свое содержимое. В то же самое время происходит слияние клеток диска между собой, а также с несущей клеткой карпогонной ветви, с богатыми содержимым клетками на дне цистокарпа и некоторыми другими. В результате этого процесса образуется крупная многоядерная клетка слияния. От нее, теперь уже по направлению к наружной стенке цистокарпа, развиваются короткие нити, конечные клетки которых превращаются в карпоспоры. Выходят карпоспоры через специальное отверстие на вершине цистокарпа (рис. 165, 5, 6).
Долгое время тетраспоры у боннемезонии были неизвестны, и поэтому полагали, что бесполая форма развития у нее совсем отсутствует. Но при изучении биологии этой водоросли в культуре оказалось, что карпоспора боннемезонии прорастает в растение, построенное из однорядных разветвленных нитей, отчасти стелющихся, отчасти прямостоячих, образующих дернины высотой около 1-3 см. Это растение-эпифит раньше было известно как водоросль с неясным систематическим положением под названием трайлиелла (Trailiella). Раннее развитие при прорастании карпоспор сопровождается образованием плотного базального диска.
Из этого подробного описания внимательный читатель может заключить, что принцип организации слоевища и особенности размножения боннемезонии заметно отличаются от описанных ранее у немалиевых. Эти отличия не случайны, и теперь уже накопилось много данных, которые позволяют усомниться в правильности систематического положения семейства боннемезониевых в порядке немалиевых. Некоторые специалисты полагают, что боннемезониевые - самостоятельный порядок, который гораздо ближе к церамиевым, чем к нема лиевым, по таким признакам, как способ прорастания и формирования слоевища, характер карпогонной ветви и сложные процессы слияния при развитии гонимобласта, наличие железистых клеток и т. п.
Рассмотренные семейства немалиевых характеризовались одноосевым типом строения. Другую эволюционную линию в порядке представляют водоросли с фонтанным типом строения. Эта линия представлена главным образом морскими водорослями - гельминтокладиевыми и хетангиевыми.
Семейство гелъминтокладиевые (Helminthocladiaceae)
Прекрасным примером строения гельминто- кладиевых служит немалион (Nemalion) - наиболее многочисленный (около 10 видов) и широко распространенный род семейства. Все гелъминтокладиевые - тепловодные организмы. Распространение большинства из них ограничено тропическими и субтропическими морями. Виды немалиона заходят в умеренные широты и потому лучше других гельминтокладиевых известны в морях СССР.
Слизистое червеобразное слоевище немалиона состоит из пучка клеточных нитей, каждая из которых растет посредством апикальной клетки. От пучка радиально расходятся многочисленные вильчато-разветвленные ветви ограниченного роста, образующие рыхлый ассимиляционный слой, погруженный в слизь. От нижних клеток ассимиляционных ветвей отходят тонкие ризоидные нити, служащие для укрепления центрального пучка (рис. 166, 2). У родственной немалиону тропической водоросли лиагоры (Liagora) слоевище местами пропитано известью.
Растения немалиона, несущие половые органы, бывают однодомными и двудомными. Мужские растения обычно меньше женских. При формировании сперматангиев на вершине ассимиляционных веточек вырастают короткие двух-, четырехклеточные нити. Каждая клетка этой нити отчленяет вокруг своей вершины 2-4 сперматангии (рис. 159, 1). Карпогонные ветви у немалиона состоят из различного числа клеток, от 4 до 7. Они образуются на месте обычных боковых веточек ассимиляционных нитей (рис. 166, 2). После оплодотворения карпогон делится поперечной перегородкой. Из двух образовавшихся клеток только верхняя участвует в образовании нитей гонимобласта. Нижняя клетка сливается с остальными клетками карпогонной ветви, включая несущую, в одну крупную, богатую содержимым клетку слияния, служащую источником питания для развивающегося гонимобласта. Зрелые гонимобласты немалиона - это четко отграниченные компактные образования, погруженные в слизь среди ассимиляционных нитей (рис. 166, 3-5). У других водорослей семейства нити гонимобласта стелются между ассимиляционными нитями, образуя негустые пучки веточек с карпоспорами на конце. Карпоспоры у немалиона конечные.
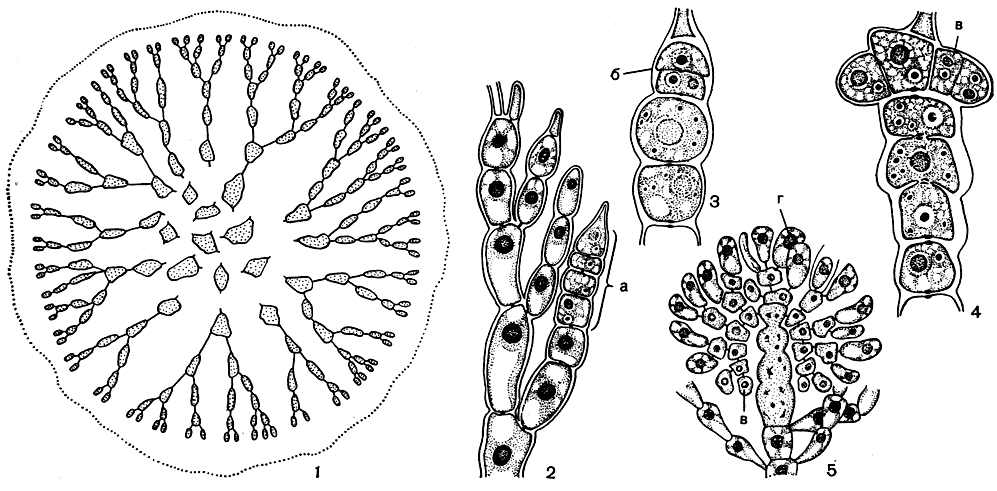
Рис. 166. Строение и размножение Nemalion: 1 - поперечный срез слоевища; 2 - карпогонная ветвь; 3-5 - стадии развития гонимобласта. а - клетки карпогонной ветви; б - карпогон после оплодотворения, в - клетки гонимобласта, г - карпоспоры
До последнего времени у представителей семейства было известно только половое размножение, и считалось, что тетраспорофиты у них отсутствуют. Однако теперь наши знания о цикле развития этих водорослей намного расширились. У немалиона червеобразного (Nemalion vermiculare) и у других видов обнаружены спорофиты - карликовые растения, внешне весьма похожие на растения акрохетиума и родохортона, несущие тетраспоры или моноспоры. Прорастание карпоспор, так же как и тетраспор, происходит с образованием вначале простых, а затем и разветвленных однорядных нитей, которые очень напоминают растения акрохетиума.
Семейство хетангиевые (Chaetangiaceae)
Представители этого семейства, близкого к гельминтокладиевым, построены также по многоосевому принципу. Главные их отличия заключаются в том, что поверхностные клетки ассимиляционных нитей слагаются в сплошной коровой слой, а нити гонимобласта окружены оберткой из тонких перепутанных нитей, в результате чего формируется мешковидный концептакул с отверстием на вершине для выхода карпоспор. Кроме того, семейство характеризуется наличием обызвествленных форм. Среди них широко известна галаксаура (Galaxaurа) - одна из самых обычных водорослей в тропических морях Мирового океана. В большей или меньшей степени обызвествленное и вильчато-разветвленное слоевище галаксауры имеет весьма характерный облик. Любопытная особенность видов этого рода (а их описано около 70) заключается в том, что гаметофит и спорофит, будучи оба макроскопическими растениями, заметно различаются по анатомическому строению. Это служило поводом для описания их как отдельных видов, и к настоящему времени систематика этого рода изрядно запутана.
|
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://plantlife.ru/ 'PlantLife.ru: Статьи и книги о растениях'